Allelic Variants of Glutamine Synthetase and Glutamate Synthase Genes in a Collection of Durum Wheat and Association with Grain Protein Content


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. PCR Condition and Sequencing
2.3. Digestion with CEL I and Revelation Fragments
2.4. Protein Content Quantification and Regression Analysis
3. Results and Discussion
3.1. Phenotypic Characterization for the Protein Content of a Collection of Tetraploid Wheat Genotypes
3.2. GS2 Allelic Variants and Relationship with GPC
3.3. GOGAT Allelic Variants and Relationship with GPC
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lawlor, D.W. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef]
- Triboi, E.; Triboi-Blondel, A.M. Productivity and grain or seed composition: A new approach to an old problem. Eur. J. Agron. 2002, 16, 163–186. [Google Scholar] [CrossRef]
- Lea, P.J.; Azevedo, R.A. Nitrogen use efficiency. 2. Amino acid metabolism. Ann. Appl. Biol. 2007, 151, 269–275. [Google Scholar] [CrossRef]
- Hirel, B.; le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Obara, M.; Sato, T.; Sasaki, S.; Kashiba, K.; Nagano, A.; Nakamura, I.; Ebitani, T.; Yano, M.; Yamaya, T. Identification and characterization of a QTL on chromosome 2 for cytosolic glutamines ynthetase content and panicle number in rice. Theor. Appl. Genet. 2004, 110, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Swarbreck, S.M.; Defoin-Platel, M.; Hindle, M.; Saqi, M.; Habash, D.Z. New perspectives on glutamine synthetase in grasses. J. Exp. Bot. 2011, 62, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.M.; Møller, A.L.B.; Dionisio, G.; Kichey, T.; Jahn, T.P.; Dubois, F.; Baudo, M.; Lopes, M.S.; Tercé-Laforgue, T.; Foyer, C.H.; et al. Gene expression, cellular localization and function of glutamine synthetase isozymes in wheat (Triticum aestivum L.). Plant. Mol. Biol. 2008, 67, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.C.; Eriksson, D.; Moller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant. Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Habash, D.Z.; Bernard, S.; Schondelmaier, J.; Weyen, J.; Quarrie, S.A. The genetics of nitrogenuse in hexaploid wheat:N utilisation, development and yield. Theor. Appl. Genet. 2007, 114, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, A.; Nigro, D.; Giancaspro, A.; Blanco, A. The glutamine synthetase (GS2) genes in relation to grain protein content of durum wheat. Funct. Integr. Genom. 2011, 11, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, A.; Nigro, D.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Blanco, A. Isolation and characterization of cytosolic glutamine synthetase (GSe) genes and association with grain protein content in durum wheat. Crop Pasture Sci. 2014, 65, 38–45. [Google Scholar] [CrossRef]
- Nigro, D.; Fortunato, S.; Giove, S.L.; Paradiso, A.; Gu, Y.Q.; Blanco, A.; de Pinto, M.C.; Gadaleta, A. Glutamine synthetase in Durum Wheat: Genotypic Variation and Relationship with Grain Protein Content. Front. Plant Sci. 2016, 7, 971. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef] [PubMed]
- Cren, M.; Hirel, B. Glutamine Synthetase in Higher Plants Regulation of Gene and Protein Expression from the Organ to the Cell. Plant Cell Physiol. 1999, 40, 1187–1193. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice. J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, F.M.; Avila, C.; Cantón, F.R.; Cañas, R.A.; de la Torre, F. Ammonium assimilation and amino acid metabolism in conifers. J. Exp. Bot. 2007, 58, 2307–2318. [Google Scholar] [CrossRef]
- Leegood, R.C.; Lea, P.J.; Adcock, M.D.; Hausler, R.E. The regulation and control of photorespiration. J. Exp. Bot. 1995, 46, 1397–1414. [Google Scholar] [CrossRef]
- Ferrario-Méry, S.; Hodges, M.; Hirel, B.; Foyer, C.H. Photorespiration dependent increases in phosphoenolpyruvate carboxylase, isocitrate dehydrogenase and glutamate dehydrogenase in transformed tobacco plants deficient in ferredoxin-dependent glutamine-alpha-ketoglutarate aminotransferase. Planta 2002, 214, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Ferrario-Méry, S.; Valadier, M.H.; Godefroy, N.; Miallier, D.; Hirel, B.; Foyer, C.H.; Suzuki, A. Diurnal changes in ammonia assimilation in transformed tobacco plants expressing ferredoxin-dependent glutamate synthase mRNA in the antisense orientation. Plant Sci. 2002, 163, 59–67. [Google Scholar] [CrossRef]
- Lancien, M.; Martin, M.; Hsieh, M.H.; Leustek, T.; Goodman, H.; Coruzzi, G.M. Arabidopsis glt1-T mutant defines a role for NADH-GOGAT in the non-photorespiratory ammonium assimilatory pathway. Plant J. 2002, 29, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Kawabata, S.; Takahashi, H.; Hase, T.; Sugiyama, T. Molecular cloning of the family of glutamine synthetase genes from maize: Expression of genes for glutamine synthetase and ferredoxin-dependent glutamate synthase in photosynthetic and non-photosynthetic tissues. Plant Cell Physiol. 1992, 33, 49–58. [Google Scholar]
- Zehnacker, C.; Becker, T.W.; Suzuki, A.; Carrayol, E.; Caboche, M.; Hirel, B. Purification and properties of tobacco ferredoxin-dependent glutamate synthase, and isolation of corresponding cDNA clones. Light-inducibility and organspecificity of gene transcription and protein expression. Planta 1992, 187, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Coschigano, K.T.; Melo-Oliveira, R.; Lim, J.; Coruzzi, G.M. Arabidopsis gls mutants and distinct Fd.-GOGAT genes: Implications for photorespiration and primary nitrogen assimilation. Plant Cell 1998, 10, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.; Márquez, A.J.; Pajuelo, P.; Cannell, M.E.; Wallsgrove, R.M.; Forde, B.G. Cloning and sequence analysis of a cDNA for barley ferredoxin-dependent glutamate synthase and molecular analysis of photorespiratory mutants deficient in the enzyme. Planta 1993, 189, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Boisson, M.; Mondon, K.; Torney, V.; Nicot, N.; Laine, A.L.; Bahrman, N.; Gouy, A.; Daniel-Vedele, F.; Hirel, B.; Sourdille, P.; et al. Partial sequences of nitrogen metabolism genes in hexaploid wheat. Theor. Appl. Genet. 2005, 110, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Nigro, D.; Gu, Y.Q.; Huo, N.; Marcotuli, I.; Blanco, A.; Gadaleta, A.; Anderson, O.D. Structural analysis of the wheat genes encoding NADH-dependent glutamine-2-oxoglutarate amidotransferases genes and correlation with grain protein content. PLoS ONE 2013, 8, e73751. [Google Scholar] [CrossRef] [PubMed]
- Nigro, D.; Blanco, A.; Anderson, O.D.; Gadaleta, A. Characterization of Ferredoxin-Dependent Glutamine-Oxoglutarate Amidotransferase (Fd-GOGAT) Genes and Their Relationship with Grain Protein Content QTL in Wheat. PLoS ONE 2014, 9, e103869. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.J.; Kreis, M.; Shewry, P.R.; Gale, M.D. Location of b-amylase sequences in wheat and its relatives. Theoret. Appl. Genet. 1988, 75, 286–290. [Google Scholar] [CrossRef]
- Freed, R.; Eisensmith, S.P.; Goetz, D.; Reicosky, D.; Smail, V.W.; Wolberg, P. MSTAT-C: A Microcomputer Program for the Design, Management, and Analysis of Agronomic Research Experiments; Michigan State University: East Lansing, MI, USA, 1991. [Google Scholar]
- Laido, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; de Vita, P. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef] [PubMed]
- Marcotuli, I.; Houston, K.; Schwerdt, J.G.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Gadaleta, A. Genetic diversity and genome wide association study of β-glucan content in tetraploid wheat grains. PLoS ONE 2016, 11, e0152590. [Google Scholar] [CrossRef] [PubMed]
- Colasuonno, P.; Lozito, M.L.; Marcotuli, I.; Nigro, D.; Giancaspro, A.; Mangini, G.; Simeone, R. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genom. 2017, 18, 122. [Google Scholar] [CrossRef] [PubMed]
- Nigro, D.; Laddomada, B.; Mita, G.; Blanco, E.; Colasuonno, P.; Simeone, R.; Blanco, A. Genome-wide association mapping of phenolic acids in tetraploid wheats. J. Cereal Sci. 2017, 75, 25–34. [Google Scholar] [CrossRef]
- Li, X.P.; Zhao, X.Q.; He, X.; Zhao, G.Y.; Li, B.; Liu, D.C.; Li, Z.S. Haplotype analysis of the genes encoding glutamine synthetase plastic isoforms and their association with nitrogen-use-and yield-related traits in bread wheat. New Phytol. 2011, 189, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Guttieri, M.J.; Frels, K.; Regassa, T.; Waters, B.M.; Baenziger, P.S. Variation for nitrogen use efficiency traits in current and historical great plains hard winter wheat. Euphytica 2017, 213, 87. [Google Scholar] [CrossRef]
- Fontaine, J.X.; Ravel, C.; Pageau, K.; Heumez, E.; Dubois, F.; Hirel, B.; Le Gouis, J. A quantitative genetic study for elucidating the contribution of glutamine synthetase, glutamate dehydrogenase and other nitrogen-related physiological traits to the agronomic performance of common wheat. Theor. Appl. Genet. 2009, 119, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Bogard, M.; Allard, V.; Martre, P.; Heumez, E.; Snape, J.W.; Orford, S.; Griffiths, S.; Gaju, O.; Foulkes, J.; Le Gouis, J. Identifying wheat genomic regions for improving grain protein concentration independently of grain yield using multiple inter-related populations. Mol. Breed. 2013, 31, 587. [Google Scholar] [CrossRef]
- Zeng, D.D.; Qin, R.; Li, M.; Alamin, M.; Jin, X.L.; Liu, Y.; Shi, C.H. The ferredoxin-dependent glutamate synthase (OsFd-GOGAT) participates in leaf senescence and the nitrogen remobilization in rice. Mol. Genet. Genom. 2017, 292, 385. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, U.M.; Abrouk, M.; Murat, F.; Pont, C.; Foucrier, S.; Desmaizieres, G.; Confolent, C.; Riviere, N.; Charmet, G.; Paux, E.; et al. Cross-genome map based dissection of a nitrogen use efficiency ortho-meta QTL in bread wheat unravels concerted cereal genome evolution. Plant J. 2011, 65, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.; Mangini, G.; Giancaspro, A.; Giove, S.; Colasuonno, P.; Simeone, R.; Signorile, A.; de Vita, P.; Mastrangelo, A.M.; Cattivelli, L.; et al. Relationships between grain protein content and grain yield components through QTL analyses in a RIL population derived from two elite durum wheat cultivars. Mol. Breed. 2012, 30, 79–92. [Google Scholar] [CrossRef]
- Lu, Y.; Luo, F.; Yang, M.; Li, X.; Lian, X. Suppression of glutamate synthase genes significantly affects carbon and nitrogen metabolism in rice (Oryza sativa L.). China Life Sci. 2011, 54, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Joppa, L.R.; Cantrell, R.G. Chromosomal location of genes for grain protein content of wild tetraploid wheat. Crop Sci. 1990, 30, 1059–1064. [Google Scholar] [CrossRef]
- Prasad, M.; Varshney, R.K.; Kumar, A.; Balyan, H.S.; Sharma, P.C.; Edwards, K.J.; Gupta, P.K. A microsatellite marker associated with a QTL for grain protein content on chromosome arm 2DL of bread wheat. Theor. Appl. Genet. 1999, 99, 341–345. [Google Scholar] [CrossRef]
- Suprayogi, Y.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Singh, A.K. Identification and validation of quantitative trait loci for grain protein concentration in adapted Canadian durum wheat populations. Theor. Appl. Genet. 2009, 119, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, P.B.; Lopez-Bellido, R.; Hawkesford, M.J. Genotypic variation in the uptake, partitioning and remobilization of nitrogen during grain-filling in wheat. Field Crops Res. 2014, 156, 242–248. [Google Scholar] [CrossRef] [PubMed]
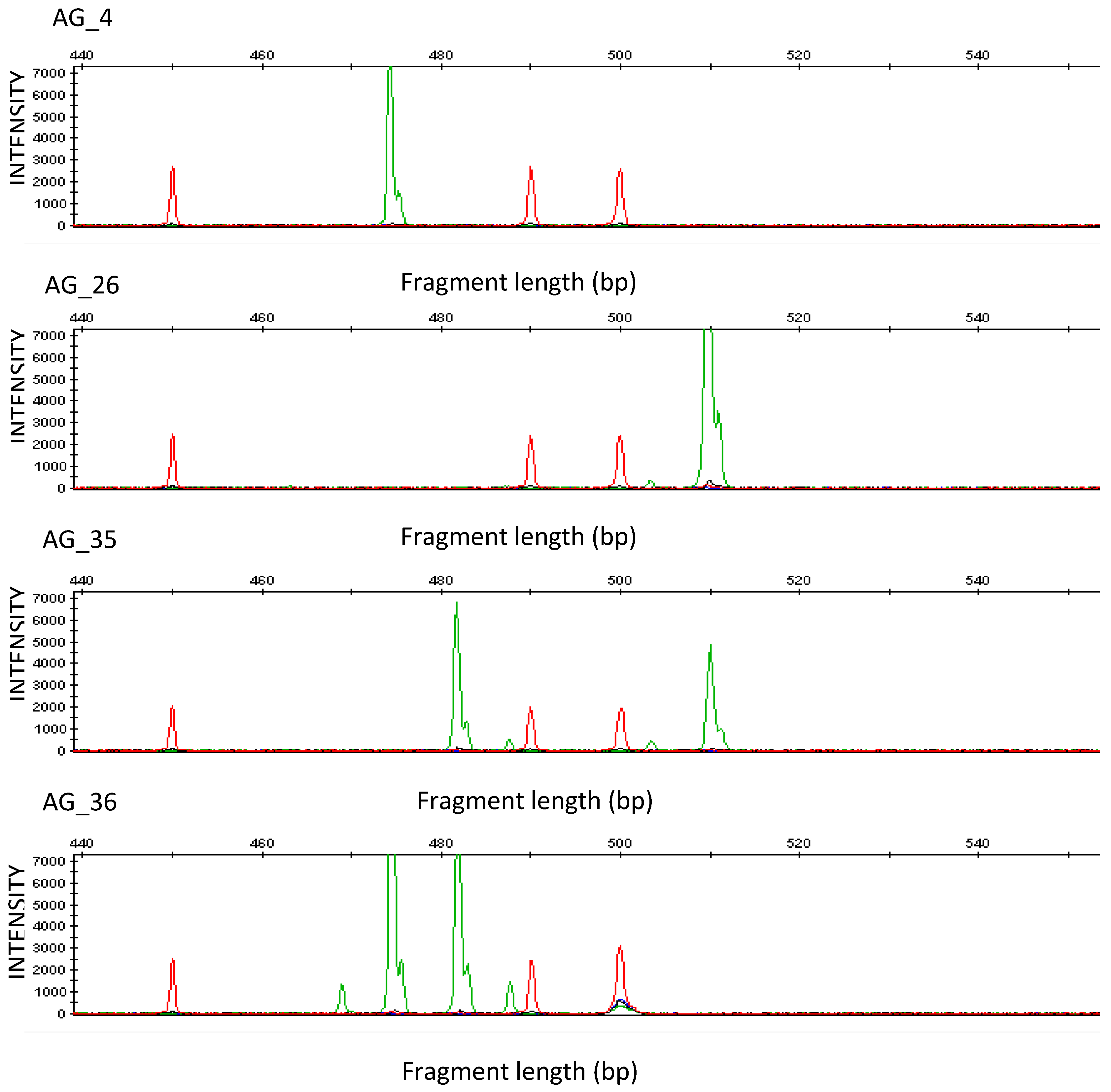

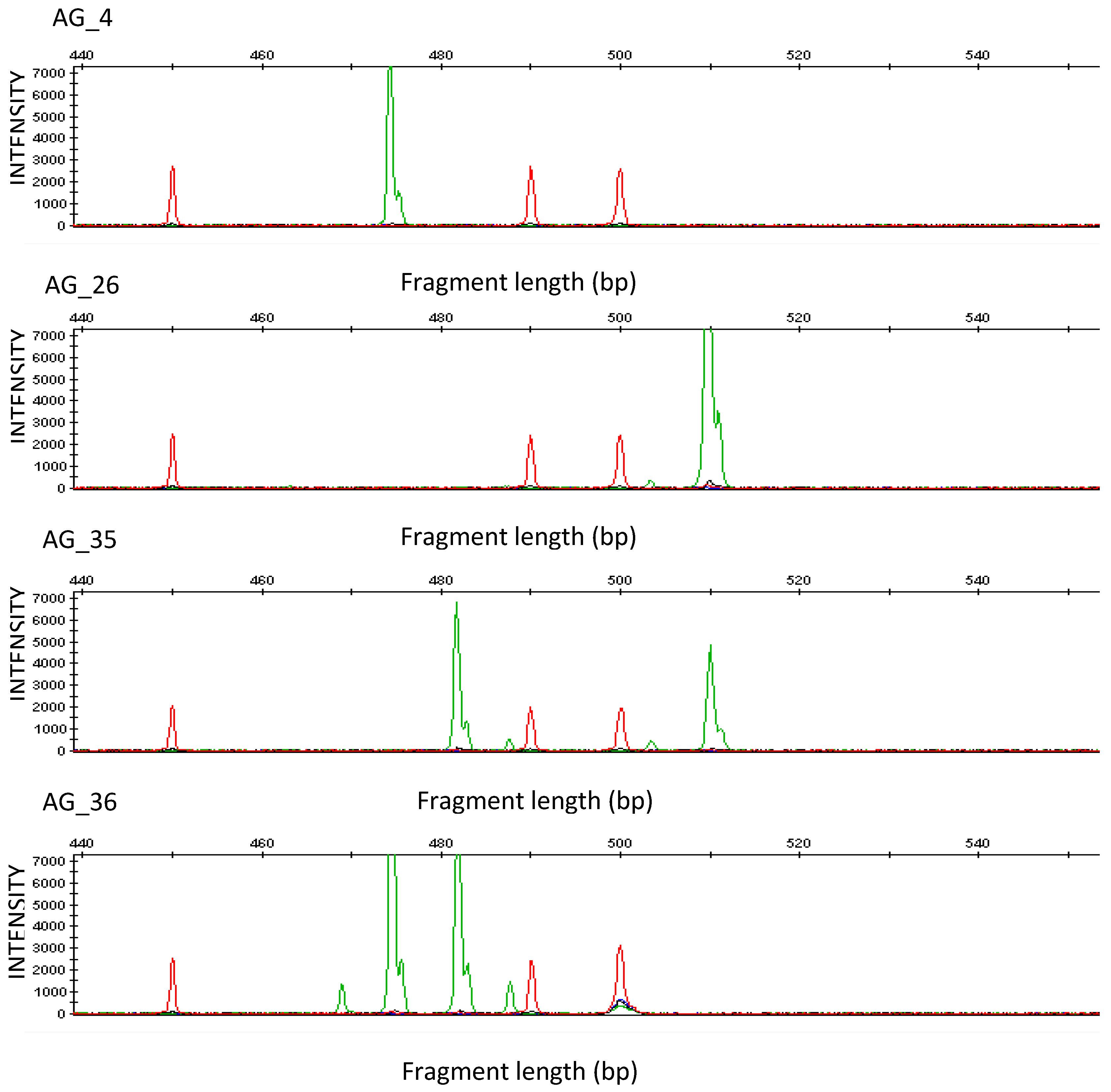{kind=link}
| Environment | Foggia 2009 | Valenzano 2009 | Valenzano 2010 |
|---|---|---|---|
| Mean | 14.1 | 16.2 | 15.5 |
| SD | 1.55 | 2.01 | 2.21 |
| Min | 10.8 | 11.8 | 12.5 |
| Max | 20.4 | 22.9 | 23.5 |
| CV (%) | 10.9 | 12.4 | 14.3 |
| Gene | Amplicons (bp) | Frequency | Environments | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Foggia 2009 | Valenzano 2009 | Valenzano 2010 | |||||||||
| log10(p) | Effect | R2 | log10(p) | Effect | R2 | −log10(p) | Effect | R2 | |||
| GS2-A2 | 480/719 | 131–90 | 5.3 *** | 0.92 | 9.2 | 2.6 * | 0.83 | 4.1 | 3.7 ** | 1.11 | 6.1 |
| GS2-B2 | 473/507 | 184–37 | 0.4 | 0.24 | 0.4 | 0.4 | −0.33 | 0.4 | 0.1 | 0.13 | 0.0 |
| Gene | Allele | Frequency | Environments | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Foggia 2009 | Valenzano 2009 | Valenzano 2010 | |||||||||
| −log10(p) | Effect | R2 | −log10(p) | Effect | R2 | −log10(p) | Effect | R2 | |||
| Fd-GOGAT-A2a | C/T | 204–17 | 4.1 *** | −1.47 | 6.9 | 6.7 *** | −2.48 | 11.7 | 8.4 *** | −3.05 | 14.6 |
| Fd-GOGAT-A2b | C/T | 115–99 | 0.2 | −0.09 | 0.1 | 0.0 | −0.02 | 0.0 | 0.2 | −0.14 | 0.1 |
| Fd-GOGAT-A2c | A/G | 100–117 | 0.3 | 0.12 | 0.2 | 0.0 | 0.02 | 0.0 | 0.2 | 0.13 | 0.1 |
| Fd-GOGAT-B2a | A/G | 181–38 | 0.9 | −0.41 | 1.1 | 2.3 | −0.98 | 3.5 | 2.7 * | −1.19 | 4.3 |
| Fd-GOGAT-B2b | A/G | 37–181 | 1.1 | 0.47 | 1.4 | 2.6 * | 1.06 | 4.0 | 3.0 * | 1.28 | 4.9 |
| Fd-GOGAT-B2c | A/G | 117–101 | 0.3 | −0.15 | 0.3 | 0.0 | 0.00 | 0.0 | 0.2 | −0.14 | 0.1 |
| Gene | Allele | Frequency | Environments | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Foggia 2009 | Valenzano 2009 | Valenzano 2010 | |||||||||
| –log10(p) | Effect | R2 | –log10(p) | Effect | R2 | –log10(p) | Effect | R2 | |||
| NADH-GOGAT-A3a | C/T | 145–70 | 2.7 * | 0.68 | 4.4 | 1.3 | 0.58 | 1.9 | 1.6 | 0.73 | 2.4 |
| NADH-GOGAT-A3b | G/T | 42–176 | 6.1 *** | −1.24 | 10.6 | 4.1 *** | −1.36 | 7.0 | 6.4 *** | −1.87 | 11.1 |
| NADH-GOGAT-A3c | A/G | 111–103 | 0.1 | −0.04 | 0.0 | 0.7 | −0.36 | 0.8 | 1.3 | −0.60 | 1.9 |
| NADH-GOGAT-B3a | G/T | 71–147 | 2.8 * | −0.68 | 4.6 | 1.5 | −0.61 | 2.1 | 1.8 | −0.76 | 2.6 |
| NADH-GOGAT-B3b | C/T | 115–103 | 0.1 | −0.06 | 0.0 | 0.9 | −0.43 | 1.1 | 1.6 | −0.68 | 2.3 |
| NADH-GOGAT-B3c | A/G | 146–72 | 2.6 * | 0.65 | 4.2 | 1.2 | 0.54 | 1.6 | 1.5 | 0.68 | 2.1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nigro, D.; Fortunato, S.; Giove, S.L.; Mangini, G.; Yacoubi, I.; Simeone, R.; Blanco, A.; Gadaleta, A. Allelic Variants of Glutamine Synthetase and Glutamate Synthase Genes in a Collection of Durum Wheat and Association with Grain Protein Content. Diversity 2017, 9, 52. https://doi.org/10.3390/d9040052
Nigro D, Fortunato S, Giove SL, Mangini G, Yacoubi I, Simeone R, Blanco A, Gadaleta A. Allelic Variants of Glutamine Synthetase and Glutamate Synthase Genes in a Collection of Durum Wheat and Association with Grain Protein Content. Diversity. 2017; 9(4):52. https://doi.org/10.3390/d9040052
Chicago/Turabian StyleNigro, Domenica, Stefania Fortunato, Stefania Lucia Giove, Giacomo Mangini, Ines Yacoubi, Rosanna Simeone, Antonio Blanco, and Agata Gadaleta. 2017. "Allelic Variants of Glutamine Synthetase and Glutamate Synthase Genes in a Collection of Durum Wheat and Association with Grain Protein Content" Diversity 9, no. 4: 52. https://doi.org/10.3390/d9040052