Deposition of Bacteriorhodopsin Protein in a Purple Membrane Form on Nitrocellulose Membranes for Enhanced Photoelectric Response

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental
2.1. Chemicals
2.2. PM preparation
2.3. Fabrication of pH Electrodes
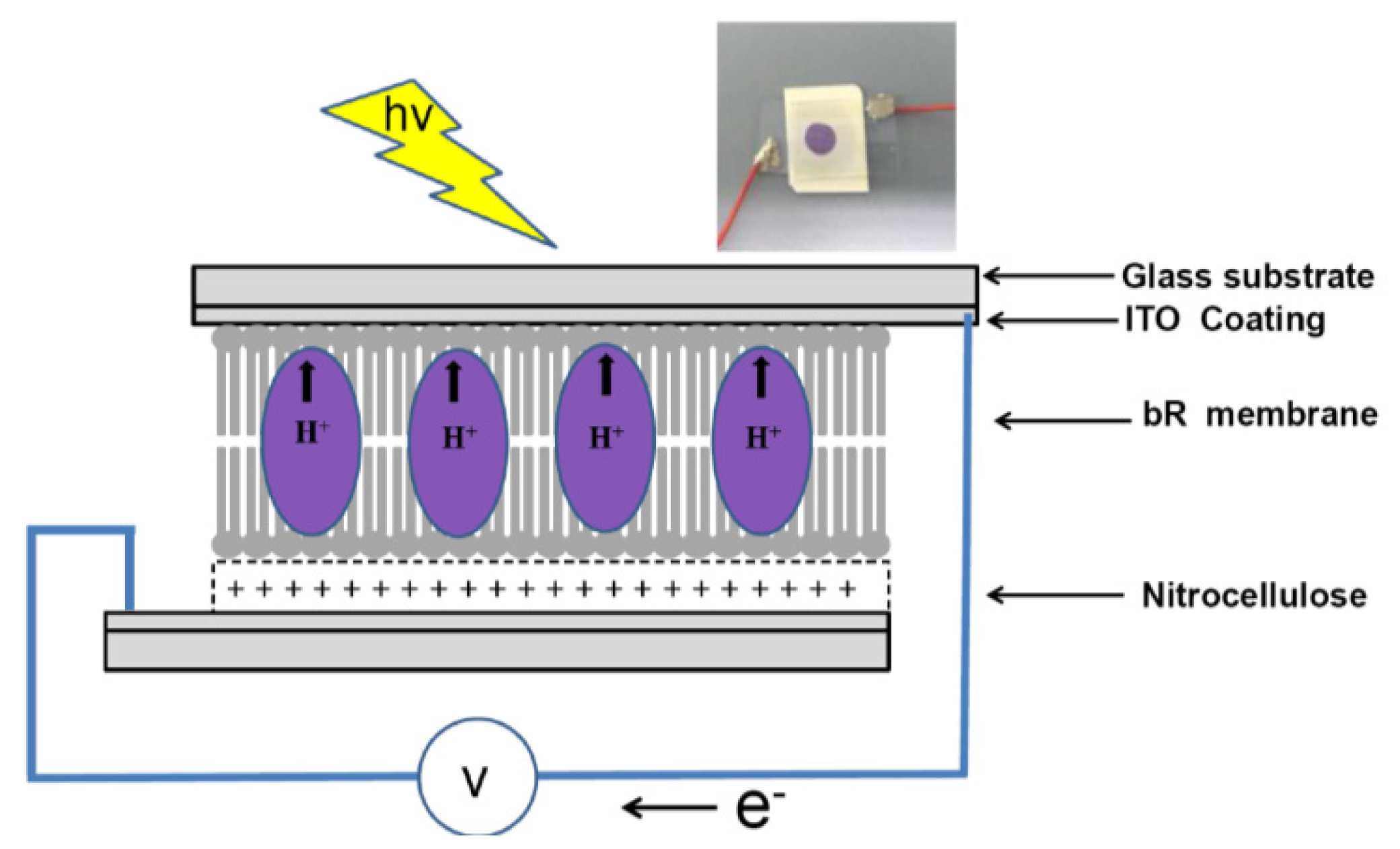
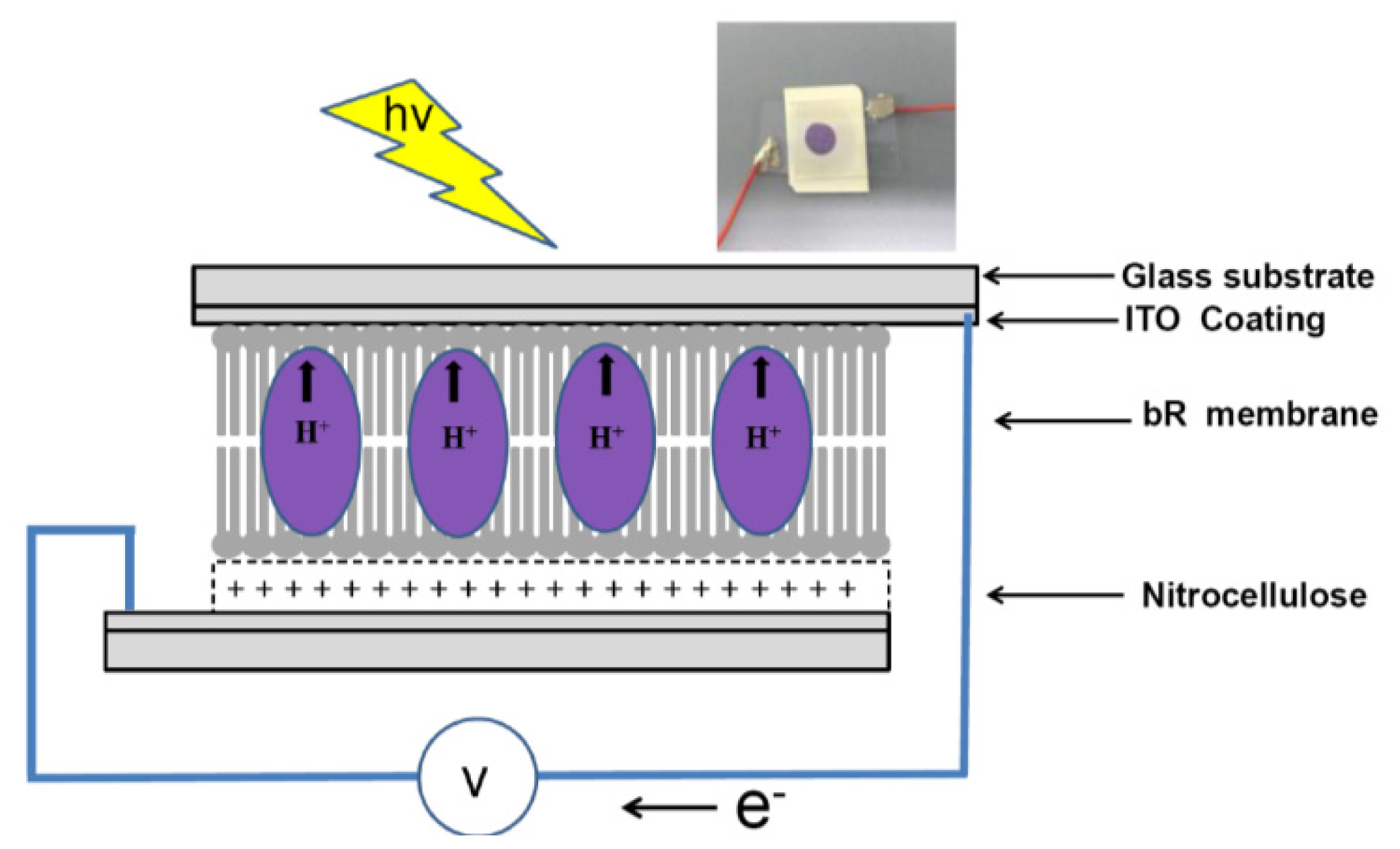
2.4. PM immobilization onto Nitrocellulose Membrane in Photovoltatic Cell
2.5. Photoelectrochemical Measurements
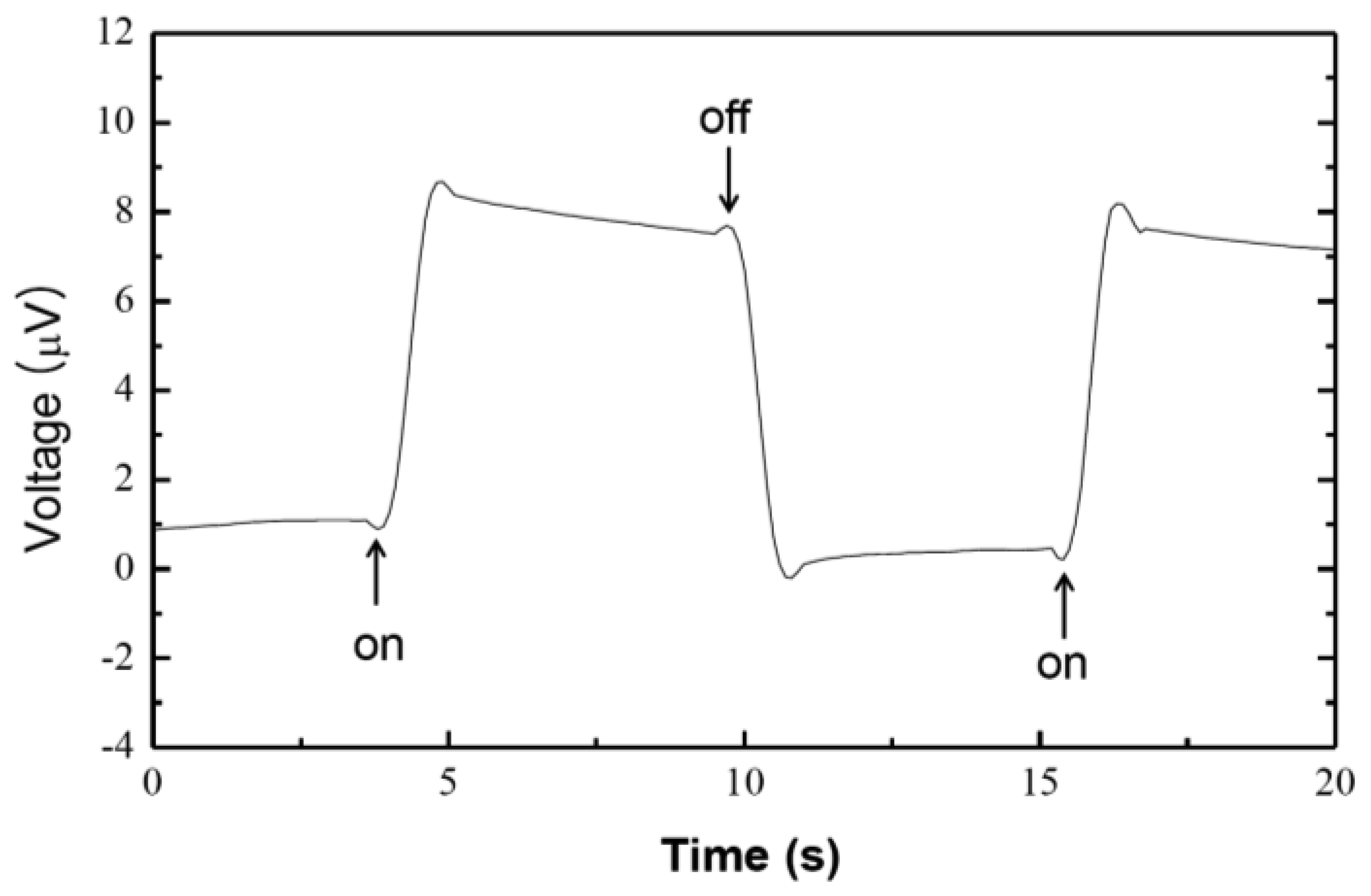
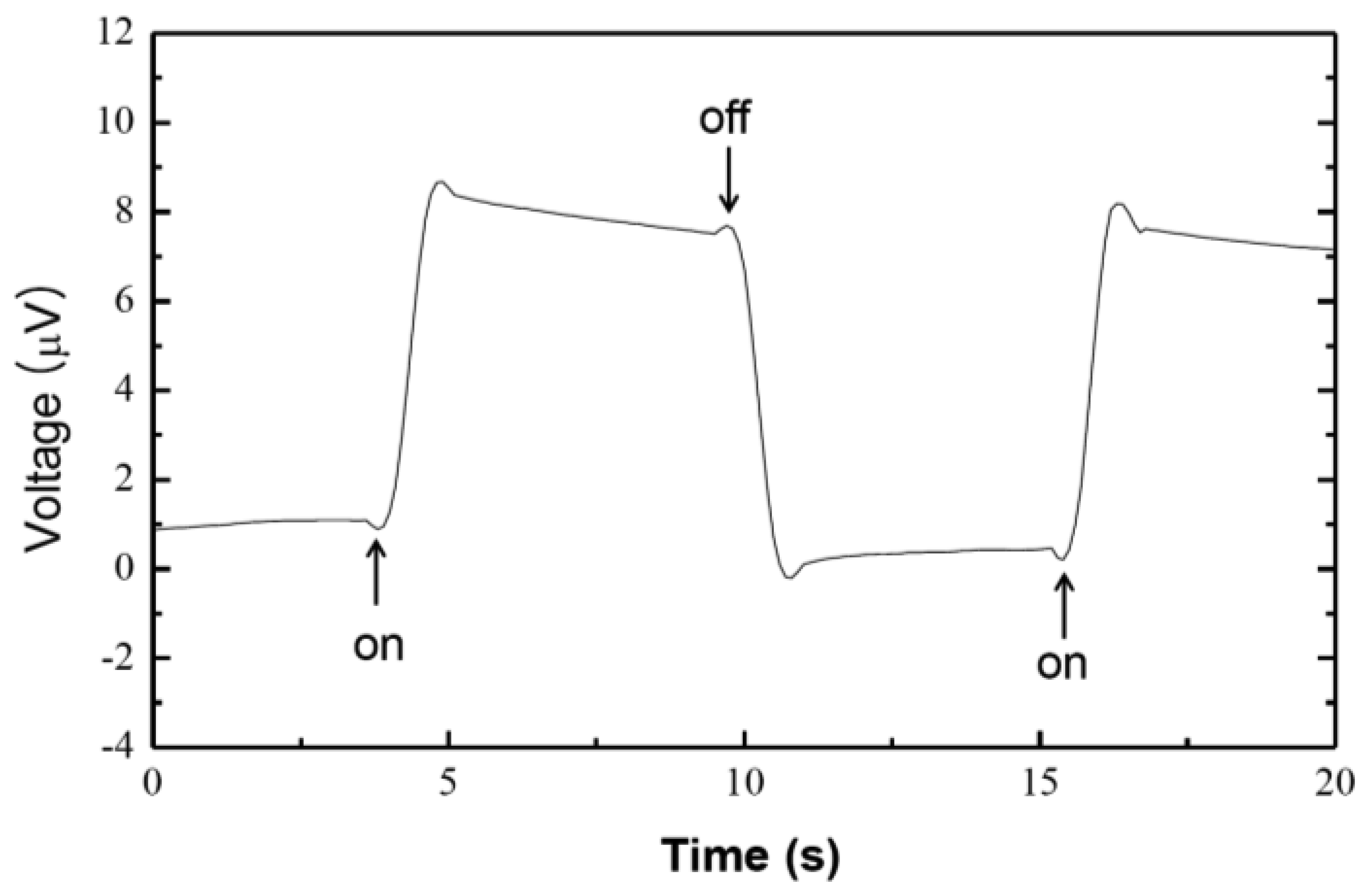
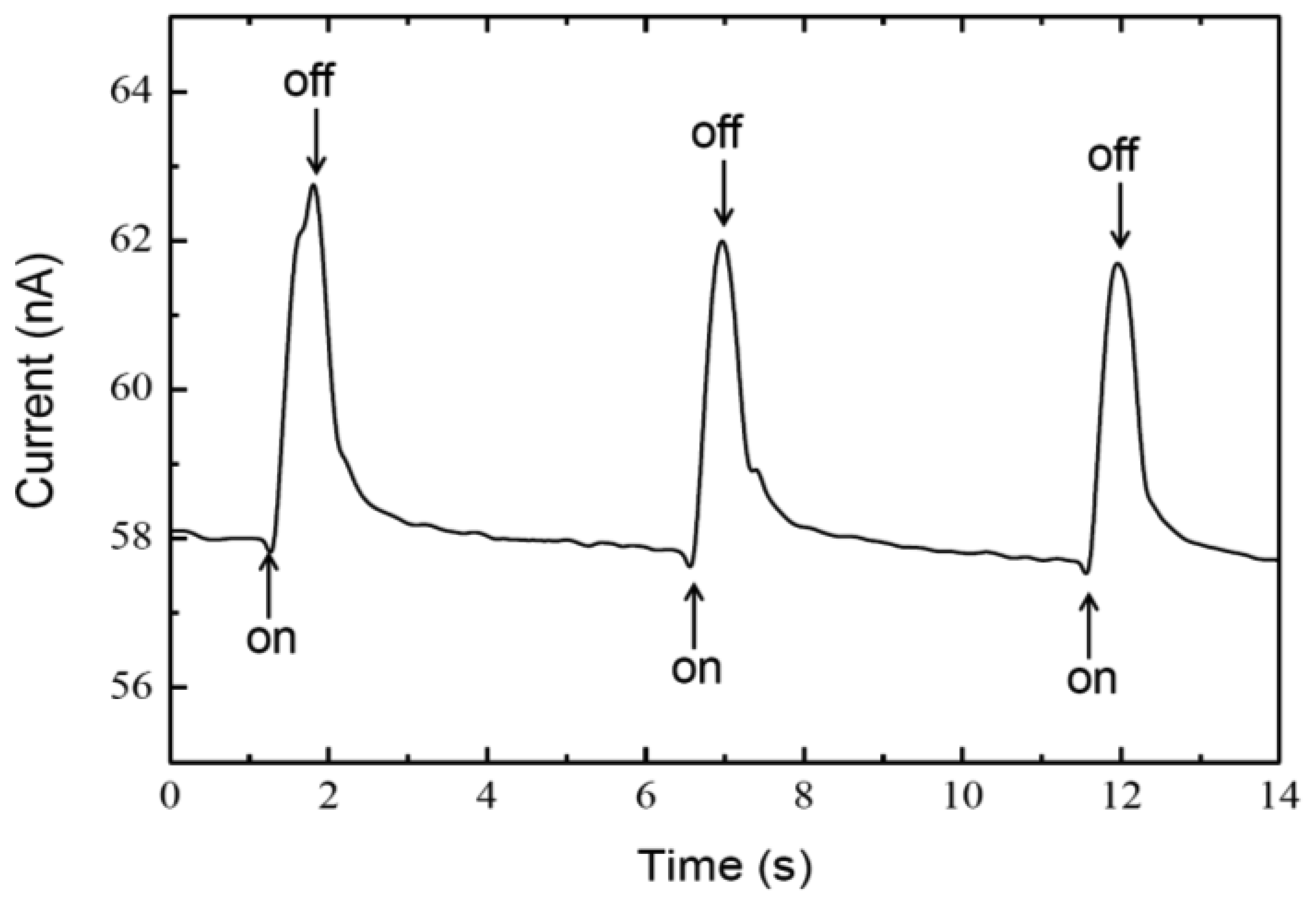
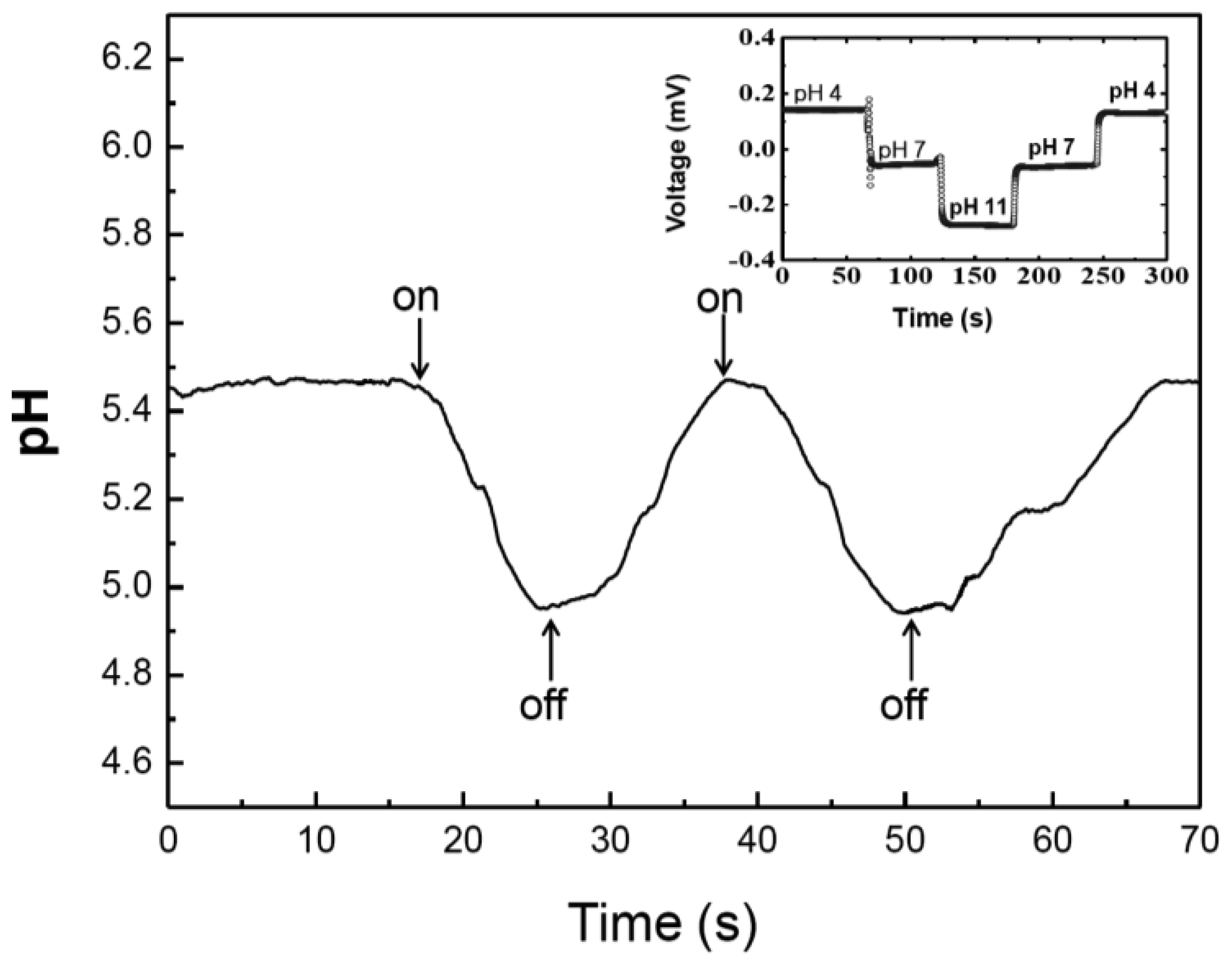
3. Results and Discussions
4. Conclusions
Acknowledgments
References
- Henderson, R.; Unwin, P.N. Three-dimensional model of purple membrane obtained by electron microscopy. Nature 1975, 257, 28–32. [Google Scholar]
- Oesterhelt, D.; Stoeckenius, W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nature 1971, 233, 149–152. [Google Scholar]
- Wang, W.W.; Knopf, G.K.; Bassi, A.S. Photoelectric properties of a detector based on dried bacteriorhodopsin film. Biosens. Bioelectron. 2006, 21, 1309–1319. [Google Scholar]
- Shen, Y.; Safinya, C.R.; Liang, K.S.; Ruppert, A.F.; Rothschild, K.J. Stabilization of the membrane protein bacteriorhodopsin to 140 C in two-dimensional films. Nature 1993, 366, 48–50. [Google Scholar]
- Choi, H.G.; Min, J.; Han, K.K.; Kim, Y.K.; Lee, W.H.; Choi, J.W. Optimal fabrication condition of bacteriorhodopsin thin films onto modified self-assembled monolayers. Mol. Cryst. Liq. Cryst. 2000, 349, 303–306. [Google Scholar]
- Jussila, T.; Li, M.; Tkachenko, N.V.; Parkkinen, S.; Li, B.; Jiang, L.; Lemmetyinen, H. Transient absorption and photovoltage study of self-assembled bacteriorhodopsin/polycation multilayer films. Biosens. Bioelectron. 2002, 17, 509–515. [Google Scholar]
- Dima, M.; Bálint, Z.; Ganea, C.; Váró, G. Wavelength dependence of the electric signals generated in dried BR samples. Rom. J. Biophys. 2007, 17, 219–224. [Google Scholar]
- Robertson, B.; Lukashev, E.P. Rapid pH change due to bacteriorhodopsin measured with a tin-oxide electrode. Biophys. J. 1995, 68, 1507–1517. [Google Scholar]
- Hampp, N. Bacteriorhodopsin as a photochromic retinal protein for optical memories. Chem. Rev. 2000, 100, 1755–1776. [Google Scholar]
- Wang, J. Vectorially oriented purple membrane: characterization by photocurrent measurement and polarized-Fourier transform infrared spectroscopy. Thin Solid Films 2000, 379, 224–229. [Google Scholar]
- Li, R.; Cui, X.; Hu, W.; Lu, Z.; Li, C.M. Fabrication of oriented poly-l-lysine/bacteriorhodopsin-embedded purple membrane multilayer structure for enhanced photoelectric response. J. Colloid. Interface Sci. 2010, 344, 150–157. [Google Scholar]
- Furuno, T.; Takimoto, K.; Kouyama, T.; Ikegami, A.; Sasabe, H. Photovoltaic properties of purple membrane Langmuir-Blodgett films. Thin Solid Films 1988, 160, 145–151. [Google Scholar]
- Keszthelyi, L.; Ormos, P. Displacement current on purple membrane fragments oriented in a suspension. Biophys. Chem. 1983, 18, 397–405. [Google Scholar]
- Brizzolara, R.A. A method for patterning purple membrane using self-assembled monolayers. Biosystems 1995, 35, 137–140. [Google Scholar]
- Chu, J.; Li, X.; Tang, J. Studies on poly (ethylenimine)/purple membrane multilayer films fabricated by layer-by-layer self-assembly. Colloid. Surface Physicochem. Eng. Aspect. 2005, 257, 41–45. [Google Scholar]
- Koyama, K.; Yamaguchi, N.; Miyasaka, T. Antibody-mediated bacteriorhodopsin orientation for molecular device architectures. Science 1994, 265, 762–765. [Google Scholar]
- Weetall, H.H. D96N mutant bacteriorhodopsin immobilized in sol-gel glass characterization. Appl. Biochem. Biotechnol. 1994, 49, 241–256. [Google Scholar]
- Chen, Z.; Chittibabu, K.G.; Marx, K.A.; Kumar, J.; Tripathy, S.K.; Samuelson, L.A.; Akkara, J.A.; Kaplan, D.L. Photodynamic protein incorporated in conducting polymer and sol-gel matrices: toward smart materials for information storage and processing. Proc. SPIE 1994. [Google Scholar] [CrossRef]
- Yamada, T.; Haruyama, Y.; Kasai, K.; Terui, T.; Tanaka, S.; Kaji, T.; Kikuchi, H.; Otomo, A. Orientation of a bacteriorhodopsin thin film deposited by dip coating technique and its chiral SHG as studied by SHG interference technique. Chem. Phys. Lett. 2012, 530, 113–119. [Google Scholar]
- Oesterhelt, D.; Stoeckenius, W. Isolation of the Cell Membrane of Halobacterium Halobium and its Fractionation into Red and Purple Membrane. Method. Enzymol. 1974, 31, 667–678. [Google Scholar]
- Yamanaka, K. Anodically Electrodeposited Iridium Oxide Films(AEIROF) from Alkaline Solutions for Electrochromic Display Devices. Jpn. J. Appl. Phys. 1989, 28, 632–637. [Google Scholar]
- Horn, C.; Steinem, C. Photocurrents generated by bacteriorhodopsin adsorbed on nano-black lipid membranes. Biophys. J. 2005, 89, 1046–1054. [Google Scholar]
- Wang, J.; Song, L.; Yoo, S.; El-Sayed, M.A. A comparison of the photoelectric current responses resulting from the proton pumping process of bacteriorhodopsin under pulsed and CW laser excitations. J. Phys. Chem. B 1997, 101, 10599–10604. [Google Scholar]
- Patil, A.V.; Premaruban, T.; Berthoumieu, O.; Watts, A.; Davis, J.J. Enhanced photocurrent in engineered bacteriorhodopsin monolayer. J. Phys. Chem. B 2011, 116, 683–689. [Google Scholar]
- Martin, C.H.; Chen, Z.P.; Birge, R.R. Towards a bacteriorhodopsin-silicon neuromorphic photosensor. Pac. Symp. Biocomput. 1997, 268–279. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, Y.J.; Neuzil, P.; Nam, C.-H.; Engelhard, M. Deposition of Bacteriorhodopsin Protein in a Purple Membrane Form on Nitrocellulose Membranes for Enhanced Photoelectric Response. Sensors 2013, 13, 455-462. https://doi.org/10.3390/s130100455
Kim YJ, Neuzil P, Nam C-H, Engelhard M. Deposition of Bacteriorhodopsin Protein in a Purple Membrane Form on Nitrocellulose Membranes for Enhanced Photoelectric Response. Sensors. 2013; 13(1):455-462. https://doi.org/10.3390/s130100455
Chicago/Turabian StyleKim, Young Jun, Pavel Neuzil, Chang-Hoon Nam, and Martin Engelhard. 2013. "Deposition of Bacteriorhodopsin Protein in a Purple Membrane Form on Nitrocellulose Membranes for Enhanced Photoelectric Response" Sensors 13, no. 1: 455-462. https://doi.org/10.3390/s130100455
APA StyleKim, Y. J., Neuzil, P., Nam, C.-H., & Engelhard, M. (2013). Deposition of Bacteriorhodopsin Protein in a Purple Membrane Form on Nitrocellulose Membranes for Enhanced Photoelectric Response. Sensors, 13(1), 455-462. https://doi.org/10.3390/s130100455