Specific siRNA Targeting the Receptor for Advanced Glycation End Products Inhibits Experimental Hepatic Fibrosis in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1 Preparation of specific siRNA targeting RAGE
- – Sequence of R1 (targeting RAGE mRNA 417–437): sense chain is:
- 5’-GATCCCAAGCCGGAAATTGTGAATCCTTTCAAGAGAAGGATTCACAATTTCCGGCTTTTTTTGGAT-3’ and antisense chain is
- 5’-AGCTATCCAAAAAAAGCCGGAAATTGTGAATCCTTCTCTTGAAAGGATTCACAATTTCCGGCTTGG-3’
- – Sequence of R2 (targeting RAGE mRNA 221–241): sense chain is
- 5’-GATCCCAAGGACTGAAGCTTGGAAGGTTTCAAGAGAACCTTCCAAGCTTCAGTCCTTTTTTTGGAT-3’ and antisense chain is
- 5’-AGCTATCCAAAAAAAGGACTGAAGCTTGGAAGGTTCTCTTGAAACCTTCCAAGCTTCAGTCCTTGG-3’
- – Sequence of R3 (targeting RAGE mRNA 534–554): sense chain is
- 5’-GATCCCCACCTCTGATTCCTGATGGCTTCAAGAGAGCCATCAGGAATCAGAGGTTTTTTGGAAA-3’ and antisense chain is
- 5’-AGCTTTTCCAAAAAACCTCTGATTCCTGATGGCTCTCTTGAAGCCATCAGGAATCAGAGGTGGG-3’
- – Sequence of C: sense chain is
- 5’-GATCCCCAATTCCAGTGGCCATCGTATTCAAGAGATACGATGGCCACTGGAATTTTTTTA-3’ and antisense chain is
- 5’-AGCTTAAAAAAATTCCAGTGGCCATCGTATCTCTTGAATACGATGGCCACTGGAATTGGG-3’
2.2 Construction of specific siRNA expression vectors
2.3 Cell culture and transfection
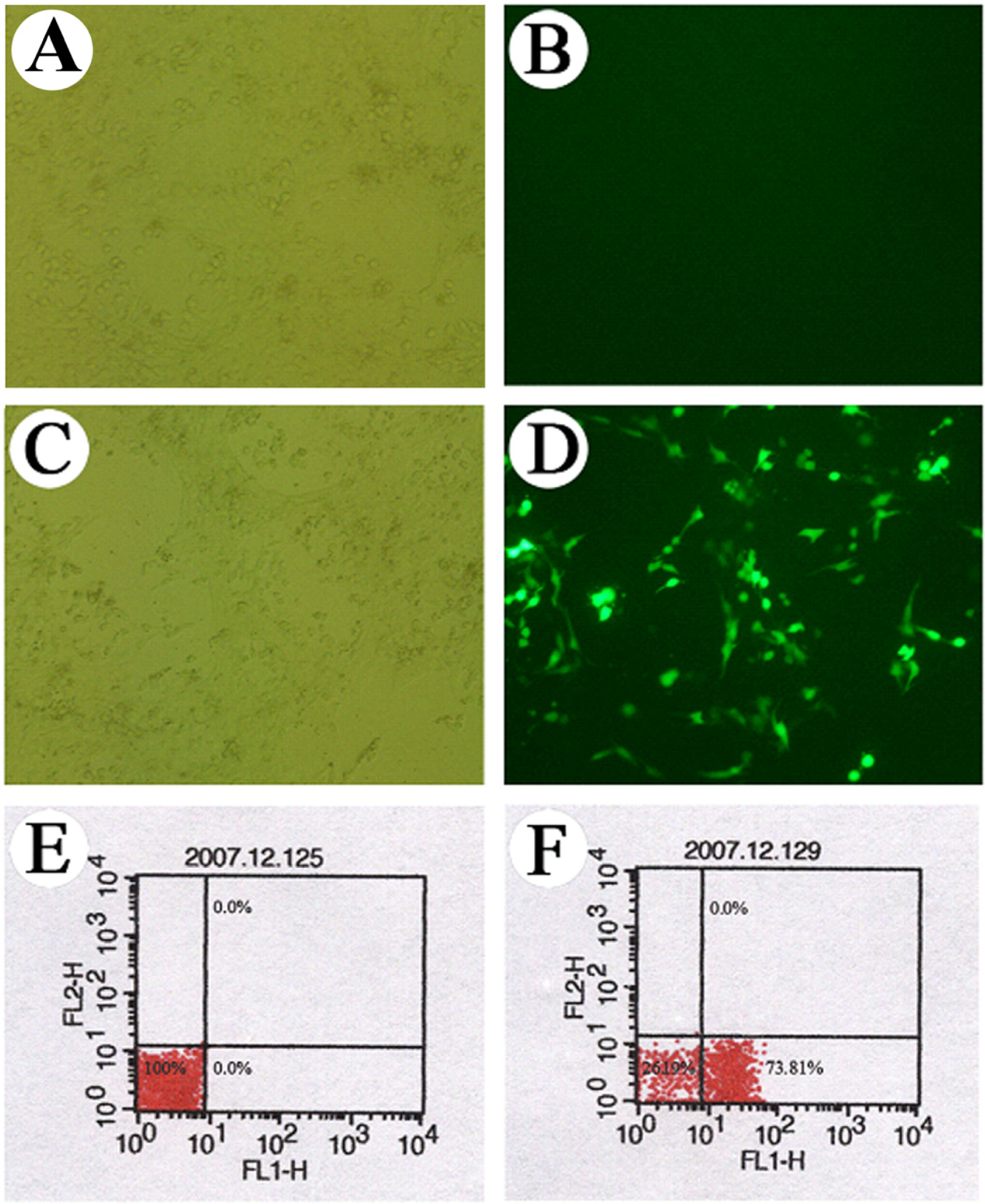
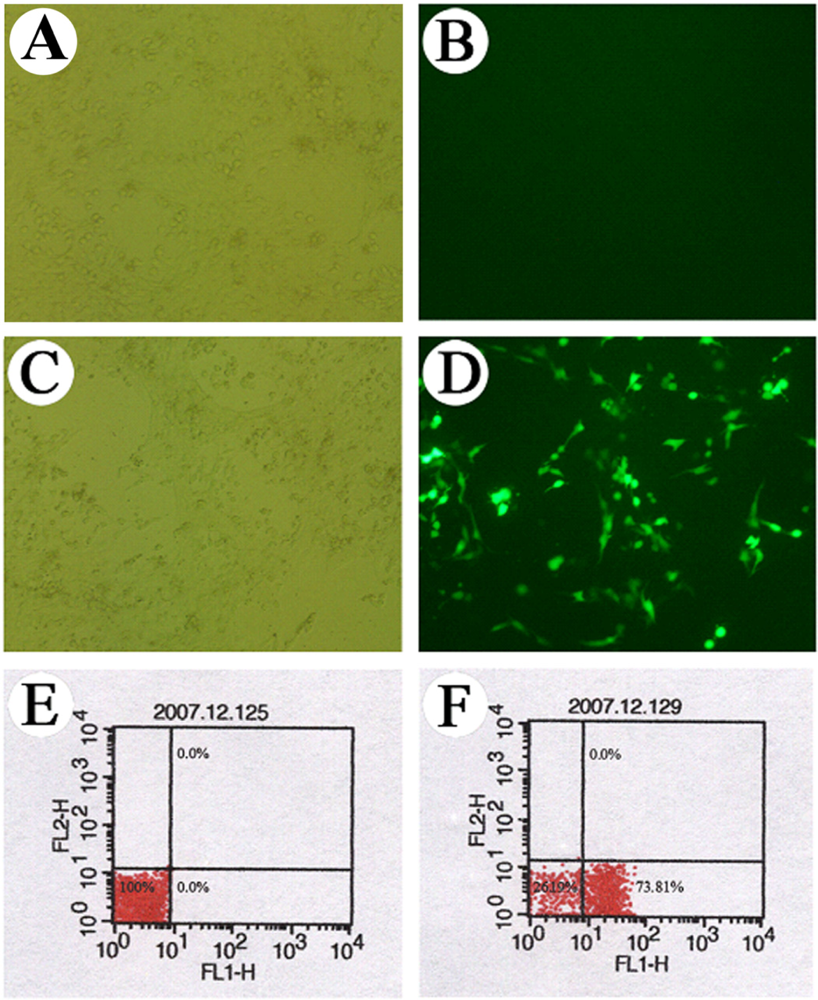
2.4 Observation of cell transfection under a fluorescence microscope
2.5 Animal model and protocol
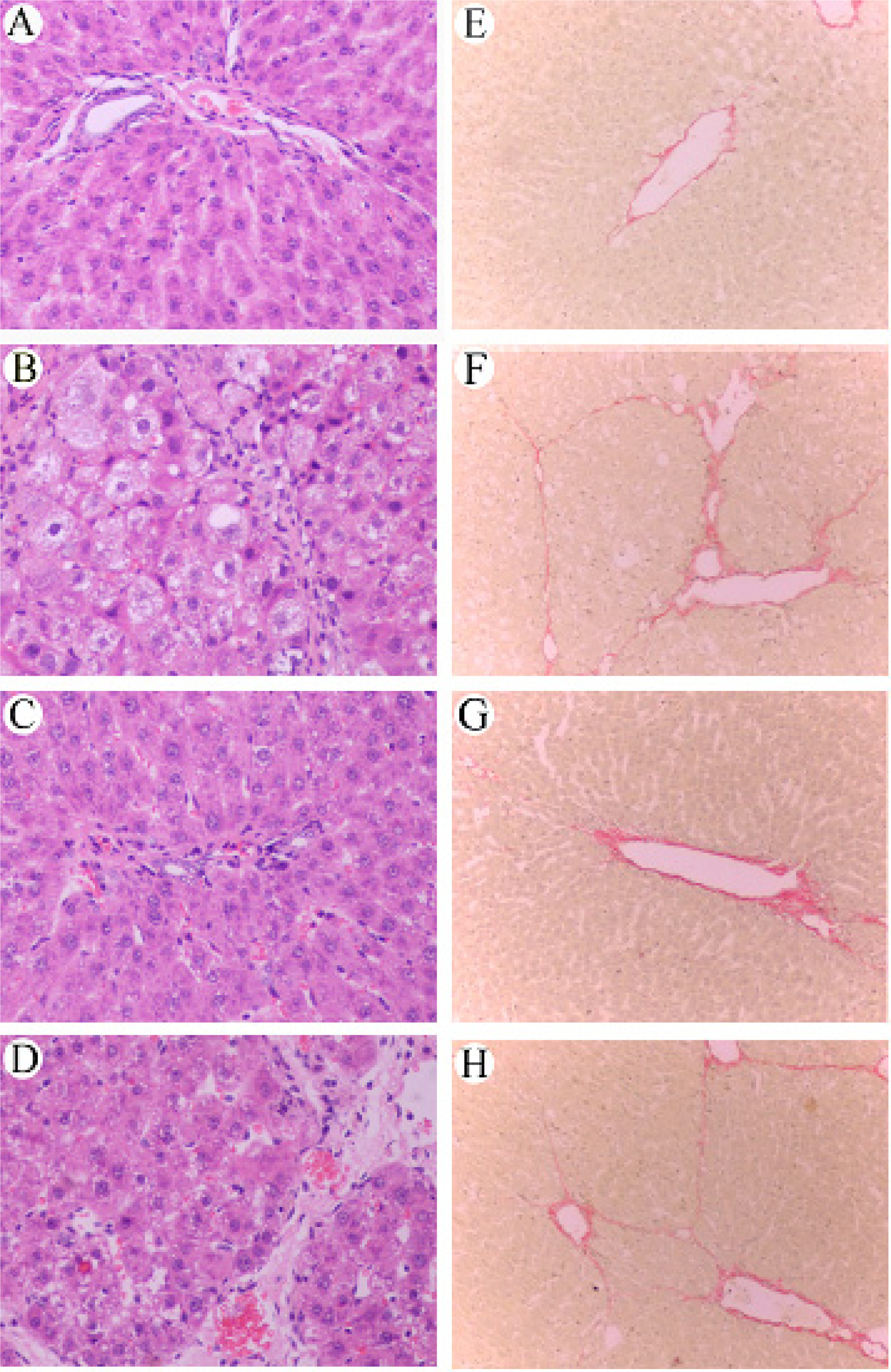
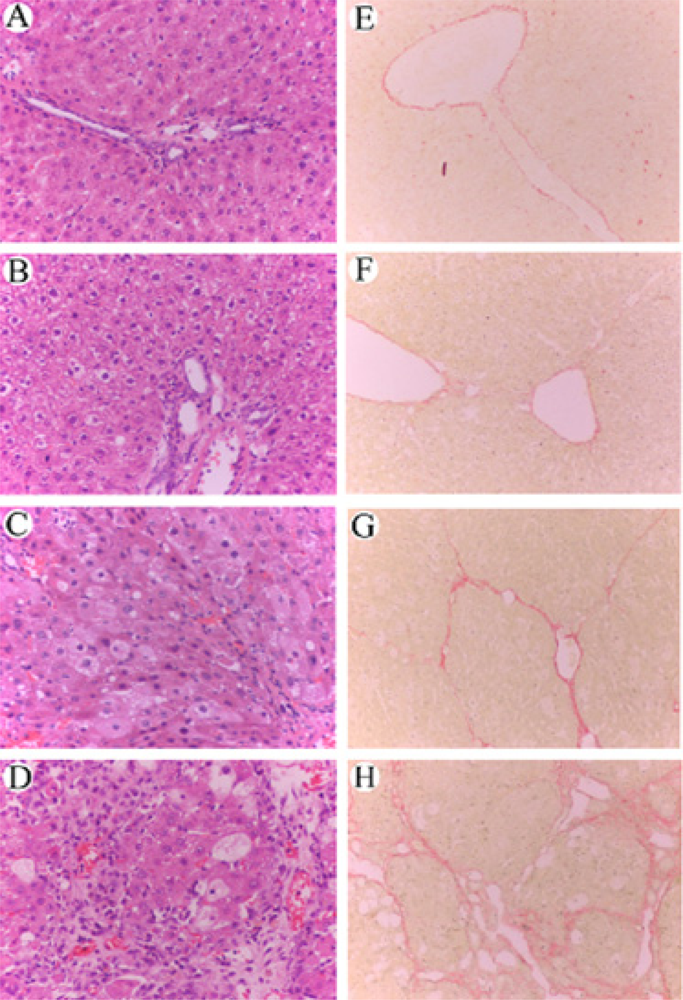
2.6 Histological examination
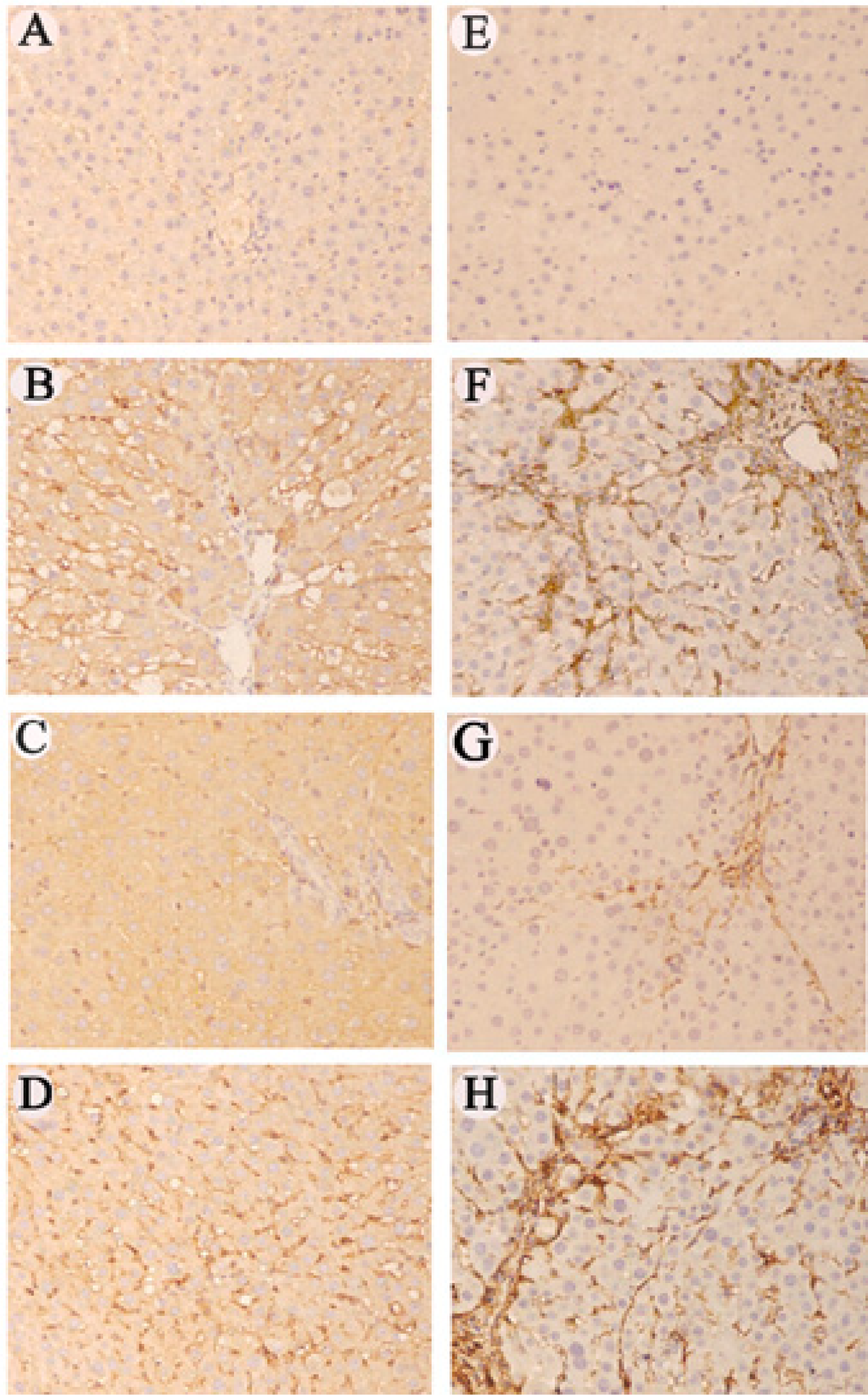
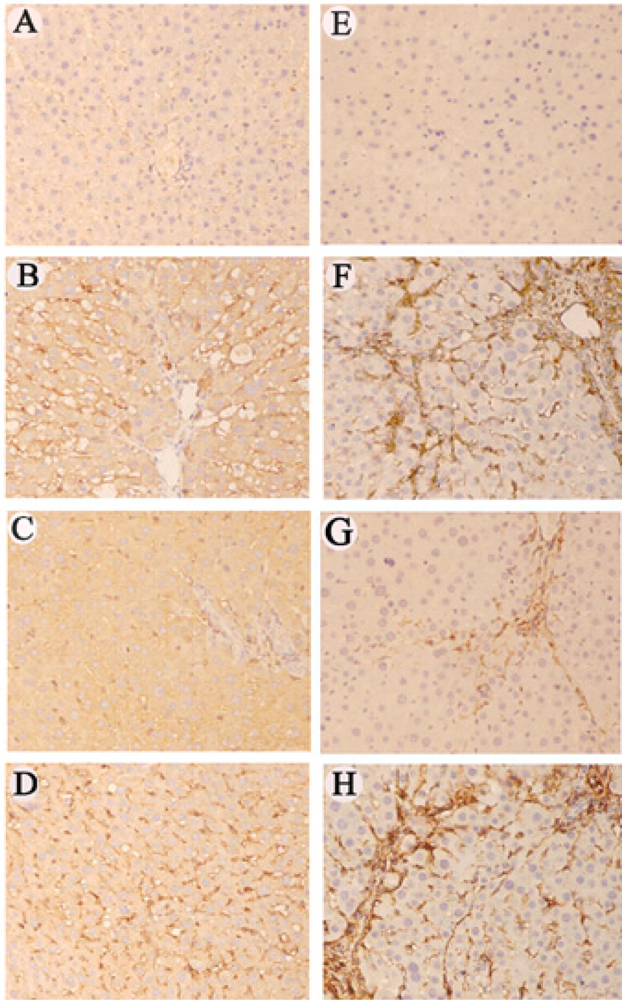
2.7 Immunohistochemical staining of RAGE and α-SMA
2.8 Transmission electron microscopy
2.9 Biochemical analysis of serum
2.10 Total RNA extraction and quantitative real-time PCR assay
2.11 Northern blot analysis
2.12 Western blot analysis
2.13 Electrophoretic mobility shift assay (EMSA)
2.14 Statistical analysis
3. Results
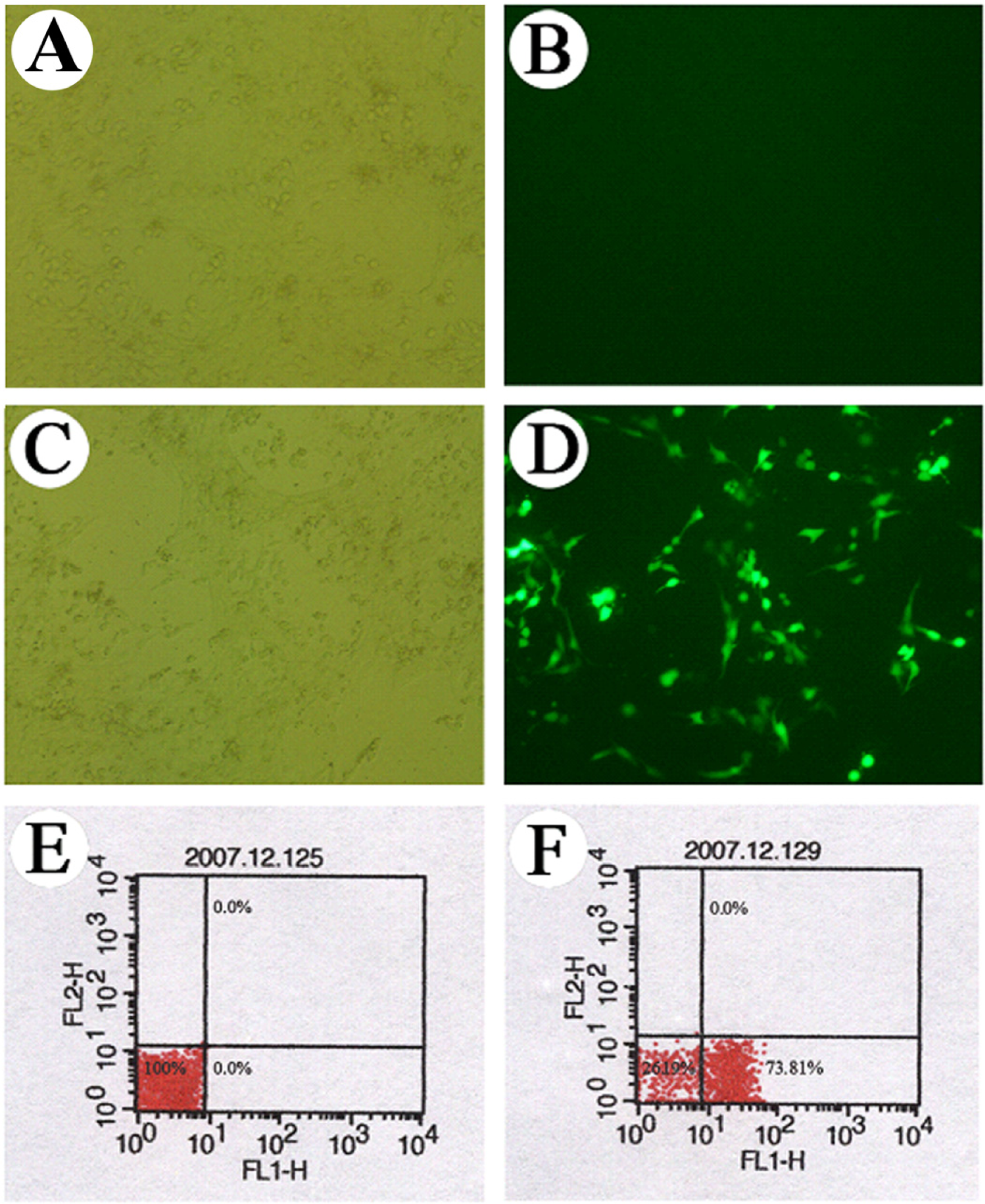
3.1 Observation of the GFP expression in the transfected cells
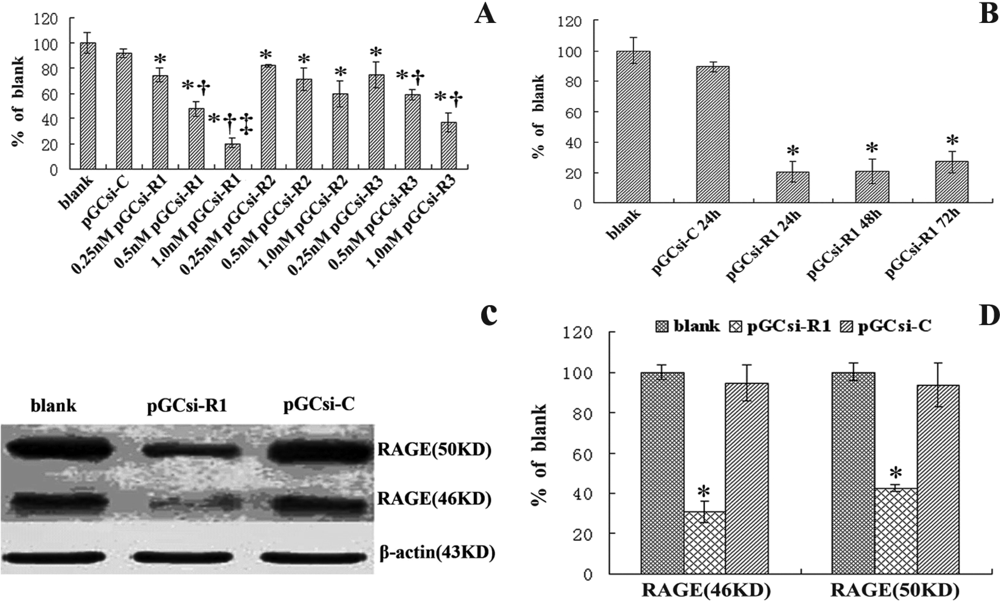
3.2 Effect of RAGE specific siRNA on RAGE expression in HSC-T6 cells
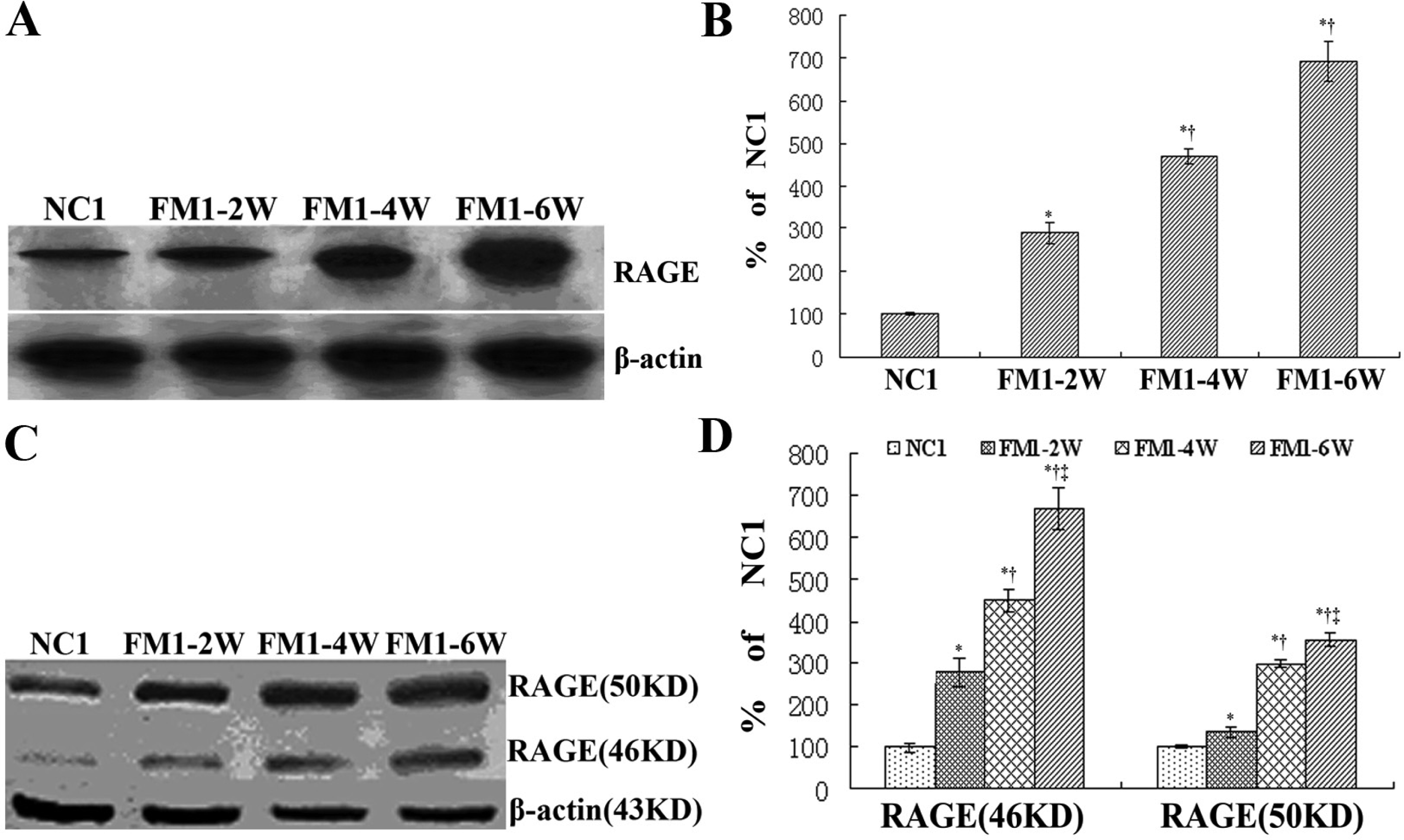
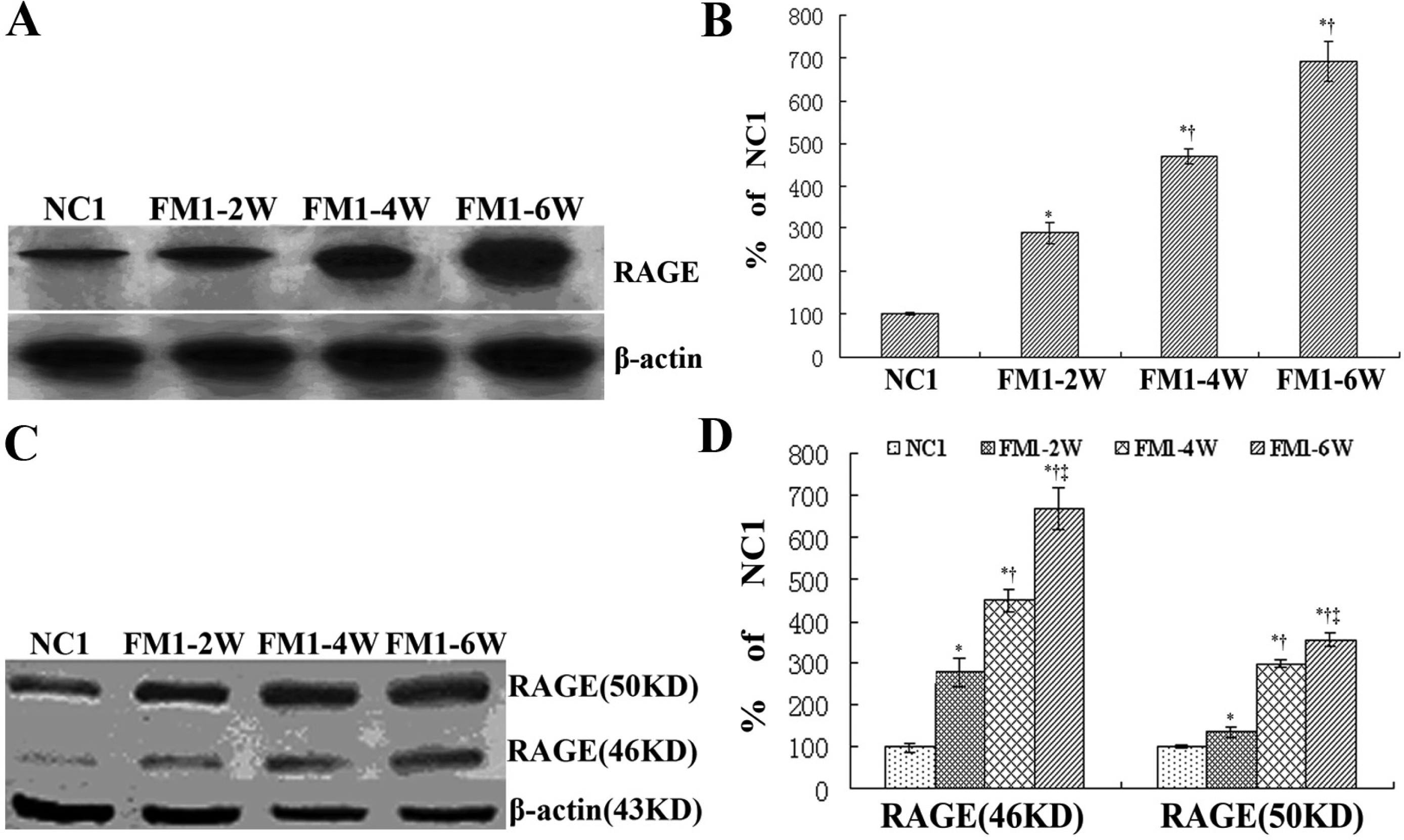
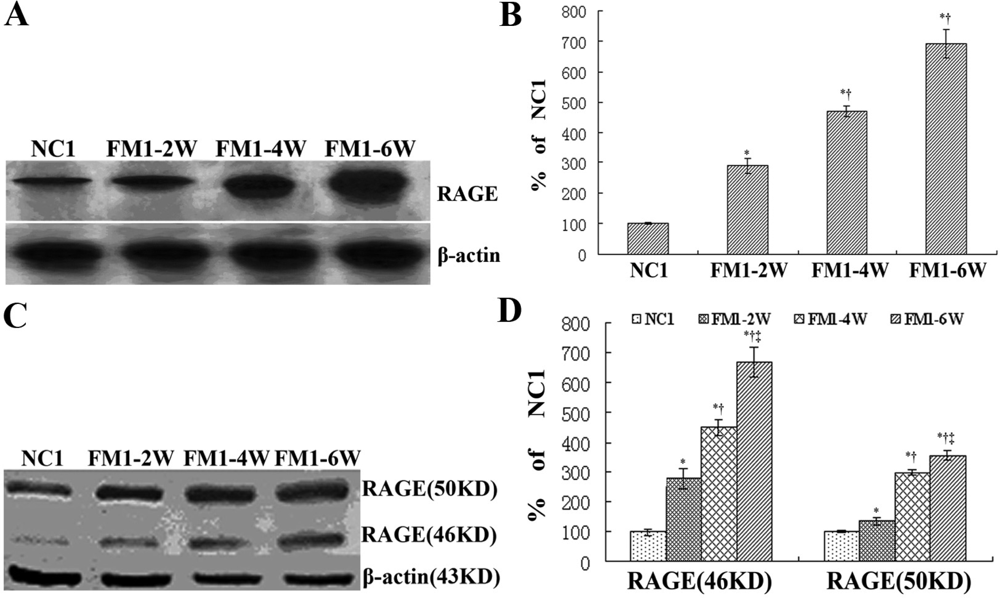
3.3 RAGE expression in the process of hepatic fibrosis
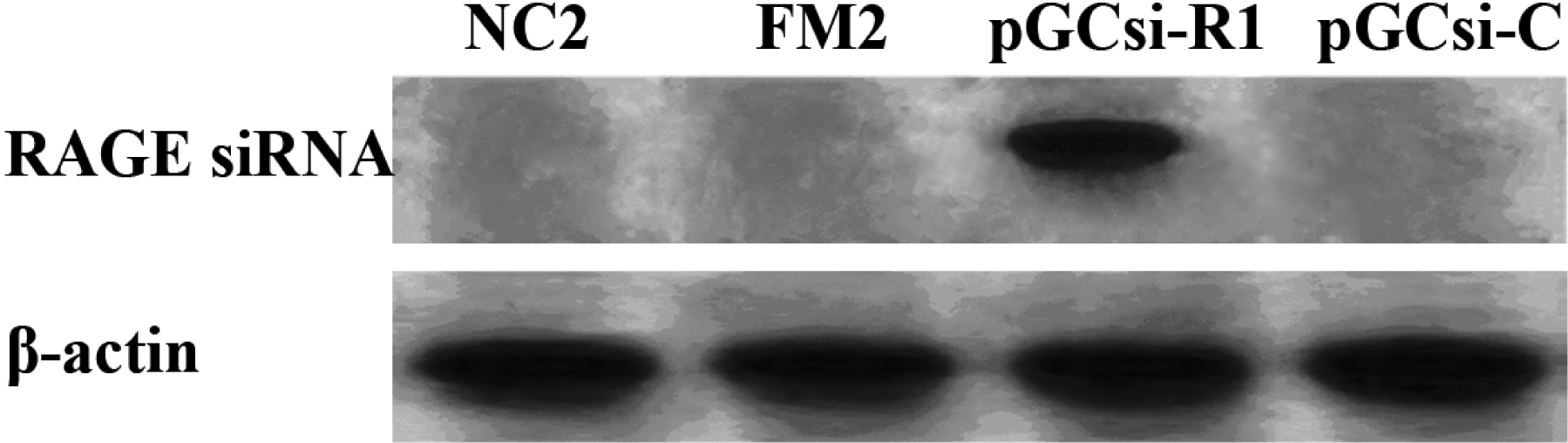
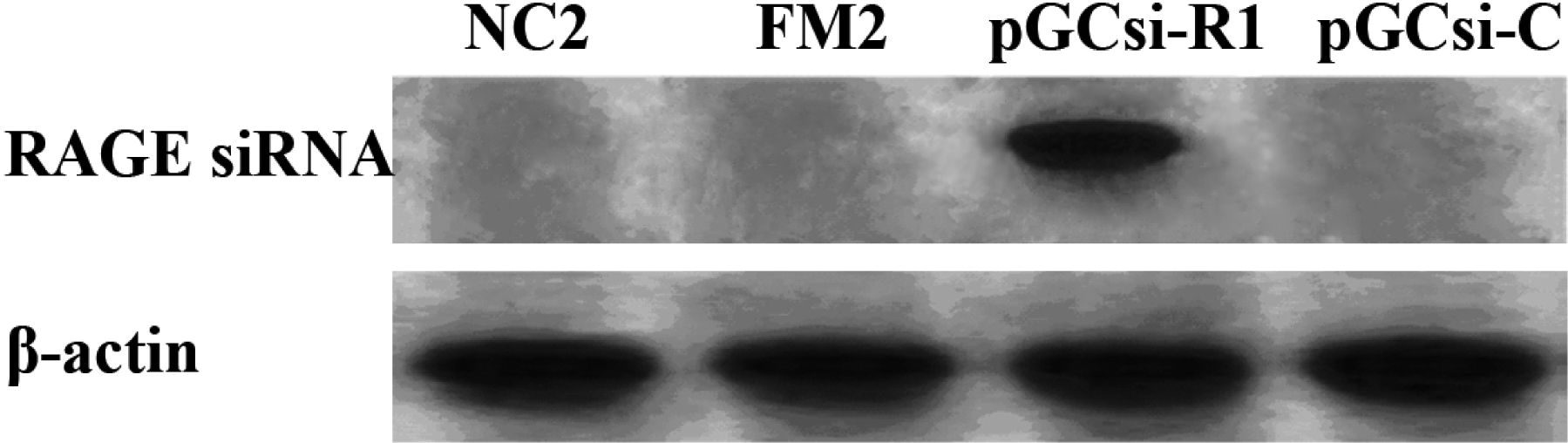
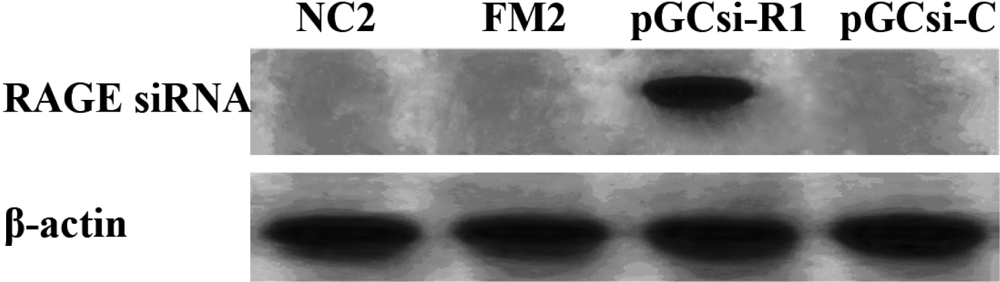
3.4 Expression of RAGE siRNA in the rat liver
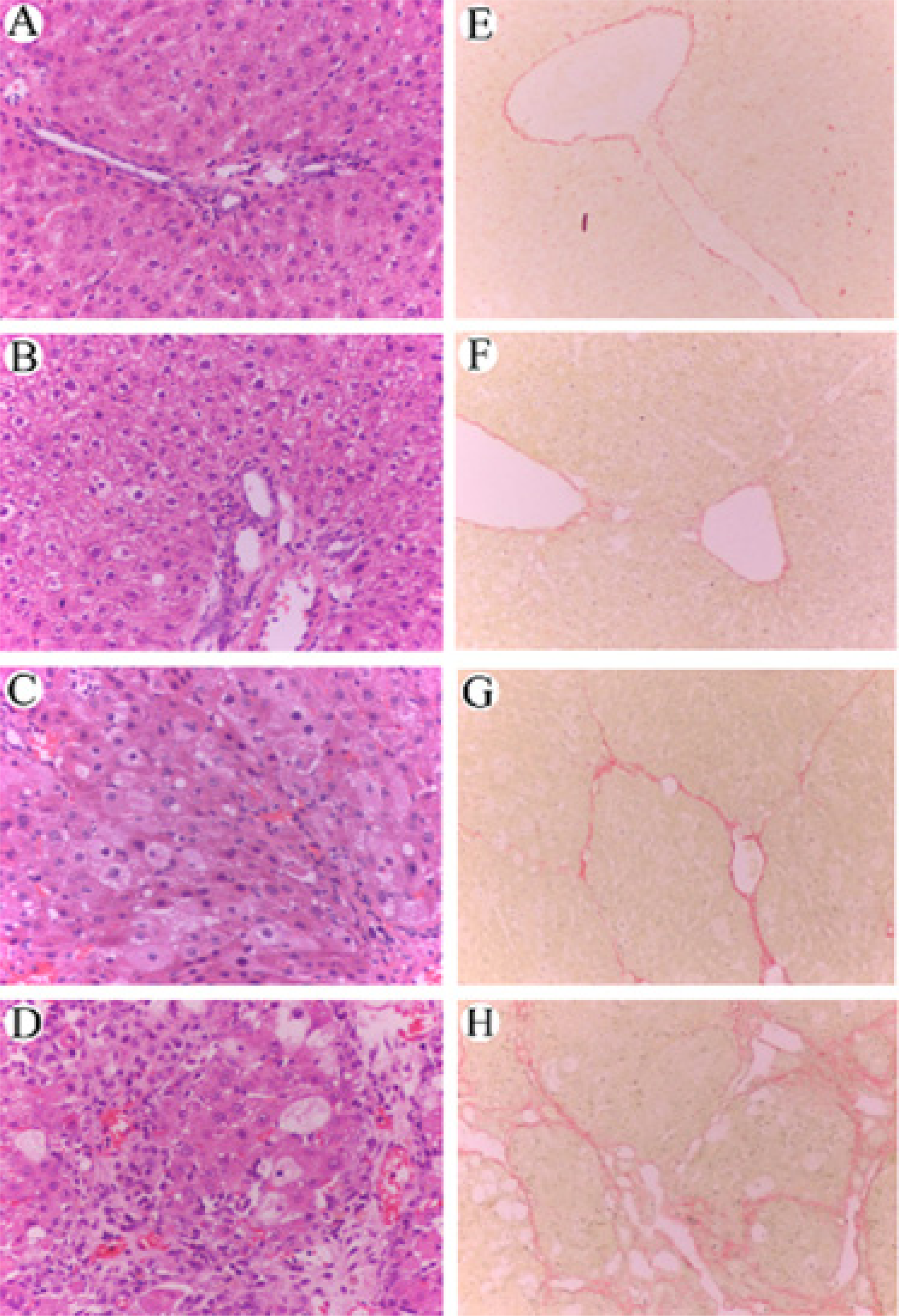
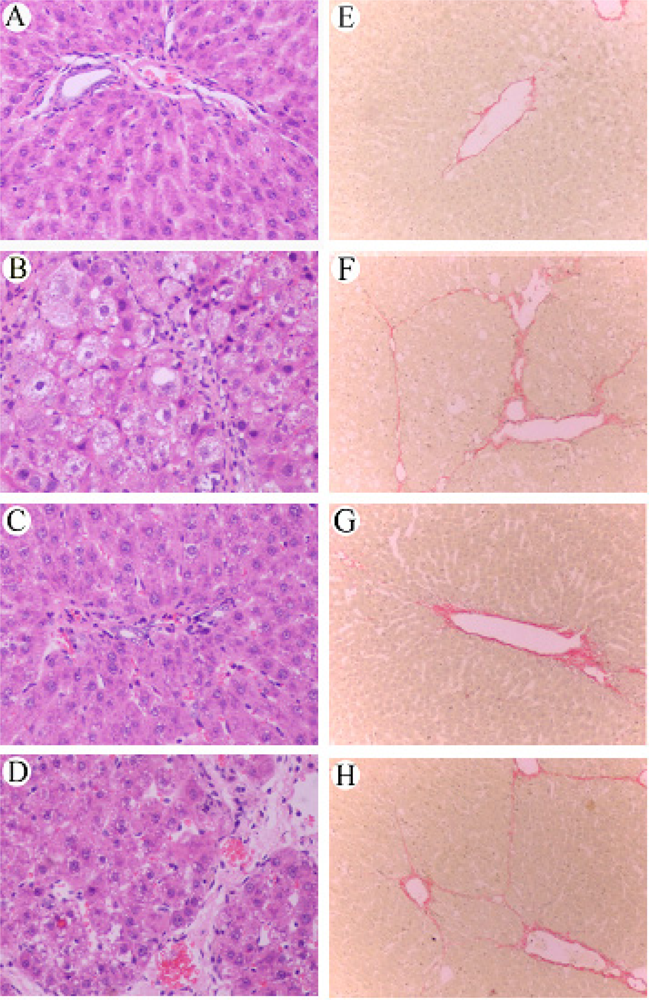
3.5 Effect of RAGE suppression on the histological changes of rat liver
3.6 Effect of RAGE suppression on the levels of ALT, ALB and TBIL in rats
3.7 RAGE siRNA significantly reduced the levels of serum fibrosis markers in rats
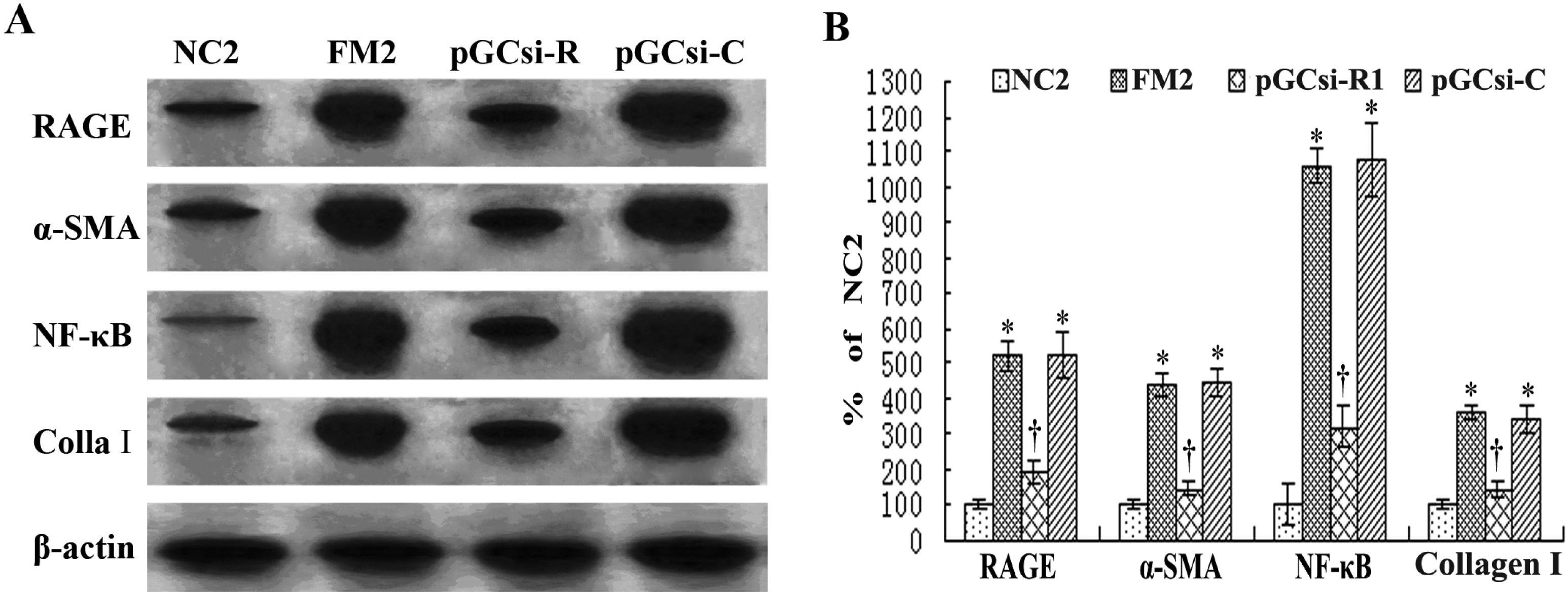
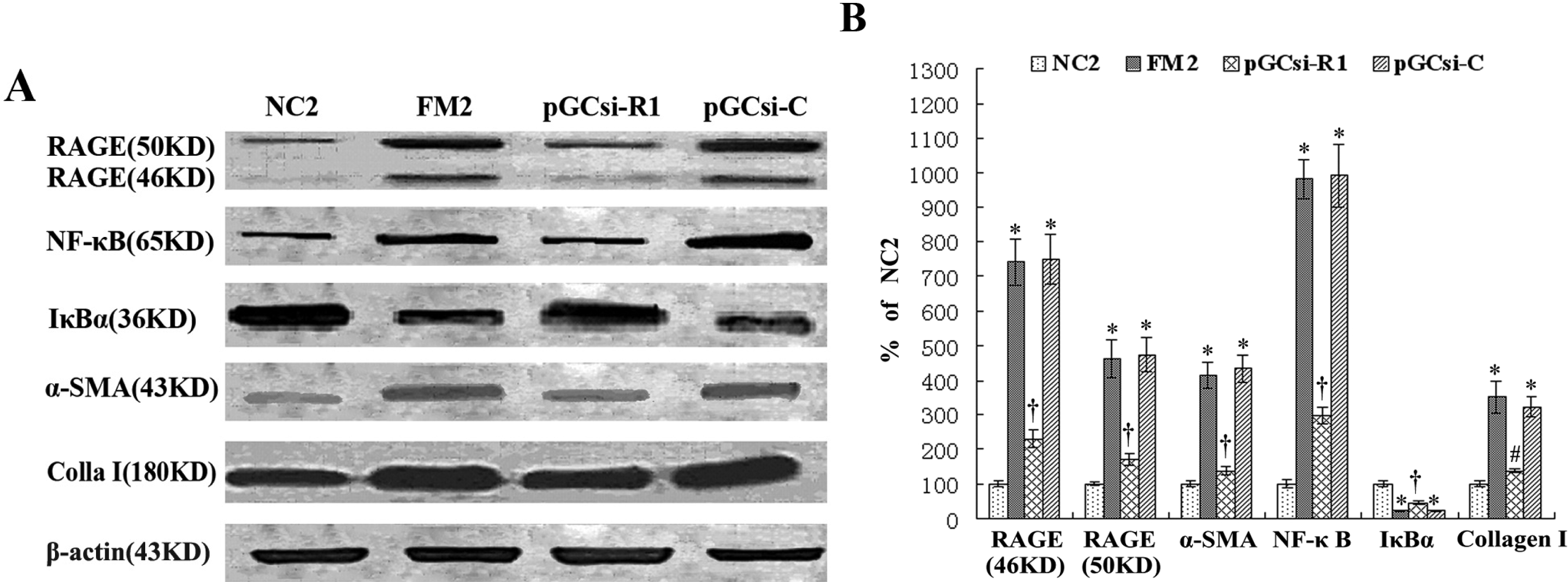
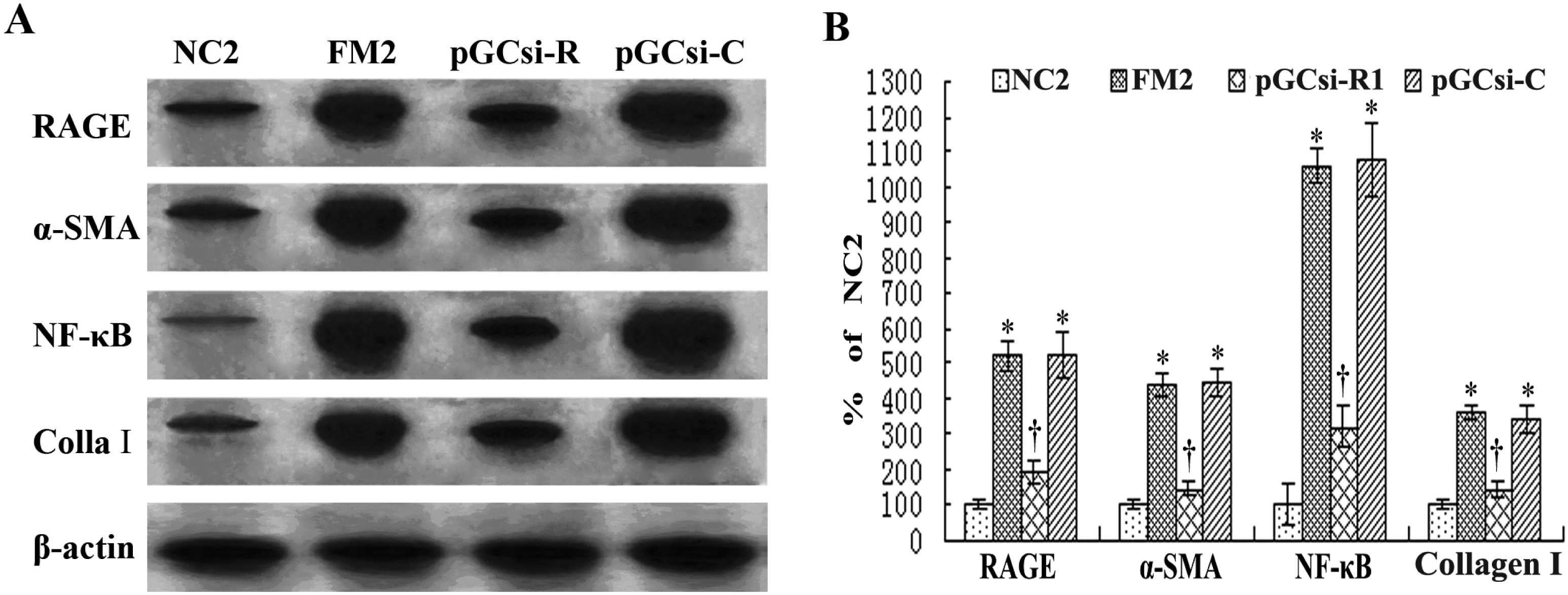
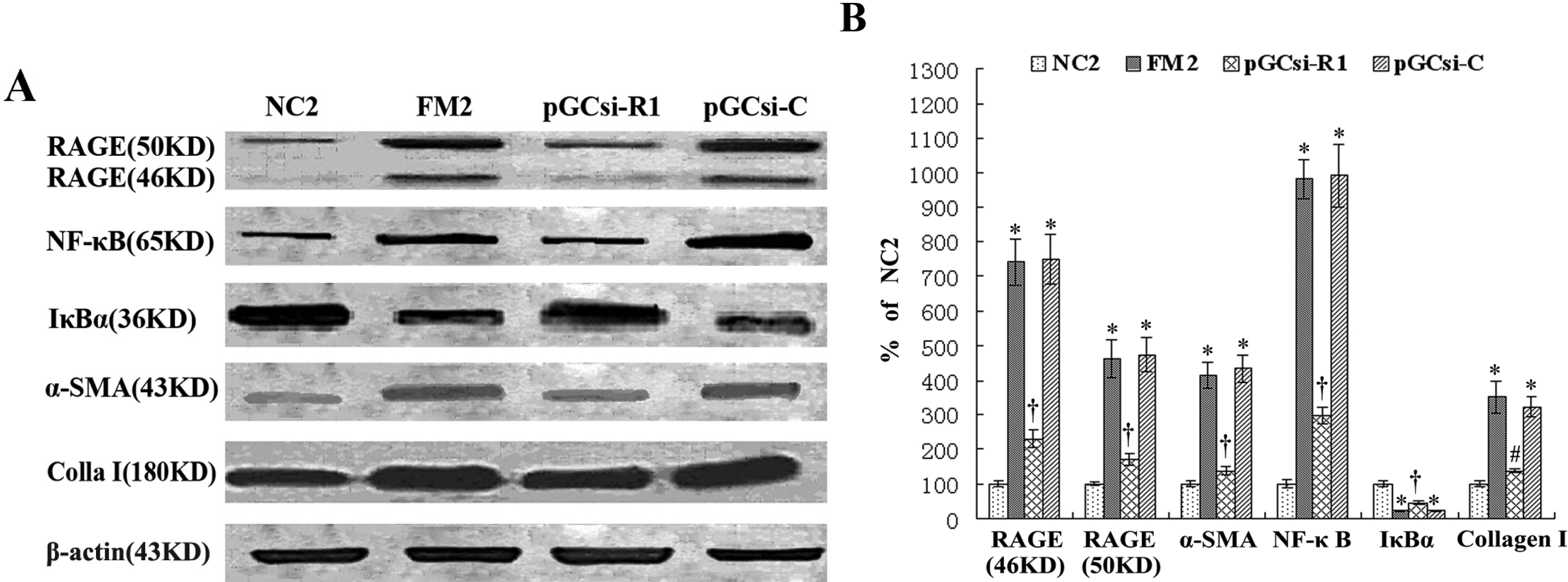
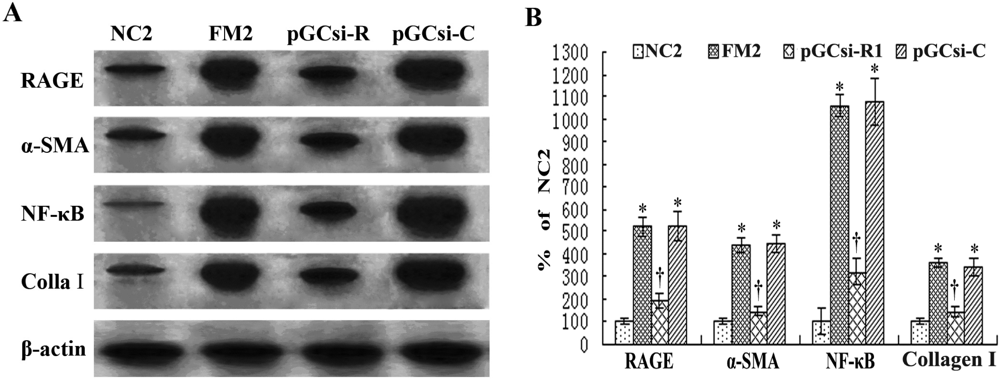
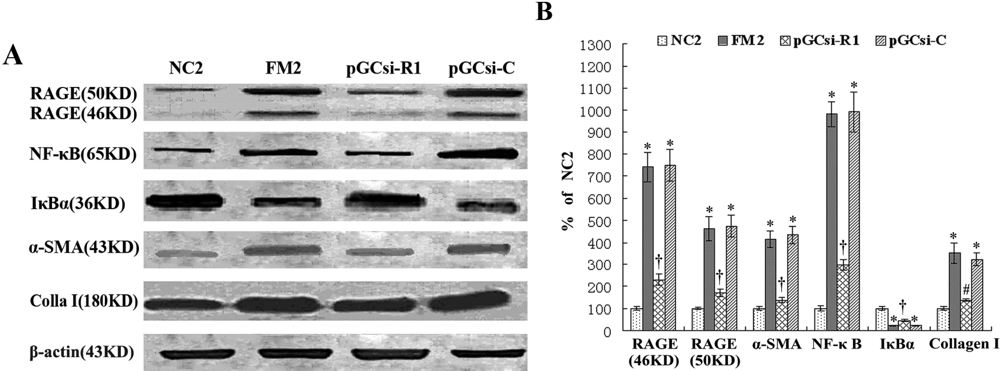
3.8 RAGE siRNA inhibited the expression of RAGE, NF-κB, IκBα, α-SMA and collagen I in rats
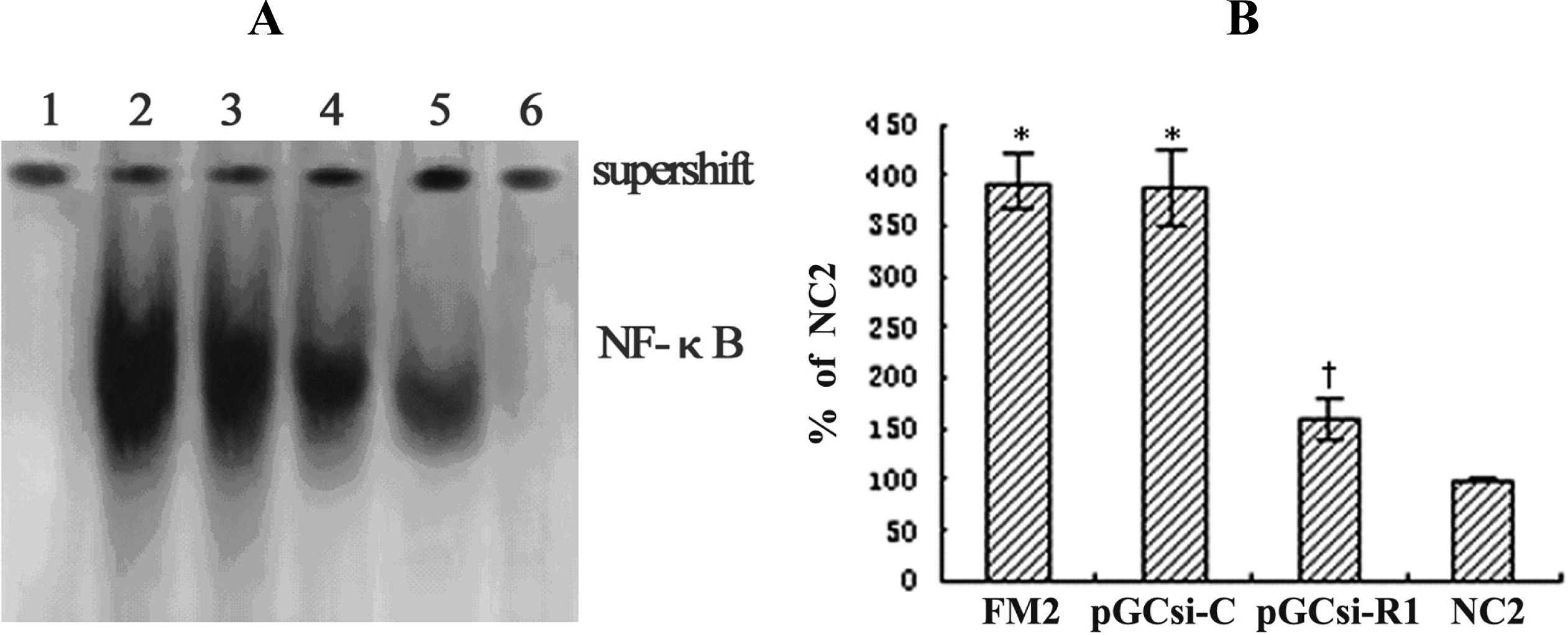
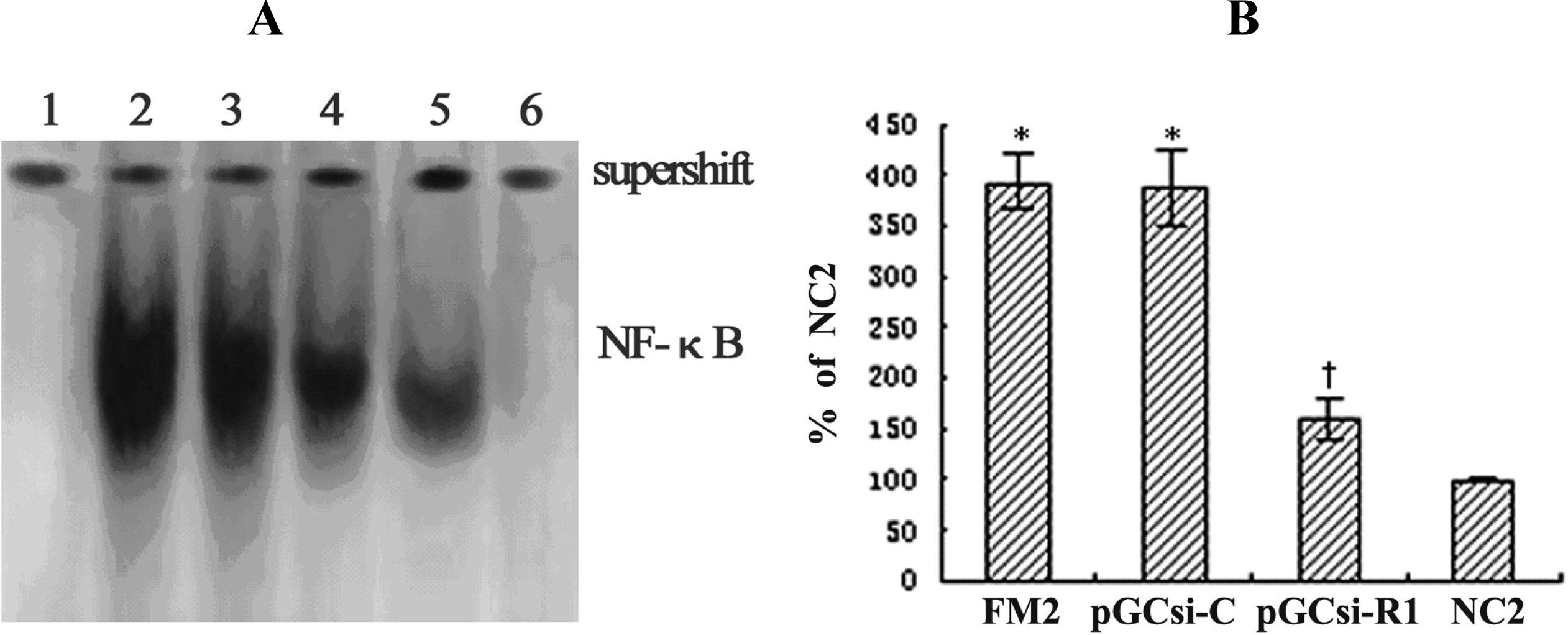
3.9 RAGE siRNA inhibited NF-κB activation in the fibrotic liver of rats
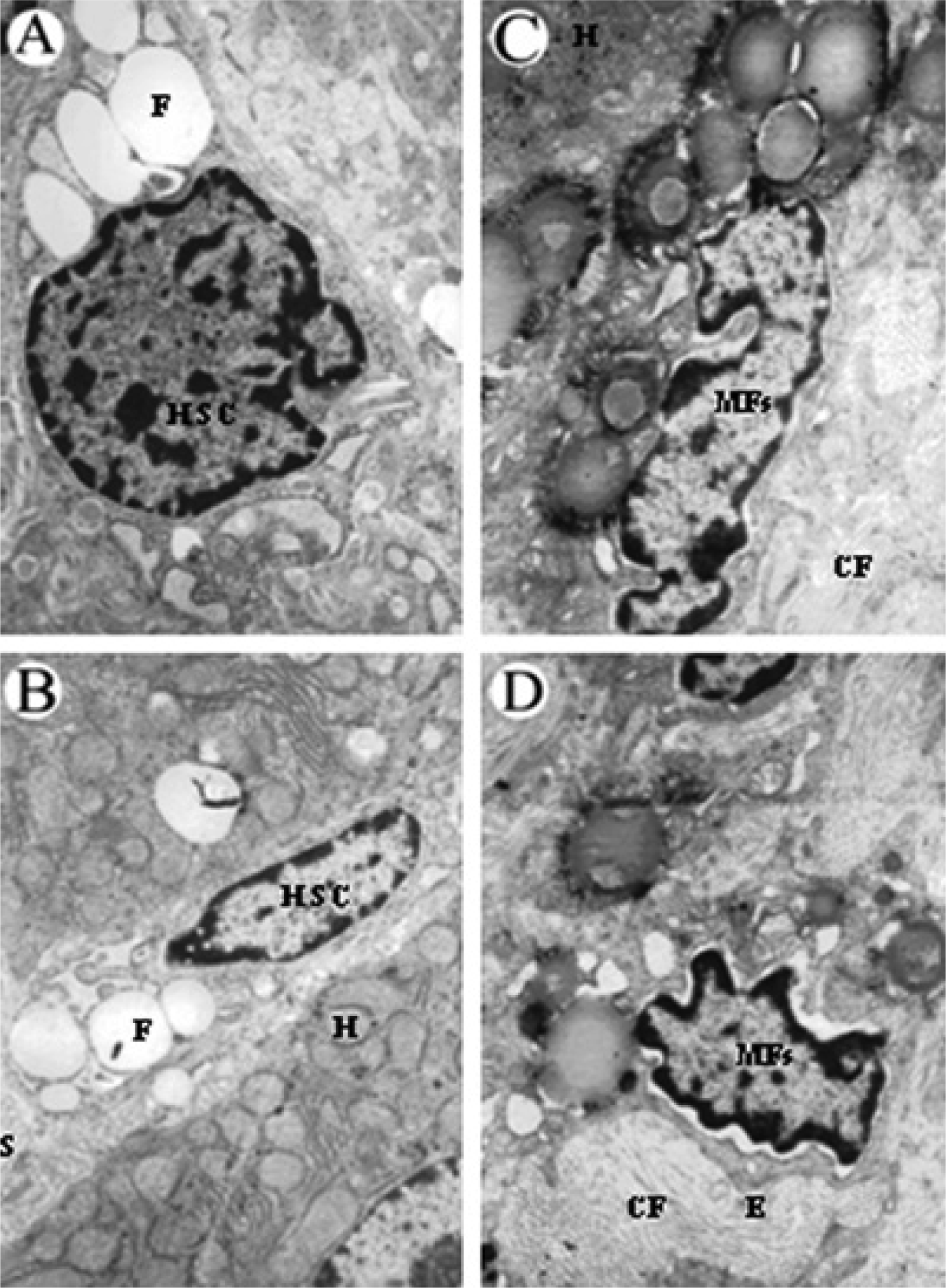
3.10 Effects of RAGE siRNA on changes in ultrastructure of rat liver
Discussion
Acknowledgments
References
- Friedman, SL. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J Biol Chem 2000, 275, 2247–2250. [Google Scholar]
- Pinzani, M. Liver fibrosis. Springer Semin Immunopathol 1999, 21, 475–490. [Google Scholar]
- Moreira, PK. Hepatic stellate cells and liver fibrosis. Arch Pathol Lab Med 2007, 131, 1728–1734. [Google Scholar]
- Reeves, HL; Friedman, SL. Activation of hepatic stellate cells-a key issue in liver fibrosis. Front Biosci 2002, 7, d808–826. [Google Scholar]
- Fehrenbach, H; Weiskirchen, R; Kasper, M; Gressner, AM. Up-regulation expression of the receptor for advanced glycation end products in cultured rat hepatic stellate cells during transdifferentiation to myofibroblasts. Hepatology 2001, 34, 943–952. [Google Scholar]
- Schmidt, AM; Yan, SD; Yan, SF; Stern, DM. The biology of the receptor for advanced glycation end products and its ligands. Biochim Biophys Acta 2000, 1498, 99–111. [Google Scholar]
- Kislinger, T; Fu, C; Huber, B; Qu, W; Taguchi, A; Du, Yan S; Hofmann, M; Yan, SF; Pischetsrieder, M; Stern, D; Schmidt, AM. N(epsilon)- (carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signalling pathways and modulate gene expression. J Biol Chem 1999, 274, 31740–31749. [Google Scholar]
- Lander, HM; Tauras, JM; Ogiste, JS; Hori, O; Moss, RA; Schmidt, AM. Activation of the receptor for advanced glycation end products triggers a p21(ras)-dependent mitogen-activated protein kinase pathway regulated by oxidant stress. J Biol Chem 1997, 272, 17810–17814. [Google Scholar]
- Yan, SD; Schmidt, AM; Anderson, GM; Chang, J; Brett, J; Zou, YS; Pinsky, D; Stern, D. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J Biol Chem 1994, 269, 9889–9897. [Google Scholar]
- Oldfield, MD; Bach, LA; Forbes, JM; Nikolic-Paterson, D; McRobert, A; Thallas, V; Atkins, RC; Osicka, T; Jerums, G; Cooper, ME. Advanced glycation end products cause epithelial-myofibroblast transdifferentiation via the receptor for advanced glycation end products (RAGE). J Clin Invest 2001, 108, 1853–1863. [Google Scholar]
- De Vriese, AS; Tilton, RG; Mortiers, S; Lameire, NH. Myofibroblast transdifferentiation of mesothelial cells is mediated by RAGE and contributes to peritoneal fibrosis in uraemia. Nephrol Dial Transplant 2006, 21, 2549–2555. [Google Scholar]
- Sebekova, K; Kupcova, V; Schinzel, R; Heidland, A. Markedly elevated levels of plasma advanced glycation end products in patients with liver cirrhosis-amelioration by liver transplantation. J Hepatol 2002, 36, 66–71. [Google Scholar]
- Ahmed, N; Luthen, R; Haussinger, D; Sebekova, K; Schinzel, R; Voelker, W; Heidland, A; Thornalley, PJ. Increased protein glycation in cirrhosis and therapeutic strategies to prevent it. Ann N Y Acad Sci 2005, 1043, 718–724. [Google Scholar]
- Yagmur, E; Tacke, F; Weiss, C; Lahme, B; Manns, MP; Kiefer, P; Trautwein, C; Gressner, AM. Elevation of Nepsilon-(carboxymethyl)lysine-modified advanced glycation end products in chronic liver disease is an indicator of liver cirrhosis. Clin Biochem 2006, 39, 39–45. [Google Scholar]
- Zeng, S; Feirt, N,; Goldstein, M; Guarrera, J; Ippagunta, N; Ekong, U; Dun, H; Lu, Y; Qu, W; Schmidt, AM; Emond, JC. Blockade of receptor for advanced glycation end products (RAGE) attenuates ischemia and reperfusion injury to the liver in mice. Hepatology 2004, 39, 422–432. [Google Scholar]
- May, MJ; Ghosh, S. Rel/NF-kappaB and I kappaB proteins: an overview. Semin Cancer Biol 1997, 8, 63–73. [Google Scholar]
- Lawrence, T; Bebien, M; Liu, GY; Nizet, V; Karin, M. IKKalpha limits macrophage NF-kappaB activation and contributes to the resolution of inflammation. Nature 2005, 434, 1138–1143. [Google Scholar]
- Wright, MC; Issa, R; Smart, DE; Trim, N; Murray, GI; Primrose, JN; Arthur, MJ; Iredale, JP; Mann, DA. Gliotoxin stimulates the apoptosis of human and rat hepatic stellate cells and enhances the resolution of liver fibrosis in rats. Gastroenterology 2001, 121, 685–698. [Google Scholar]
- Oakley, F; Meso, M; Iredale, JP; Green, K; Marek, CJ; Zhou, X; May, MJ; Millward-Sadler, H; Wright, MC; Mann, DA. Inhibition of inhibitor of kappaB kinases stimulates hepatic stellate cell apoptosis and accelerated recovery from rat liver fibrosis. Gastroenterology 2005, 128, 108–120. [Google Scholar]
- Bierhaus, A; Schiekofer, S; Schwaninger, M; Andrassy, M; Humpert, PM; Chen, J; Hong, M; Luther, T; Henle, T; Kloting, I; Morcos, M; Hofmann, M; Tritschler, H; Weigle, B; Kasper, M; Smith, M; Perry, G; Schmidt, AM; Stern, DM; Haring, HU; Schleicher, E; Nawroth, PP. Diabetes-associated sustained activation of the transcription factor nuclear factor-kappa B. Diabetes 2001, 50, 2792–2808. [Google Scholar]
- Knodell, RG; Ishak, KG; Black, WC; Chen, TS; Craig, R; Kaplowitz, N; Kiernan, TW; Wollman, J. Formulation and application of a numerical scoring system for assessing histological activity in asymptomatic chronic active hepatitis. Hepatology 1981, 1, 431–435. [Google Scholar]
- Ishak, K; Baptista, A; Bianchi, L; Callea, F; De Groote, J; Gudat, F; Denk, H; Desmet, V; Korb, G; MacSween, RN; et al. Histological grading and staging of chronic hepatitis. J Hepatol 1995, 22, 696–699. [Google Scholar]
- Ryhanen, L; Stenback, F; Ala-Kokko, L; Savolainen, ER. The effect of malotilate on type III and type IV collagen, laminin and fibronectin metabolism in dimethylnitrosamine-induced liver fibrosis in the rat. J Hepatol 1996, 24, 238–245. [Google Scholar]
- Gabrielli, GB; Capra, F; Casaril, M; Squarzoni, S; Tognella, P; Dagradi, R; De Maria, E; Colonbari, R. Serum laminin and type III precollagen in chronic hepatitis C. Diagnostic value in the assessment of disease activity and fibrosis. Chin Chim Acta 1997, 265, 21–31. [Google Scholar]
- Castera, L; Hartmann, DJ; Chapel, F; Guettier, C; Mall, F; Lons, T; Richardet, JP; Grimbert, S; Morassi, O; Beaugrand, M; Trinchet, JC. Serum laminin and type IV collagen are accurate markers of histologically severe alcoholic hepatitis in patients with cirrhosis. J Hepatol 2000, 32, 412–418. [Google Scholar]
- Liu, Z; Yu, Y; Jiang, Y; Li, J. Growth hormone increases lung NF-kappaB activation and lung microvascular injury induced by lipopolysaccharide in rats. Ann Clin Lab Sci 2002, 32, 164–170. [Google Scholar]
- Li, X; Meng, Y; Wu, P; Zhang, Z; Yang, X. Angiotensin II and Aldosterone stimulating NF-kappaB and AP-1 activation in hepatic fibrosis of rat. Regul Pept 2007, 138, 15–25. [Google Scholar]
- Bierhaus, A; Hofmann, MA; Ziegler, R; Nawroth, PP. AGEs and their interaction with AGE-receptors in vascular disease and diabetes mellitus. I. The AGE concept. Cardiovasc Res 1998, 37, 586–600. [Google Scholar]
- Baeuerle, PA; Henkel, T. Function and activation of NF-kappaB in the immune system. Annu Rev Immunol 1994, 12, 141–179. [Google Scholar]
- Yamamoto, Y; Kato, I; Doi, T; Yonekur, H; Ohashi, S; Takeuchi, M; Watanabe, T; Yamageshi, S; Sakurai, S; Takasawa, S; Okamoto, H; Yamamoto, H. Development and prevention of advanced diabetic nepropathy in RAGE-overexpressing mice. J Clin Invest 2001, 108, 261–268. [Google Scholar]
- Singh, R; Barden, A; Mori, T; Beilin, L. Advanced glycation end-products:a review. Diabetologia 2001, 44, 129–146. [Google Scholar]
- Kim, KH; Kim, HC; Hwang, MY; Oh, HK; Lee, TS; Chang, HJ; Song, HJ; Won, NH; Park, KK. The antifibrotic effect of TGF-β1 siRNAs in murine modle of liver cirrhosis. Biochem Biophys Res Commun 2006, 343, 1072–1078. [Google Scholar]
- Hu, YB; Li, DG; Lu, HM. Modified synthetic siRNA targeting tissue inhibitor of metalloproteinase-2 inhibits hepatic fibrogenesis in rats. J Gene Med 2007, 9, 217–229. [Google Scholar]
- Wautier, MP; Chappey, O; Corda, S; Stern, DM; Schmidt, AM; Wautier, JL. Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. Am J Physiol Endocrinol Metab 2001, 280, E685–694. [Google Scholar]
- Zill, H; Bek, S; Hofmann, T; Huber, J; Frank, O; Lindenmeier, M; Weigle, B; Erbersdobler, HF; Scheidler, S; Busch, AE; Faist, V. RAGE-mediated MAPK activation by food-derived AGE and non-AGE products. Biochen Biophys Res Commun 2003, 300, 311–315. [Google Scholar]
- Taguchi, A; Blood, DC; del Toro, G; Canet, A; Lee, DC; Qu, W; Tanji, N; Lu, Y; Lalla, E; Fu, C; Hofmann, MA; Kislinger, T; Ingram, M; Lu, A; Tanaka, H; Hori, O; Ogawa, S; Stern, DM; Schmidt, AM. Blockade of RAGE-amphoterin signalling suppresses tumour growth and metastases. Nature 2000, 405, 354–360. [Google Scholar]
- Bataller, R; Brenner, DA. Liver fibrosis. J Clin Invest 2005, 115, 209–218. [Google Scholar]
- Schwabe, RF; Schnabl, B; Kweon, YO; Brenner, DA. CD40 activates NF-kappaB and c-Jun N-terminal kinase and enhances chemokine secretion on activated human hepatic stellate cells. J Immunol 2001, 166, 6812–6819. [Google Scholar]
- Luedde, T; Beraza, N; Trautwein, C. Evaluation of the role of nuclear factor-kappaB signaling in liver injury using genetic animal models. J Gastroenterol Hepatol 2006, (Suppl 3), S43–S46. [Google Scholar]
- Dang, SS; Wang, BF; Cheng, YA; Song, P; Liu, ZG; Li, ZF. Inhibitory effects of saikosaponin-d on CCl4-induced hepatic fibrogenesis in rats. World J Gastroenterol 2007, 13, 557–563. [Google Scholar]
- Reynaert, H; Thompson, MG; Thomas, T; Geerts, A. Hepatic stellate cells: role in microcirculation and pathophysiology of portal hypertension. Gut 2002, 50, 571–581. [Google Scholar]
- Shi, GF; Li, Q. Effects of oxymatrine on experimental hepatic fibrosis and its mechanism in vivo. World J Gastroenterol 2005, 11, 268–271. [Google Scholar]
- Singh, KP; Gerard, HC; Hudson, AP; Boros, DL. Expression of matrix metalloproteinases and their inhibitors during the resorption of schistosome egg-induced fibrosis in praziquantel-treated mice. Immunology 2004, 111, 343–352. [Google Scholar]
- Okamoto, K; Mimura, K; Murawaki, Y; Yuasa, I. Association of functional gene polymorphisms of matrix metalloproteinase (MMP)-1, MMP-3 and MMP-9 with the progression of chronic liver disease. J Gastroenterol Hepatol 2005, 20, 1102–1108. [Google Scholar]
- Solis-Herruzo, JA; de la Torre, P; Munoz-Yague, MT. Hepatic stellate cells (HSC): architects of hepatic fibrosis. Rev Esp Enferm Dig 2003, 95. [Google Scholar]
- Urtasun, R; Nieto, N. Hepatic stellate cells and oxidative stress. Rev Esp Enferm Dig 2007, 99, 223–230. [Google Scholar]
- Wang, JH; Batey, RG; George, J. Role of ethanol in the regulation of hepatic stellate cell function. World J Gastroenterol 2006, 12, 6926–6932. [Google Scholar]
- Friedman, SL. Liver fibrosis-from bench to bedside. J Hepatol 2003, 38, S38–S53. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Inflammatory gradation
| Average rank | Fibrosis staging | Average rank | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | I | II | III | IV | 0 | I | II | III | IV | ||||
| NC2 | 6 | 6 | 0 | 0 | 0 | 0 | 4.50 | 6 | 0 | 0 | 0 | 0 | 4.50 |
| FM2 | 9 | 0 | 0 | 2 | 3 | 4 | 24.67* | 0 | 0 | 2 | 3 | 4 | 24.78* |
| pGCsi-R1 | 9 | 2 | 5 | 2 | 0 | 0 | 10.78*† | 2 | 6 | 1 | 0 | 0 | 10.56*† |
| pGCsi-C | 9 | 0 | 0 | 2 | 4 | 3 | 23.89* | 0 | 0 | 2 | 4 | 3 | 24.00* |
| Group | n | ALT (u/L) | ALB (g/L) | TBIL (μmol/L) |
|---|---|---|---|---|
| NC2 | 6 | 37.83±3.49 | 18.08±0.58 | 5.83±0.39 |
| FM2 | 6 | 363.17±56.95* | 11.52±0.68* | 25.25±1.54* |
| pGCsi-R1 | 9 | 46.00±1.79*† | 17.33±0.75† | 10.00±1.70*† |
| pGCsi-C | 9 | 373.50±59.73* | 11.90±0.85* | 25.50±3.27* |
| Group | n | PCIII (μg/L) | HA(μg/L) | LN(μg/L) |
|---|---|---|---|---|
| NC2 | 6 | 127.33±15.98 | 102.16±7.99 | 99.90±14.00 |
| FM2 | 6 | 342.14±19.04* | 424.63±31.82* | 221.83±20.06* |
| pGCsi-R1 | 9 | 180.72±19.19*† | 180.89±40.01*† | 150.41±13.25*† |
| pGCsi-C | 9 | 338.06±16.50* | 416.30±34.13* | 220.36±26.16* |
| Group | n | PA(μm2) | IOD | AOD |
|---|---|---|---|---|
| NC2 | 6 | 37074±2778 | 8434±857 | 0.228±0.019 |
| FM2 | 6 | 73722±4674* | 148164±3998* | 0.653±0.038* |
| pGCsi-R1 | 9 | 40724±2877† | 12486±1431#† | 0.307±0.033*† |
| pGCsi-C | 9 | 73809±5450* | 48704±3483* | 0.648±0.073* |
Share and Cite
Xia, J.-R.; Liu, N.-F.; Zhu, N.-X. Specific siRNA Targeting the Receptor for Advanced Glycation End Products Inhibits Experimental Hepatic Fibrosis in Rats. Int. J. Mol. Sci. 2008, 9, 638-661. https://doi.org/10.3390/ijms9040638
Xia J-R, Liu N-F, Zhu N-X. Specific siRNA Targeting the Receptor for Advanced Glycation End Products Inhibits Experimental Hepatic Fibrosis in Rats. International Journal of Molecular Sciences. 2008; 9(4):638-661. https://doi.org/10.3390/ijms9040638
Chicago/Turabian StyleXia, Jin-Rong, Nai-Feng Liu, and Nai-Xun Zhu. 2008. "Specific siRNA Targeting the Receptor for Advanced Glycation End Products Inhibits Experimental Hepatic Fibrosis in Rats" International Journal of Molecular Sciences 9, no. 4: 638-661. https://doi.org/10.3390/ijms9040638