Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes In Vitro


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- PDGF-BB and AA increase the proliferation of tenocytes.
- PDGF-BB and AA enhance extracellular matrix synthesis.
- PDGF-BB and AA change the gene expression of tendon markers.
- PDGF-BB and AA released from bioactive scaffolds support tenocyte growth and ECM synthesis.
2. Results
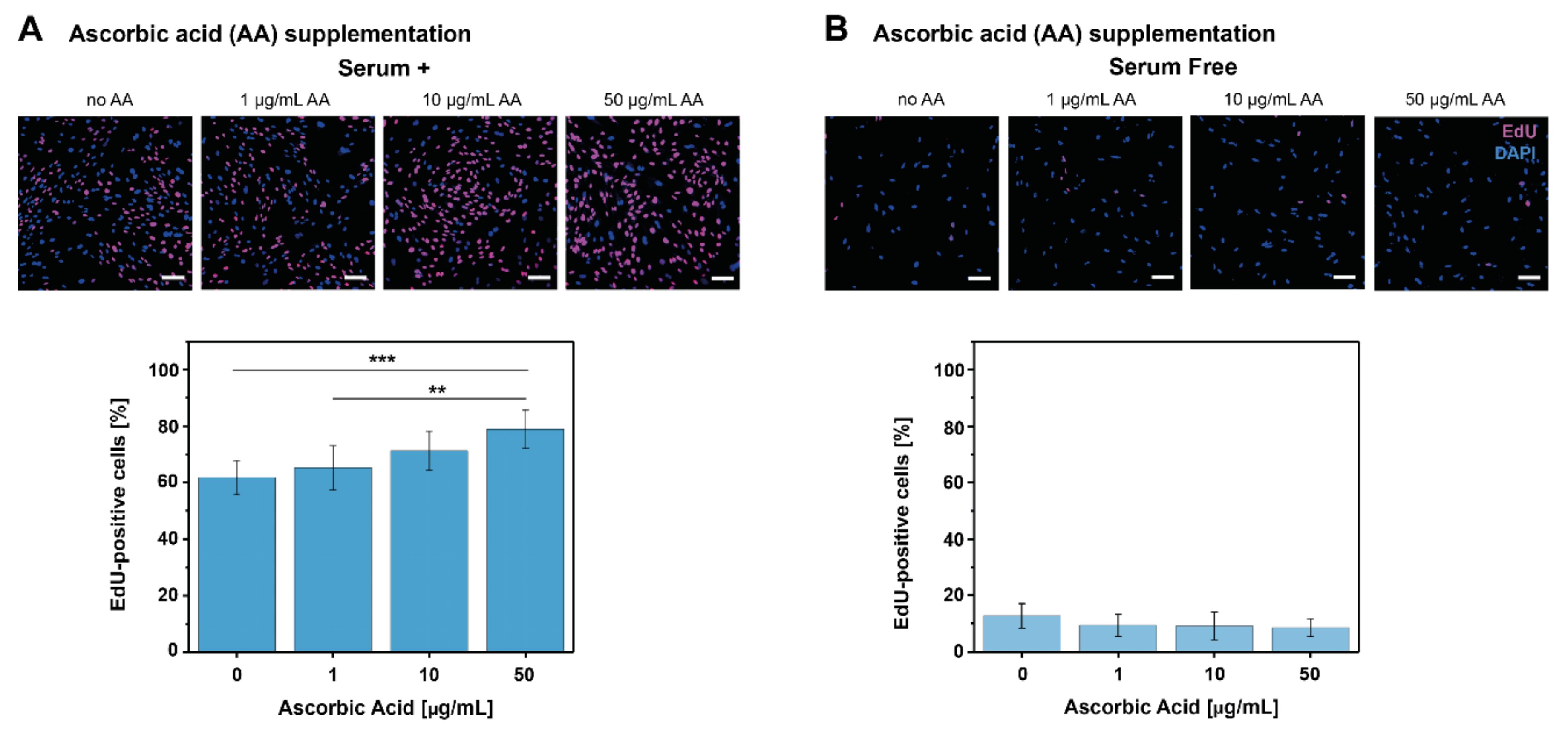
2.1. AA and PDGF-BB Supplementation Increase Tenocyte Proliferation
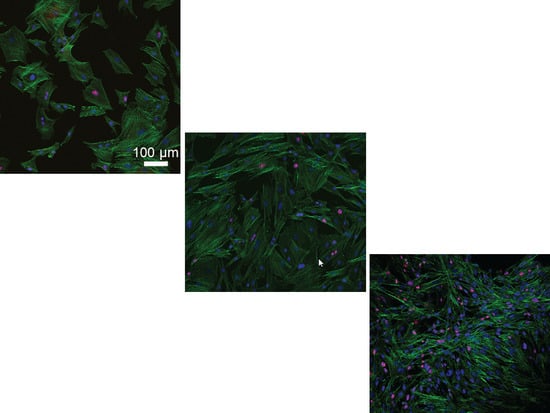
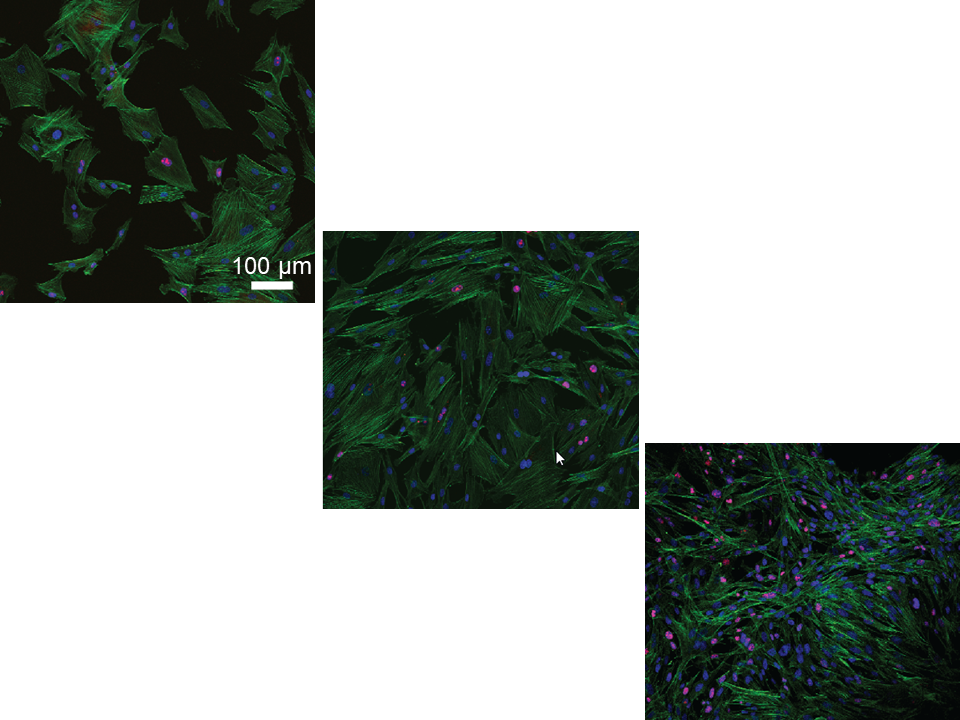
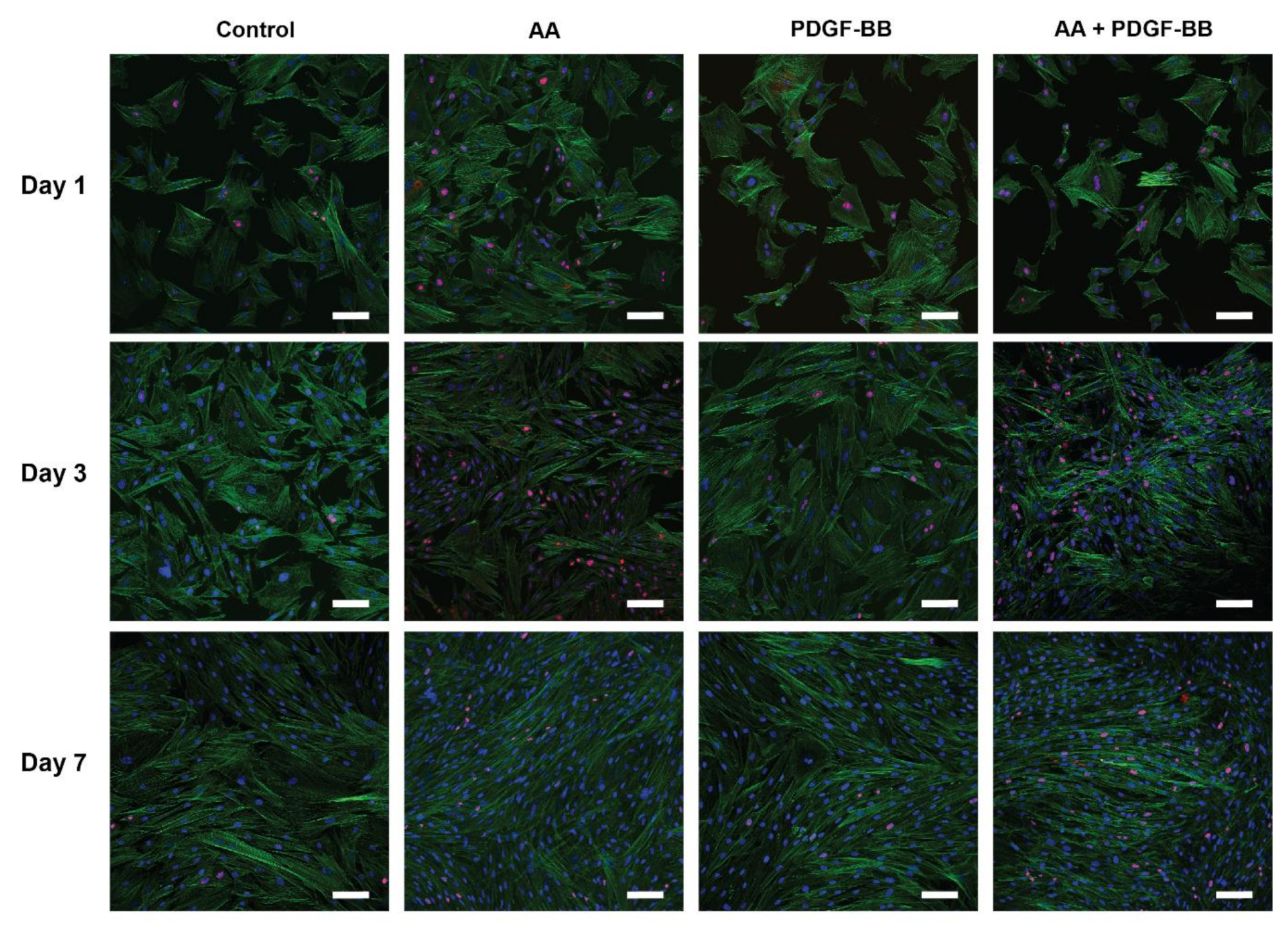
2.2. Tenocyte Morphology and ki-67 Expression Changes under Different Conditions over Time
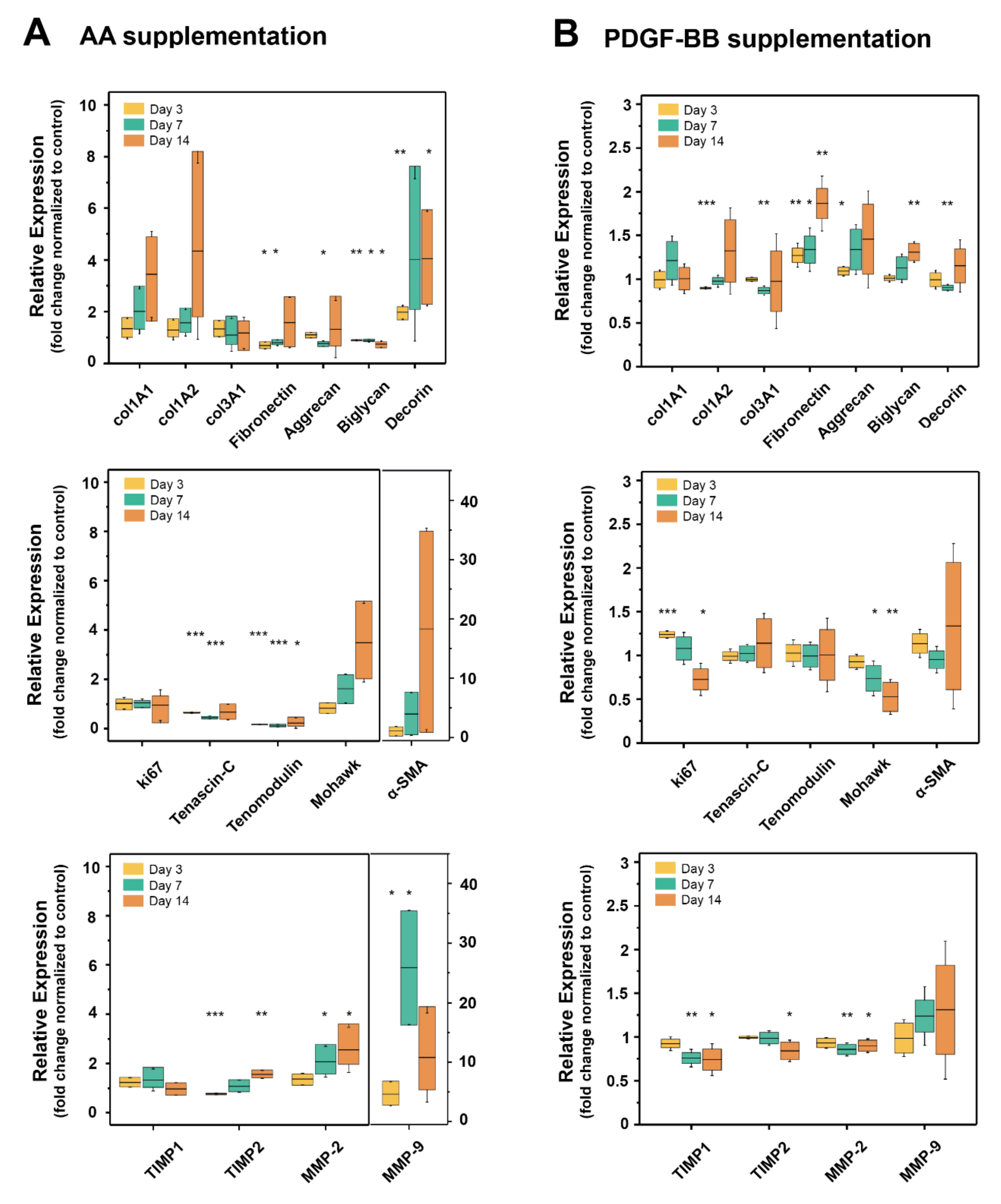
2.3. AA and PDGF-BB Supplementation Differently Affected Expression of Tendon Specific Markers and ECM Proteins
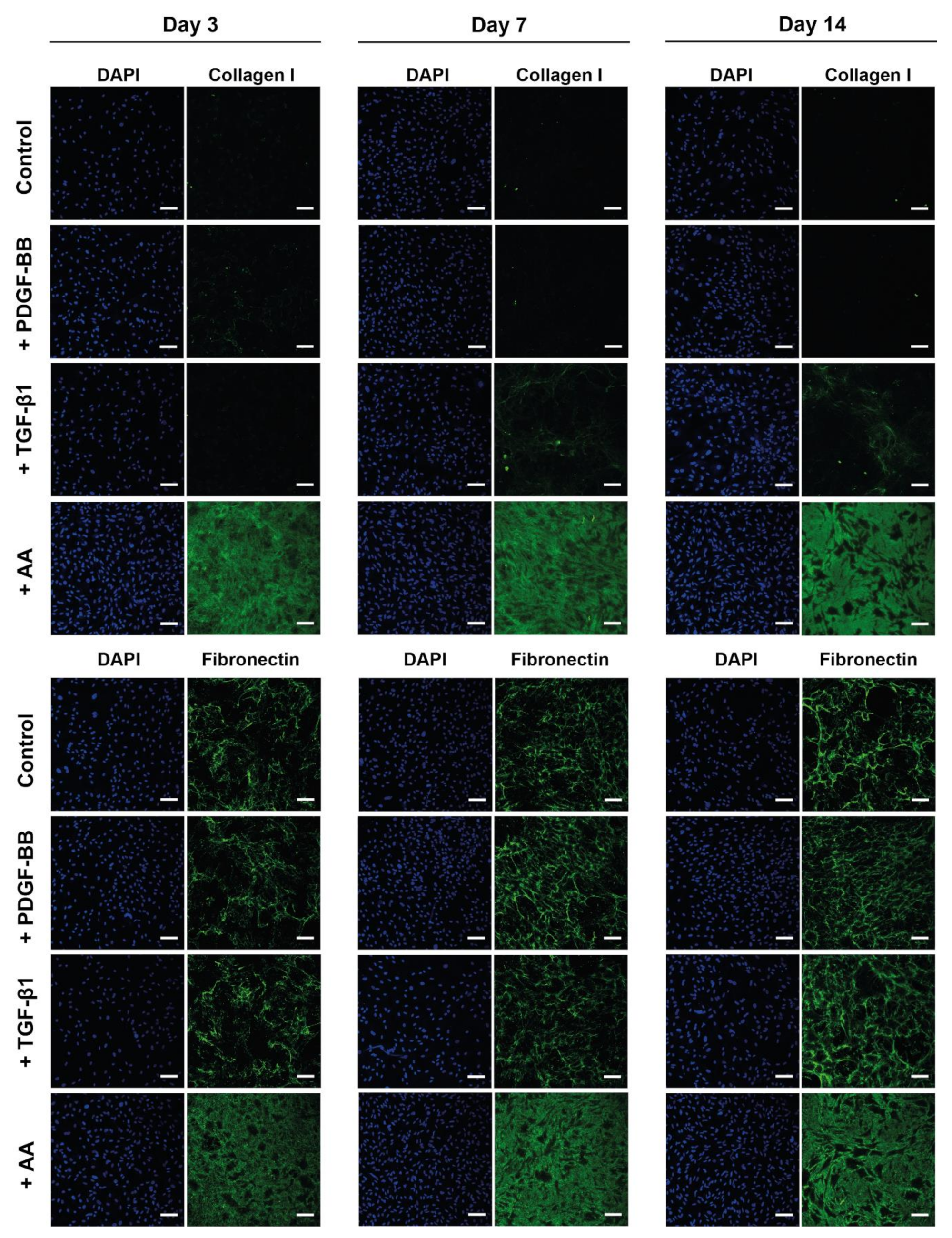
2.4. AA Supplementation Increased ECM Deposition by Tenocytes
3. Discussion
4. Materials and Methods
4.1. Isolation of Rabbit Achilles Tenocytes
4.2. EdU Proliferation Assay
4.3. Tenocyte Morphology under Different Conditions
4.4. RNA Isolation and Quantitative Real-Time PCR Analysis
4.5. Extracellular Matrix Production under Different Conditions
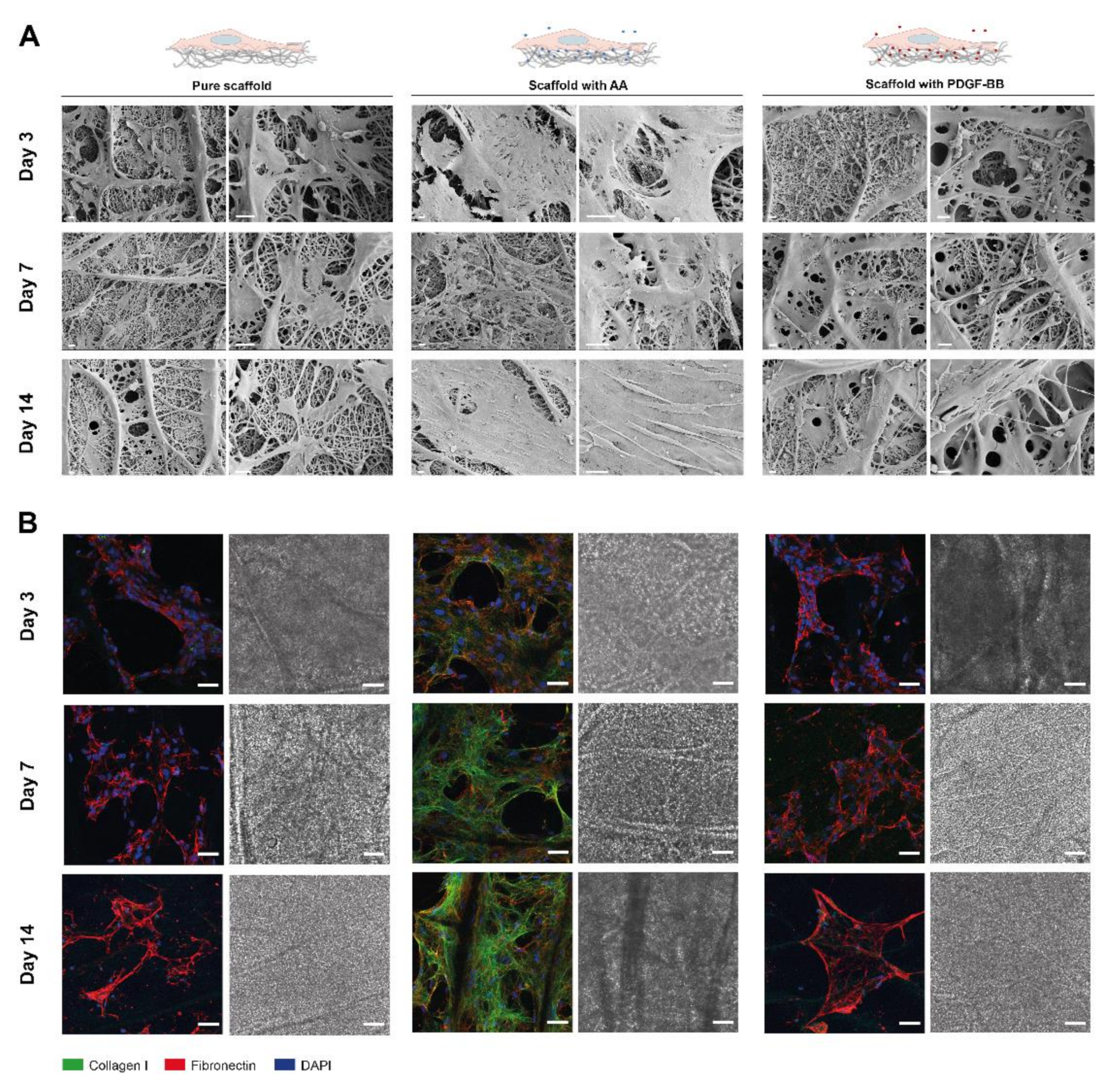
4.6. Tenocyte Culture on Electrospun Scaffolds with Incorporated AA or PDGF-BB
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Ascorbic acid |
| bFGF | Basic fibroblast growth factor |
| IGF | Insulin-like growth factor |
| MSCs | Mesenchymal stem cells |
| MMPs | Matrix metalloproteases |
| PDGF-BB | Platelet-derived growth factor-BB |
| TGF-β1 | Transforming growth factor-beta 1 |
| TIMPs | Tissue inhibitors of metalloproteases |
| VEGF | Vascular endothelial growth factor |
| DP | DegraPol® polymer |
References
- Kryger, G.S.; Chong, A.K.; Costa, M.; Pham, H.; Bates, S.J.; Chang, J. A comparison of tenocytes and mesenchymal stem cells for use in flexor tendon tissue engineering. J. Hand Surg. 2007, 32, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yin, Z.; Wu, T.; Li, Y.; Luo, X.; Xu, M.; Duan, L.; Li, J. Enhanced Effect of Tendon Stem/Progenitor Cells Combined With Tendon-Derived Decellularized Extracellular Matrix on Tendon Regeneration. Cell Transplant. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquette, C.; Slimani, S.; Kahn, C.J.; Tran, N.; Rahouadj, R.; Wang, X. A poly(lactic-co-glycolic acid) knitted scaffold for tendon tissue engineering: An in vitro and in vivo study. J. Biomater. Sci. Polym. Ed. 2010, 21, 1737–1760. [Google Scholar] [CrossRef] [PubMed]
- Kew, S.J.; Gwynne, J.H.; Enea, D.; Abu-Rub, M.; Pandit, A.; Zeugolis, D.; Brooks, R.A.; Rushton, N.; Best, S.M.; Cameron, R.E. Regeneration and repair of tendon and ligament tissue using collagen fibre biomaterials. Acta Biomater. 2011, 7, 3237–3247. [Google Scholar] [CrossRef]
- Gaspar, D.; Spanoudes, K.; Holladay, C.; Pandit, A.; Zeugolis, D. Progress in cell-based therapies for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 240–256. [Google Scholar] [CrossRef]
- Leong, D.J.; Sun, H.B. Mesenchymal stem cells in tendon repair and regeneration: Basic understanding and translational challenges. Ann. N. Y. Acad. Sci. 2016, 1383, 88–96. [Google Scholar] [CrossRef]
- Costa-Almeida, R.; Calejo, I.; Gomes, M.E. Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. Int. J. Mol. Sci. 2019, 20, 3002. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Luo, Q.; Deng, B.; Morita, Y.; Ju, Y.; Song, G. Construction of tendon replacement tissue based on collagen sponge and mesenchymal stem cells by coupled mechano-chemical induction and evaluation of its tendon repair abilities. Acta Biomater. 2018, 74, 247–259. [Google Scholar] [CrossRef]
- Orr, S.B.; Chainani, A.; Hippensteel, K.J.; Kishan, A.; Gilchrist, C.; Garrigues, N.W.; Ruch, D.S.; Guilak, F.; Little, D. Aligned multilayered electrospun scaffolds for rotator cuff tendon tissue engineering. Acta Biomater. 2015, 24, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yu, Q.; Wu, B.; Lin, Z.; Pavlos, N.J.; Xu, J.; Ouyang, H.; Wang, A.; Zheng, M.H. Autologous tenocyte therapy for experimental Achilles tendinopathy in a rabbit model. Tissue Eng. Part A 2011, 17, 2037–2048. [Google Scholar] [CrossRef]
- Pietschmann, M.F.; Frankewycz, B.; Schmitz, P.; Docheva, D.; Sievers, B.; Jansson, V.; Schieker, M.; Muller, P.E. Comparison of tenocytes and mesenchymal stem cells seeded on biodegradable scaffolds in a full-size tendon defect model. J. Mater. Sci. Mater. Med. 2013, 24, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Spanoudes, K.; Gaspar, D.; Pandit, A.; Zeugolis, D.I. The biophysical, biochemical, and biological toolbox for tenogenic phenotype maintenance in vitro. Trends Biotechnol. 2014, 32, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Breidahl, W.; Mackie, K.E.; Lin, Z.; Qin, A.; Chen, J.; Zheng, M.H. Autologous tenocyte injection for the treatment of severe, chronic resistant lateral epicondylitis: A pilot study. Am. J. Sports Med. 2013, 41, 2925–2932. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Mackie, K.; Breidahl, W.; Wang, T.; Zheng, M.H. Evidence for the Durability of Autologous Tenocyte Injection for Treatment of Chronic Resistant Lateral Epicondylitis: Mean 4.5-Year Clinical Follow-up. Am. J. Sports Med. 2015, 43, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Bucher, T.A.; Ebert, J.R.; Smith, A.; Breidahl, W.; Fallon, M.; Wang, T.; Zheng, M.-H.; Janes, G.C. Autologous Tenocyte Injection for the Treatment of Chronic Recalcitrant Gluteal Tendinopathy: A Prospective Pilot Study. Orthop. J. Sports Med. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Brandt, L.; Schubert, S.; Scheibe, P.; Brehm, W.; Franzen, J.; Gross, C.; Burk, J. Tenogenic Properties of Mesenchymal Progenitor Cells Are Compromised in an Inflammatory Environment. Int. J. Mol. Sci. 2018, 19, 2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perikamana, S.K.M.; Lee, J.; Ahmad, T.; Kim, E.M.; Byun, H.; Lee, S.; Shin, H. Harnessing biochemical and structural cues for tenogenic differentiation of adipose derived stem cells (ADSCs) and development of an in vitro tissue interface mimicking tendon-bone insertion graft. Biomaterials 2018, 165, 79–93. [Google Scholar] [CrossRef]
- Shen, H.; Gelberman, R.H.; Silva, M.J.; Sakiyama-Elbert, S.E.; Thomopoulos, S. BMP12 induces tenogenic differentiation of adipose-derived stromal cells. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Barsby, T.; Bavin, E.P.; Guest, D.J. Three-Dimensional Culture and Transforming Growth Factor Beta3 Synergistically Promote Tenogenic Differentiation of Equine Embryo-Derived Stem Cells. Tissue Eng. Part A 2014, 20, 2604–2613. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Bestwick, C.S.; Bestwick, L.A.; Maffulli, N.; Aspden, R.M. Phenotypic drift in human tenocyte culture. Tissue Eng. 2006, 12, 1843–1849. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar] [PubMed]
- Raimondi, M.T.; Lagana, M.; Conci, C.; Crestani, M.; Di Giancamillo, A.; Gervaso, F.; Deponti, D.; Boschetti, F.; Nava, M.M.; Scandone, C.; et al. Development and biological validation of a cyclic stretch culture system for the ex vivo engineering of tendons. Int. J. Artif. Organs 2018, 41, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Theiss, F.; Mirsaidi, A.; Mhanna, R.; Kummerle, J.; Glanz, S.; Bahrenberg, G.; Tiaden, A.N.; Richards, P.J. Use of biomimetic microtissue spheroids and specific growth factor supplementation to improve tenocyte differentiation and adaptation to a collagen-based scaffold in vitro. Biomaterials 2015, 69, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Djalali-Cuevas, A.; Garnica-Galvez, S.; Rampin, A.; Gaspar, D.; Skoufos, I.; Tzora, A.; Prassinos, N.; Diakakis, N.; Zeugolis, D.I. Preparation and Characterization of Tissue Surrogates Rich in Extracellular Matrix Using the Principles of Macromolecular Crowding. In The Extracellular Matrix: Methods and Protocols; Vigetti, D., Theocharis, A.D., Eds.; Springer: New York, NY, USA, 2019; pp. 245–259. [Google Scholar]
- Testa, S.; Costantini, M.; Fornetti, E.; Bernardini, S.; Trombetta, M.; Seliktar, D.; Cannata, S.; Rainer, A.; Gargioli, C. Combination of biochemical and mechanical cues for tendon tissue engineering. J. Cell. Mol. Med. 2017, 21, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Zhou, Z.P.; Taub, P.J.; Ramcharan, M.; Li, Y.H.; Akinbiyi, T.; Maharam, E.R.; Leong, D.J.; Laudier, D.M.; Ruike, T.; et al. BMP-12 Treatment of Adult Mesenchymal Stem Cells In Vitro Augments Tendon-Like Tissue Formation and Defect Repair In Vivo. PLoS ONE 2011, 6, 1–7. [Google Scholar] [CrossRef]
- Lee, C.; Jun, Y.; Kao, K. Harnessing Endogenous Stem/Progenitor Cells for Tendon Regeneration. Tissue Eng. Part A 2015, 21, S72. [Google Scholar] [CrossRef]
- Evrova, O.; Buschmann, J. In vitro and in vivo effects of PDGF-BB delivery strategies on tendon healing: A review. Eur. Cells Mater. 2017, 34, 15–39. [Google Scholar] [CrossRef]
- Chiu, C.-H.; Chen, P.; Chen, A.C.-Y.; Chan, Y.-S.; Hsu, K.-Y.; Rei, H.; Lei, K.F. Real-Time Monitoring of Ascorbic Acid-Mediated Reduction of Cytotoxic Effects of Analgesics and NSAIDs on Tenocytes Proliferation. Dose Response 2019, 17, 1–9. [Google Scholar] [CrossRef]
- Orfei, C.P.; Vigano, M.; Pearson, J.R.; Colombini, A.; De Luca, P.; Ragni, E.; Santos-Ruiz, L.; de Girolamo, L. In vitro induction of tendon-specific markers in tendon cells, adipose- and bone marrow-derived stem cells is dependent on TGF3, BMP-12 and ascorbic acid stimulation. Int. J. Mol. Sci. 2019, 20, 149. [Google Scholar] [CrossRef] [Green Version]
- Hakimi, O.; Poulson, R.; Thakkar, D.; Yapp, C.; Carr, A. Ascorbic acid is essential for significant collagen deposition by human tenocytes in vitro. Oxid. Antioxid. Med. Sci. 2014, 3, 119–127. [Google Scholar] [CrossRef] [Green Version]
- James, R.; Kumbar, S.G.; Laurencin, C.T.; Balian, G.; Chhabra, A.B. Tendon tissue engineering: Adipose-derived stem cell and GDF-5 mediated regeneration using electrospun matrix systems. Biomed. Mater. 2011, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Tao, X.; Chen, L.; Han, W.; Zhou, Y.; Tang, K. CTGF positively regulates BMP12 induced tenogenic differentiation of tendon stem cells and signaling. Cell Physiol. Biochem. 2015, 35, 1831–1845. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Guo, J.; Wu, T.Y.; Chen, X.; Xu, L.L.; Lin, S.E.; Sun, Y.X.; Chan, K.M.; Ouyang, H.; Li, G. Stepwise differentiation of mesenchymal stem cells augments tendon-like tissue formation and defect repair in vivo. Stem Cells Transl. Med. 2016, 5, 1106–1116. [Google Scholar] [CrossRef] [PubMed]
- Stanco, D.; Caprara, C.; Ciardelli, G.; Mariotta, L.; Gola, M.; Minonzio, G.; Soldati, G. Tenogenic differentiation protocol in xenogenic-free media enhances tendon-related marker expression in ASCs. PLoS ONE 2019, 14, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Falcon, N.D.; Riley, G.P.; Saeed, A. Induction of tendon-specific markers in adipose-derived stem cells in serum-free culture conditions. Tissue Eng. Part C Methods 2019, 25, 389–400. [Google Scholar] [CrossRef]
- Kraus, A.; Sattler, D.; Wehland, M.; Luetzenberg, R.; Abuagela, N.; Infanger, M. Vascular endothelial growth factor enhances proliferation of human tenocytes and promotes tenogenic gene expression. Plast. Reconstr. Surg. 2018, 142, 1240–1247. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Berardocco, M.; Gallorini, M.; Oliva, F.; Colosimo, A.; Cataldi, A.; Maffulli, N.; Berardi, A.C. Combined supplementation of ascorbic acid and thyroid hormone T3 affects tenocyte proliferation. The effect of ascorbic acid in the production of nitric oxide. Muscles Ligaments Tendons J. 2017, 7, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.A.; Wu, C.; Pham, B.V.; Chong, A.K.S.; Pham, H.M.; Chang, J. Tissue engineering of flexor tendons: Optimization of tenocyte proliferation using growth factor supplementation. Tissue Eng. 2006, 12, 1937–1943. [Google Scholar] [CrossRef]
- Huegle, T.; Majewski, M.; Valderrabano, V. Zell-und molekularbiologische Abläufe bei der Heilung von Sehnenverletzungen. Schweiz. Z. Med. Traumatol. 2012, 60, 49–51. [Google Scholar]
- Chang, J.; Tunder, R.; Most, D.; Longaker, M.T.; Lineaweaver, W.C. Studies in flexor tendon wound healing: Neutralizing antibody to TGF-beta1 increases postoperative range of motion. Plast. Reconstr. Surg. 2000, 105, 148–155. [Google Scholar] [CrossRef]
- Galatz, L.M.; Sandell, L.J.; Rothermich, S.Y.; Das, R.; Mastny, A.; Havlioglu, N.; Silva, M.J.; Thomopoulos, S. Characteristics of the rat supraspinatus tendon during tendon-to-bone healing after acute injury. J. Orthop. Res. 2006, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Penn, J.W.; Grobbelaar, A.O.; Rolfe, K.J. The role of the TGF-β family in wound healing, burns and scarring: A review. Int. J. Burn. Trauma 2012, 2, 18–28. [Google Scholar]
- Canty, E.G.; Kadler, K.E. Procollagen trafficking, processing and fibrillogenesis. J. Cell Sci. 2005, 118, 1341–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, C.J.; Bailey, A.J.; Prynne, C.J.; Levene, C.I. The effect of ascorbic acid on the synthesis of collagen precursor secreted by 3T6 mouse fibroblasts in culture. Biochim. Et. Biophys. Acta 1972, 278, 372–390. [Google Scholar] [CrossRef]
- Ömeroğlu, S.; Peker, T.; Türközkan, N.; Ömeroğlu, H.J.A.O.O.; Surgery, T. High-dose vitamin C supplementation accelerates the Achilles tendon healing in healthy rats. Arch. Orthop. Trauma Surg. 2009, 129, 281–286. [Google Scholar] [CrossRef]
- Evrova, O.; Houska, J.; Welti, M.; Bonavoglia, E.; Calcagni, M.; Giovanoli, P.; Vogel, V.; Buschmann, J. Bioactive, elastic, and biodegradable emulsion electrospun DegraPol tube delivering PDGF-BB for tendon rupture repair. Macromol. Biosci. 2016, 16, 1048–1063. [Google Scholar] [CrossRef]
- Jiang, Y.; Shi, Y.; He, J.; Zhang, Z.; Zhou, G.; Zhang, W.; Cao, Y.; Liu, W. Enhanced tenogenic differentiation and tendon-like tissue formation by tenomodulin overexpression in murine mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2017, 11, 2525–2536. [Google Scholar] [CrossRef]
- Docheva, D.; Hunziker, E.B.; Fassler, R.; Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell. Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Shukunami, C.; Yoshimoto, Y.; Takimoto, A.; Yamashita, H.; Hiraki, Y. Molecular characterization and function of tenomodulin, a marker of tendons and ligaments that integrate musculoskeletal components. Jpn. Dent. Sci. Rev. 2016, 52, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Toriuchi, N.; Yoshitaka, T.; Ueno-Kudoh, H.; Sato, T.; Yokoyama, S.; Nishida, K.; Akimoto, T.; Takahashi, M.; Miyaki, S.; et al. The mohawk homeobox gene is a critical regulator of tendon differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 10538–10542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Ito, Y.; Shinohara, M.; Yamashita, S.; Ichinose, S.; Kishida, A.; Oyaizu, T.; Kayama, T.; Nakamichi, R.; Koda, N.; et al. Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 7840–7845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milet, C.; Duprez, D. The Mkx homeoprotein promotes tenogenesis in stem cells and improves tendon repair. Ann. Transl. Med. 2015, 3, S33. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.A.; Sun, M.; Barnum, C.E.; Weiss, S.N.; Huegel, J.; Shetye, S.S.; Lin, L.; Saez, D.; Adams, S.M.; Iozzo, R.V.; et al. Decorin and biglycan are necessary for maintaining collagen fibril structure, fiber realignment, and mechanical properties of mature tendons. Matrix Biol. 2017, 64, 81–93. [Google Scholar] [CrossRef]
- Minkwitz, S.; Schmock, A.; Kurtoglu, A.; Tsitsilonis, S.; Manegold, S.; Wildemann, B.; Klatte-Schulz, F. Time-dependent alterations of MMPs, TIMPs and tendon structure in human achilles tendons after acute rupture. Int. J. Mol. Sci. 2017, 18, 2199. [Google Scholar] [CrossRef]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-β signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Ignotz, R.A.; Massague, J. Transforming growth factor-beta stimulates the expression of fibronectin and collagen and their incorporation into the extracellular matrix. J. Biol. Chem. 1986, 261, 4337–4345. [Google Scholar]
- Smith, M.L.; Gourdon, D.; Little, W.C.; Kubow, K.E.; Eguiluz, R.A.; Luna-Morris, S.; Vogel, V. Force-induced unfolding of fibronectin in the extracellular matrix of living cells. PLoS Biol. 2007, 5, e268. [Google Scholar] [CrossRef]
- Kang, H.J.; Kang, E.S. Ideal concentration of growth factors in rabbit’s flexor tendon culture. Yonsei. Med. J. 1999, 40, 26–29. [Google Scholar] [CrossRef]
- Raghavan, S.S.; Woon, C.Y.L.; Kraus, A.; Megerle, K.; Hung, P.; Chang, J. Optimization of human tendon tissue engineering: Synergistic effects of growth factors for use in tendon scaffold repopulation. Plast. Reconstr. Surg. 2012, 129, 479–489. [Google Scholar] [CrossRef]
- DePhillipo, N.N.; Aman, Z.S.; Kennedy, M.I.; Begley, J.P.; Moatshe, G.; LaPrade, R.F. Efficacy of vitamin c supplementation on collagen synthesis and oxidative stress after musculoskeletal injuries: A systematic review. Orthop. J. Sports Med. 2018, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, C.; Cermola, F.; Patriarca, E.J.; Minchiotti, G. Vitamin C in stem cell biology: Impact on extracellular matrix homeostasis and epigenetics. Stem Cells Int. 2017, 8936156, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herchenhan, A.; Uhlenbrock, F.; Eliasson, P.; Weis, M.; Eyre, D.; Kadler, K.E.; Magnusson, S.P.; Kjaer, M. Lysyl oxidase activity is required for ordered collagen fibrillogenesis by tendon cells. J. Biol. Chem. 2015, 290, 16440–16450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, F.; Maffulli, N.; Gissi, C.; Veronesi, F.; Calciano, L.; Fini, M.; Brogini, S.; Gallorini, M.; Passeri, C.A.L.; Bernardini, R.; et al. Combined ascorbic acid and T-3 produce better healing compared to bone marrow mesenchymal stem cells in an Achilles tendon injury rat model: A proof of concept study. J. Orthop. Surg. Res. 2019, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, W.-K.; Sung, Y.K.; Kwack, M.H.; Song, S.Y.; Choi, J.-S.; Park, S.G.; Yi, T.; Lee, H.-J.; Kim, D.-D.; et al. The molecular mechanism underlying the proliferating and preconditioning effect of vitamin C on adipose-derived stem cells. Stem Cells Dev. 2014, 23, 1364–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moteki, H.; Shimamura, Y.; Kimura, M.; Ogihara, M. Signal transduction pathway for L-ascorbic acid- and L-ascorbic acid 2-glucoside-induced DNA synthesis and cell proliferation in primary cultures of adult rat hepatocytes. Eur. J. Pharmacol. 2012, 683, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Banes, A.J.; Tsuzaki, M.; Hu, P.; Brigman, B.; Brown, T.; Almekinders, L.; Lawrence, W.T.; Fischer, T. PDGF-BB, IGF-I and mechanical load stimulate DNA synthesis in avian tendon fibroblasts in vitro. J. Biomech. 1995, 28, 1505–1513. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, X.; Zhang, Y.; Carr, A.J.; Zhu, L.; Xia, Z.; Sabokbar, A. Development of a refined tenocyte expansion culture technique for tendon tissue engineering. J. Tissue Eng. Regen. Med. 2014, 8, 955–962. [Google Scholar] [CrossRef]
- Best, K.T.; Loiselle, A.E. Scleraxis lineage cells contribute to organized bridging tissue during tendon healing and identify a subpopulation of resident tendon cells. FASEB J. 2019, 33, 8578–8587. [Google Scholar] [CrossRef]
- Piran, M.; Vakilian, S.; Piran, M.; Mohammadi-Sangcheshmeh, A.; Hosseinzadeh, S.; Ardeshirylajimi, A. In vitro fibroblast migration by sustained release of PDGF-BB loaded in chitosan nanoparticles incorporated in electrospun nanofibers for wound dressing applications. Artif. Cells Nanomed. Biotechnol. 2018, 46, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, C.; Zhu, S.; Lu, P.; Zhu, T.; Gong, X.; Zhang, Z.; Hu, J.; Yin, Z.; Heng, B.C.; et al. Mohawk promotes the tenogenesis of mesenchymal stem cells through activation of the tgf beta signaling pathway. Stem Cells 2015, 33, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Hill, A.; Canty-Laird, E.G. Collagen fibrillogenesis: Fibronectin, integrins, and minor collagens as organizers and nucleators. Curr. Opin. Cell Biol. 2008, 20, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Kubow, K.E.; Vukmirovic, R.; Zhe, L.; Klotzsch, E.; Smith, M.L.; Gourdon, D.; Luna, S.; Vogel, V. Mechanical forces regulate the interactions of fibronectin and collagen I in extracellular matrix. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sottile, J.; Shi, F.; Rublyevska, I.; Chiang, H.Y.; Lust, J.; Chandler, J. Fibronectin-dependent collagen I deposition modulates the cell response to fibronectin. Am. J. Physiol. Cell Physiol. 2007, 293, C1934–C1946. [Google Scholar] [CrossRef] [Green Version]
- Halper, J.; Kjaer, M. Basic Components of Connective Tissues and Extracellular Matrix: Elastin, Fibrillin, Fibulins, Fibrinogen, Fibronectin, Laminin, Tenascins and Thrombospondins. In Progress in Heritable Soft Connective Tissue Diseases; Halper, J., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 31–47. [Google Scholar]
- Midwood, K.S.; Orend, G. The role of tenascin-C in tissue injury and tumorigenesis. J. Cell Commun. Signal 2009, 3, 287–310. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.L.; Jones, F.S. Tenascin-C in development and disease: Gene regulation and cell function. Matrix Biol. 2000, 19, 581–596. [Google Scholar] [CrossRef]
- Wu, H.; Wu, Y.; Ai, Z.; Yang, L.; Gao, Y.; Du, J.; Guo, Z.; Zhang, Y. Vitamin C enhances nanog expression via activation of the jak/stat signaling pathway. Stem Cells. 2014, 32, 166–176. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, L.; Chen, L.; Wang, X.; Wu, H.; Ai, Z.; Du, J.; Liu, Y.; Shi, X.; Wu, Y.; et al. Vitamin C facilitates pluripotent stem cell maintenance by promoting pluripotency gene transcription. Biochimie 2013, 95, 2107–2113. [Google Scholar] [CrossRef]
- Mundre, R.S.; Koka, P.; Dhanaraj, P.; Khatri, N.; Vig, S.; Chandramohan, Y.; Dhanasekaran, A. Synergistic role of 5-azacytidine and ascorbic acid in directing cardiosphere derived cells to cardiomyocytes in vitro by downregulating Wnt signaling pathway via phosphorylation of β-catenin. PLoS ONE 2017, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Rharass, T.; Lantow, M.; Gbankoto, A.; Weiss, D.G.; Panáková, D.; Lucas, S. Ascorbic acid alters cell fate commitment of human neural progenitors in a WNT/β-catenin/ROS signaling dependent manner. J. Biomed. Sci. 2017, 24, 24–78. [Google Scholar] [CrossRef] [Green Version]
- Sha, H.; Zhang, D.Z.; Zhang, Y.F.; Wen, Y.H.; Wang, Y.C. ATF3 promotes migration and M1/M2 polarization of macrophages by activating tenascin-C via Wnt/beta-catenin pathway. Mol. Med. Rep. 2017, 16, 3641–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, H.J.; Park, D.W.; Seo, J.-Y.; Moon, J.-Y.; Kim, T.H.; Sohn, J.W.; Shin, D.H.; Yoon, H.J.; Park, S.S.; Kim, S.-H. The Wnt/β-catenin signaling pathway regulates the development of airway remodeling in patients with asthma. Exp. Mol. Med. 2015, 47, e198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popov, C.; Burggraf, M.; Kreja, L.; Ignatius, A.; Schieker, M.; Docheva, D. Mechanical stimulation of human tendon stem/progenitor cells results in upregulation of matrix proteins, integrins and MMPs, and activation of p38 and ERK1/2 kinases. BMC Mol. Biol. 2015, 16, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisman, E.; Lu, A.; Jamil, S.; Mousavizadeh, R.; McCormack, R.; Roberts, C.; Scott, A. Influence of repetitive mechanical loading on MMP2 activity in tendon fibroblasts. J. Orthop. Res. 2016, 34, 1991–2000. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Tanzil, G.; Al-Sadi, O.; Wiegand, E.; Ertel, W.; Busch, C.; Kohl, B.; Pufe, T. The role of pro-inflammatory and immunoregulatory cytokines in tendon healing and rupture: New insights. Scand. J. Med. Sci. Sports 2011, 21, 337–351. [Google Scholar] [CrossRef]
- Henriet, P.; Emonard, H. Matrix metalloproteinase-2: Not (just) a “hero” of the past. Biochimie 2019. [Google Scholar] [CrossRef]
- Karagiannis, E.D.; Popel, A.S. A theoretical model of type I collagen proteolysis by matrix metalloproteinase (MMP) 2 and membrane type 1 MMP in the presence of tissue inhibitor of metalloproteinase 2. J. Biol. Chem. 2004, 279, 39105–39114. [Google Scholar] [CrossRef] [Green Version]
- Deryugina, E.I.; Ratnikov, B.; Monosov, E.; Postnova, T.I.; DiScipio, R.; Smith, J.W.; Strongin, A.Y. MT1-MMP initiates activation of pro-MMP-2 and integrin alpha v beta 3 promotes maturation of MMP-2 in breast carcinoma cells. Exp. Cell Res. 2001, 263, 209–223. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evrova, O.; Kellenberger, D.; Calcagni, M.; Vogel, V.; Buschmann, J. Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes In Vitro. Int. J. Mol. Sci. 2020, 21, 458. https://doi.org/10.3390/ijms21020458
Evrova O, Kellenberger D, Calcagni M, Vogel V, Buschmann J. Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes In Vitro. International Journal of Molecular Sciences. 2020; 21(2):458. https://doi.org/10.3390/ijms21020458
Chicago/Turabian StyleEvrova, Olivera, Damian Kellenberger, Maurizio Calcagni, Viola Vogel, and Johanna Buschmann. 2020. "Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes In Vitro" International Journal of Molecular Sciences 21, no. 2: 458. https://doi.org/10.3390/ijms21020458