Functional Mechanisms Underlying the Antimicrobial Activity of the Oryza sativa Trx-like Protein

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
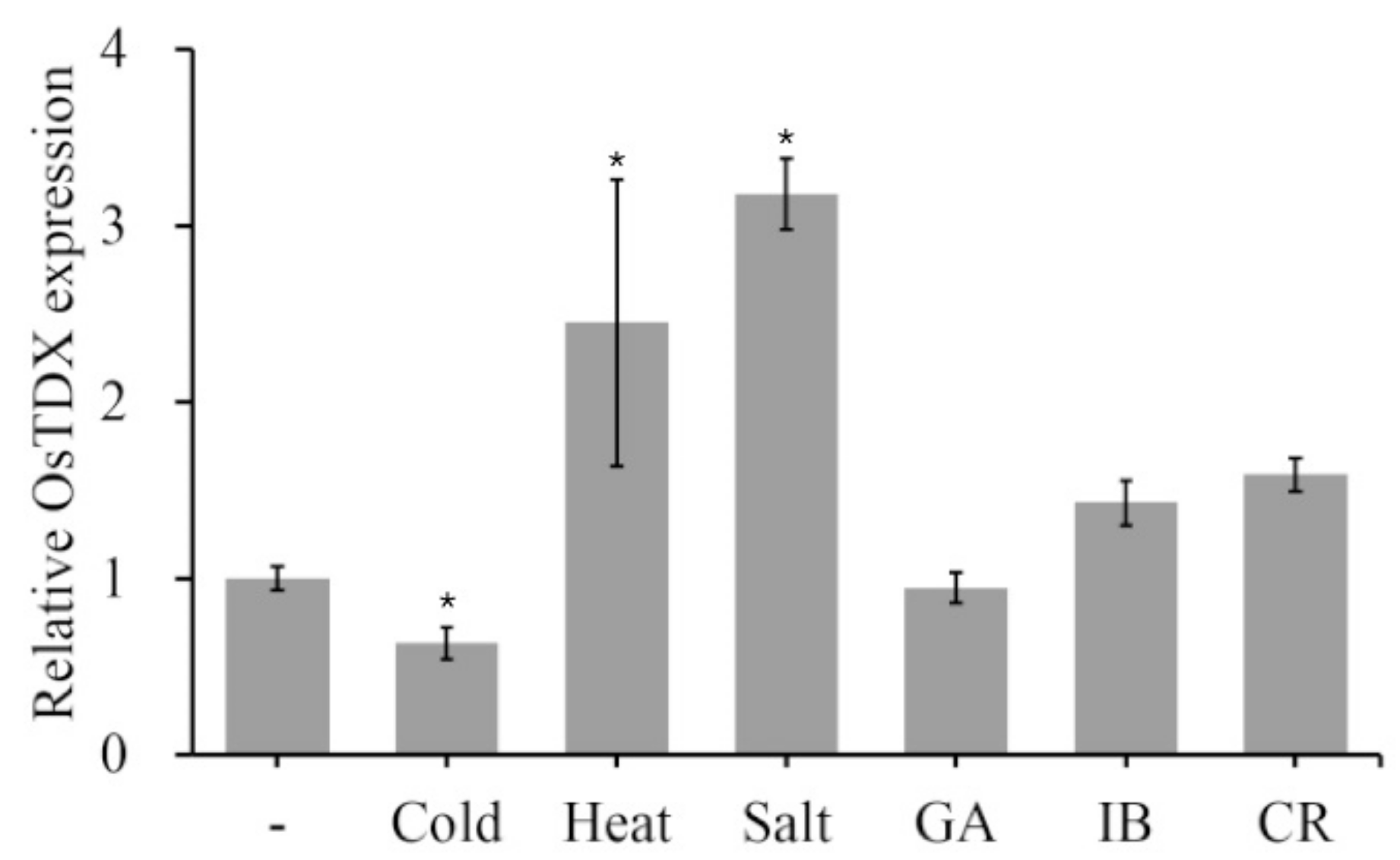
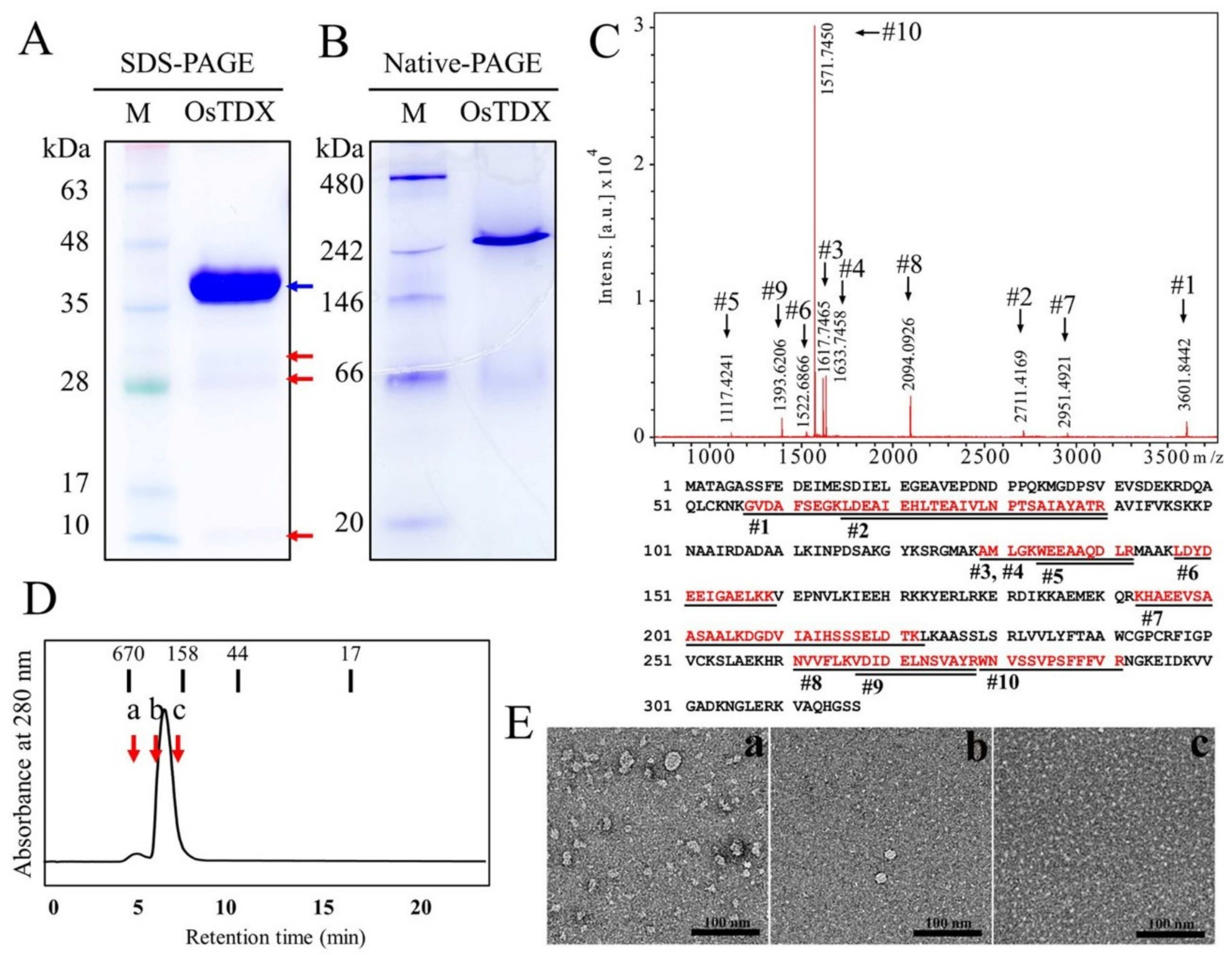
2.1. Isolation and Functional Characterization of Oryza sativa Tetratricopeptide Domain-Containing Thioredoxin (OsTDX) Protein
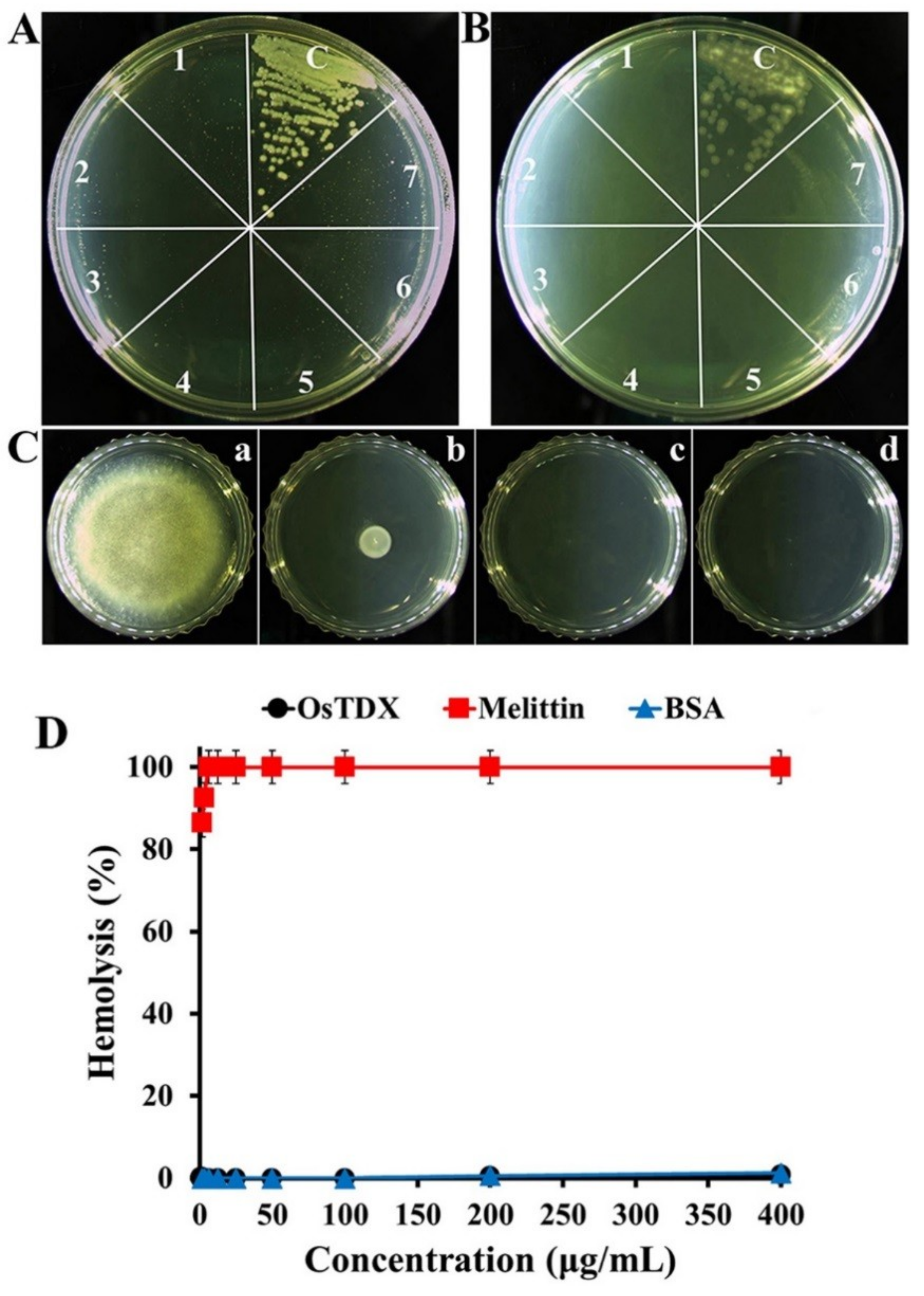
2.2. Antifungal and Cytotoxic Effects of Recombinant OsTDX
2.3. Molecular Mechanism of OsTDX in Fungal Cells
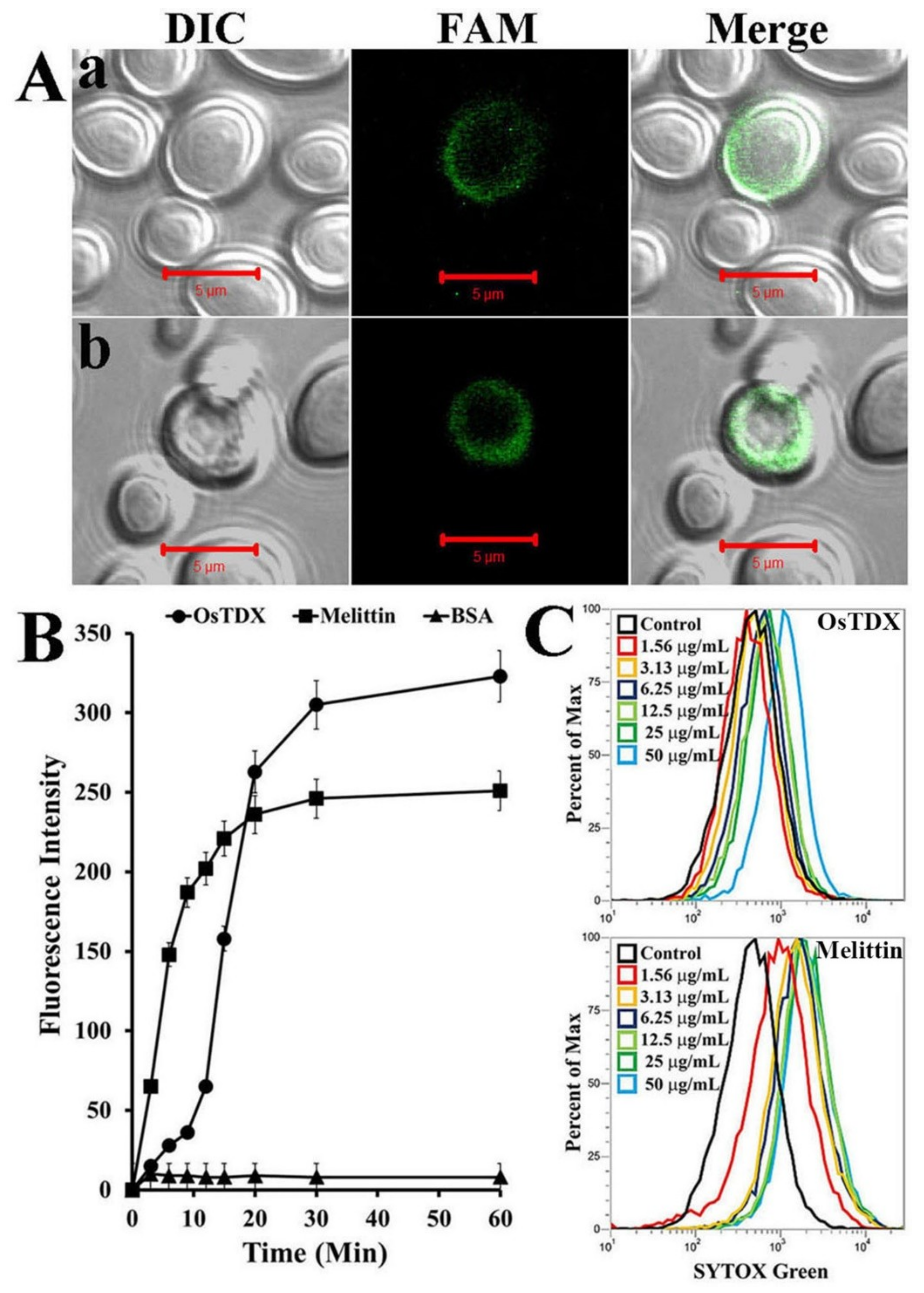
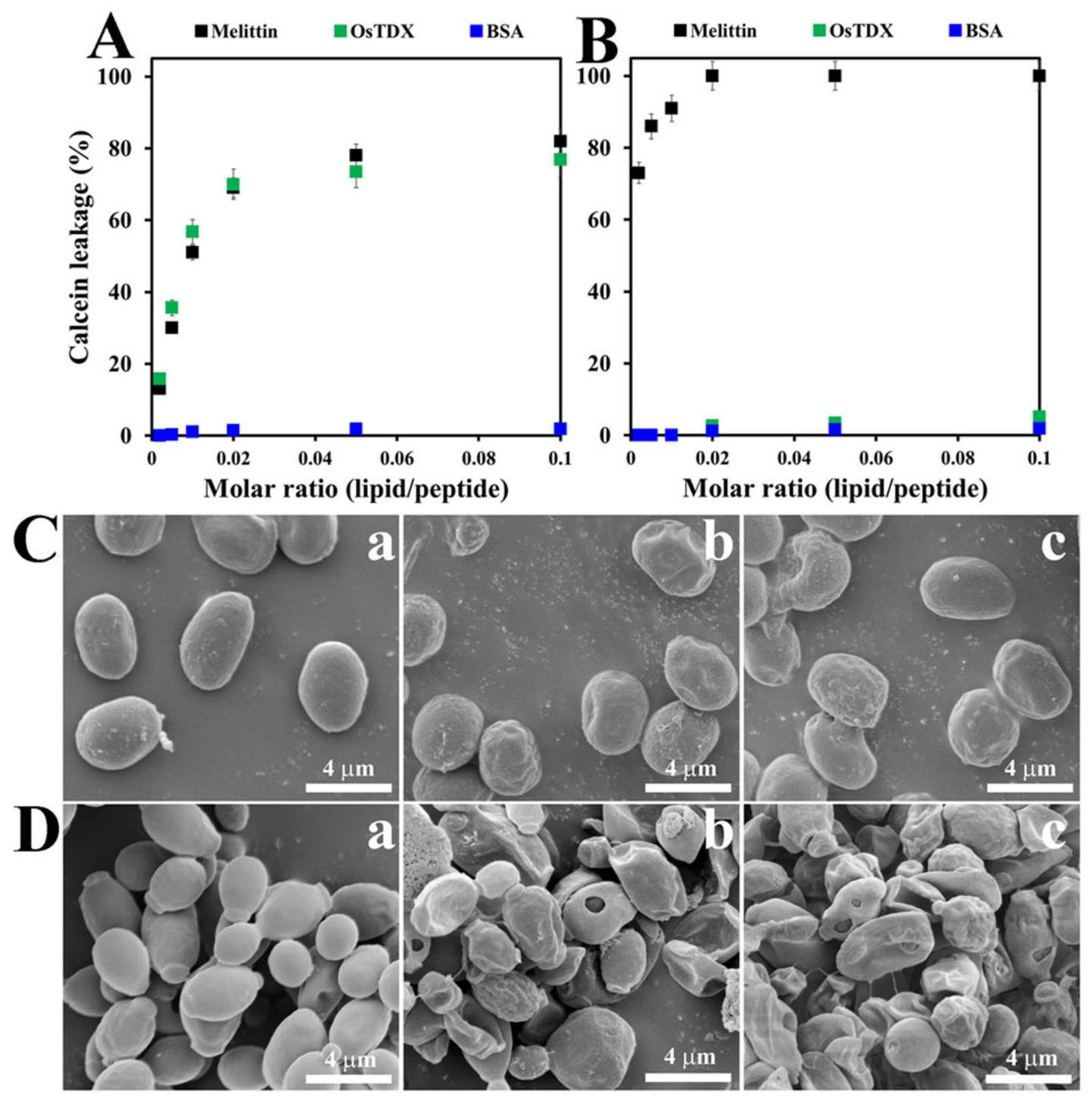
2.4. Effects of OsTDX on Membrane Permeability
2.5. Morphological Alterations Caused by OsTDX in Fungal Cells
3. Materials and Methods
3.1. Materials
3.2. Fungal Cells
3.3. RNA Isolation and qRT-PCR
3.4. Cloning of the OsTDX Gene and Protein Expression in E. coli
3.5. Purification and Structural Characterization of OsTDX Protein
3.6. Antifungal Assay
3.7. Hemolytic Effect of OsTDX in Rat Red Blood Cells (rRBCs)
3.8. Cellular Distribution of FAM-Labeled OsTDX
3.9. SYTOX-Green Uptake Assay
3.10. Calcein Leakage
3.11. Morphological Observation by Scanning Electron Microscopy (SEM)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Austin, M.J.; Muskett, P.; Kahn, K.; Feys, B.J.; Jones, J.D.; Parker, J.E. Regulatory role of SGT1 in early R gene-mediated plant defenses. Science 2002, 295, 2077–2080. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.H.; Chang, H.S.; Gupta, R.; Wang, X.; Zhu, T.; Luan, S. Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis. Plant Physiol. 2002, 129, 661–677. [Google Scholar] [CrossRef]
- Singh, K.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant-pathogen interactions. Curr. Opin. Plant Biol. 2005, 8, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Narusaka, Y.; Narusaka, M.; Seki, M.; Umezawa, T.; Ishida, J.; Nakajima, M.; Enju, A.; Shinozaki, K. Crosstalk in the responses to abiotic and biotic stresses in Arabidopsis: Analysis of gene expression in cytochrome P450 gene superfamily by cDNA microarray. Plant Mol. Biol. 2004, 55, 327–342. [Google Scholar] [CrossRef]
- Dooki, A.D.; Mayer-Posner, F.J.; Askari, H.; Zaiee, A.A.; Salekdeh, G.H. Proteomic responses of rice young panicles to salinity. Proteomics 2006, 6, 6498–6507. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress--Contribution of proteomics studies to understanding plant stress response. J. Proteomics 2011, 74, 1301–1322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Zhao, B.C.; Ge, W.N.; Zhang, Y.F.; Song, Y.; Sun, D.Y.; Guo, Y. An apoplastic h-type thioredoxin is involved in the stress response through regulation of the apoplastic reactive oxygen species in rice. Plant Physiol. 2011, 157, 1884–1899. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.G.; Chung, B.Y.; Kim, J.H.; Baek, M.H.; Yang, D.H.; Lee, J.W.; Kim, J.S. Ultrastructural changes of cell organelles in Arabidopsis stems after gamma irradation. J. Plant Biol. 2005, 48, 195–200. [Google Scholar] [CrossRef]
- Gicquel, M.; Esnault, M.A.; Jorrín-Novo, J.V.; Cabello-Hurtado, F. Application of proteomics to the assessment of the response to ionising radiation in Arabidopsis thaliana. J. Proteomic 2011, 74, 1364–1377. [Google Scholar] [CrossRef]
- Esnault, M.-A.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Potters, G.; Horemans, N.; Jansen, M.A. The cellular redox state in plant stress biology—A charging concept. Plant Physiol. Biochem. 2010, 48, 292–300. [Google Scholar] [CrossRef]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational changes of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef]
- Park, S.C.; Jung, Y.J.; Kim, I.R.; Lee, Y.; Kim, Y.M.; Jang, M.K.; Lee, J.R. Functional characterization of thioredoxin h type 5 with antimicrobial activity from Arabidopsis thaliana. Biotechnol. Bioprocess Eng. 2017, 22, 129–135. [Google Scholar] [CrossRef]
- Kim, S.G.; Chi, Y.H.; Lee, J.S.; Schlesinger, S.R.; Zabet-Moghaddam, M.; Chung, J.S.; Knaff, D.B.; Kim, S.T.; Lee, S.Y.; Kim, S.K. Redox properties of a thioredoxin-like Arabidopsis protein, AtTDX. Biochim. Biophys. Acta 2010, 1804, 2213–2221. [Google Scholar] [CrossRef]
- Xie, G.; Kato, H.; Sasaki, K.; Imai, R. A cold-induced thioredoxin h of rice, OsTrx23, negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro. FEBS Lett. 2009, 583, 2734–2738. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, J.Y.; Shin, S.O.; Jeong, C.Y.; Kim, M.H.; Shin, S.Y.; Cheong, G.W.; Park, Y.; Hahm, K.S. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 2006, 343, 222–228. [Google Scholar] [CrossRef]
- Mihajlovic, M.; Lazaridis, T. Antimicrobial peptides bind more strongly to membrane pores. Biochim. Biophys. Acta 2010, 1798, 1494–1502. [Google Scholar] [CrossRef]
- Erwig, L.P.; Gow, N.A. Interactions of fungal pathogens with phagocytes. Nat. Rev. Microbiol. 2016, 14, 163–176. [Google Scholar] [CrossRef]
- Caldana, C.; Scheible, W.R.; Mueller-Roeber, B.; Ruzicic, S. A quantitative RT-PCR platform for high-throughput expression profiling of 2500 rice transcription factors. Plant Methods 2007, 3, 7. [Google Scholar] [CrossRef]
- Lee, J.R.; Xie, X.; Yang, K.; Zhang, J.; Lee, S.Y.; Shippen, D.E. Dynamic interactions of Arabidopsis TEN1: Stabilizing telomeres in response to heat stress. Plant Cell 2016, 28, 2212–2224. [Google Scholar] [CrossRef]
- Park, S.-C.; Kim, I.R.; Kim, J.-Y.; Lee, Y.; Kim, E.-J.; Jung, J.H.; Jung, Y.J.; Jang, M.-K.; Lee, J.R. Molecular mechanism of Arabidopsis thaliana profilins as antifungal proteins. Biochim. Biophys. Acta (BBA) Gen. Subj. 2018, 1862, 2545–2554. [Google Scholar] [CrossRef]
- McLellan, T. Electrophoresis buffers for polyacrylamide gels at various pH. Anal. Biochem. 1982, 126, 94–99. [Google Scholar] [CrossRef]
- Moon, J.C.; Park, S.-C.; Shin, S.Y.; Jung, Y.J.; Jung, J.H.; Choi, W.; Jang, M.-K.; Lee, S.Y.; Lee, J.R. Deletion of the carboxyl terminal of thioredoxin reductase C of Arabidopsis facilitates oligomerization. Biotechnol. Bioprocess Eng. 2016, 21, 641–645. [Google Scholar] [CrossRef]
- Park, S.-C.; Kim, Y.-M.; Lee, J.-K.; Kim, N.-H.; Kim, E.-J.; Heo, H.; Lee, M.-Y.; Lee, J.R.; Jang, M.-K. Targeting and synergistic action of an antifungal peptide in an antibiotic drug-delivery system. J. Control. Release 2017, 256, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kim, J.Y.; Lee, J.K.; Hwang, I.; Cheong, H.; Nah, J.W.; Hahm, K.S.; Park, Y. Antifungal mechanism of a novel antifungal protein from pumpkin rinds against various fungal pathogens. J. Agric. Food Chem. 2009, 57, 9299–9304. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Lee, J.R.; Shin, S.O.; Park, Y.; Lee, S.Y.; Hahm, K.S. Characterization of a heat-stable protein with antimicrobial activity from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2007, 362, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Park, S.C.; Park, H.K.; Shin, S.Y.; Kim, Y.; Hahm, K.S. Structure-activity relationship of HP (2-20) analog peptide: Enhanced antimicrobial activity by N-terminal random coil region deletion. Biopolymers 2007, 88, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.D.; Morrell, J.L.; Hamer, J.E. Identification and characterization of Aspergillus nidulans mutants defective in cytokinesis. Genetics 1994, 136, 517–532. [Google Scholar] [PubMed]
- Gopal, R.; Park, S.C.; Ha, K.J.; Cho, S.J.; Kim, S.W.; Song, P.I.; Nah, J.W.; Park, Y.; Hahm, K.S. Effect of Leucine and Lysine substitution on the antimicrobial activity and evaluation of the mechanism of the HPA3NT3 analog peptide. J. Pept. Sci. 2009, 15, 589–594. [Google Scholar] [CrossRef]
- Fisher, D.K.; Higgins, T.J. A sensitive, high-volume, colorimetric assay for protein phosphatases. Pharm. Res. 1994, 11, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cleland, L.G. Serum-induced leakage of liposome contents. Biochim. Biophys. Acta 1980, 597, 418–426. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, M.H.; Hossain, M.A.; Shin, S.Y.; Kim, Y.; Stella, L.; Wade, J.D.; Park, Y.; Hahm, K.S. Amphipathic alpha-helical peptide, HP (2-20), and its analogues derived from Helicobacter pylori: Pore formation mechanism in various lipid compositions. Biochim. Biophys. Acta 2008, 1778, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kim, J.Y.; Kim, E.J.; Cheong, G.W.; Lee, Y.; Choi, W.; Lee, J.R.; Jang, M.K. Hydrophilic linear peptide with histidine and lysine residues as a key factor affecting antifungal activity. Int. J. Mol. Sci. 2018, 19, 3781. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | MIC (µg/mL) | MIC (µM) | ||
|---|---|---|---|---|
| OsTDX | Melittin | OsTDX | Melittin | |
| Mold | ||||
| C. gloeosporioides | 100 | 25 | 2.86 | 8.78 |
| F. graminearum | 50 | 25 | 1.43 | 8.78 |
| F. oxysporum | 50 | 50 | 1.43 | 17.57 |
| F. solani | 100 | 50 | 2.86 | 17.57 |
| G. zeae | 100 | 100 | 2.86 | 35.14 |
| Yeast | ||||
| C. albicans | 1.56 | 6.25 | 0.04 | 2.2 |
| C. krusei | 1.56 | 6.25 | 0.04 | 2.2 |
| C. parapsilosis | 6.25 | 12.5 | 0.18 | 4.39 |
| T. beigelii | 6.25 | 12.5 | 0.18 | 4.39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-C.; Kim, I.R.; Hwang, J.E.; Kim, J.-Y.; Jung, Y.J.; Choi, W.; Lee, Y.; Jang, M.-K.; Lee, J.R. Functional Mechanisms Underlying the Antimicrobial Activity of the Oryza sativa Trx-like Protein. Int. J. Mol. Sci. 2019, 20, 1413. https://doi.org/10.3390/ijms20061413
Park S-C, Kim IR, Hwang JE, Kim J-Y, Jung YJ, Choi W, Lee Y, Jang M-K, Lee JR. Functional Mechanisms Underlying the Antimicrobial Activity of the Oryza sativa Trx-like Protein. International Journal of Molecular Sciences. 2019; 20(6):1413. https://doi.org/10.3390/ijms20061413
Chicago/Turabian StylePark, Seong-Cheol, Il Ryong Kim, Jung Eun Hwang, Jin-Young Kim, Young Jun Jung, Wonkyun Choi, Yongjae Lee, Mi-Kyeong Jang, and Jung Ro Lee. 2019. "Functional Mechanisms Underlying the Antimicrobial Activity of the Oryza sativa Trx-like Protein" International Journal of Molecular Sciences 20, no. 6: 1413. https://doi.org/10.3390/ijms20061413