Robust Identification of Suitable T-Cell Subsets for Personalized CMV-Specific T-Cell Immunotherapy Using CD45RA and CD62L Microbeads

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Cell Selection Strategy for Naive T-Cell Depletion Using CD45RA and CD62L Microbeads
2.2. Multiparametric Evaluation of Cell Fractions Following Naive T-Cell Depletion with CD45RA and CD62L Microbeads
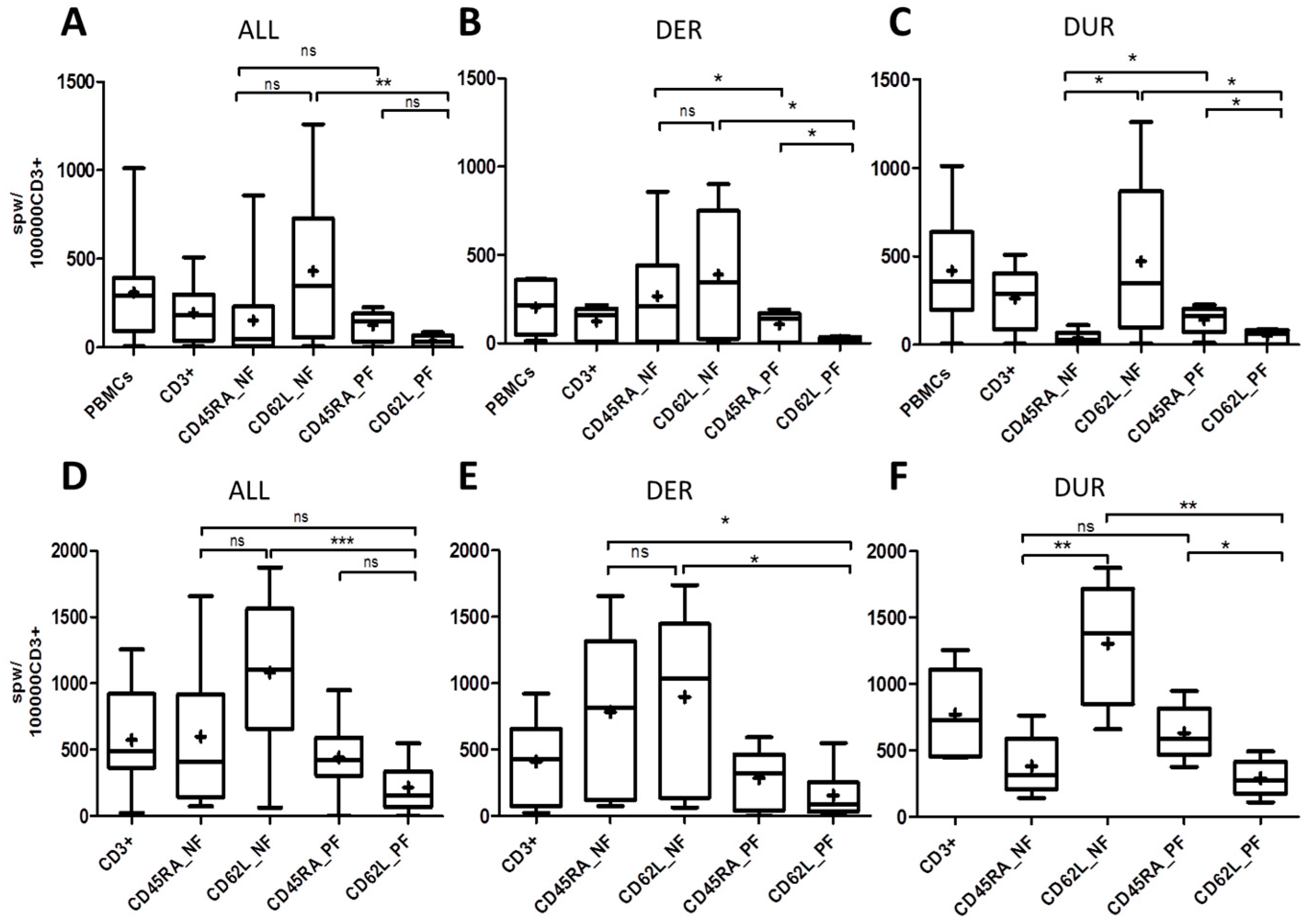
2.3. T-Cell Response to ppCMV_pp65 by IFN-γ Enzyme-Linked ImmunoSpot (ELISpot) Assay
2.4. Correlations between T-Cell Frequencies, Phenotypes and Auxiliary T Cells among Donors with Expected and Unexpected Response
2.5. Confirmation of CMV-Specific T-Cell Responses by Intracellular IFN-γ and Tumor Necrosis Factor-α (TNF-α) Cytokine Staining
3. Discussion
3.1. Dynamics of Sequential Cellular Composition within the Memory and Naive T-Cell Fractions
3.2. Dynamics of Sequential Cellular Composition within the Memory and Naive T-Cell Fractions
3.3. Dichotomy of CD45RA and CD62L Naive T-Cell Depletion Characteristics
3.4. Paradigm Shift in the Selection of T-Cell Products in Clinical Practice
4. Materials and Methods
4.1. Isolation of PBMCs and T Cells
4.2. Flow Cytometry Analysis
4.3. CMV-Specific T-Cell Response Determined by IFN-γ ELISpot
4.4. Detection of IFN-γ and TNF-α by Intracellular Cytokine Staining
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DER | Donors with expected response |
| DUR | Donors with unexpected response |
| CCS | Cytokine capture system |
| CMV | Cytomegalovirus |
| EBV | Epstein–Barr virus |
| GCSF | Granulocyte-colony stimulating factor |
| GvHD | Graft-versus-host disease |
| HSCT | Hematopoietic stem-cell transplantation |
| ICS | Intracellular cytokine staining |
| VST | Virus-specific T cell |
| NF | Negative fraction |
| PF | Positive fraction |
| Treg | Regulatory T cell |
References
- Papadopoulou, A.; Gerdemann, U.; Katari, U.L.; Tzannou, I.; Liu, H.; Martinez, C.; Leung, K.; Carrum, G.; Gee, A.P.; Vera, J.F.; et al. Activity of broad-spectrum T cells as treatment for AdV, EBV, CMV, BKV, and HHV6 infections after HSCT. Sci. Transl. Med. 2014, 6, 242ra83. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Cooper, M.L.; Staser, K.; Ashami, K.; Vij, K.R.; Wang, B.; Marsala, L.; Niswonger, J.; Ritchey, J.; Alahmari, B.; et al. Baricitinib-induced blockade of interferon gamma receptor and interleukin-6 receptor for the prevention and treatment of graft-versus-host disease. Leukemia 2018, 32, 2483–2494. [Google Scholar] [CrossRef]
- Papanicolaou, G.A.; Lee, Y.J.; Young, J.W.; Seshan, S.V.; Boruchov, A.M.; Chittick, G.; Mommeja-Marin, H.; Glezerman, I.G. Brincidofovir for polyomavirus-associated nephropathy after allogeneic hematopoietic stem cell transplantation. Am. J. Kidney Dis. 2015, 65, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Cheng, M.P. Antiviral prophylaxis for cytomegalovirus infection in allogeneic hematopoietic cell transplantation. Blood Adv. 2018, 2, 2159–2175. [Google Scholar] [CrossRef] [PubMed]
- Maffini, E.; Giaccone, L.; Festuccia, M.; Brunello, L.; Busca, A.; Bruno, B. Treatment of CMV infection after allogeneic hematopoietic stem cell transplantation. Expert Rev. Hematol. 2016, 9, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.; Satterwhite, L.E.; Ercolani, R.J. New Locus of Drug Resistance in the Human Cytomegalovirus UL56 Gene Revealed by In Vitro Exposure to Letermovir and Ganciclovir. Antimicrob Agents Chemother. 2018, 62, e00922-18. [Google Scholar] [CrossRef] [PubMed]
- Goldner, T.; Hewlett, G.; Ettischer, N.; Ruebsamen-Schaeff, H.; Zimmermann, H.; Lischka, P. The novel anticytomegalovirus compound AIC246 (Letermovir) inhibits human cytomegalovirus replication through a specific antiviral mechanism that involves the viral terminase. J. Virol. 2011, 85, 10884–10893. [Google Scholar] [CrossRef] [PubMed]
- Tzannou, I.; Papadopoulou, A.; Naik, S.; Leung, K.; Martinez, C.A.; Ramos, C.A.; Carrum, G.; Sasa, G.; Lulla, P.; Watanabe, A.; et al. Off-the-Shelf Virus-Specific T Cells to Treat BK Virus, Human Herpesvirus 6, Cytomegalovirus, Epstein-Barr Virus, and Adenovirus Infections After Allogeneic Hematopoietic Stem-Cell Transplantation. J. Clin. Oncol 2017, 35, 3547–3557. [Google Scholar] [CrossRef] [PubMed]
- Heslop, H.E.; Leen, A.M. T-cell therapy for viral infections. Hematol. Am. Soc. Hematol. Educ. Program. 2013, 2013, 342–347. [Google Scholar] [CrossRef]
- Frey, N.V.; Porter, D.L. Graft-versus-host disease after donor leukocyte infusions: Presentation and management. Best Pract Res. Clin. Haematol. 2008, 21, 205–222. [Google Scholar] [CrossRef]
- de Lima, M.; Bonamino, M.; Vasconcelos, Z.; Colares, M.; Diamond, H.; Zalcberg, I.; Tavares, R.; Lerner, D.; Byington, R.; Bouzas, L.; et al. Prophylactic donor lymphocyte infusions after moderately ablative chemotherapy and stem cell transplantation for hematological malignancies: High remission rate among poor prognosis patients at the expense of graft-versus-host disease. Bone Marrow Transplant. 2001, 27, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Feuchtinger, T.; Opherk, K.; Bethge, W.A.; Topp, M.S.; Schuster, F.R.; Weissinger, E.M.; Mohty, M.; Or, R.; Maschan, M.; Schumm, M.; et al. Adoptive transfer of pp65-specific T cells for the treatment of chemorefractory cytomegalovirus disease or reactivation after haploidentical and matched unrelated stem cell transplantation. Blood 2010, 116, 4360–4367. [Google Scholar] [CrossRef] [PubMed]
- Tischer, S.; Priesner, C.; Heuft, H.-G.; Goudeva, L.; Mende, W.; Barthold, M.; Kloeß, S.; Arseniev, L.; Aleksandrova, K.; Maecker-Kolhoff, B.; et al. Rapid generation of clinical-grade antiviral T cells: Selection of suitable T-cell donors and GMP-compliant manufacturing of antiviral T cells. J. Transl. Med. 2014, 12, 336. [Google Scholar] [CrossRef]
- Priesner, C.; Esser, R.; Tischer, S.; Marburger, M.; Aleksandrova, K.; Maecker-Kolhoff, B.; Heuft, H.G.; Goudeva, L.; Blasczyk, R.; Arseniev, L.; et al. Comparative Analysis of Clinical-Scale IFN-gamma-Positive T-Cell Enrichment Using Partially and Fully Integrated Platforms. Front. Immunol. 2016, 7, 393. [Google Scholar] [CrossRef]
- Casalegno-Garduno, R.; Schmitt, A.; Yao, J.; Wang, X.; Xu, X.; Freund, M.; Schmitt, M. Multimer technologies for detection and adoptive transfer of antigen-specific T cells. Cancer Immunol. Immunother. 2010, 59, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Knabel, M.; Franz, T.J.; Schiemann, M.; Wulf, A.; Villmow, B.; Schmidt, B.; Bernhard, H.; Wagner, H.; Busch, D.H. Reversible MHC multimer staining for functional isolation of T-cell populations and effective adoptive transfer. Nat. Med. 2002, 8, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Neuenhahn, M.; Albrecht, J.; Odendahl, M.; Schlott, F.; Dossinger, G.; Schiemann, M.; Lakshmipathi, S.; Martin, K.; Bunjes, D.; Harsdorf, S.; et al. Transfer of minimally manipulated CMV-specific T cells from stem cell or third-party donors to treat CMV infection after allo-HSCT. Leukemia 2017, 31, 2161–2171. [Google Scholar] [CrossRef]
- Gerdemann, U.; Vera, J.F.; Rooney, C.M.; Leen, A.M. Generation of multivirus-specific T cells to prevent/treat viral infections after allogeneic hematopoietic stem cell transplant. J. Vis. Exp. 2011. [Google Scholar] [CrossRef]
- Geyeregger, R.; Freimuller, C.; Stemberger, J.; Artwohl, M.; Witt, V.; Lion, T.; Fischer, G.; Lawitschka, A.; Ritter, J.; Hummel, M.; et al. First-in-man clinical results with good manufacturing practice (GMP)-compliant polypeptide-expanded adenovirus-specific T cells after haploidentical hematopoietic stem cell transplantation. J. Immunother. 2014, 37, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Saglio, F.; Hanley, P.J.; Bollard, C.M. The time is now: Moving toward virus-specific T cells after allogeneic hematopoietic stem cell transplantation as the standard of care. Cytotherapy 2014, 16, 149–159. [Google Scholar] [CrossRef]
- Fuji, S.; Löffler, J.; Einsele, H.; Kapp, M. Immunotherapy for opportunistic infections: Current status and future perspectives. Virulence 2016, 7, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Fuji, S.; Kapp, M.; Grigoleit, G.U.; Einsele, H. Adoptive immunotherapy with virus-specific T cells. Best Pract. Res. Clin. Haematol. 2011, 24, 413–419. [Google Scholar] [CrossRef]
- Lindemann, M.; Eiz-Vesper, B.; Steckel, N.K.; Tischer, S.; Fiedler, M.; Heinold, A.; Klisanin, V.; Maecker-Kolhoff, B.; Blasczyk, R.; Horn, P.A.; et al. Adoptive transfer of cellular immunity against cytomegalovirus by virus-specific lymphocytes from a third-party family donor. Bone Marrow Transplant. 2018, 53, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Leen, A.; Meij, P.; Redchenko, I.; Middeldorp, J.; Bloemena, E.; Rickinson, A.; Blake, N. Differential immunogenicity of Epstein-Barr virus latent-cycle proteins for human CD4(+) T-helper 1 responses. J. Virol. 2001, 75, 8649–8659. [Google Scholar] [CrossRef] [PubMed]
- Distler, E.; Bloetz, A.; Albrecht, J.; Asdufan, S.; Hohberger, A.; Frey, M.; Schnurer, E.; Thomas, S.; Theobald, M.; Hartwig, U.F.; et al. Alloreactive and leukemia-reactive T cells are preferentially derived from naive precursors in healthy donors: Implications for immunotherapy with memory T cells. Haematologica 2011, 96, 1024–1032. [Google Scholar] [CrossRef]
- Triplett, B.M.; Muller, B.; Kang, G.; Li, Y.; Cross, S.J.; Moen, J.; Cunningham, L.; Janssen, W.; Mamcarz, E.; Shook, D.R.; et al. Selective T-cell depletion targeting CD45RA reduces viremia and enhances early T-cell recovery compared with CD3-targeted T-cell depletion. Transpl. Infect. Dis. 2018, 20, e12823. [Google Scholar] [CrossRef]
- Bleakley, M.; Heimfeld, S.; Jones, L.A.; Turtle, C.; Krause, D.; Riddell, S.R.; Shlomchik, W. Engineering human peripheral blood stem cell grafts that are depleted of naive T cells and retain functional pathogen-specific memory T cells. Biol. Blood Marrow Transplant. 2014, 20, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Touzot, F.; Neven, B.; Dal-Cortivo, L.; Gabrion, A.; Moshous, D.; Cros, G.; Chomton, M.; Luby, J.M.; Terniaux, B.; Magalon, J.; et al. CD45RA depletion in HLA-mismatched allogeneic hematopoietic stem cell transplantation for primary combined immunodeficiency: A preliminary study. J. Allergy Clin. Immunol. 2015, 135, 1303–1309.e3. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, W.A.; Ramanathan, M. Review on Haploidentical Hematopoietic Cell Transplantation in Patients with Hematologic Malignancies. Adv. Hematol. 2016, 2016, 5726132. [Google Scholar] [CrossRef] [PubMed]
- Teschner, D.; Distler, E.; Wehler, D.; Frey, M.; Marandiuc, D.; Langeveld, K.; Theobald, M.; Thomas, S.; Herr, W. Depletion of naive T cells using clinical grade magnetic CD45RA beads: A new approach for GVHD prophylaxis. Bone Marrow Transplant. 2014, 49, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Verfuerth, S.; Sousa, P.S.; Beloki, L.; Murray, M. Generation of memory T cells for adoptive transfer using clinical-grade anti-CD62L magnetic beads. Bone Marrow Transplant. 2015, 50, 1358–1364. [Google Scholar] [CrossRef]
- Nikolich-Zugich, J.; Slifka, M.K.; Messaoudi, I. The many important facets of T-cell repertoire diversity. Nat. Rev. Immunol. 2004, 4, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Inman, C.F.; Eldershaw, S.A.; Croudace, J.E.; Davies, N.J.; Sharma-Oates, A.; Rai, T.; Pearce, H.; Sirovica, M.; Chan, Y.L.T.; Verma, K.; et al. Unique features and clinical importance of acute alloreactive immune responses. JCI Insight 2018, 3, e97219. [Google Scholar] [CrossRef] [PubMed]
- Okada, R.; Kondo, T.; Matsuki, F.; Takata, H.; Takiguchi, M. Phenotypic classification of human CD4+ T cell subsets and their differentiation. Int Immunol. 2008, 20, 1189–1199. [Google Scholar] [CrossRef]
- Appay, V.; van Lier, R.A.; Sallusto, F.; Roederer, M. Phenotype and function of human T lymphocyte subsets: Consensus and issues. Cytometry A 2008, 73, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.; Price, D.A.; Papagno, L.; Arkoub, Z.A.; Sauce, D.; Bornstein, E.; Asher, T.E.; Samri, A.; Schnuriger, A.; Theodorou, I.; et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J. Exp. Med. 2007, 204, 2473–2485. [Google Scholar] [CrossRef] [PubMed]
- Sacre, K.; Carcelain, G.; Cassoux, N.; Fillet, A.-M.; Costagliola, D.; Vittecoq, D.; Salmon, D.; Amoura, Z.; Katlama, C.; Autran, B. Repertoire, diversity, and differentiation of specific CD8 T cells are associated with immune protection against human cytomegalovirus disease. J. Exp. Med. 2005, 201, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Appay, V.; Dunbar, P.R.; Callan, M.; Klenerman, P.; Gillespie, G.M.; Papagno, L.; Ogg, G.S.; King, A.; Lechner, F.; Spina, C.A.; et al. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat. Med. 2002, 8, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.; Lugli, E. The who’s who of T-cell differentiation: Human memory T-cell subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef] [PubMed]
- Feuchtinger, T.; Lang, P.; Hamprecht, K.; Schumm, M.; Greil, J.; Jahn, G.; Niethammer, D.; Einsele, H. Isolation and expansion of human adenovirus-specific CD4+ and CD8+ T cells according to IFN-gamma secretion for adjuvant immunotherapy. Exp. Hematol. 2004, 32, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Babor, M.; Lane, J.; Schulten, V.; Patil, V.S.; Seumois, G. Unique phenotypes and clonal expansions of human CD4 effector memory T cells re-expressing CD45RA. Nat. Commun. 2017, 8, 1473. [Google Scholar] [CrossRef] [PubMed]
- Crough, T.; Khanna, R. Immunobiology of human cytomegalovirus: From bench to bedside. Clin. Microbiol. Rev. 2009, 22, 76–98. [Google Scholar] [CrossRef] [PubMed]
- Ljungman, P.; Hakki, M.; Boeckh, M. Cytomegalovirus in hematopoietic stem cell transplant recipients. Hematol. Oncol. Clin. N. Am. 2011, 25, 151–169. [Google Scholar] [CrossRef]
- Hebart, H.; Einsele, H. Clinical aspects of CMV infection after stem cell transplantation. Hum. Immunol. 2004, 65, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Teira, P.; Battiwalla, M.; Ramanathan, M.; Barrett, A.J.; Ahn, K.W.; Chen, M.; Green, J.S.; Saad, A.; Antin, J.H.; Savani, B.N.; et al. Early cytomegalovirus reactivation remains associated with increased transplant-related mortality in the current era: A CIBMTR analysis. Blood 2016, 127, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Quinn, M.; Snyder, C.M. CMV-Specific CD8 T Cell Differentiation and Localization: Implications for Adoptive Therapies. Front. Immunol. 2016, 7, 352. [Google Scholar] [CrossRef] [PubMed]
- Khairallah, C.; Déchanet-Merville, J.; Capone, M. γδ T Cell-Mediated Immunity to Cytomegalovirus Infection. Front. Immunol. 2017, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Marusina, A.I.; Ono, Y.; Merleev, A.A.; Shimoda, M.; Ogawa, H.; Wang, E.A.; Kondo, K.; Olney, L.; Luxardi, G.; Miyamura, Y.; et al. CD4(+) virtual memory: Antigen-inexperienced T cells reside in the naive, regulatory, and memory T cell compartments at similar frequencies, implications for autoimmunity. J. Autoimmun. 2017, 77, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Beres, A.J.; Drobyski, W.R. The role of regulatory T cells in the biology of graft versus host disease. Front. Immunol. 2013, 4, 163. [Google Scholar] [CrossRef]
- Muller, N.; Landwehr, K.; Langeveld, K.; Stenzel, J.; Pouwels, W.; van der Hoorn, M.; Seifried, E.; Bonig, H. Generation of alloreactivity-reduced donor lymphocyte products retaining memory function by fully automatic depletion of CD45RA-positive cells. Cytotherapy 2018, 20, 532–542. [Google Scholar] [CrossRef]
- Antoine, P.; Olislagers, V.; Huygens, A.; Lecomte, S.; Liesnard, C.; Donner, C.; Marchant, A. Functional exhaustion of CD4+ T lymphocytes during primary cytomegalovirus infection. J. Immunol. 2012, 189, 2665–2672. [Google Scholar] [CrossRef]
- Klenerman, P.; Oxenius, A. T cell responses to cytomegalovirus. Nat. Rev. Immunol. 2016, 16, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Shariff, N.; Cobbold, M.; Bruton, R.; Ainsworth, J.A.; Sinclair, A.J.; Nayak, L.; Moss, P.A. Cytomegalovirus seropositivity drives the CD8 T cell repertoire toward greater clonality in healthy elderly individuals. J. Immunol. 2002, 169, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.M.; Vukmanovic-Stejic, M.; Dunne, P.J.; Birch, K.E.; Cook, J.E.; Jackson, S.E.; Salmon, M.; Rustin, M.H.; Akbar, A.N. Cytomegalovirus-specific CD4+ T cells in healthy carriers are continuously driven to replicative exhaustion. J. Immunol. 2005, 175, 8218–8225. [Google Scholar] [CrossRef]
- Luo, X.H.; Huang, X.J.; Liu, K.Y.; Xu, L.P.; Liu, D.H. Protective immunity transferred by infusion of cytomegalovirus-specific CD8(+) T cells within donor grafts: Its associations with cytomegalovirus reactivation following unmanipulated allogeneic hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2010, 16, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.L.; Masters, J.E.; De Lara, C.M.; Henson, S.M.; Worth, A.; Zhang, Y.; Kumar, S.R.; Beverley, P.C.; Akbar, A.N.; Macallan, D.C. Human cytomegalovirus-specific CD8(+) T-cell expansions contain long-lived cells that retain functional capacity in both young and elderly subjects. Immunology 2011, 132, 27–38. [Google Scholar] [CrossRef]
- Scheinberg, P.; Melenhorst, J.J.; Brenchley, J.M.; Hill, B.J.; Hensel, N.F.; Chattopadhyay, P.K.; Roederer, M.; Picker, L.J.; Price, D.A.; Barrett, A.J.; et al. The transfer of adaptive immunity to CMV during hematopoietic stem cell transplantation is dependent on the specificity and phenotype of CMV-specific T cells in the donor. Blood 2009, 114, 5071–5080. [Google Scholar] [CrossRef]
- Waller, E.C.P.; McKinney, N.; Hicks, R.; Carmichael, A.J.; Sissons, J.G.P.; Wills, M.R. Differential costimulation through CD137 (4–1BB) restores proliferation of human virus-specific “effector memory” (CD28− CD45RAHI) CD8+ T cells. Blood 2007, 110, 4360–4366. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Matte-Martone, C.; Jain, D.; McNiff, J.; Shlomchik, W.D. Central memory CD8+ T cells induce graft-versus-host disease and mediate graft-versus-leukemia. J. Immunol. 2009, 182, 5938–5948. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Berger, C.; Wong, C.W.; Forman, S.J.; Riddell, S.R.; Jensen, M.C. Engraftment of human central memory-derived effector CD8+ T cells in immunodeficient mice. Blood 2011, 117, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, A.; Smith, A.L.; Takahashi, Y.; Wong, S.; Bahceci, E.; Cook, L.; Ramos, C.; Tawab, A.; McCoy, J.P., Jr.; Read, E.J.; et al. Differences in the phenotype, cytokine gene expression profiles, and in vivo alloreactivity of T cells mobilized with plerixafor compared with G-CSF. J. Immunol. 2013, 191, 6241–6249. [Google Scholar] [CrossRef] [PubMed]
- Bunse, C.E.; Borchers, S.; Varanasi, P.R.; Tischer, S.; Figueiredo, C.; Immenschuh, S.; Kalinke, U.; Kohl, U.; Goudeva, L.; Maecker-Kolhoff, B.; et al. Impaired functionality of antiviral T cells in G-CSF mobilized stem cell donors: Implications for the selection of CTL donor. PLoS ONE 2013, 8, e77925. [Google Scholar] [CrossRef] [PubMed]
- Sukdolak, C.; Tischer, S.; Dieks, D.; Figueiredo, C.; Goudeva, L.; Heuft, H.G.; Verboom, M.; Immenschuh, S.; Heim, A.; Borchers, S.; et al. CMV-, EBV- and ADV-specific T cell immunity: Screening and monitoring of potential third-party donors to improve post-transplantation outcome. Biol Blood Marrow Transplant. 2013, 19, 1480–1492. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T-Cell Subset | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
|---|---|---|---|---|---|---|
| CD3+ [%] | 52.39 (32.7–73.8) | 96.35 (91.9–98.9) | 98.08 (95.3–99.7) | 96.19 (91.2–98.5) | 94.83 (86.8–99.0) | 96.11 (90.4–98.9) |
| TN [%] | 43.46 (19.6–63.2) | 43.6 (14.7–65.8) | 1.06 (0–8.3) | 1.63 (0.0–13.0) | 65.97 (35.0–91.9) | 54.78 (32.2–74.5) |
| TCM [%] | 28.62 (153–49.0) | 25.13 (10.6–44.8) | 56.99 (31.4–74.2) | 1.77 (0.0–15.4) | 1.08 (0.2–4.6) | 41.18 (22.9–67.0) |
| TEM [%] | 19.18 (9.3–34.7) | 20.4 (8.0–40.8) | 40.77 (24.0–60.4) | 66.15 (32.7–90.3) | 1.1 (0.2–4.0) | 2.6 (0.2–9.4) |
| TEMRA [%] | 9.72 (1.4–28.1) | 11.57 (17–32.7) | 1.11 (0.0–17.8) | 28.7 (7.1–64.5) | 33.39 (7.2–62.6) | 1.15 (0.0–6.3) |
| CD4+ [%] | 77.97 (51.3–93.2) | 68.28 (46.7–87.7) | 82.81 (71.3–92.6) | 48.98 (31.9–81.3) | 59.84 (36.8–86.6) | 73.99 (53.4–94.1) |
| CD8+ [%] | 18.85 (3.4–48.7) | 30.29 (8.9–50.3) | 16.34 (7.2–27.9) | 48.47 (16.1–63.9) | 38.15 (9.8–63.2) | 24.66 (3.6–46.6) |
| CD4/8 ratio | 4.13 | 2.25 | 5.08 | 1.01 | 1.56 | 3.0 |
| Tregs [%] | 5.7 (3.1–9.3) | 5.8 (2.7–11.5) | 6.58 (1.2–13.4) | 4.85 (1.1–12.1) | 5.1 (1.7–9.7) | 5.13 (2.5–7.4) |
| γδ T cells [%] | 3.16 (0.5–6.1) | 1.9 (0.5–3.7) | 1.41 (0.3–4.0) | 2.63 (0.6–5.7) | 2.7 (0.5–4.3) | 2.02 (0.5–5.7) |
| T-Cell Subset | PBMC | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
|---|---|---|---|---|---|---|
| CD3+ [%] | 54.52 (32.7–73.8) | 96.15 (92.4–98.4) | 98.25 (96.7–99.7) | 96.08 (92.0–98.0) | 94.48 (88.3–98.7) | 95.79 (90.4–98.8) |
| TN [%] | 43.85 (19.6–63.2) | 43.77 (23–65.8) | 0.88 (0.0–8.3) | 0.9 (0.0–6.8) | 67.28 (35–91.9) | 59.49 (32.2–74.5) |
| TCM [%] | 26.70 (15.3–49.0) | 22.96 (10.6–44.8) | 54.08 (31.4–74.2) | 0.82 (0.0–8.6) | 0.57 (0.2–1.1) | 38.72 (25.3–67) |
| TEM [%] | 19.52 (9.3–30) | 21.05 (10.9–31.7) | 43.48 (25.6–58.2) | 65.08 (34.5–89.3) | 0.78 (0.3–1.8) | 1.042 (0.2–3.1) |
| TEMRA [%] | 9.93 (1.4–28.1) | 12.24 (3.1–32.7) | 1.55 (0.0–17.8) | 33.17 (10.6–64.5) | 31.37 (7.2–65.6) | 0.76 (0–2.7) |
| CD4+ [%] | 77.21 (51.3–93.2) | 65.41 (51.3–82.9) | 80.58 (74.8–90.7) | 44.49 (32.6–56.4) | 56.86 (36.8–76.1) | 71.98 (53.4–89.2) |
| TN [%] | 44.08 (13.6–60.8) | 42.11 (13–63) | 0.33 (0.0–1.8) | 2.07 (0.0–11.0) | 83.78 (64.2–96.7) | 52.7 (17.2–72.7) |
| TCM [%] | 33.76 (21.1–72.0) | 33.02 (16.6–59.7) | 59.97 (44.9–74.3) | 8.42 (0–77.8) | 1.13 (0.1–3.3) | 45.72 (27.1–82.4) |
| TEM [%] | 17.40 (10.9–35.8) | 20.1 (10.0–35.3) | 39.51 (25.5–55) | 77.29 (11.5–98.8) | 1.05 (0.2–2.6) | 1.275 (0.2–4) |
| TEMRA [%] | 4.77 (0.4–20.8) | 4.2 (0.2–21.2) | 0.19 (0.0–1.6) | 12.23 (0.6–45.2) | 14.03 (2.5–33.6) | 0.29 (0.0–1.0) |
| CD8+ [%] | 20.08 (6.3–48.7) | 33.39 (15.9–48.7) | 18.80 (9.0–25.2) | 53.12 (42.0–63.0) | 41.31 (22.1–63.2) | 27.03 (9.9–46.6) |
| TN [%] | 35.7 (0.8–81.9) | 39.63 (22.1–80.8) | 0.44 (0.1–1.5) | 0.92 (0.0–3.6) | 42.23 (17.4–89.6) | 66.41 (34.3–93.7) |
| TCM [%] | 7.99 (0.2–25.8) | 10.26 (1.8–35.7) | 31.57 (13.3–62.8) | 0.43 (0.0–4.0) | 0.88 (0.1–6.2) | 27.78 (2.8–60.9) |
| TEM [%] | 25.73 (3.2–51.5) | 29.96 (2.9–48.8) | 67.58 (36.6–86.2) | 49.33 (7.1–77.7) | 0.94 (0.1–2.1) | 3.47 (0.3–13.3) |
| TEMRA [%] | 30.58 (7.4–72.1) | 20.16 (3.1–51) | 0.39 (0.0–1.5) | 49.29 (20.3–91.2) | 55.94 (9.6–81.7) | 2.33 (0.1–6.4) |
| CD4/8 ratio | 3.85 | 1.96 | 4.29 | 0.84 | 1.38 | 2.66 |
| Tregs [%] | 6.43 (3.3–9.3) | 6.63 (4.8–11.5) | 7.62 (1.2–13.4) | 4.42 (1.1–12.1) | 5.7 (1.7–9.7) | 5.57 (2.5–7.4) |
| γδ T cells [%] | 2.93 (0.5–4.6) | 1.73 (0.5–3.2) | 1.16 (0.3–2.9) | 2.22 (0.6–4.30) | 2.68 (0.5–3.8) | 2.3 (0.5–5.7) |
| A | ||||||
| ppCMV_pp65 (ALL) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.28 (0.1–0.5) | 0.72 (0.2–1.3) | 0.42 (0.2–0.7) | 2.15 (0.4–5.9) | 0.84 (0.2–2.2) | 0.18 (0.0–0.4) |
| CD4+IFN-γ+ | 0.26 (0.1–0.6) | 0.3 (0.1–0.6) | 0.2 (0.0–0.4) | 1.33 (0.2–2.9) | 0.44 (0.1–0.8) | 0.13 (0.0–0.2) |
| CD8+IFN-γ+ | 0.54 (0.2–1.0) | 1.44 (0.3–2.8) | 1.82 (0.3–3.4) | 3 (0.6–8.4) | 1.38 (0.3–3.4) | 0.38 (0.0–0.9) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ+ | 0.74 (0.7–1.4) | 1.06 (0.3–2.2) | 0.58 (0.2–1.1) | 2.65 (0.5–7.1) | 1.06 (0.3–2.2) | 0.33 (0.2–0.5) |
| CD4+TNF-ɑ+ | 0.76 (0.2–2.0) | 0.66 (0.2–1.3) | 0.3 (0.1–0.7) | 1.73 (0.4–3.6) | 0.52 (0.3–1.0) | 0.23 (0.1–0.4) |
| CD8+TNF-ɑ+ | 1.04 (0.7–1.4) | 2.02 (0.8–3.9) | 2.3 (0.7–3.8) | 3.35 (0.9–9.0) | 1.76 (0.5–3.5) | 0.63 (0.3–1.2) |
| ppCMV_IE1 (ALL) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.26 (0.1–0.6) | 0.64 (0.1–2.0) | 0.36 (0.1–0.9) | 0.45 (0.0–1.5) | 1.13 (0.0–5.8) | 0.13 (0.0–0.3) |
| CD4+IFN-γ+ | 0.20 (0.0–0.7) | 0.18 (0.0–0.5) | 0.06 (0.0–0.1) | 0.08 (0.0–0.1) | 0.64 (0.0–2.5) | 0.05 (0.0–0.1) |
| CD8+IFN-γ+ | 0.38 (0.1–0.8) | 1.18 (0.1–3.7) | 1.42 (0.1–4.1) | 0.93 (0.0–3.2) | 2.16 (0.0–9.1) | 0.25 (0.0–0.7) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ+ | 0.66 (0.2–1.0) | 1.06 (0.1–3.3) | 0.52 (0.1–1.3) | 3.05 (0.3–7.3) | 1.6 (0.2–6.2) | 0.23 (0.1–0.5) |
| CD4+TNF-ɑ+ | 0.62 (0.1–2.0) | 0.52 (0.1–1.3) | 0.14 (0.0–0.2) | 0.23 (0.1–0.4) | 0.76 (0.1–2.1) | 0.13 (0.1–0.2) |
| CD8+TNF-ɑ+ | 1.06 (0.5–1.9) | 2.04 (0.5–5.5) | 2.3 (0.3–6.0) | 4.88 (0.6–11.4) | 2.84 (0.3–10.1) | 0.45 (0.2–1.0) |
| B | ||||||
| ppCMV_pp65 in donors with expected response (DER) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.23 (0.1–0.4) | 0.47 (0.2–0.9) | 0.47 (0.3–0.7) | 0.9 (0.4–1.7) | 0.4 (0.2–0.6) | 0.1 (0.0–0.2) |
| CD4+IFN-γ+ | 0.17 (0.1–0.2) | 0.2 (0.1–0.4) | 0.3 (0.1–0.4) | 0.8 (0.2–1.4) | 0.33 (0.1–0.5) | 0.1 (0.1–0.2) |
| CD8+IFN-γ+ | 0.57 (0.2–1.0) | 0.97 (0.3–2.1) | 2.3 (0.5–3.4) | 1.2 (0.9–2.4) | 0.5 (0.3–0.7) | 0.2 (0.0–0.4) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ+ | 0.53 (0.3–0.9) | 0.53 (0.3–1.0) | 0.7 (0.5–1.1) | 1.17 (0.5–2.2) | 0.57 (0.3–1.0) | 0.27 (0.2–0.4) |
| CD4+TNF-ɑ+ | 0.43 (0.2–0.7) | 0.33 (0.2–0.5) | 0.43 (0.2–0.7) | 1.1 (0.4–2) | 0.53 (0.3–1) | 0.2 (0.1–0.4) |
| CD8+TNF-ɑ+ | 0.93 (0.7–1.3) | 1.3 (0.8–2.2) | 2.67 (0.7–3.8) | 1.47 (0.9–2.4) | 0.83 (0.5–1.0) | 0.43 (0.3–0.5) |
| ppCMV_IE1 in donors with expected response (DER) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.1 (0.1–0.1) | 0.1 (0.1–0.2) | 0.1 (0.1–0.4) | 0.2 (0.1–0.4) | 0.1 (0.0–0.2) | 0.07 (0.0–0.1) |
| CD4+IFN-γ+ | 0.1 (0.1–0.2) | 0.03 (0.0–0.1) | 0.05 (0.0–0.12) | 0.1 (0.1–0.2) | 0.1 (0.0–0.1) | 0.03 (0.0–0.1) |
| CD8+IFN-γ+ | 0.2 (0.1–0.3) | 0.17 (0.1–0.3) | 0.1 (0.1–0.1) | 0.3 (0.2–0.4) | 0.2 (0.1–0.3) | 0.13 (0.0–0.3) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ+ | 0.27 (0.2–0.3) | 0.2 (0.1–0.3) | 0.1 (0.1–0.5) | 0.37 (0.3–0.5) | 0.43 (0.2–0.4) | 0.13 (0.1–0.2) |
| CD4+TNF-ɑ+ | 0.2 (0.1–0.4) | 0.17 (0.1–0.3) | 0.05 (0.2–0.3) | 0.23 (0.1–0.4) | 0.43 (0.2–0.3) | 0.1 (0.1–0.1) |
| CD8+TNF-ɑ+ | 0.63 (0.5–0.9) | 0.47 (0.4–0.5) | 0.35 (0.3–0.4) | 0.7 (0.6–0.8) | 0.57 (0.3–0.8) | 0.23 (0.2–0.3) |
| C | ||||||
| ppCMV_pp65 in donors with unexpected response (DUR) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.35 (0.2–0.5) | 1.1 (0.9–1.3) | 0.35 (0.2–0.5) | 5.9 (5.9–5.9) | 1.5 (0.8–2.2) | 0.4 (0.4–0.4) |
| CD4+IFN-γ+ | 0.4 (0.2–0.6) | 0.45 (0.3–0.6) | 0.05 (0.0–0.1) | 2.9 (2.9–2.9) | 0.6 (0.4–0.8) | 0.2 (0.2–0.2) |
| CD8+IFN-γ+ | 0.5 (0.4–0.6) | 2.15 (1.5–2.8) | 1.1 (0.3–1.9) | 8.4 (8.4–8.4) | 2.7 (2.0–3.4) | 0.9 (0.9–0.9) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ+ | 1.05 (0.7–1.4) | 1.85 (1.5–2.2) | 0.4 (0.2–0.6) | 7.1 (7.1–7.1) | 1.8 (1.4–2.2) | 0.5 (0.5–0.5) |
| CD4+TNF-ɑ+ | 1.25 (0.5–2.0) | 1.15 (1.0–1.3) | 0.1 (0.1–0.1) | 3.6 (3.6–3.6) | 0.5 (0.5–0.5) | 0.3 (0.3–0.3) |
| CD8+TNF-ɑ+ | 1.2 (1.0–1.4) | 3.1 (2.3–3.9) | 1.75 (0.9–2.6) | 9.0 (9.0–9.0) | 3.15 (2.8–3.5) | 1.2 (1.2–1.2) |
| ppCMV_IE1 in donors with unexpected response (DUR) | ||||||
| IFN-γ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+IFN-γ+ | 0.37 (0.1–0.6) | 1.0 (0.4–2.0) | 0.53 (0.3–0.9) | 0.75 (0.0–1.5) | 2.17 (0.0–5.8) | 0.2 (0.1–0.3) |
| CD4+IFN-γ+ | 0.27 (0.0–0.7) | 0.3 (0.0–0.5) | 0.07 (0.0–0.1) | 0.07 (0.0–0.1) | 1.03 (0.0–2.5) | 0.07 (0.0–0.1) |
| CD8+IFN-γ+ | 0.53 (0.2–0.8) | 1.9 (0.8–3.7) | 2.3 (1.4–4.1) | 1.6 (0.0–3.2) | 3.5 (0.0–9.1) | 0.45 (0.2–0.7) |
| TNF-ɑ | PBMCs | CD3+ | CD45RA_NF | CD62L_NF | CD45RA_PF | CD62L_PF |
| CD3+TNF-ɑ | 0.97 (0.5–1.7) | 1.63 (0.7–3.3) | 0.8 (0.5–1.3) | 5.8 (4.3–7.3) | 2.5 (0.4–6.2) | 0.35 (0.2–0.5) |
| CD4+TNF-ɑ+ | 0.93 (0.4–2.0) | 0.73 (0.1–1.3) | 0.2 (0.2–0.2) | 0.2 (0.2–0.2) | 1.1 (0.1–2.1) | 0.15 (0.1–0.2) |
| CD8+TNF-ɑ+ | 1.3 (0.5–1.9) | 3.07 (1.8–5.5) | 3.6 (2.0–6.0) | 9.1 (6.8–11.4) | 4.43 (1.0–10.1) | 0.7 (0.4–1.0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangare, C.; Tischer-Zimmermann, S.; Riese, S.B.; Dragon, A.C.; Prinz, I.; Blasczyk, R.; Maecker-Kolhoff, B.; Eiz-Vesper, B. Robust Identification of Suitable T-Cell Subsets for Personalized CMV-Specific T-Cell Immunotherapy Using CD45RA and CD62L Microbeads. Int. J. Mol. Sci. 2019, 20, 1415. https://doi.org/10.3390/ijms20061415
Mangare C, Tischer-Zimmermann S, Riese SB, Dragon AC, Prinz I, Blasczyk R, Maecker-Kolhoff B, Eiz-Vesper B. Robust Identification of Suitable T-Cell Subsets for Personalized CMV-Specific T-Cell Immunotherapy Using CD45RA and CD62L Microbeads. International Journal of Molecular Sciences. 2019; 20(6):1415. https://doi.org/10.3390/ijms20061415
Chicago/Turabian StyleMangare, Caroline, Sabine Tischer-Zimmermann, Sebastian B. Riese, Anna C. Dragon, Immo Prinz, Rainer Blasczyk, Britta Maecker-Kolhoff, and Britta Eiz-Vesper. 2019. "Robust Identification of Suitable T-Cell Subsets for Personalized CMV-Specific T-Cell Immunotherapy Using CD45RA and CD62L Microbeads" International Journal of Molecular Sciences 20, no. 6: 1415. https://doi.org/10.3390/ijms20061415