
Extracting Plant Phenology Metrics in a Great Basin Watershed: Methods and Considerations for Quantifying Phenophases in a Cold Desert

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
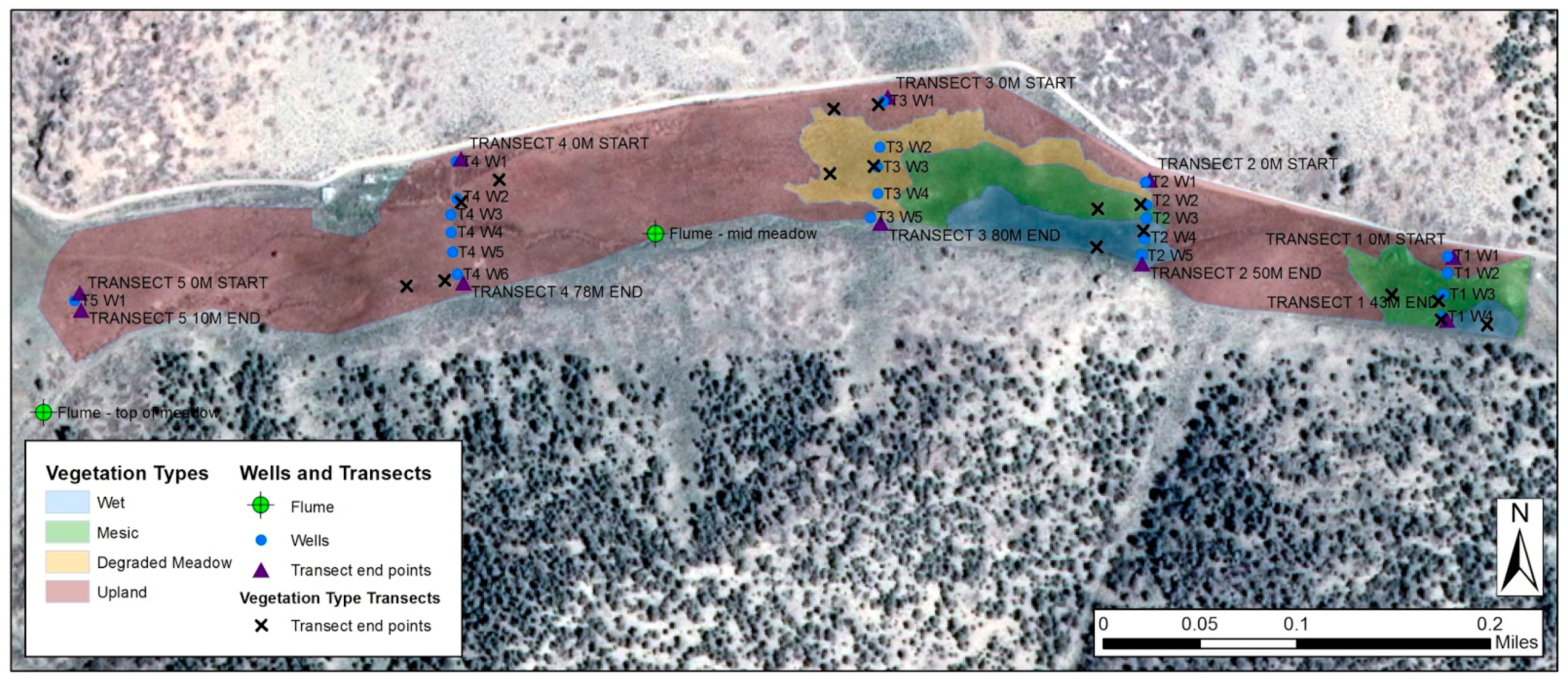
2.1. Study Area
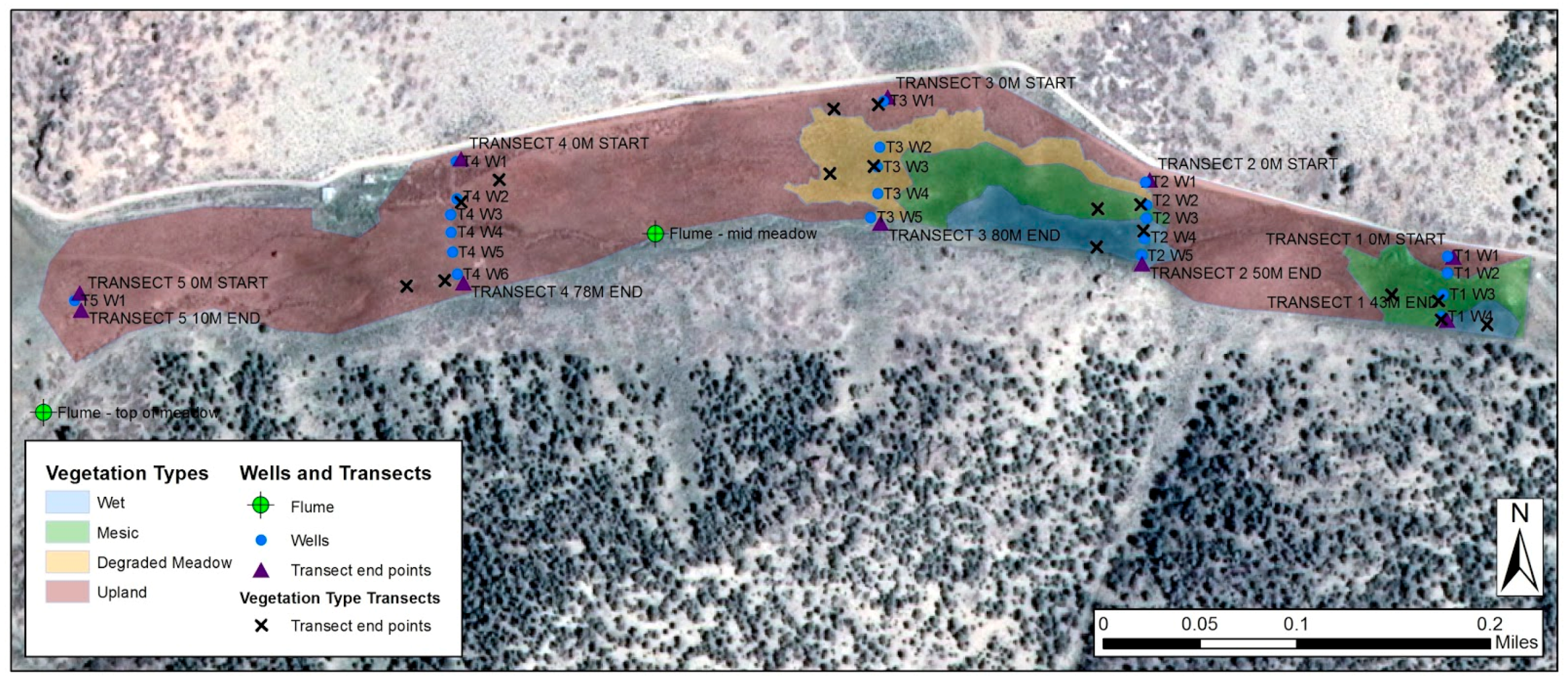
2.2. Camera Sites and Overview
2.3. Vegetation
2.4. Environmental Measurements
2.5. Camera Setup
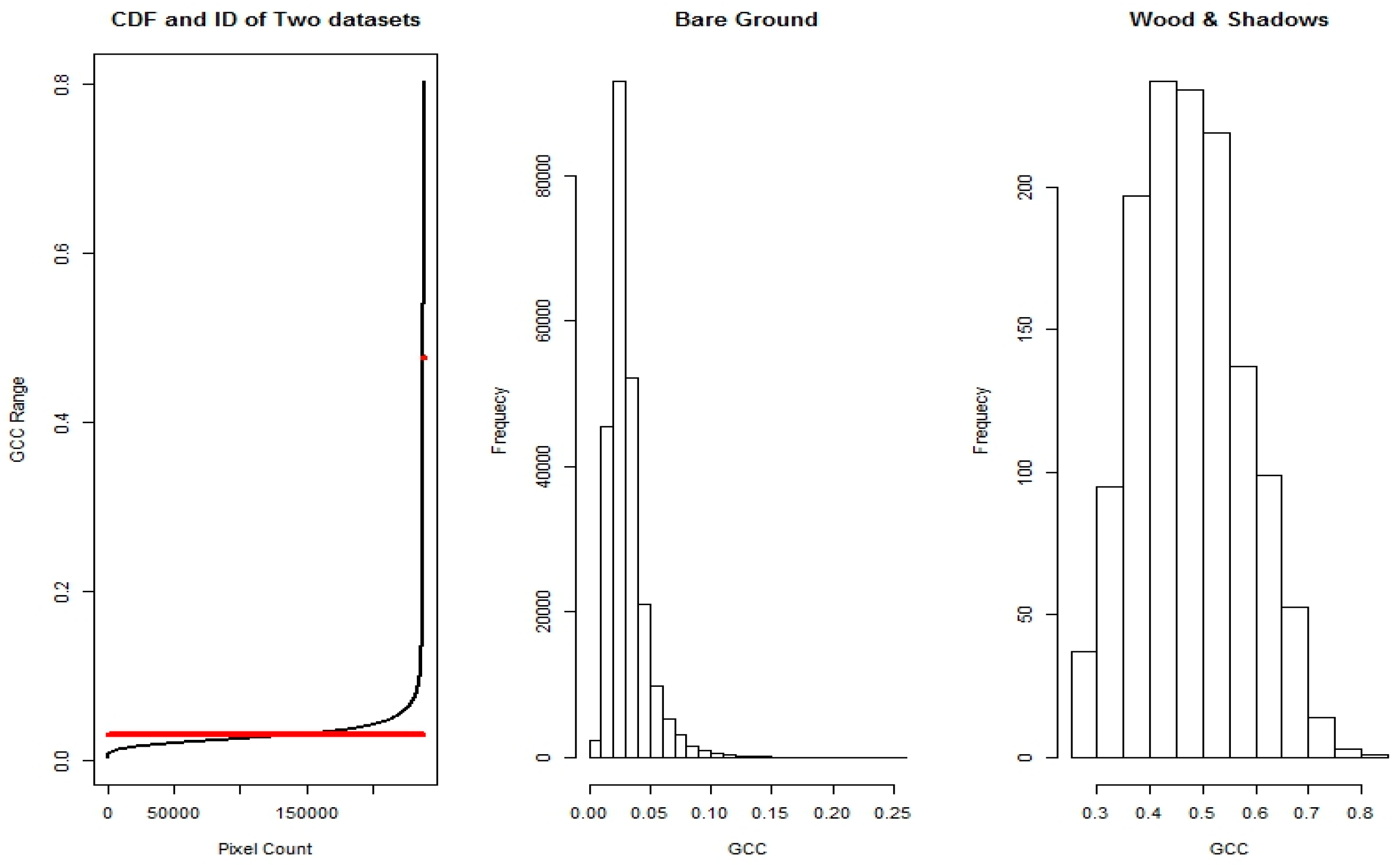
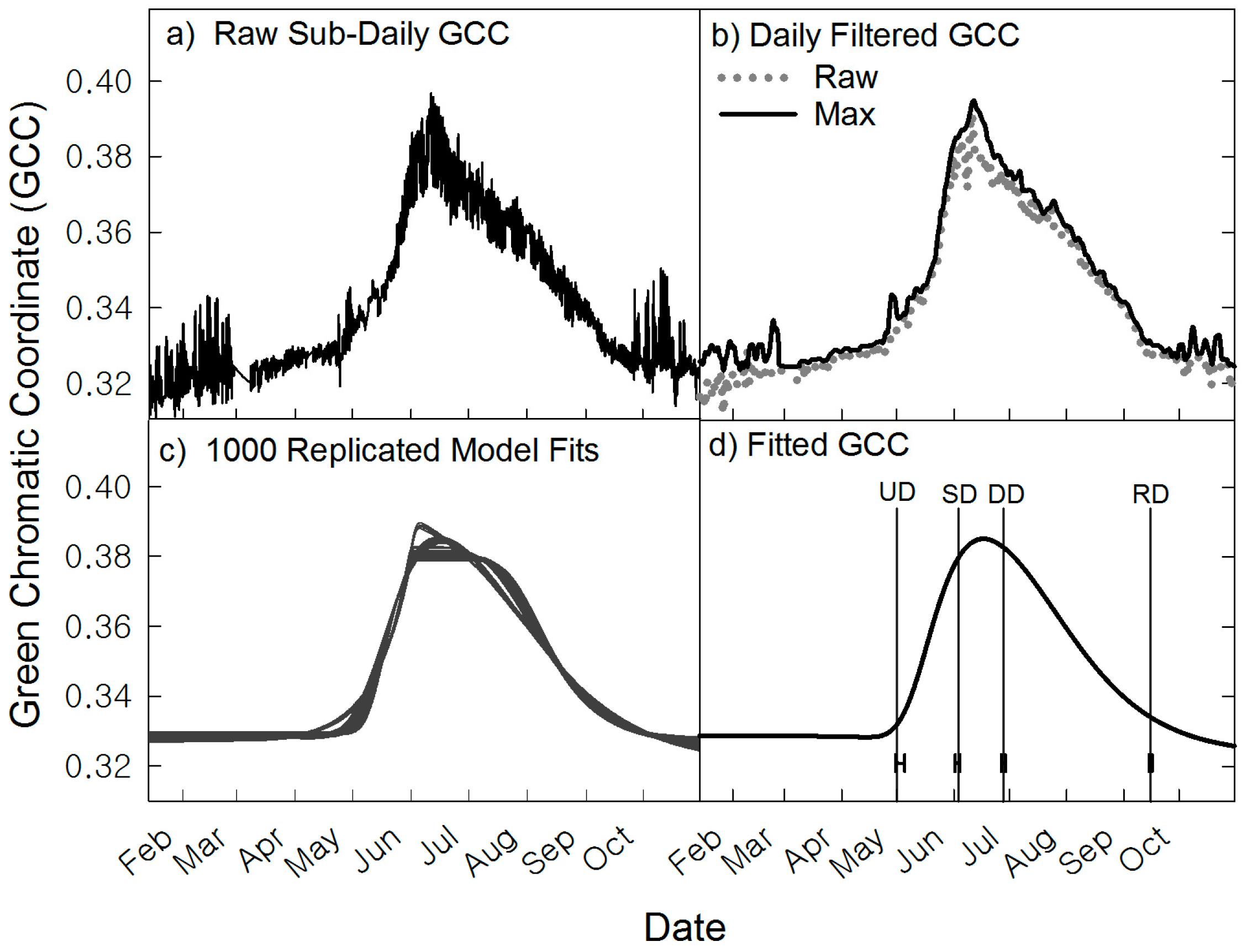
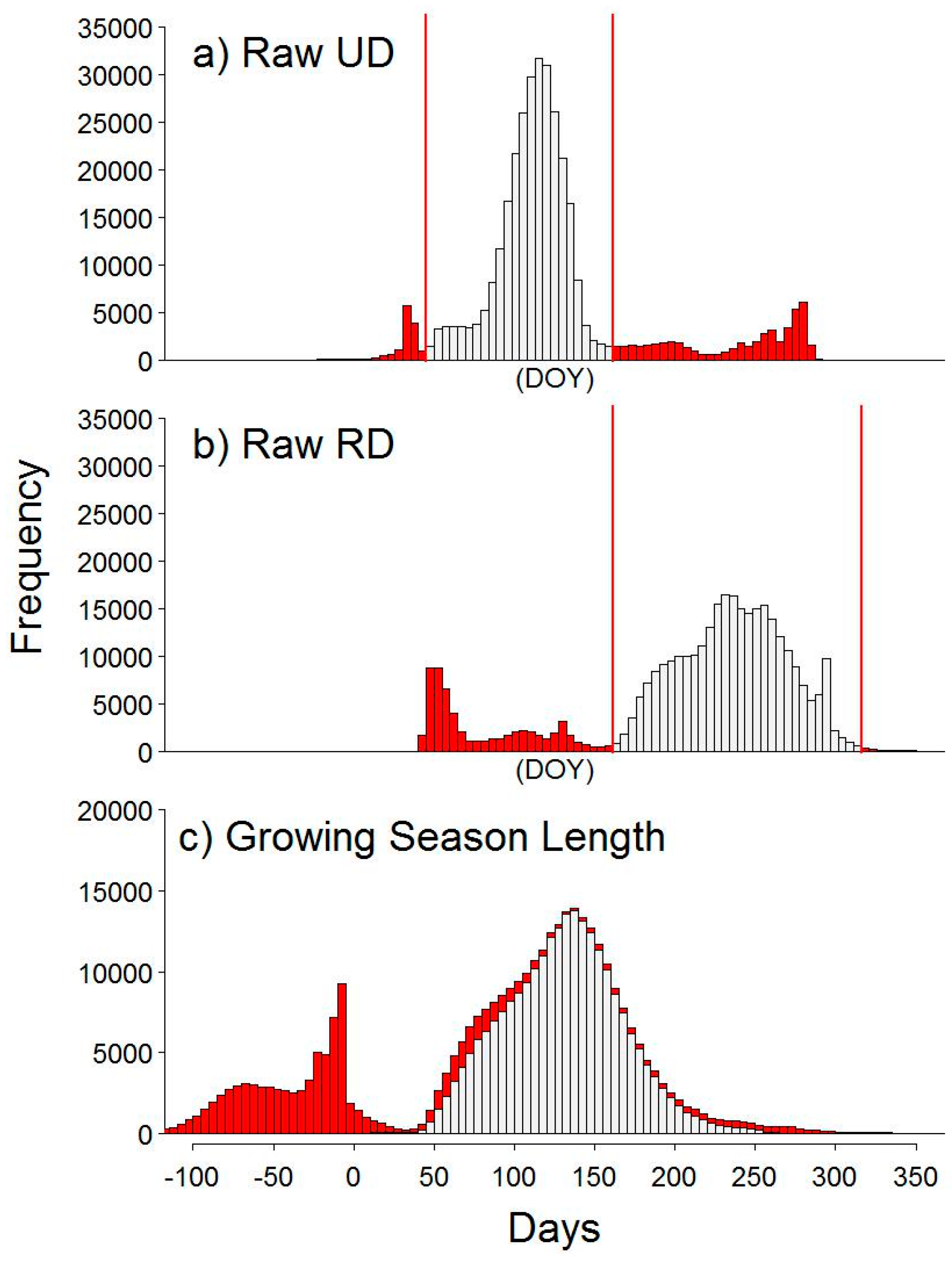
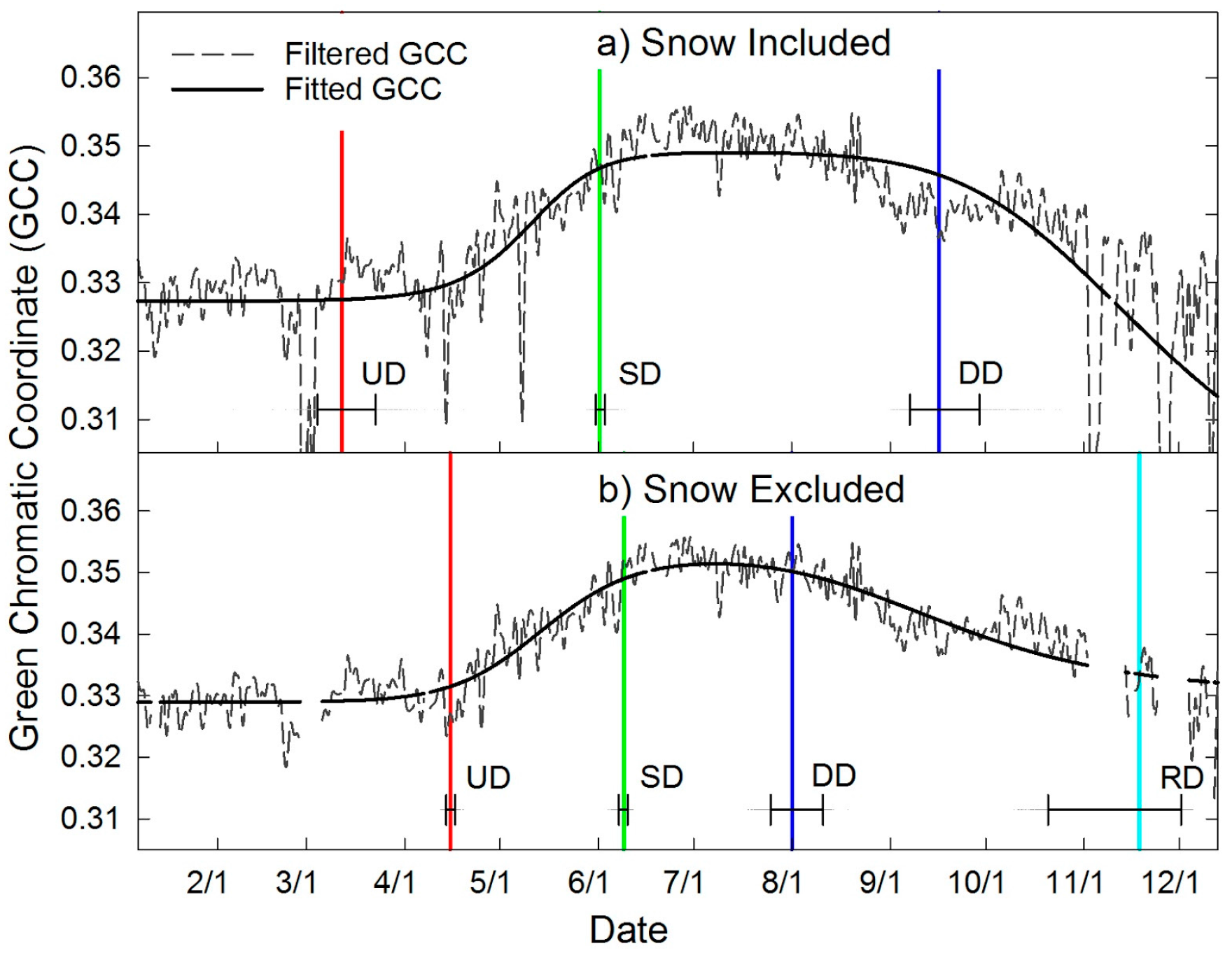
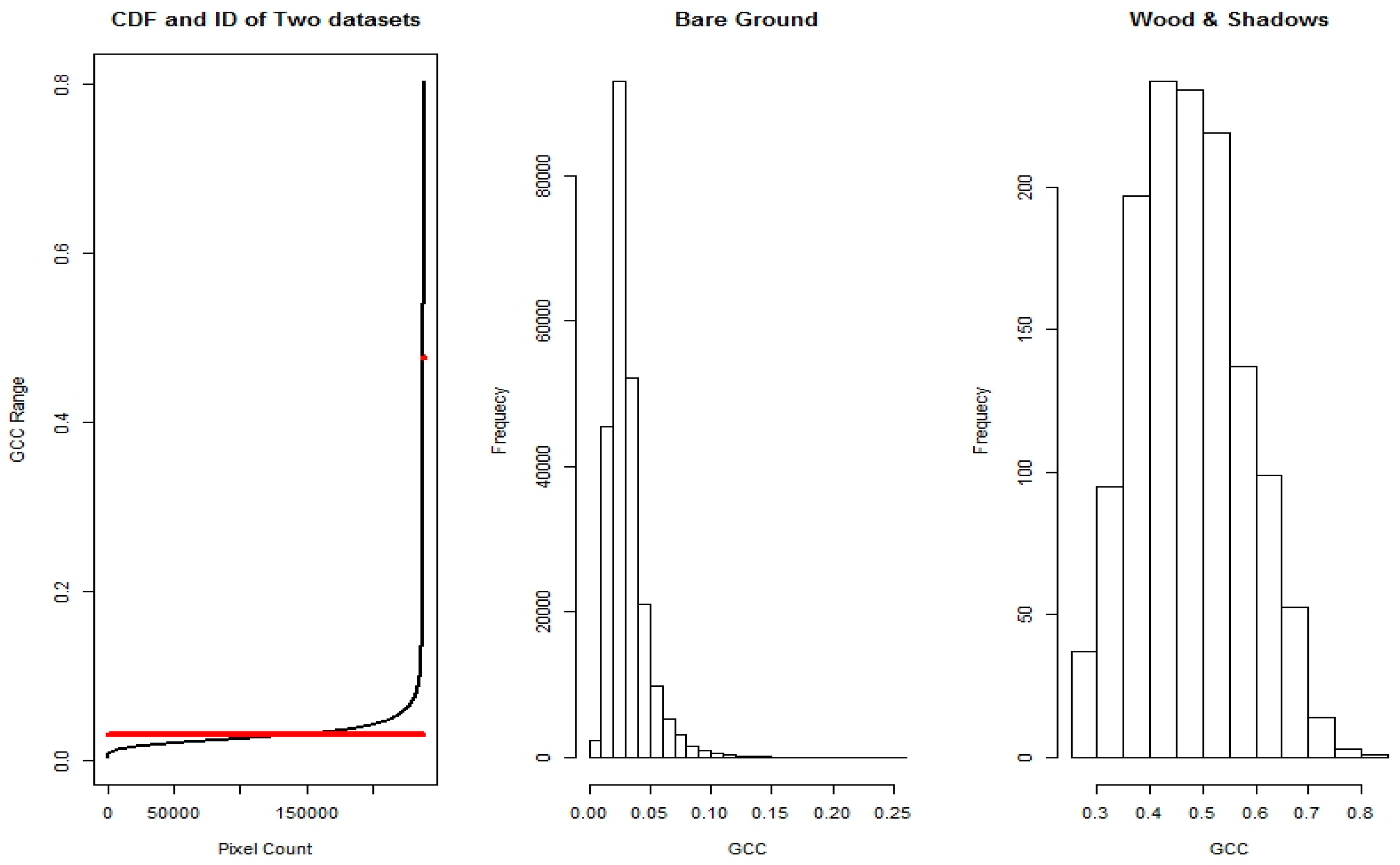
2.6. Image Data Processing
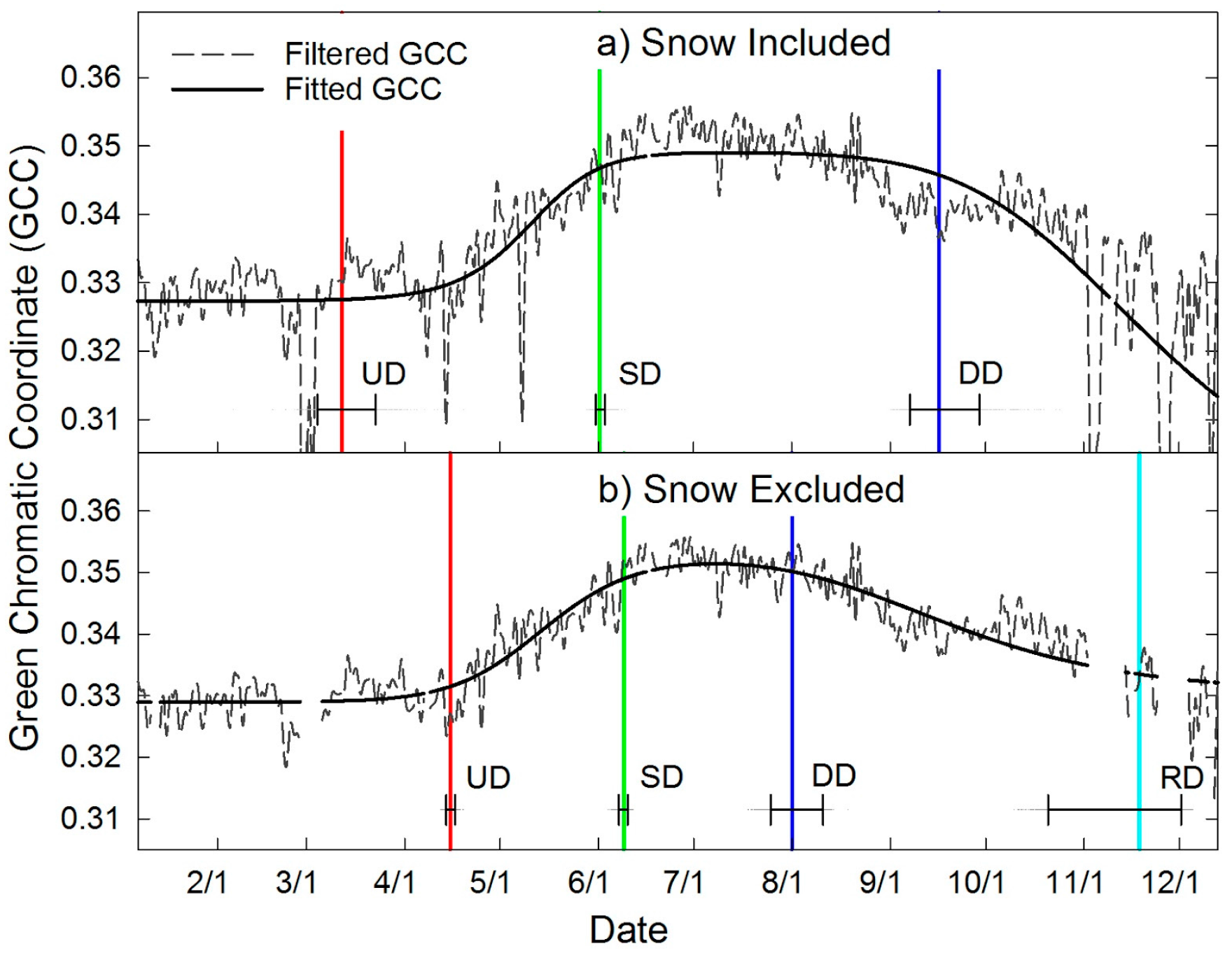
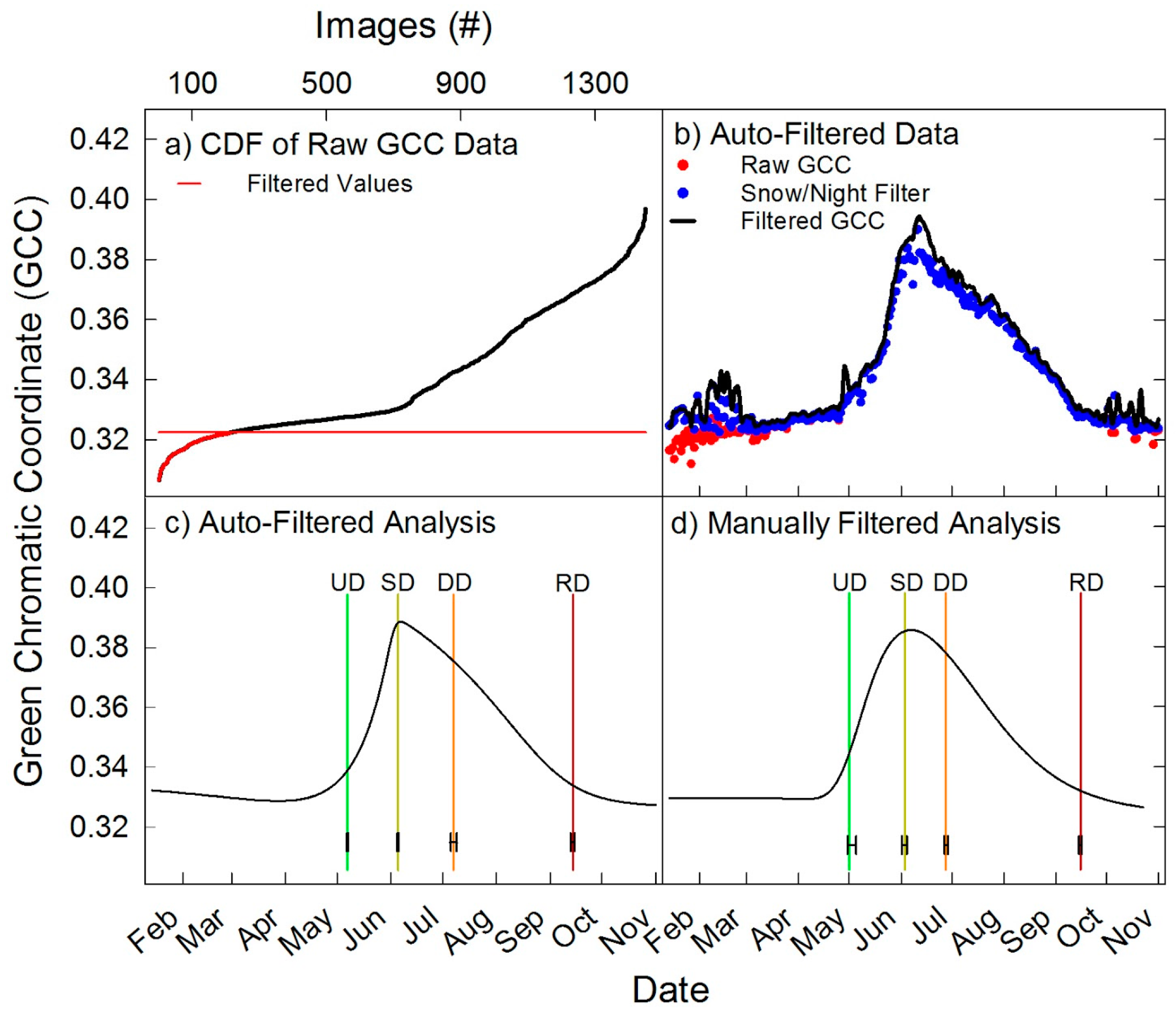
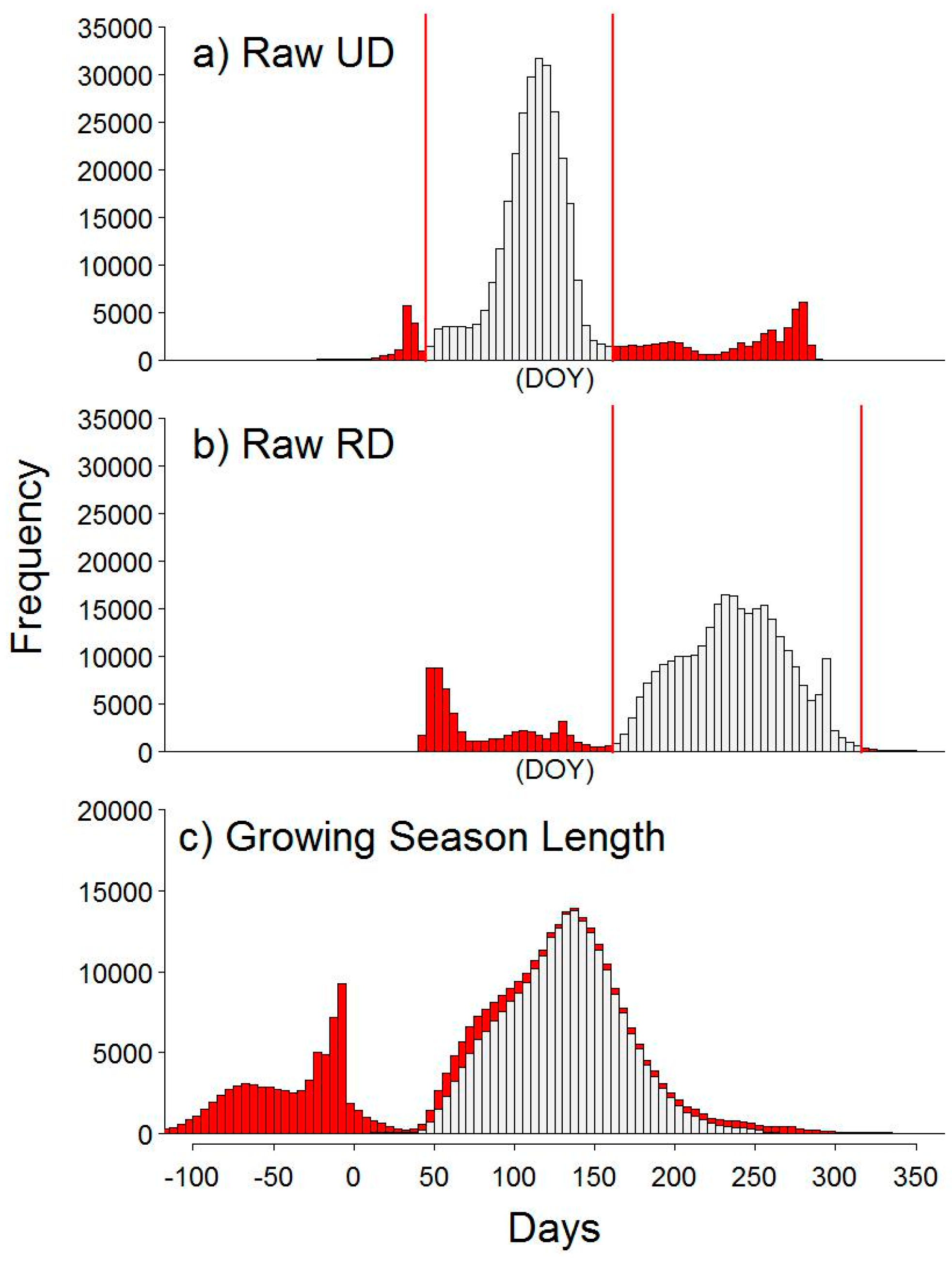
2.7. Automated Snow vs. Manual Snow Filtering
3. Results
3.1. Community Structure
3.2. Averaged ROIs
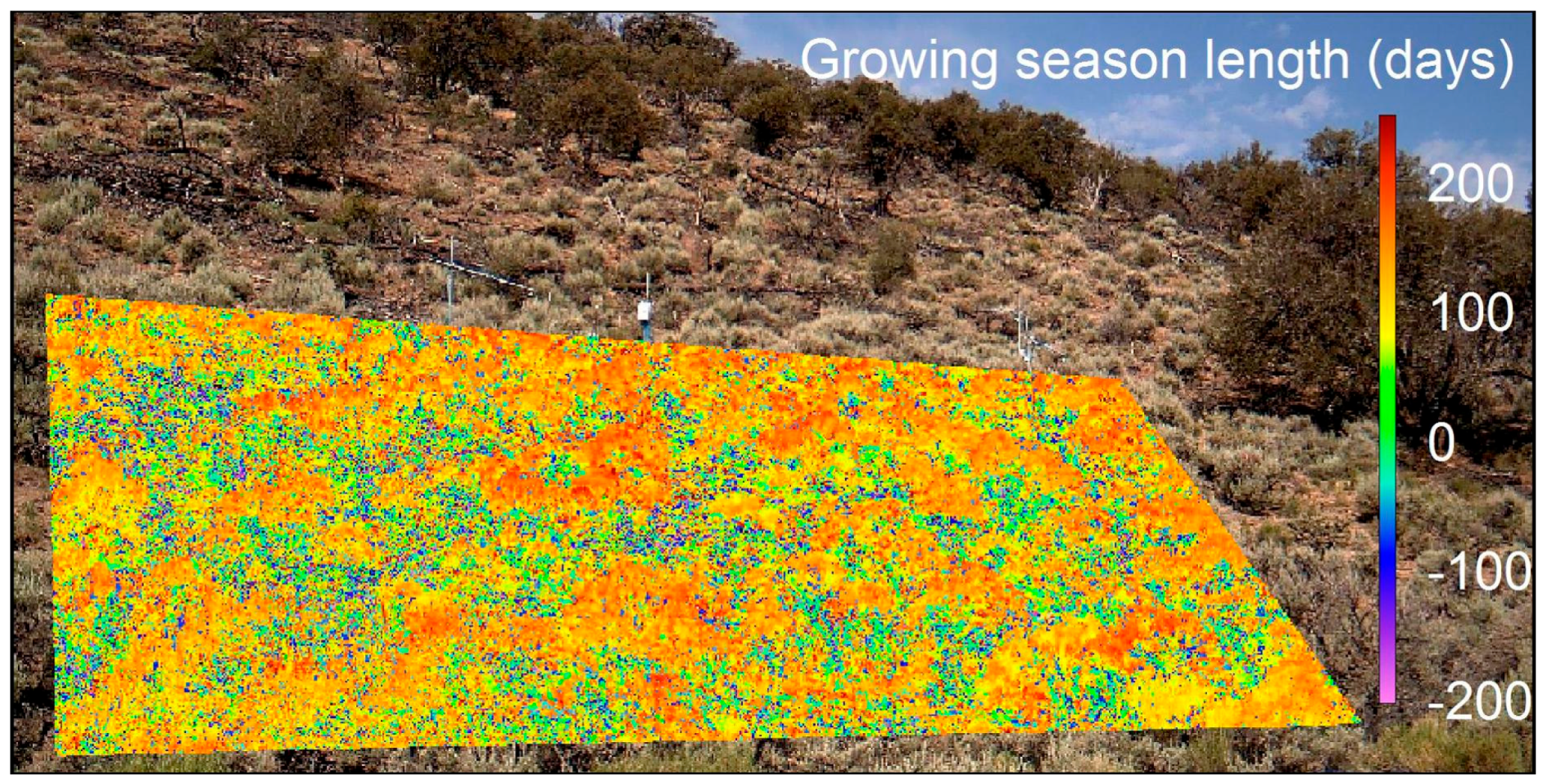
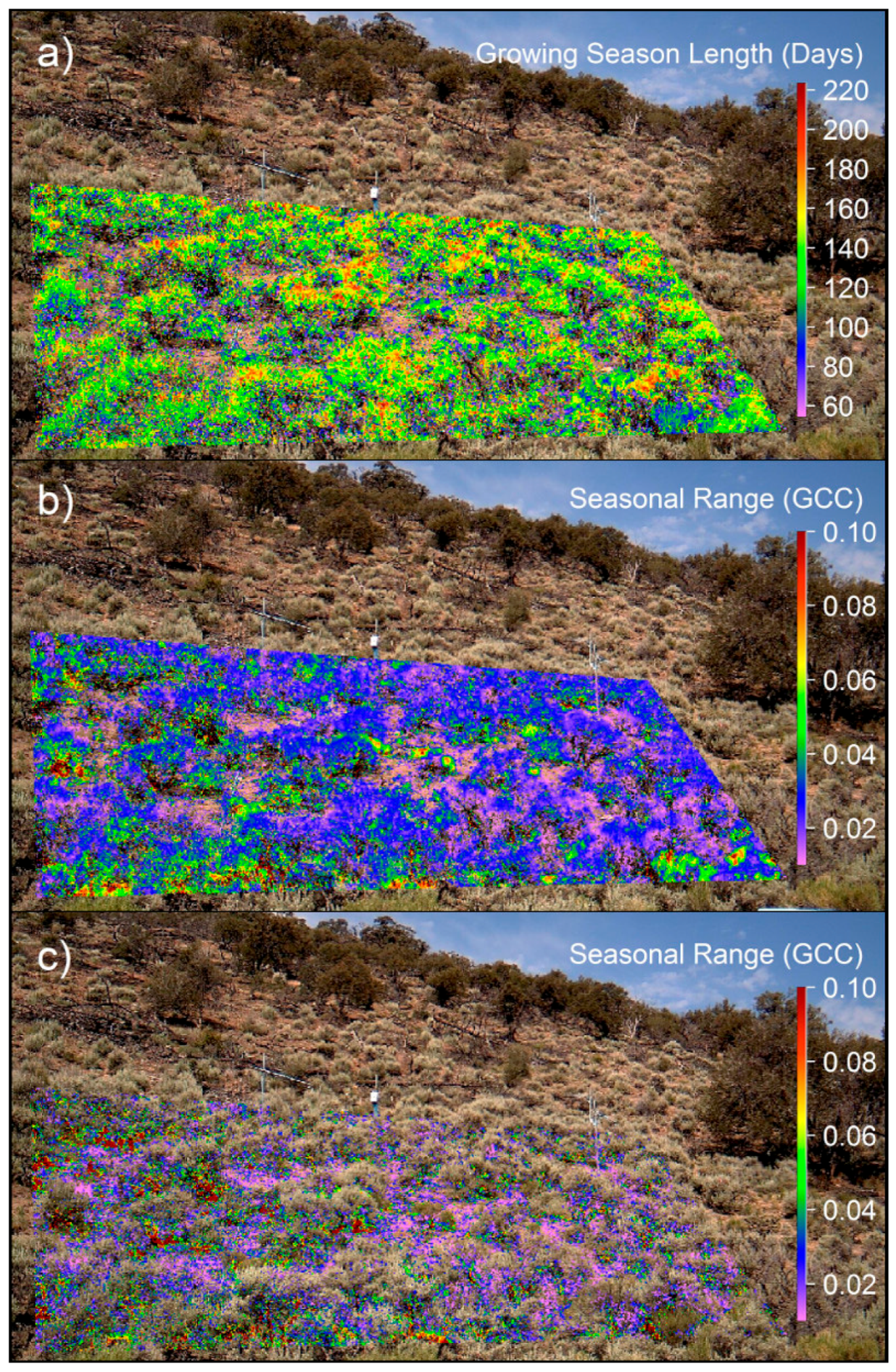
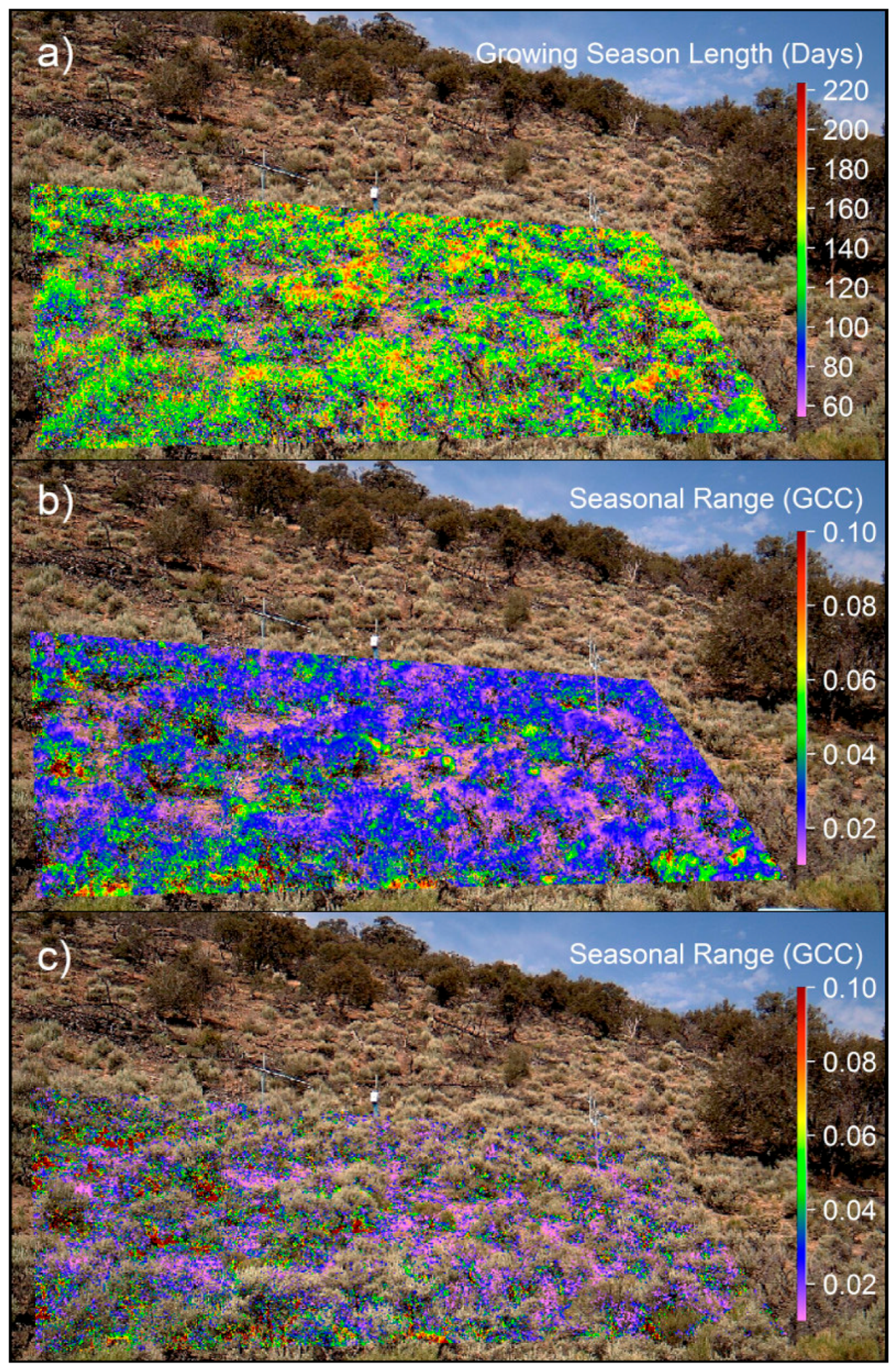
3.3. Spatiotemporal Analyses
4. Discussion
4.1. Methodological Considerations
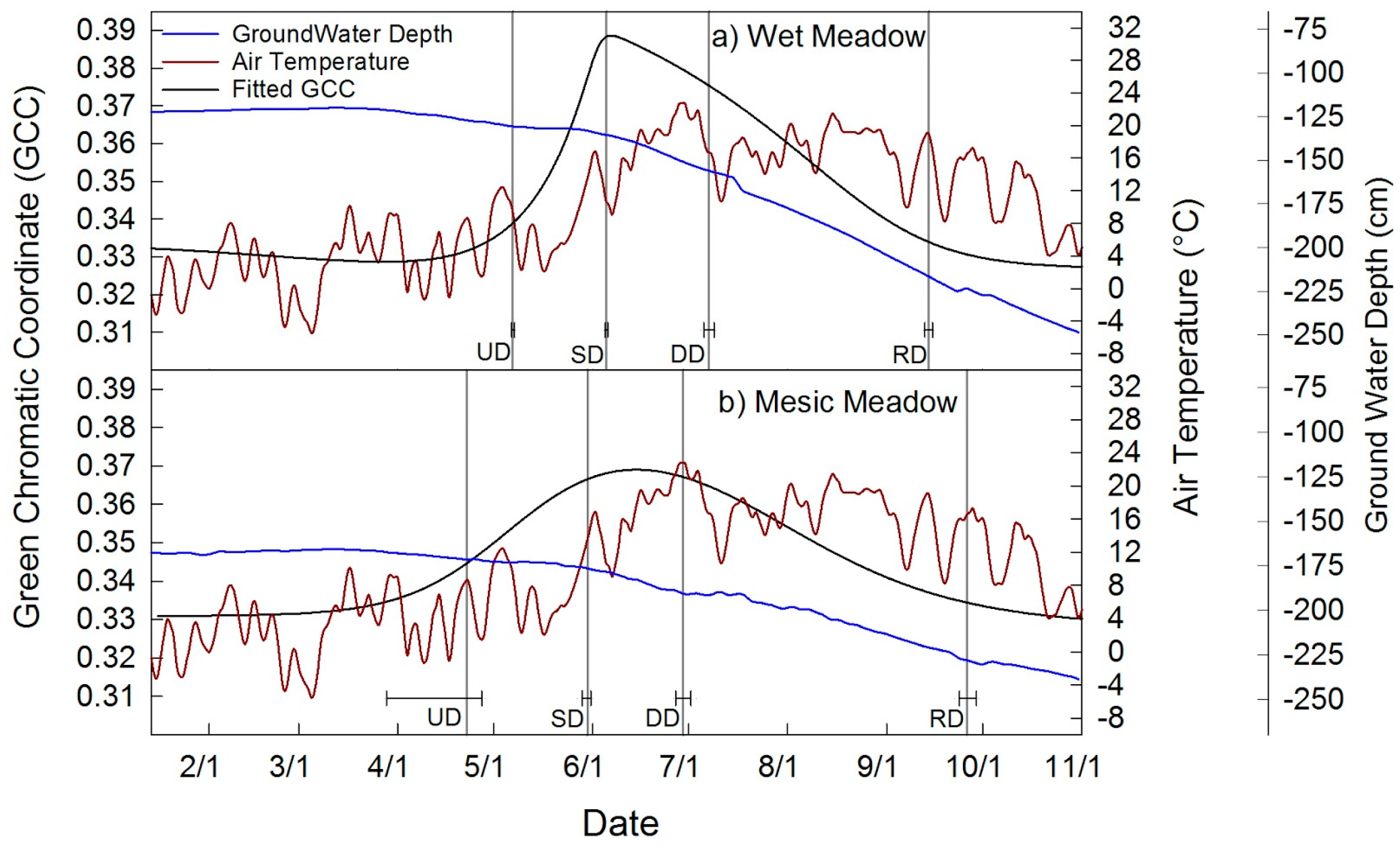
4.2. Meadow Phenology
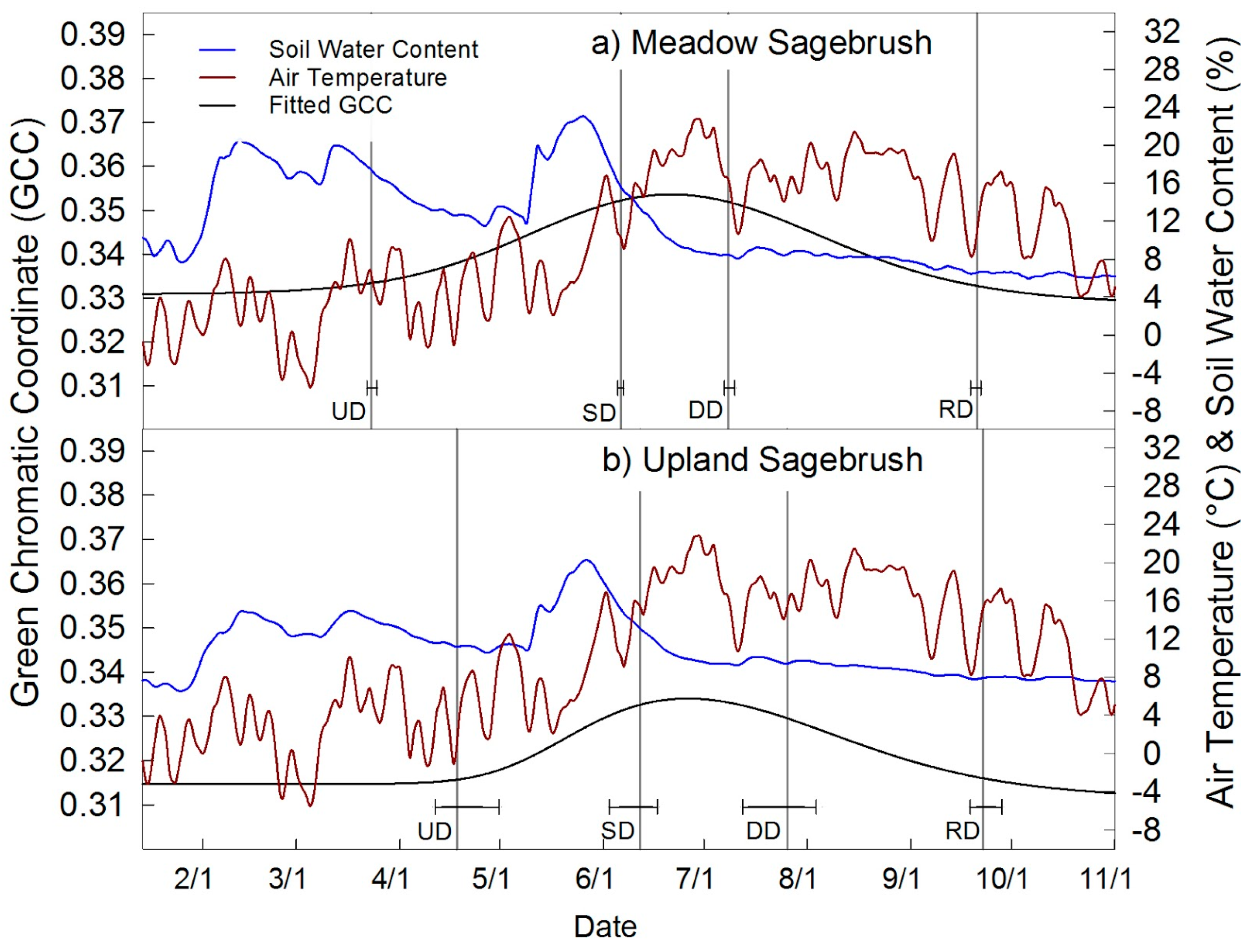
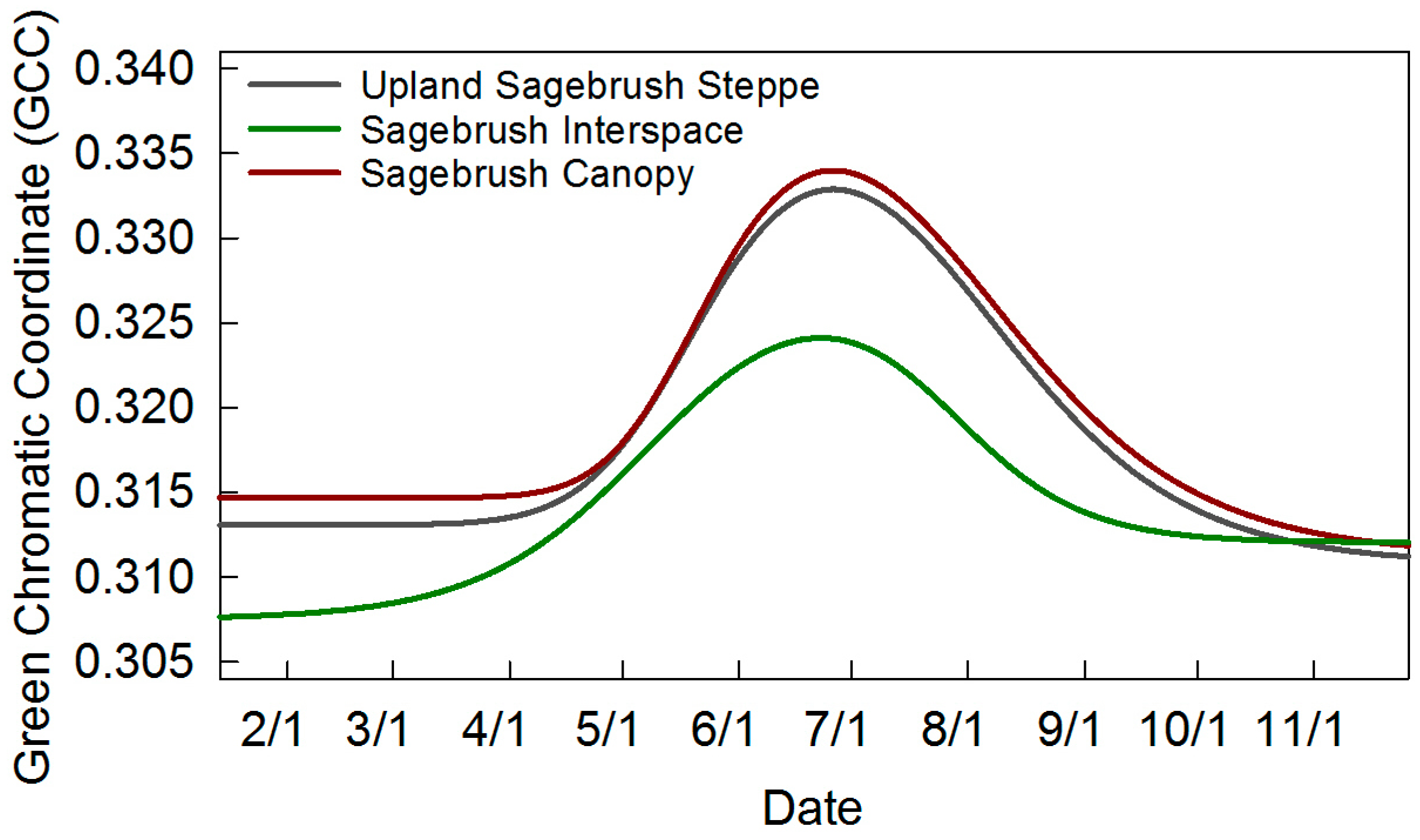
4.3. Sagebrush Phenology
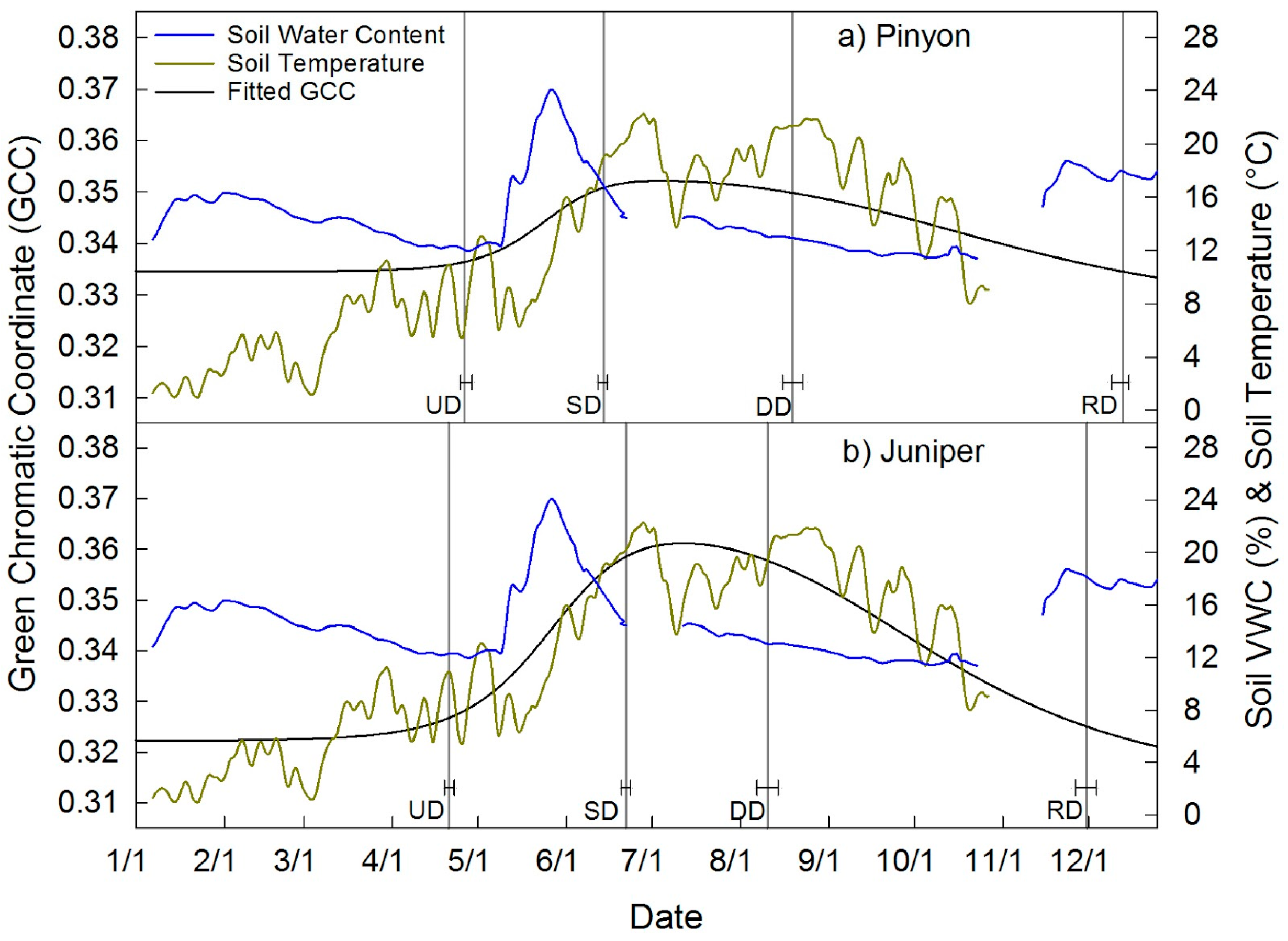
4.4. Pinyon and Juniper Phenology
4.5. Summary
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
References
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Kerr, J.T.; Ostrovsky, M. From space to species: Ecological applications for remote sensing. Trends Ecol. Evol. 2003, 18, 299–305. [Google Scholar] [CrossRef]
- Cohen, W.B.; Goward, S.N. Landsat’s role in ecological applications of remote sensing. Bioscience 2004, 54, 535–545. [Google Scholar] [CrossRef]
- Richardson, A.D.; Jenkins, J.P.; Braswell, B.H.; Hollinger, D.Y.; Ollinger, S.V.; Smith, M.L. Use of digital webcam images to track spring green-up in a deciduous broadleaf forest. Oecologia 2007, 152, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Gitelson, A.A.; Nguy-Robertson, A.L.; Arkebauer, T.J.; Wardlow, B.D.; Suyker, A.E.; Verma, S.B.; Shibayama, M. An alternate method using digital cameras for continuous monitoring of crop status. Agric. For. Meteorol. 2012, 154–155, 113–126. [Google Scholar] [CrossRef]
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Dworak, V.; Selbeck, J.; Dammer, K.; Hoffmann, M.; Zarezadeh, A.A.; Bobda, C. Strategy for the Development of a Smart NDVI Camera System for Outdoor Plant Detection and Agricultural Embedded Systems. Sensors 2013, 13, 1523–1538. [Google Scholar] [CrossRef] [PubMed]
- Filippa, G.; Cremonese, E.; Galvagno, M.; Migliavacca, M.; di Cella, U.M.; Petey, M.; Siniscalco, C. Five years of phenological monitoring in a mountain grassland: Inter-annual patterns and evaluation of the sampling protocol. Int. J. Biometeorol. 2015, 59, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.B.; Hultine, K.R.; Steltzer, H.; Denny, E.G.; Denslow, M.W.; Granados, J.; Henderson, S.; Moore, D.; Nagai, S.; SanClements, M.; et al. Using phenocams to monitor our changing earth: Toward a global phenocam network. Front. Ecol. Environ. 2016, 14, 84–93. [Google Scholar] [CrossRef]
- Baghzouz, M.; Devitt, D.A.; Fenstermaker, L.F.; Young, M.H. Monitoring vegetation phenological cycles in two different semi-arid environmental settings using a ground-based NDVI system: A potential approach to improve satellite data interpretation. Remote Sens. 2010, 2, 990–1013. [Google Scholar] [CrossRef]
- Castelli, R.M.; Chambers, J.C.; Tausch, R.J. Soil-plant relations along a soil-water gradient in great basin riparian meadows. Wetlands 2000, 20, 251–266. [Google Scholar] [CrossRef]
- Reed, B.C.; Schwartz, M.D.; Xiao, X. Remote sensing phenology. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 231–246. [Google Scholar]
- Jacobs, N.; Burgin, W.; Fridrich, N.; Abrams, A.; Miskell, K.; Braswell, B.H.; Richardson, A.D.; Pless, R. The global network of outdoor webcams. In Proceedings of the 17th ACM SIGSPATIAL International Conference on Advanced Geographic Information Systems & Web Services, Seattle, WA, USA, 4–6 November 2009.
- Graham, E.A.; Riordan, E.C.; Yuen, E.M.; Estrin, D.; Rundel, P.W. Public internet-connected cameras used as a cross-continental ground-based plant phenology monitoring system. Glob. Chang. Biol. 2010, 16, 3014–3023. [Google Scholar] [CrossRef]
- Petach, A.R.; Toomey, M.; Aubrecht, D.M.; Richardson, A.D. Monitoring vegetation phenology using an infrared-enabled security camera. Agric. For. Meteorol. 2014, 195, 143–151. [Google Scholar] [CrossRef]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite remote sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Toomey, M.; Friedl, M.A.; Frolking, S.; Hufkens, K.; Klosterman, S.; Sonnentag, O.; Baldocchi, D.D.; Bernacchi, C.J.; Biraud, S.C.; Bohrer, G.; et al. Greenness indices from digital cameras predict the timing and seasonal dynamics of canopy-scale photosynthesis. Ecol. Appl. 2015, 25, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Melaas, E.K.; Friedl, M.A.; Richardson, A.D. Multiscale modeling of spring phenology across deciduous forests in the eastern United States. Glob. Chang. Biol. 2016, 22, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Julitta, T.; Cremonese, E.; Migliavacca, M.; Colombo, R.; Galvagno, M.; Siniscalco, C.; Rossini, M.; Fava, F.; Cogliati, S.; di Cella, U.M.; et al. Using digital camera images to analyse snowmelt and phenology of a subalpine grassland. Agric. For. Meteorol. 2014, 198, 116–125. [Google Scholar] [CrossRef]
- Wu, J.; Albert, L.P.; Lopes, A.P.; Restrepo-Coupe, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. Science 2016, 351, 972–976. [Google Scholar] [CrossRef] [PubMed]
- Kurc, S.A.; Benton, L.M. Digital image-derived greenness links deep soil moisture to carbon uptake in a creosotebush-dominated shrubland. J. Arid Environ. 2010, 74, 585–594. [Google Scholar] [CrossRef]
- Inoue, T.; Nagai, S.; Kobayashi, H.; Koizumi, H. Utilization of ground-based digital photography for the evaluation of seasonal changes in the aboveground green biomass and foliage phenology in a grassland ecosystem. Ecol. Inform. 2015, 25, 1–9. [Google Scholar] [CrossRef]
- Bradley, B.A.; Mustard, J.F. Comparison of phenology trends by land cover class: A case study in the Great Basin, USA. Glob. Chang. Biol. 2008, 14, 334–346. [Google Scholar] [CrossRef]
- Chambers, J.C.; Bradley, B.A.; Brown, C.S.; D’Antonio, C.; Germino, M.J.; Grace, J.B.; Hardegree, S.P.; Miller, R.F.; Pyke, D.A. Resilience to stress and disturbance, and resistance to Bromus tectorum invasion in cold desert shrublands of western north america. Ecosystems 2014, 17, 360–375. [Google Scholar] [CrossRef]
- Chambers, J.C.; Devoe, N.; Evenden, A. Collaborative Management and Research in the Great Basin—Examining the Issues and Developing a Framework for Action; General Technical Report (GTR): Reno, NV, USA, 2006. [Google Scholar]
- Petersen, S.L.; Stringham, T.K. Intercanopy community structure across a heterogeneous landscape in a western juniper-encroached ecosystem. J. Veg. Sci. 2009, 20, 1163–1175. [Google Scholar] [CrossRef]
- Pierson, F.B.; Williams, C.J.; Kormos, P.R.; Hardegree, S.P.; Clark, P.E.; Rau, B.M. Hydrologic vulnerability of sagebrush steppe following pinyon and juniper encroachment. Rangel. Ecol. Manag. 2010, 63, 614–629. [Google Scholar] [CrossRef]
- Kormos, P.R.; Marks, D.; Pierson, F.B.; Williams, C.J.; Hardegree, S.P.; Havens, S.; Hedrick, A.; Bates, J.D.; Svejcar, T.J. Ecosystem Water availability in juniper versus sagebrush snow-dominated rangelands. Rangel. Ecol. Manag. 2016, in press. [Google Scholar] [CrossRef]
- Stringham, T.K.; Krueger, W.C.; Thomas, D.R. Application of non-equilibrium ecology to rangeland riparian zones. J. Range Manag. 2001, 54, 210–217. [Google Scholar] [CrossRef]
- Klos, P.Z.; Link, T.E.; Abatzoglou, J.T. Extent of the rain-snow transition zone in the western U.S. under historic and projected climate. Geophys. Res. Lett. 2014, 41, 4560–4568. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.H.; Rudolf, V.H.W. Phenology, ontogeny and the effects of climate change on the timing of species interactions. Ecol. Lett. 2010, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jett, C.E. Soil Survey of Lander County, Nevada, South Part, USDA-NRCS, 1991, 1. Available online: http://www.nrcs.usda.gov/Internet/FSE_MANUSCRIPTS/nevada/NV768/0/LanderSouth.pdf (accessed on 22 September 2016).
- Soil Survey Staff. Keys to Soil Taxonomy. Available online: http://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs142p2_051546.pdf (accessed on 22 September 2016).
- Schlaepfer, D.R.; Lauenroth, W.K.; Bradford, J.B. Ecohydrological niche of sagebrush ecosystems. Ecohydrology 2012, 5, 453–466. [Google Scholar] [CrossRef]
- Daly, C.; Neilson, R.P.; Phillips, D.L. A Statistical-topographic model for mapping climatological precipitation over mountainous terrain. J. Appl. Meteorol. 1994, 33, 140–158. [Google Scholar] [CrossRef]
- R-Forge Phenopix. Available online: https://r-forge.r-project.org/projects/phenopix/ (accessed on 20 September 2016).
- Filippa, G.; Cremonese, E.; Migliavacca, M.; Galvagno, M.; Forkel, M.; Wingate, L.; Tomelleri, E.; di Cella, U.M.; Richardson, A.D. Phenopix: A R package for image-based vegetation phenology. Agric. For. Meteorol. 2016, 220, 141–150. [Google Scholar] [CrossRef]
- Canfield, R.H. Application of the line interception method in sampling range vegetation. J. For. 1941, 39, 388–394. [Google Scholar]
- PhenoCam Software Tools. Available online: https://phenocam.sr.unh.edu/webcam/tools/ (accessed on 20 September 2016).
- Snyder, K.A.; Stringham, T.K. Meadow community structure and depth to groundwater, a multi-year analyses. Unpublished work, in preparation.
- Migliavacca, M.; Galvagno, M.; Cremonese, E.; Rossini, M.; Meroni, M.; Sonnentag, O.; Cogliati, S.; Manca, G.; Diotri, F.; Busetto, L.; et al. Using digital repeat photography and eddy covariance data to model grassland phenology and photosynthetic CO2 uptake. Agric. For. Meteorol. 2011, 151, 1325–1337. [Google Scholar] [CrossRef]
- Gu, L.; Post, W.M.; Baldocchi, D.D.; Black, T.A.; Suyker, A.E.; Verma, S.B.; Vesala, T.; Wofsy, S.C. Characterizing the seasonal dynamics of plant community photosynthesis across a range of vegetation types. In Phenology of Ecosystem Processes; Springer: New York, NY, USA, 2009. [Google Scholar]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Elmore, A.J.; Guinn, S.M.; Minsley, B.J.; Richardson, A.D. Landscape controls on the timing of spring, autumn, and growing season length in mid-Atlantic forests. Glob. Chang. Biol. 2012, 18, 656–674. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using phenocam imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef]
- Forkel, M.; Migliavacca, M.; Thonicke, K.; Reichstein, M.; Schaphoff, S.; Weber, U.; Carvalhais, N. Codominant water control on global interannual variability and trends in land surface phenology and greenness. Glob. Chang. Biol. 2015, 21, 3414–3435. [Google Scholar] [CrossRef] [PubMed]
- CRAN—Package Strucchange. Available online: https://cran.r-project.org/web/packages/strucchange/index.html (accessed on 20 September 2016).
- NRCS Porter Canyon Station. Available online: http://wcc.sc.egov.usda.gov/nwcc/site?sitenum=2170 (accessed on 20 September 2016).
- CRAN—Package Changepoints. Available online: https://cran.r-project.org/web/packages/changepoint/index.html (accessed on 20 September 2016).
- Gu, L.; Baldocchi, D.; Verma, S.B.; Black, T.A.; Vesala, T.; Falge, E.M.; Dowty, P.R. Advantages of diffuse radiation for terrestrial ecosystem productivity. J. Geophys. Res. Atmos. 2002, 107. [Google Scholar] [CrossRef]
- Hufkens, K.; Keenan, T.F.; Flanagan, L.B.; Scott, R.L.; Bernacchi, C.J.; Joo, E.; Brunsell, N.A.; Verfaillie, J.; Richardson, A.D. Productivity of North Amersican grasslands is increased under future climate scenarios despite rising aridity. Nat. Clim. Chang. 2016, 6, 710–714. [Google Scholar] [CrossRef]
- Westergaard-Nielsen, A.; Lund, M.; Hansen, B.U.; Tamstorf, M.P. Camera derived vegetation greenness index as proxy for gross primary production in a low arctic wetland area. ISPRS J. Photogram. Remote Sens. 2013, 86, 89–99. [Google Scholar] [CrossRef]
- Ratliff, R.D.; Westfall, S.E. Nebraska sedge (carex-nebrascensis dewey)—Phenology and life-history at tule meadow, sierra national forest, california. J. Range Manag. 1992, 45, 210–215. [Google Scholar] [CrossRef]
- Engineer Research and Development Center, Environmental Lab, Vicksburg, MS. Regional supplement to the corps of engineers wetland delineation manual: Arid west region. Water Environ. Technol. 2011, 804, 25–28. [Google Scholar]
- Sala, A.; Nowak, R.S. Ecophysiological responses of three riparian graminoids to changes in the soil water table. Int. J. Plant Sci. 1997, 158, 835–843. [Google Scholar] [CrossRef]
- Williams, D.G.; Ehleringer, J.R. Intra- and interspecific variation for summer precipitation use in pinyon-juniper woodlands. Ecol. Monogr. 2000, 70, 517–537. [Google Scholar]
- Bradley, B.A.; Jacob, R.W.; Hermance, J.F.; Mustard, J.F. A curve fitting procedure to derive inter-annual phenologies from time series of noisy satellite NDVI data. Remote Sens. Environ. 2007, 106, 137–145. [Google Scholar] [CrossRef]
- Bradley, B.A.; Mustard, J.F. Identifying land cover variability distinct from land cover change: Cheatgrass in the Great Basin. Remote Sens. Environ. 2005, 94, 204–213. [Google Scholar] [CrossRef]
- Mote, P.W.; Hamlet, A.F.; Clark, M.P.; Lettenmaier, D.P. Declining mountain snowpack in western North America. Bull. Am. Meteorol. Soc. 2005, 86, 39–49. [Google Scholar] [CrossRef]
- NOAA National Centers for Environmental Information. Climate at a Glance: U.S. Time Series. Available online: http://www.ncdc.noaa.gov/cag/ (accessed on 17 August 2016).
- Sonnentag, O.; Detto, M.; Vargas, R.; Ryu, Y.; Runkle, B.R.K.; Kelly, M.; Baldocchi, D.D. Tracking the structural and functional development of a perennial pepperweed (Lepidium latifolium L.) infestation using a multi-year archive of webcam imagery and eddy covariance measurements. Agric. For. Meteorol. 2011, 151, 916–926. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude (dd) | Longitude (dd) | Area (m2) |
|---|---|---|---|
| Wet Meadow | 39.46587 N | 117.613008 W | 300 |
| Mesic Meadow | 39.46605 N | 117.612967 W | 168 |
| Meadow Sagebrush | 39.46623 N | 117.612894 W | 110 |
| Upland Sagebrush Steppe | 39.46599 N | 117.622114 W | 528 |
| Upland PJ Canopy | 39.46333 N | 117.623339 W | 72 per spp. |
| NRCS SNOTEL | 39.46542 N | 117.620694 W | - |
| Site | UD | SD | DD | RD | GSL | Min | Max | Range | Figure |
|---|---|---|---|---|---|---|---|---|---|
| DOY ± Days | Days | GCC | # | ||||||
| Wet Meadow | 127 ± 0.5 | 156 ± 0.5 | 188 ± 2 | 257 ± 1 | 130 ± 1 | 0.327 | 0.389 | 0.062 | 4a |
| Mesic Meadow | 113 ± 15 | 150 ± 1 | 180 ± 2 | 269 ± 3 | 157 ± 14 | 0.329 | 0.369 | 0.040 | 4b |
| Meadow Sagebrush Canopy | 82 ± 1 | 157 ± 1 | 189 ± 2 | 264 ± 2 | 181 ± 2 | 0.329 | 0.354 | 0.025 | 5a |
| Upland Sagebrush Canopy | 108 ± 10 | 163 ± 7 | 207 ± 11 | 265 ± 5 | 157 ± 14 | 0.312 | 0.334 | 0.022 | 5b, 7 |
| Upland Pinyon | 116 ± 2 | 165 ± 2 | 231 ± 4 | 347 ± 3 | 230 ± 4 | 0.333 | 0.352 | 0.019 | 6a |
| Upland Juniper | 111 ± 2 | 173 ± 2 | 223 ± 4 | 334 ± 4 | 224 ± 4 | 0.321 | 0.361 | 0.040 | 6b |
| Upland Sagebrush Steppe | 99 ± 22 | 167 ± 17 | 194 ± 22 | 269 ± 8 | 170 ± 28 | 0.311 | 0.333 | 0.022 | 7 |
| Upland Sagebrush Interspace | 119 ± 24 | 144 ± 14 | 223 ± 5 | 265 ± 10 | 146 ± 15 | 0.308 | 0.324 | 0.016 | 7 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snyder, K.A.; Wehan, B.L.; Filippa, G.; Huntington, J.L.; Stringham, T.K.; Snyder, D.K. Extracting Plant Phenology Metrics in a Great Basin Watershed: Methods and Considerations for Quantifying Phenophases in a Cold Desert. Sensors 2016, 16, 1948. https://doi.org/10.3390/s16111948
Snyder KA, Wehan BL, Filippa G, Huntington JL, Stringham TK, Snyder DK. Extracting Plant Phenology Metrics in a Great Basin Watershed: Methods and Considerations for Quantifying Phenophases in a Cold Desert. Sensors. 2016; 16(11):1948. https://doi.org/10.3390/s16111948
Chicago/Turabian StyleSnyder, Keirith A., Bryce L. Wehan, Gianluca Filippa, Justin L. Huntington, Tamzen K. Stringham, and Devon K. Snyder. 2016. "Extracting Plant Phenology Metrics in a Great Basin Watershed: Methods and Considerations for Quantifying Phenophases in a Cold Desert" Sensors 16, no. 11: 1948. https://doi.org/10.3390/s16111948
APA StyleSnyder, K. A., Wehan, B. L., Filippa, G., Huntington, J. L., Stringham, T. K., & Snyder, D. K. (2016). Extracting Plant Phenology Metrics in a Great Basin Watershed: Methods and Considerations for Quantifying Phenophases in a Cold Desert. Sensors, 16(11), 1948. https://doi.org/10.3390/s16111948