Interfacial Recognition of Acetylcholine by an Amphiphilic p-Sulfonatocalix[8]arene Derivative Incorporated into Dimyristoyl Phosphatidylcholine Vesicles

Abstract
:
1. Introduction
2. Results and Discussion
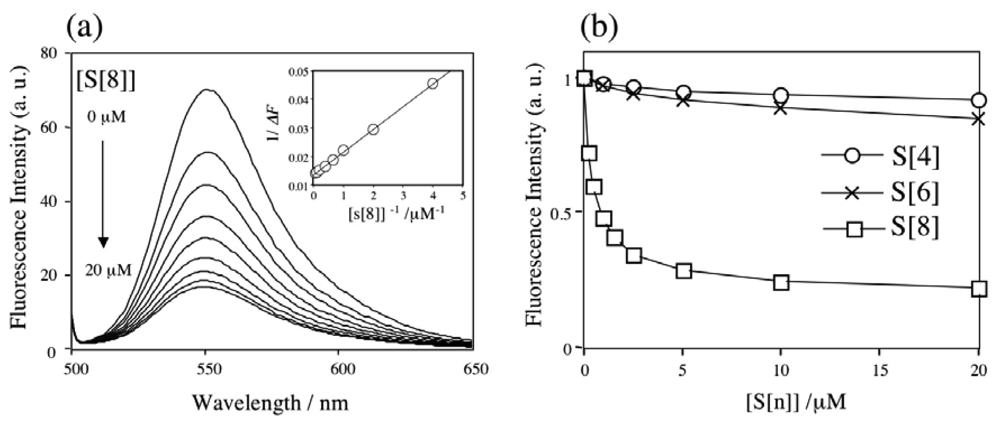
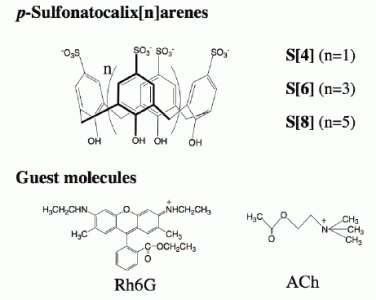
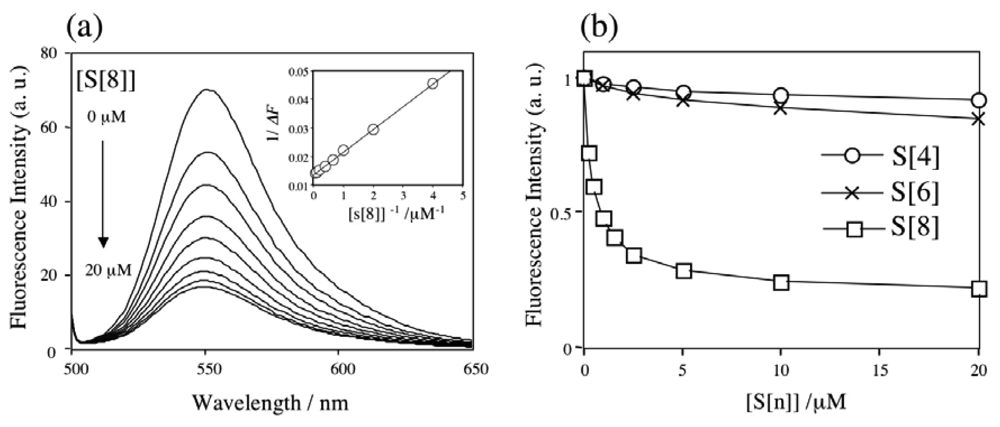
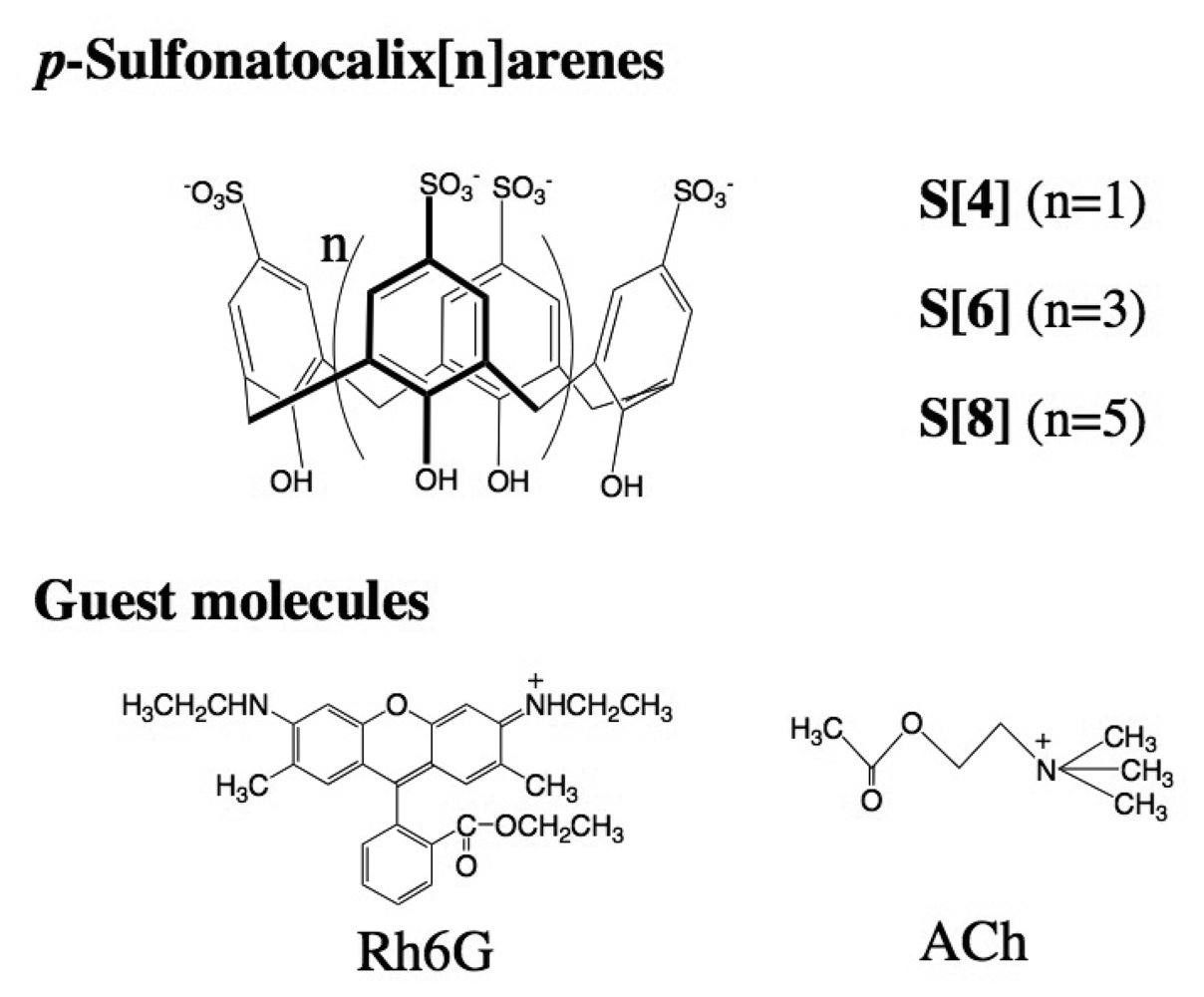
2.1 The binding ability and the stoichiometry of binding of parent p-sulfonatocalix[n]arenes for Rh6G and ACh in aqueous solution
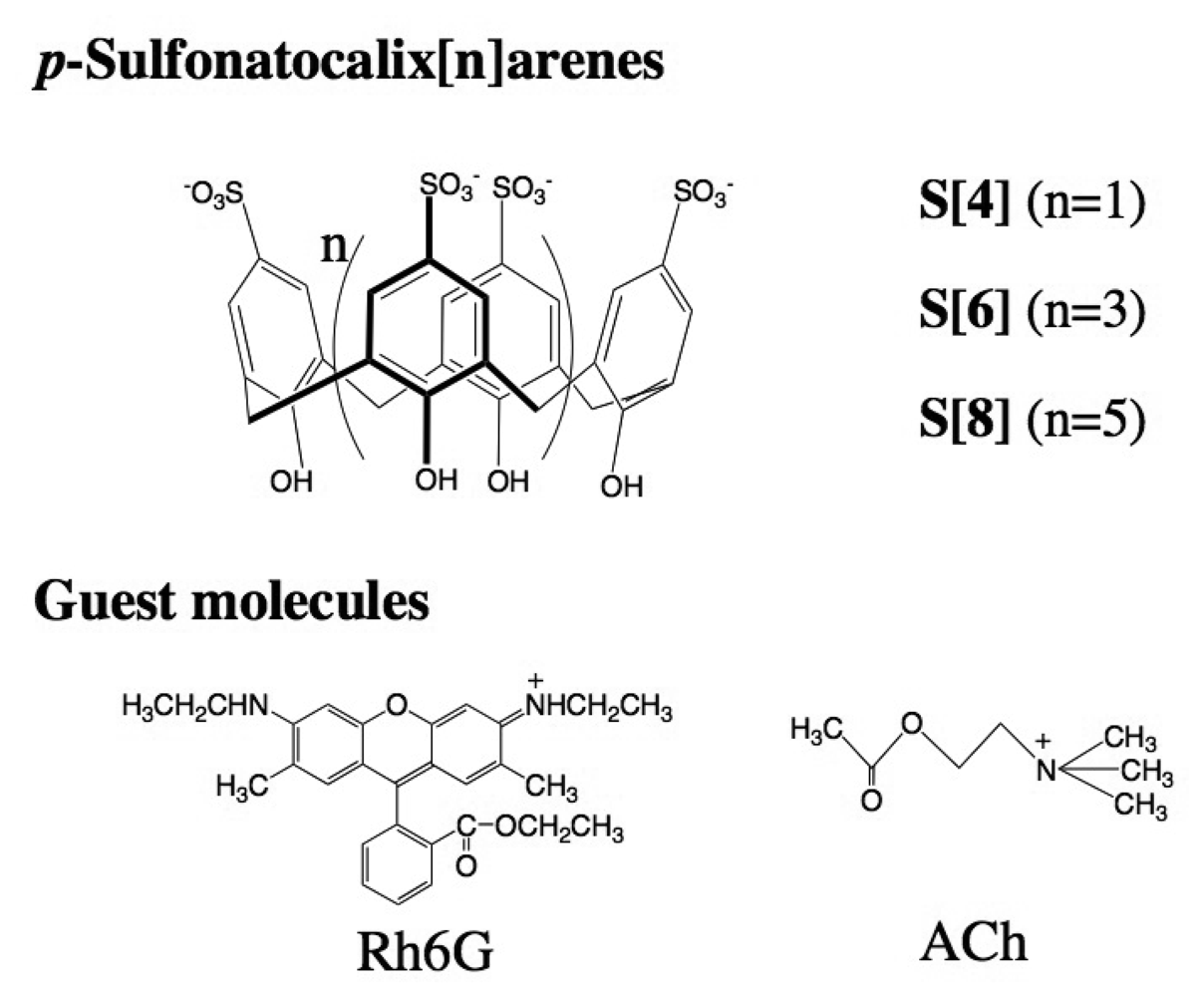
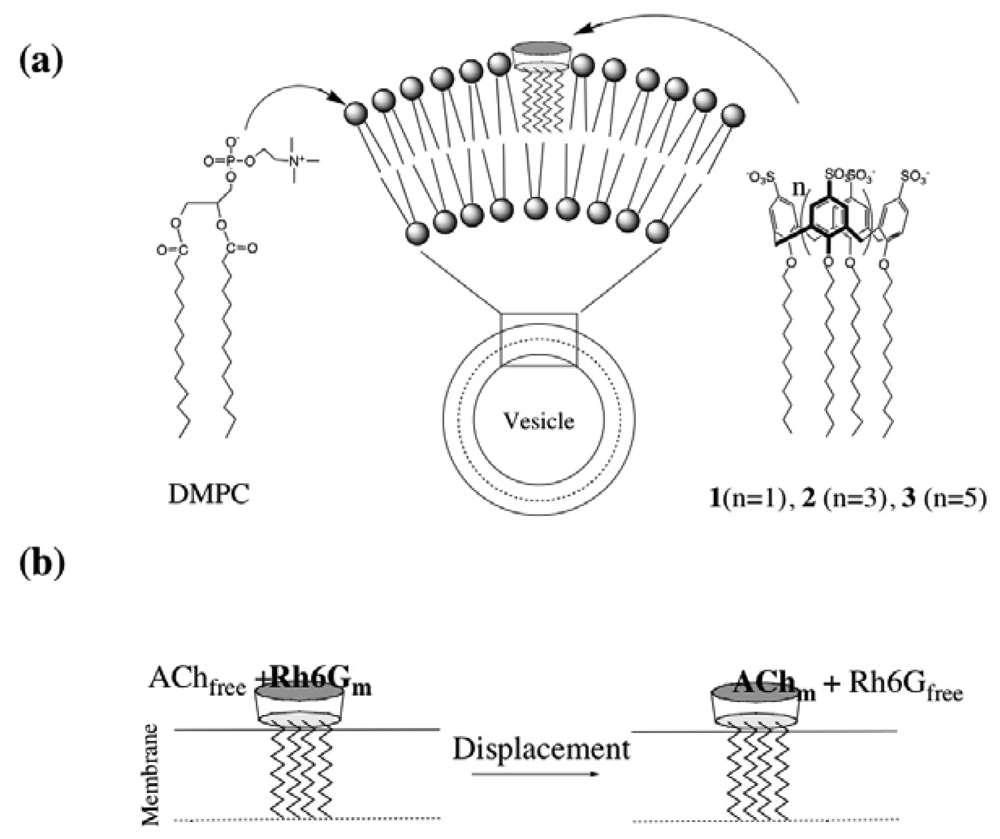
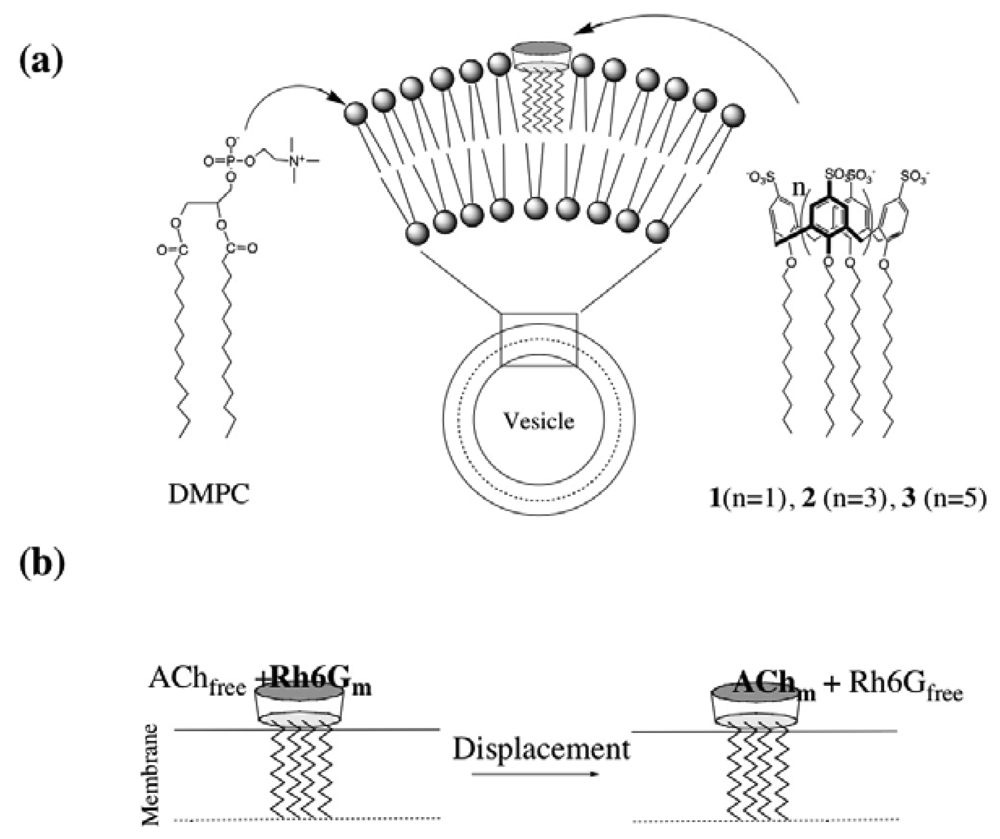
2.2 Characterization of DMPC vesicles containing dodecyl ether derivatives of p-sulfonato-calix[n]arenes 1-3
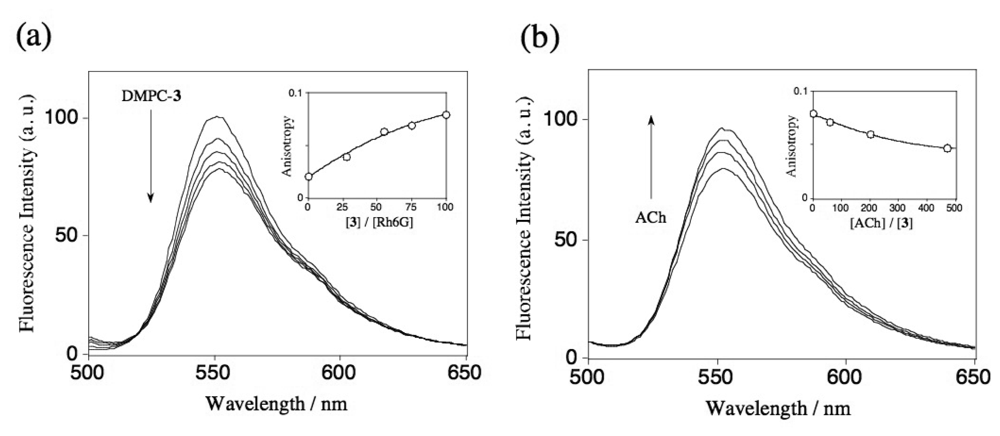
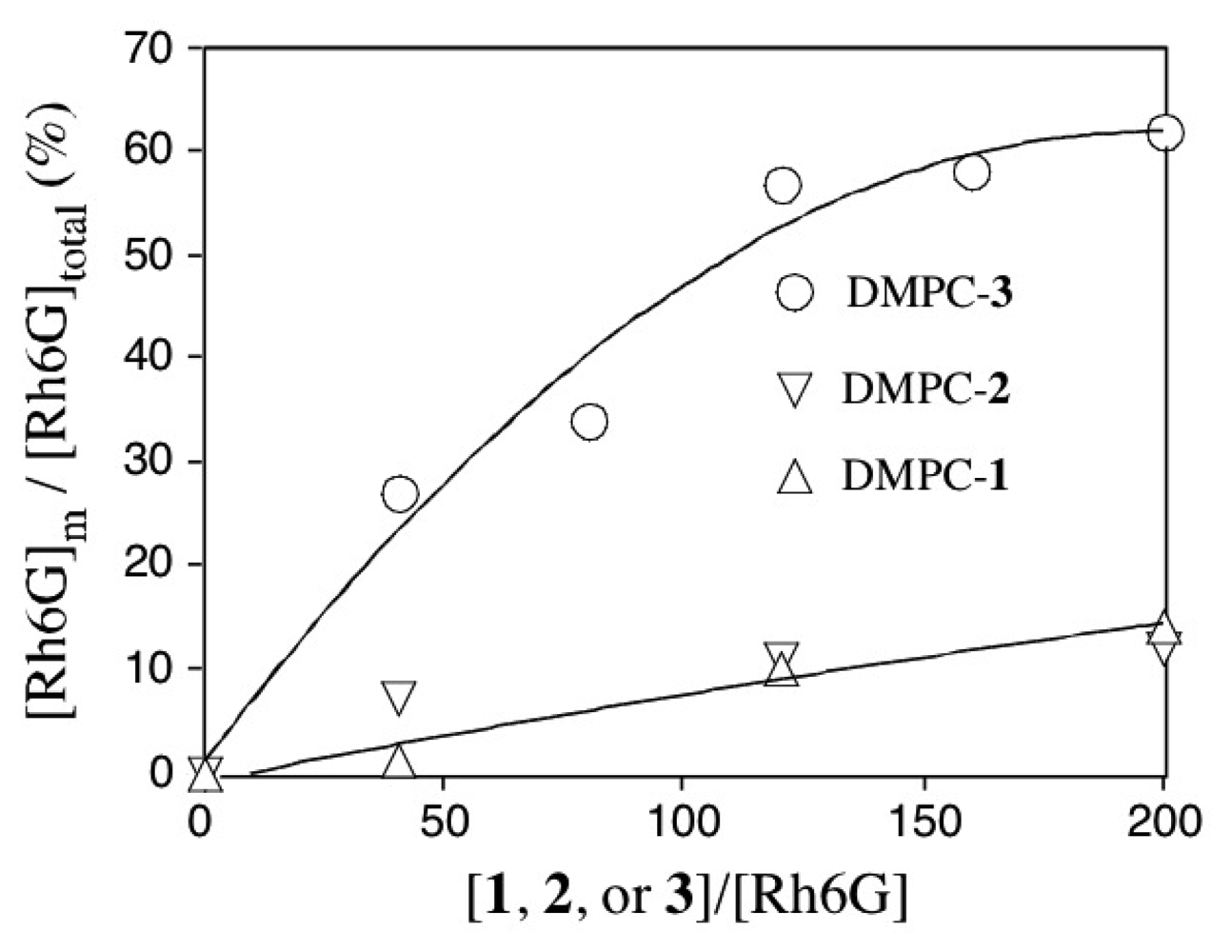
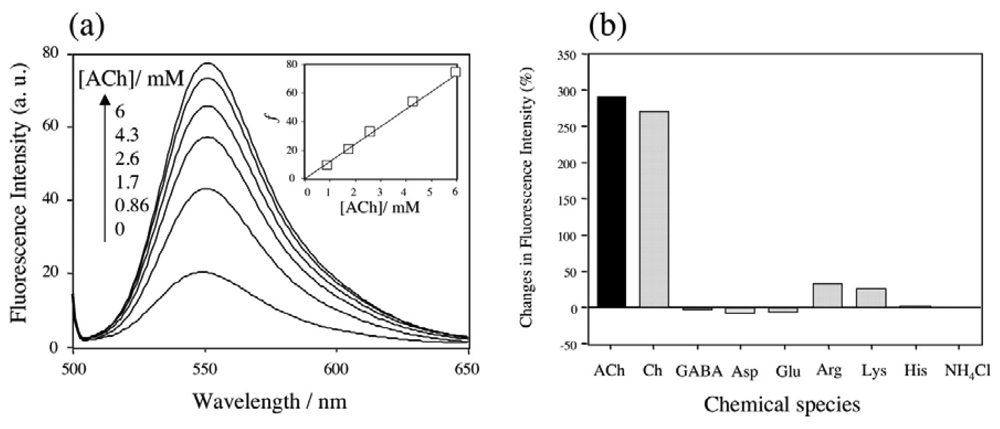
2.3 Fluorescence spectra and fluorescence anisotropy measurements for the binding of Rh6G and ACh to DMPC vesicles containing 1-3
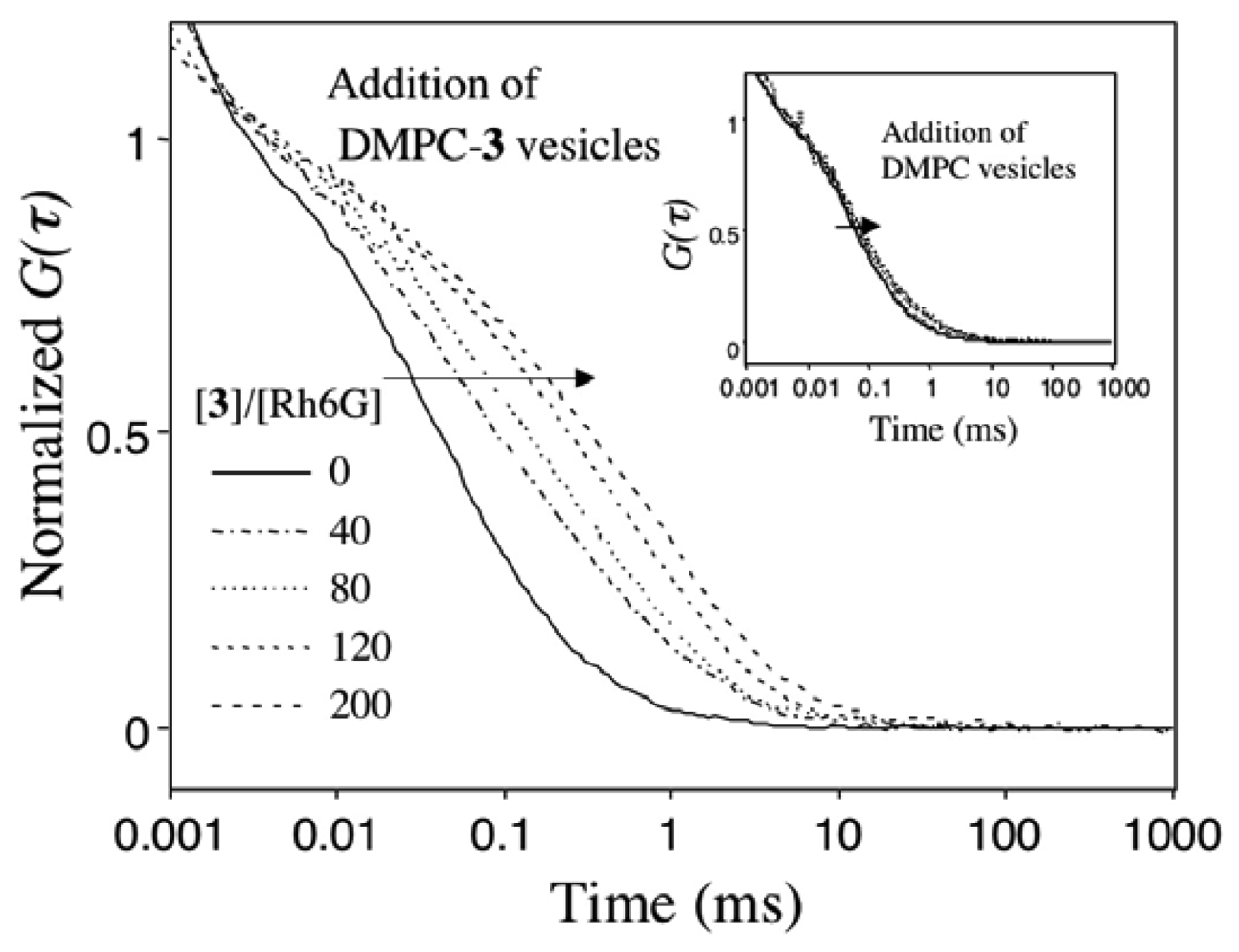
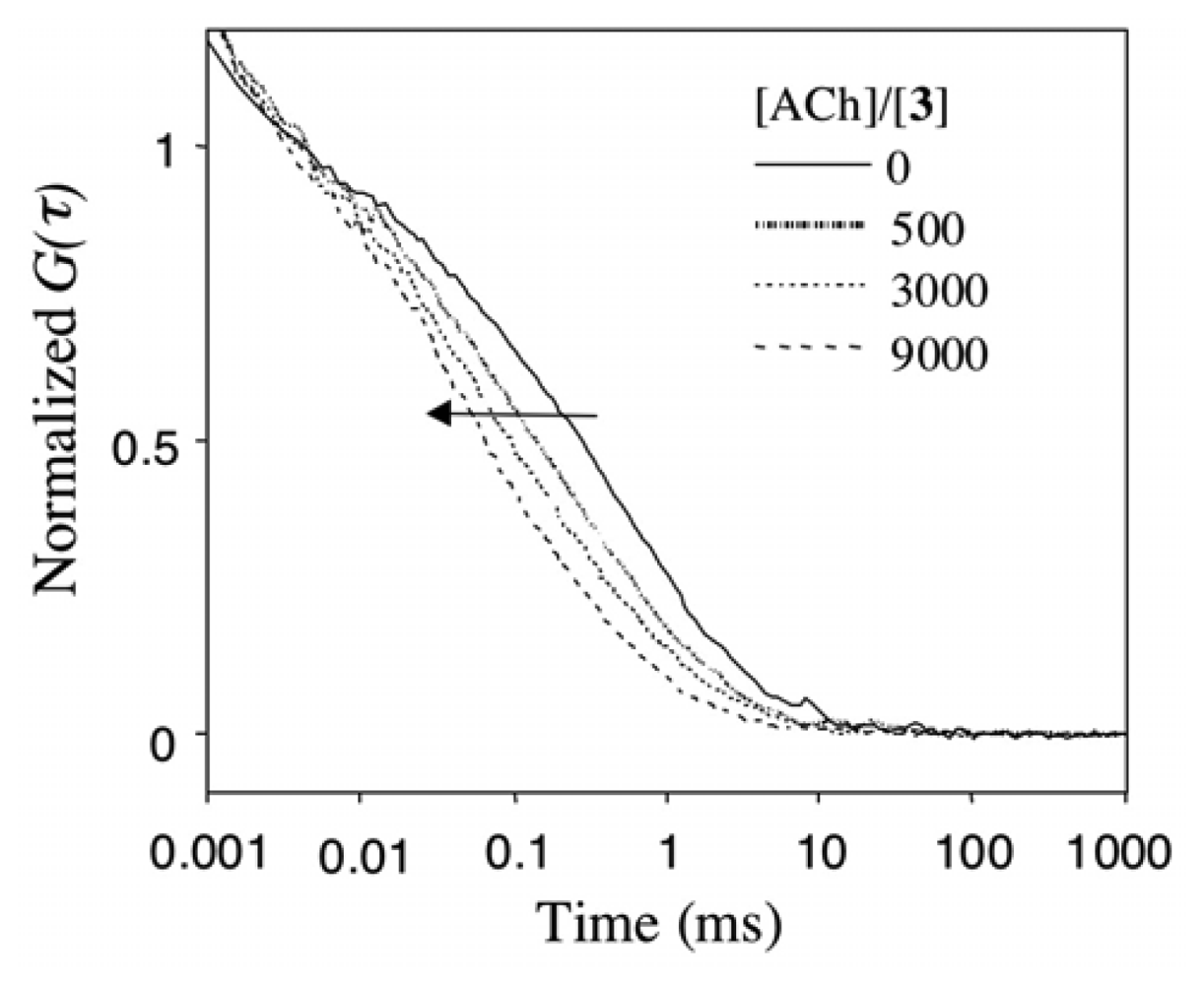
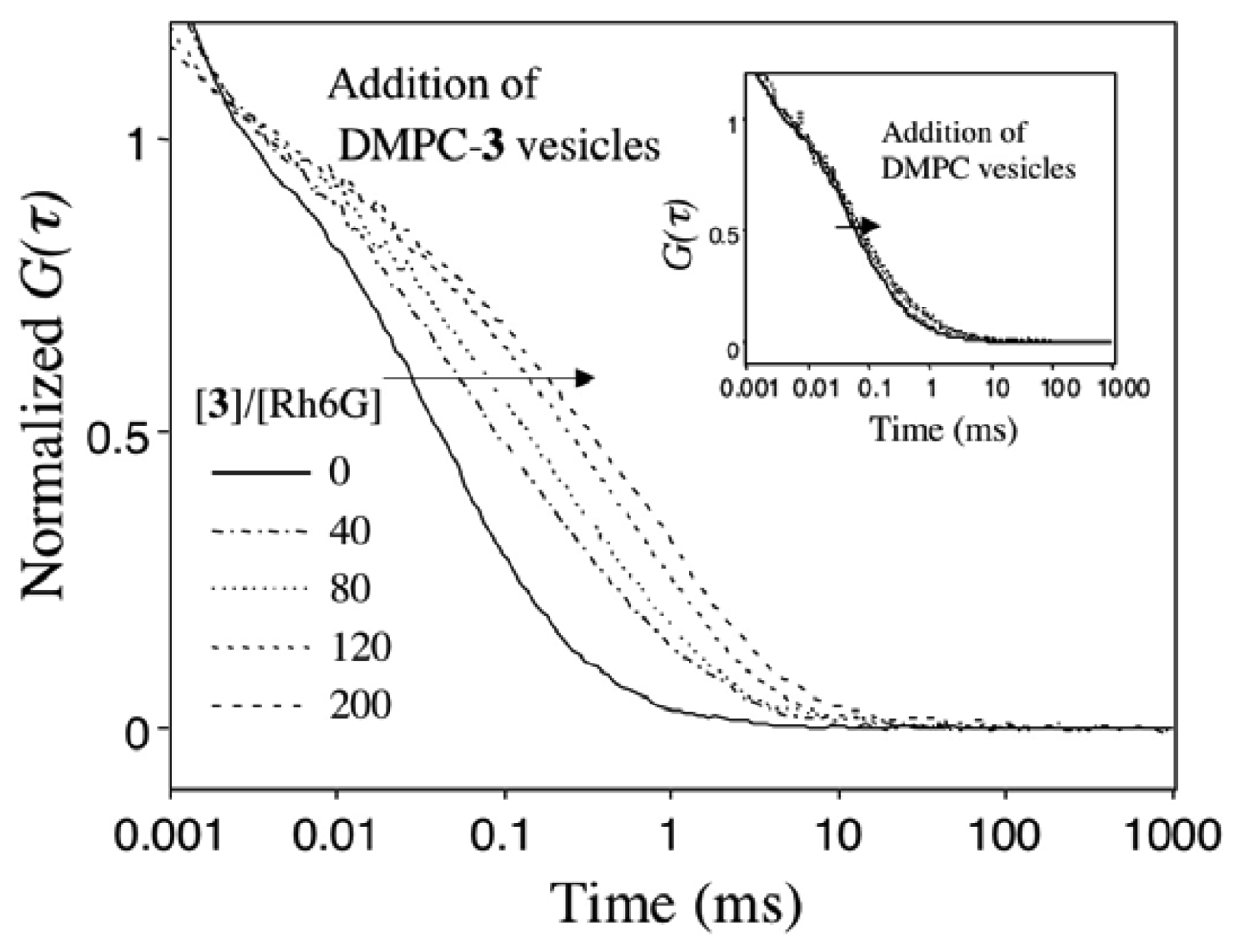
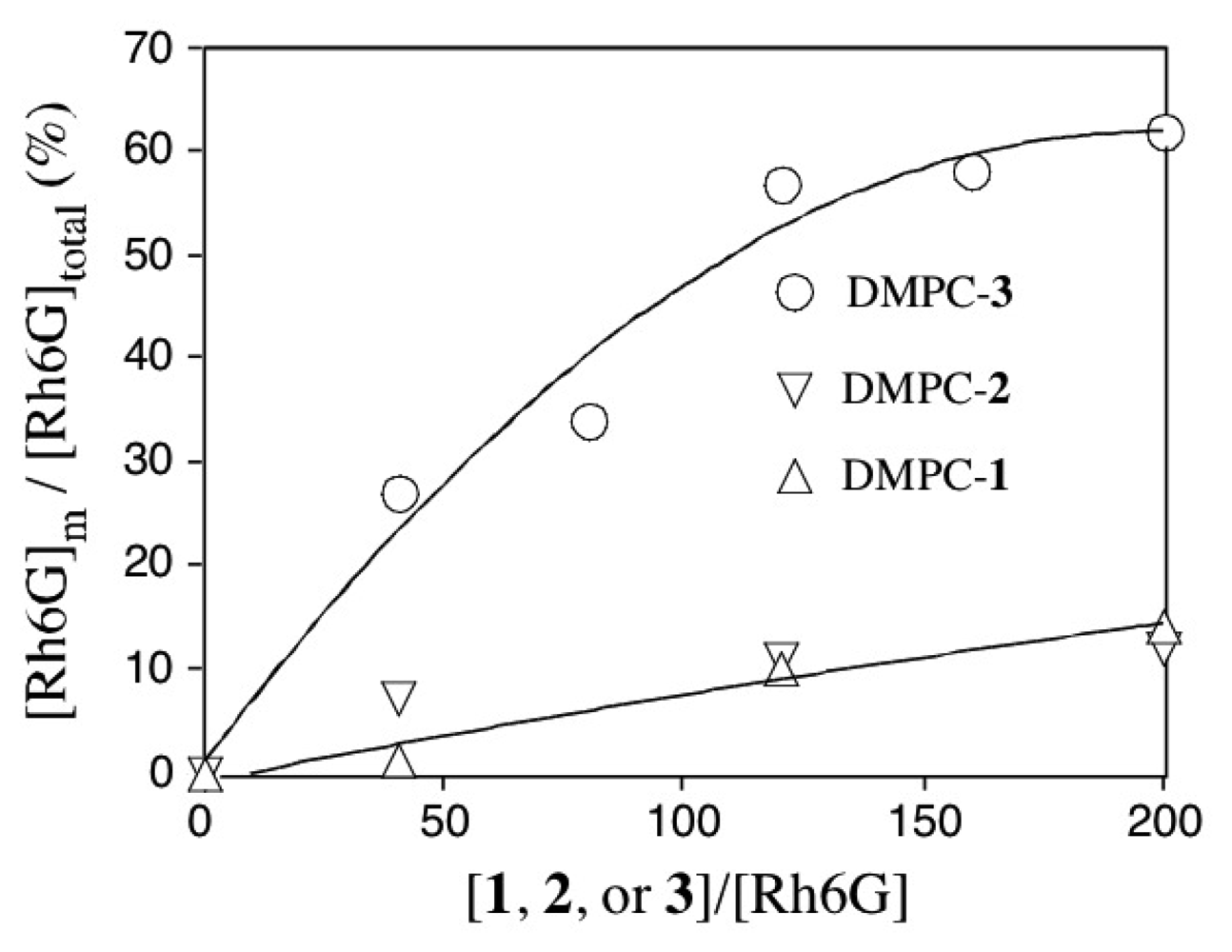
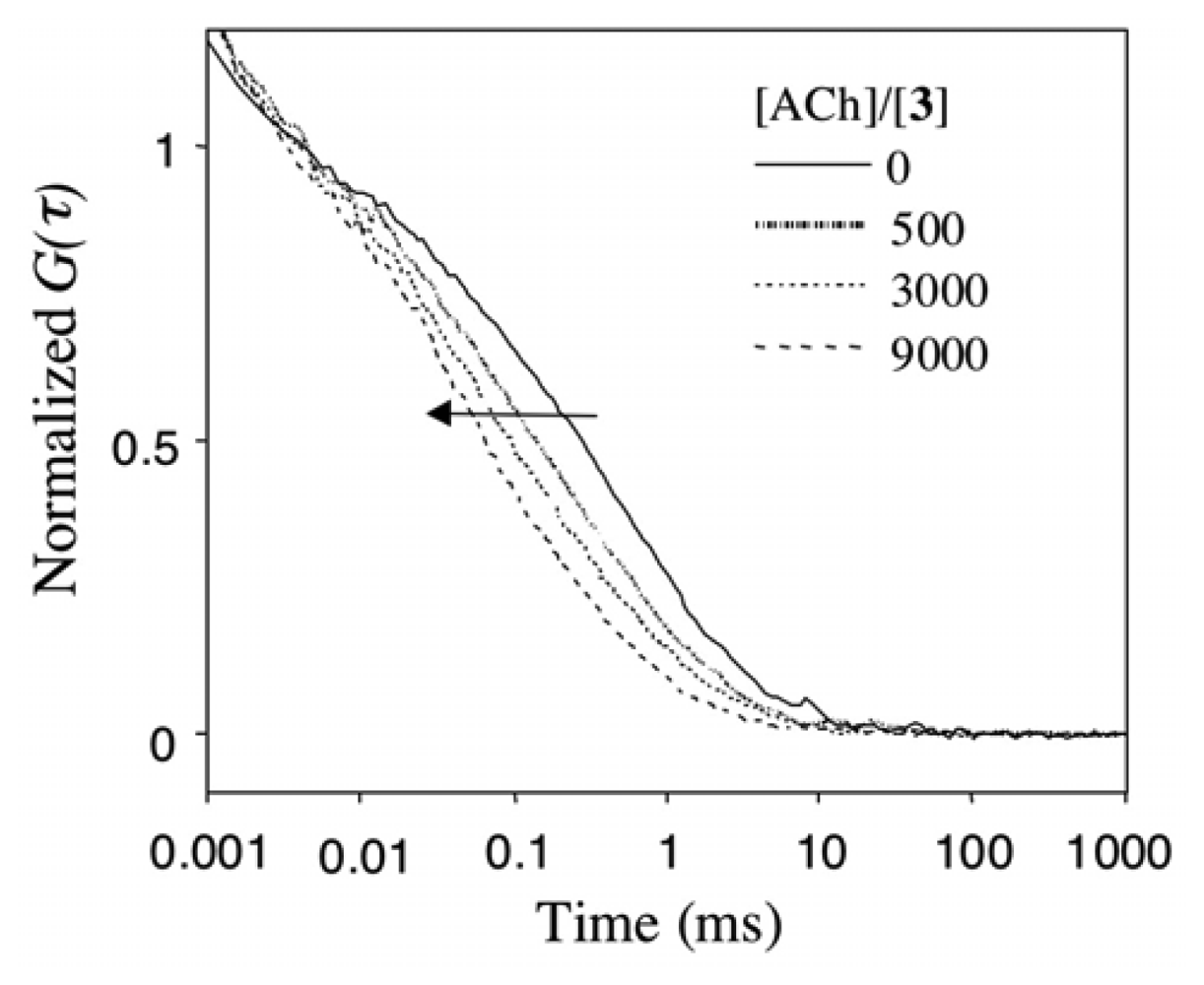
2.4 Fluorescence correlation spectroscopy for the binding of Rh6G and ACh to DMPC vesicles containing 1-3
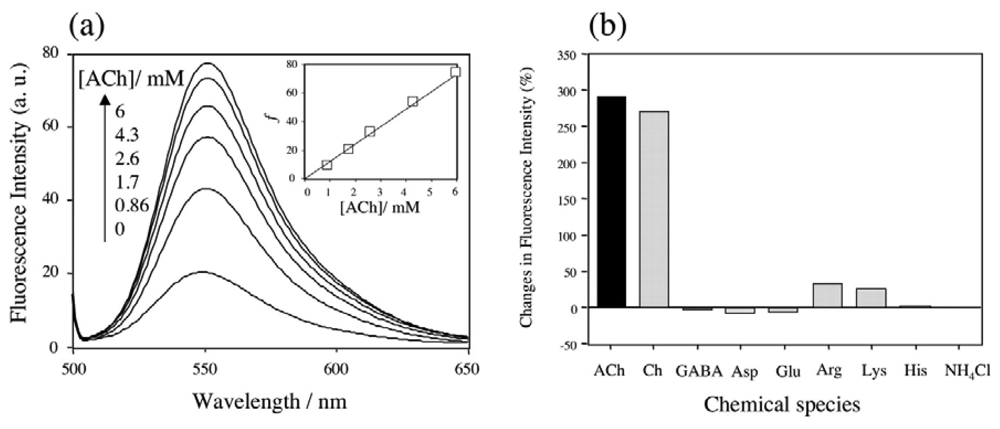
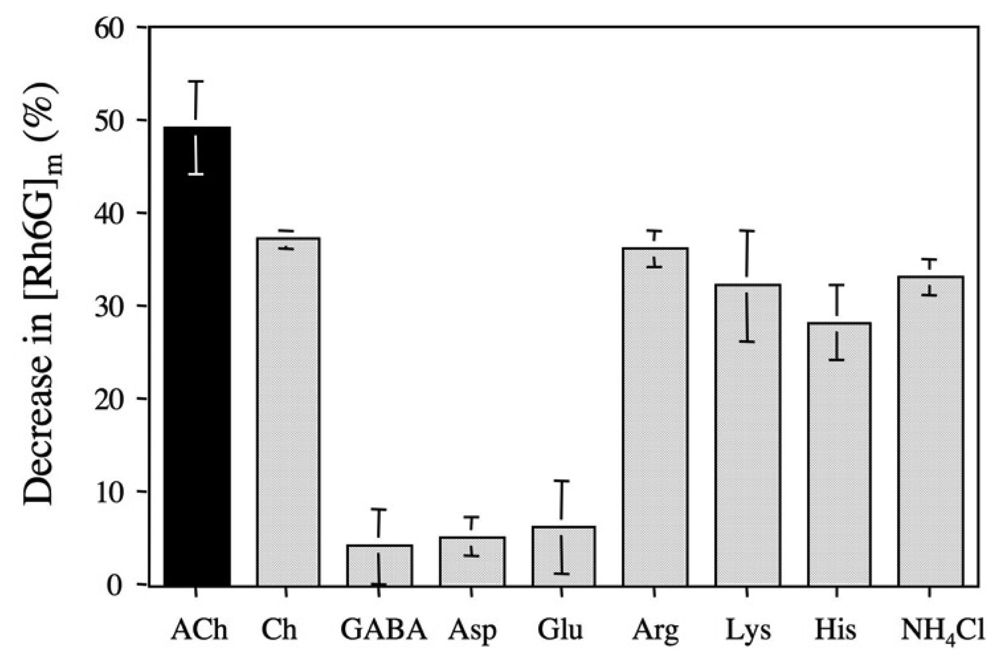
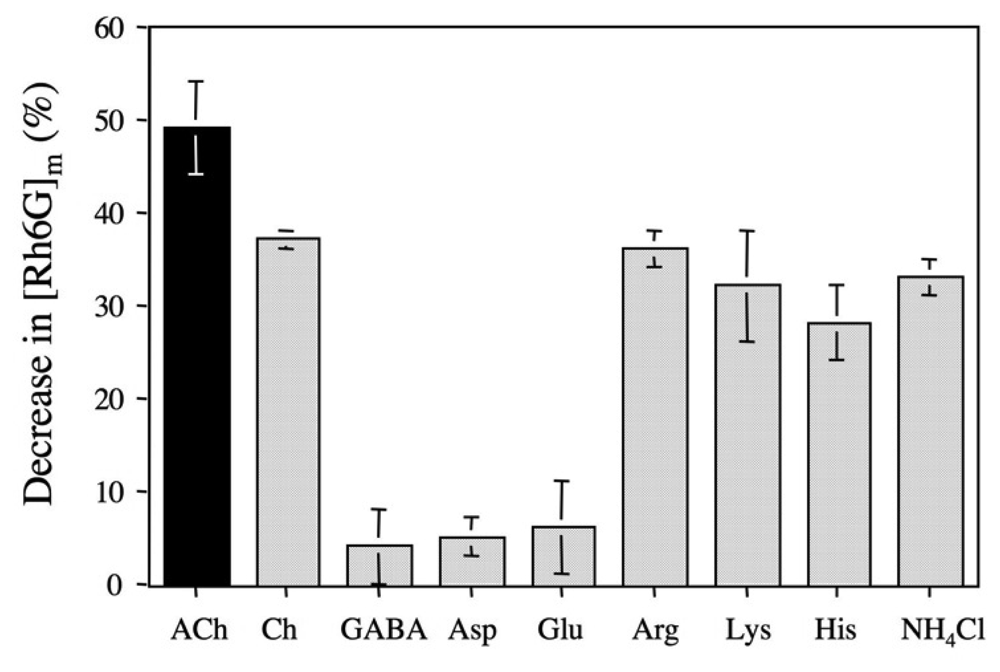
2.5 The binding selectivity of DMPC-3 vesicle
3. Experimental Section
3.1. Materials and Methods
3.2. Synthesis
3.3. Preparation of DMPV vesicles containing 1-3
3.4. Fluorescence spectra and fluorescence anisotropy measurements
3.5. Vesicle size and zeta potential measurements
3.6. Fluorescence autocorrelation curve measurements
4. Conclusions
Acknowledgments
References and Notes
- Albert, B.; Johnson, A.; Lewis, J.; Laff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 5th Edition ed; Garland Science: New York, 2008; Chapter 11; pp. 684–686. [Google Scholar]
- Méric, R; Vigneron, J.-P; Lehn, J.-M. Efficient complexation of quaternary ammonium compounds by a new water-soluble macrobicyclic receptor molecule. Chem. Commun. 1993, 129–131. [Google Scholar]
- Garel, L; Lozach, B; Dutasta, J.-P; Collet, A. Remarkable effect of receptor size in the binding of acetylcholine and related ammonium ions to water-soluble cryptophanes. J. Am. Chem. Soc. 1993, 115, 11652–11653. [Google Scholar]
- Lehn, J.-M; Meric, R; Vigneron, J. -P; Cesario, M; Guilhem, J; Pascard, C; Asfari, Z; Vicens, J. Binding of acetylcholine and other quaternary ammonium cations by sulfonated calixarenes. Crystal structure of a [choline-tetrasulfonated calix[4]arene] complex. Supramol. Chem. 1995, 5, 97–103. [Google Scholar]
- Koh, K.N; Araki, K; Ikeda, A; Otsuka, H; Shinkai, S. Reinvestigation of calixarene-based artificial-signaling acetylcholine receptors useful in neutral aqueous (water/methanol) solution. J. Am. Chem. Soc. 1996, 118, 755–758. [Google Scholar]
- Zhang, Y.-J; Cao, W.-X; Xu, J. Interaction of sulfonated calix[n]arenes with rhodamine B and its application to determine acetylcholine in a real neutral aqueous medium. Chin. J. Chem. 2002, 20, 322–326. [Google Scholar]
- Jin, T. A new fluorometric method for the detection of the neurotransmitter acetylcholine in water using a dansylcholine complex with p-sulfonated calix[8]arene. J. Inclus. Phenom. Macrocycl. Chem. 2003, 45, 195–201. [Google Scholar]
- Wang, L.-H; Guo, D.-S; Chen, Y; Liu, Y. Thermodynamics of interactions between organic ammonium ions and sulfonatocalixarenes. Thermochim. Acta 2006, 443, 132–135. [Google Scholar]
- Bakirci, H; Nau, W.M. Fluorescence regenerations as a signaling principle for choline and carnitine binding: a refined supramolecular sensor system based on a fluorescent azoalkane. Adv. Funct. Mater. 2006, 16, 237–242. [Google Scholar]
- Korbakov, N; Timmerman, P; Lidich, N; Urbach, B; Sa'ar, A; Yitzchaik, S. Acetylcholine detection at micromolar concentrations with the use of an artificial receptor-based fluorescence switch. Langmuir 2008, 24, 2580–2587. [Google Scholar]
- Inouye, M; Hashimoto, K; Isagawa, K. Nondestructive detection of acetylcholine in protic media: artificial-signaling acetylcholine receptors. J. Am. Chem. Soc. 1994, 116, 5517–5518. [Google Scholar]
- Maruyama, K; Aoki, K. Molecular recognition involving multiple cation-π interactions: the inclusion of the acetylcholine trimethylammonium moiety in resorcin[4]arene. Chem. Commun. 1997, 119–120. [Google Scholar]
- Ballester, P; Shivanyuk, A; Far, A.R; Rebek, J., Jr. A synthetic receptor for choline and carnitine. J. Am. Chem. Soc. 2002, 124, 14014–14016. [Google Scholar]
- Hof, F; Trembleau, L; Ullrich, E.C; Rebek, J., Jr. Acetylcholine recognition by a deep, biomimetic pocket. Angew. Chem. Int. Ed. 2003, 42, 3150–3153. [Google Scholar]
- Tan, S.-D; Chen, W.-H; Satake, A; Wang, B; Xu, Z.-L; Kobuke, Y. Tetracyanoresorcin[4]arene as a pH dependent acetylcholine receptor. Org. Biomol. Chem. 2004, 2, 2719–2721. [Google Scholar]
- Shinkai, S; Araki, K; Manabe, O. NMR determination of association constants for calixarene complexes. Evidence for the formation of a 1:2 complex with calix[8]arene. J. Am. Chem. Soc. 1988, 110, 7214–7215. [Google Scholar]
- Shinkai, S; Araki, K; Matsuda, T; Nishiyama, N; Ikeda, H; Takasu, I; Iwamoto, M. NMR and crystallographic studies of a p-sulfonatocalix[4]arene-guest complex. J. Am. Chem. Soc. 1990, 112, 9053–9058. [Google Scholar]
- Arena, G; Casnati, A; Contino, A; Gulino, F. G; Sciotto, D; Ungaro, R. Entropic origin of the sulfonate groups' electrostatic assistance in the complexion of quaternary ammonium cations by water soluble calix[4]arenes. J. Chem. Soc, Perkin Trans. 2 2000, 419–423. [Google Scholar]
- Arduini, A; Demuru, D; Pochini, A; Secchi, A. Recognition of quaternary ammonium cations by calix[4]arene derivatives supported on gold nanoparticles. Chem. Commun. 2005, 645–647. [Google Scholar]
- Perret, F; Lazar, A.N; Coleman, A.W. Biochemistry of the para-sulfonato-calixarenes. Chem. Commun. 2006, 2425–2438. [Google Scholar]
- Jin, T; Kinjo, M; Koyama, T; Kobayashi, Y; Hirata, H. Selective Na+ transport through phospholipid bilayer membrane by a synthetic calix[4]arene carrier. Langumuir 1996, 12, 2684–2689. [Google Scholar]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd Edition ed; Springer: New York, 2006; Chapter 24; pp. 798–840. [Google Scholar]
- Riddell, F.G.; Arumugam, S.; Brophy, P.J.; Cox, B.G.; Payne, M.C.H.; Southon, T.E. The nigericin-mediated transport of sodium and potassium ions through phospholipids bilayers studied by 23Na and 39K nmr spectroscopy. J. Am. Chem. Soc. 1988, 110, 734–738. [Google Scholar]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd Edition ed; Springer: New York, 2006; Chapter 10; pp. 353–381. [Google Scholar]
- G(τ)is defined as <I(t)I(t+ τ)>/<I(t)>2, where I (t)is the fluorescence intensity at the time t.
- Albert, B.; Johnson, A.; Lewis, J.; Laff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 5th Edition ed; Garland Science: New York, 2008; chapter 2; p. 60. [Google Scholar]
- Shinkai, S; Mori, S; Koreishi, H; Tsubaki, T; Manabe, O. Hexasulfonated calix[6]arene derivatives: a new class of catalysts, surfactants, and host molecules. J. Am. Chem. Soc. 1986, 108, 2409–2416. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zeta potentials (mV) | |||
|---|---|---|---|
| DMPC | DMPC-1 | DMPC-2 | DMPC-3 |
| 1.6 ± 0.6 | -10.4±2.3 | -8.2±1.0 | -7.1±0.8 |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jin, T.; Fujii, F.; Ooi, Y. Interfacial Recognition of Acetylcholine by an Amphiphilic p-Sulfonatocalix[8]arene Derivative Incorporated into Dimyristoyl Phosphatidylcholine Vesicles. Sensors 2008, 8, 6777-6790. https://doi.org/10.3390/s8106777
Jin T, Fujii F, Ooi Y. Interfacial Recognition of Acetylcholine by an Amphiphilic p-Sulfonatocalix[8]arene Derivative Incorporated into Dimyristoyl Phosphatidylcholine Vesicles. Sensors. 2008; 8(10):6777-6790. https://doi.org/10.3390/s8106777
Chicago/Turabian StyleJin, Takashi, Fumihiko Fujii, and Yasuhiro Ooi. 2008. "Interfacial Recognition of Acetylcholine by an Amphiphilic p-Sulfonatocalix[8]arene Derivative Incorporated into Dimyristoyl Phosphatidylcholine Vesicles" Sensors 8, no. 10: 6777-6790. https://doi.org/10.3390/s8106777