Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants



Abstract
1. Introduction
2. Harmful Effects of TEs and Their Consequence on Plant Organisms
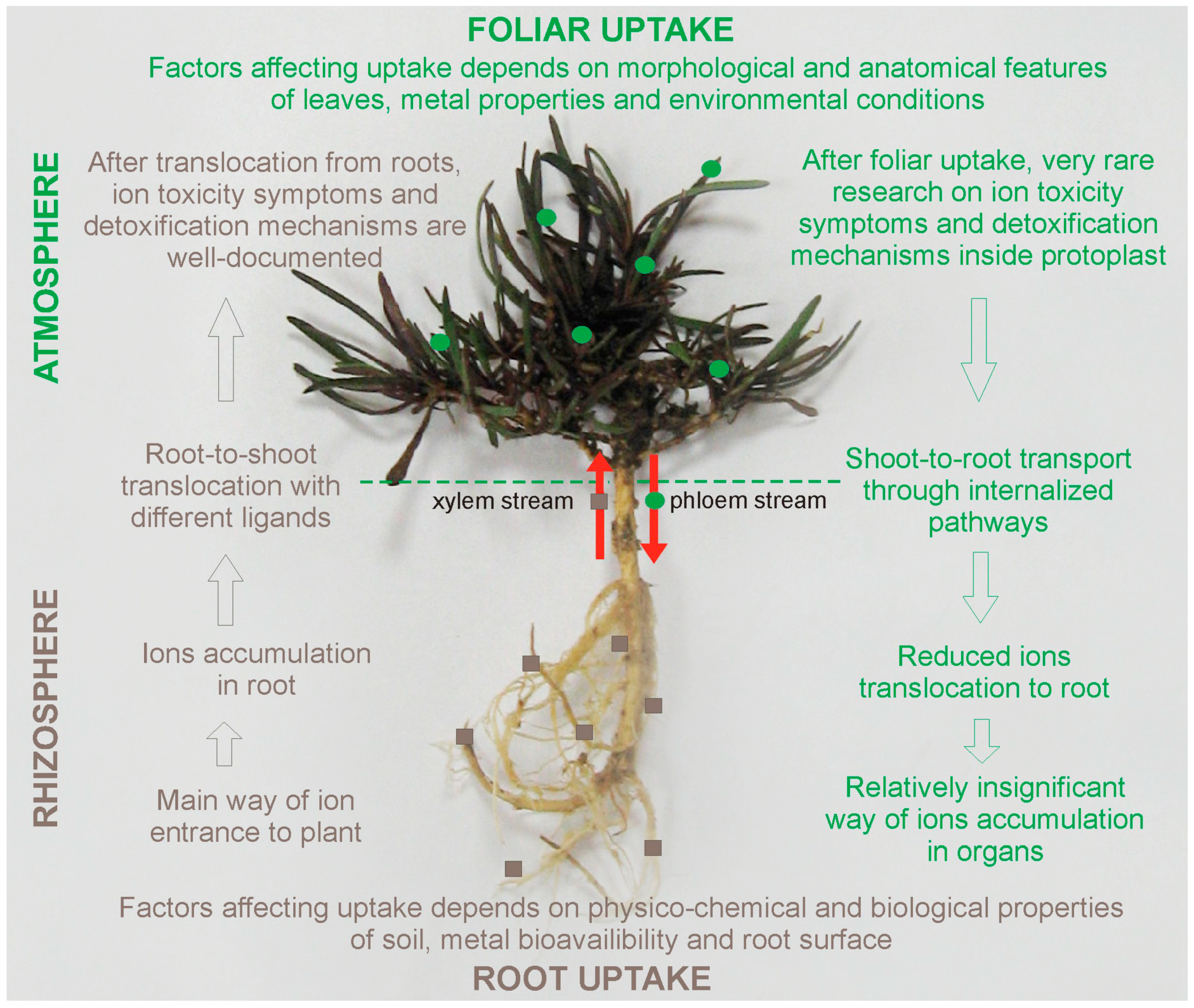
2.1. Possible Pathways of TEs Uptake by Plants
2.2. The confrontation of Root and Foliar Pathways in TEs Interaction with Plants
2.3. Structural and Metabolic Modifications under Metallic Elements Exposure
3. Second Face of TEs—Beneficial Significance in Abiotic Stress Mitigation by ROS Scavenger Activation
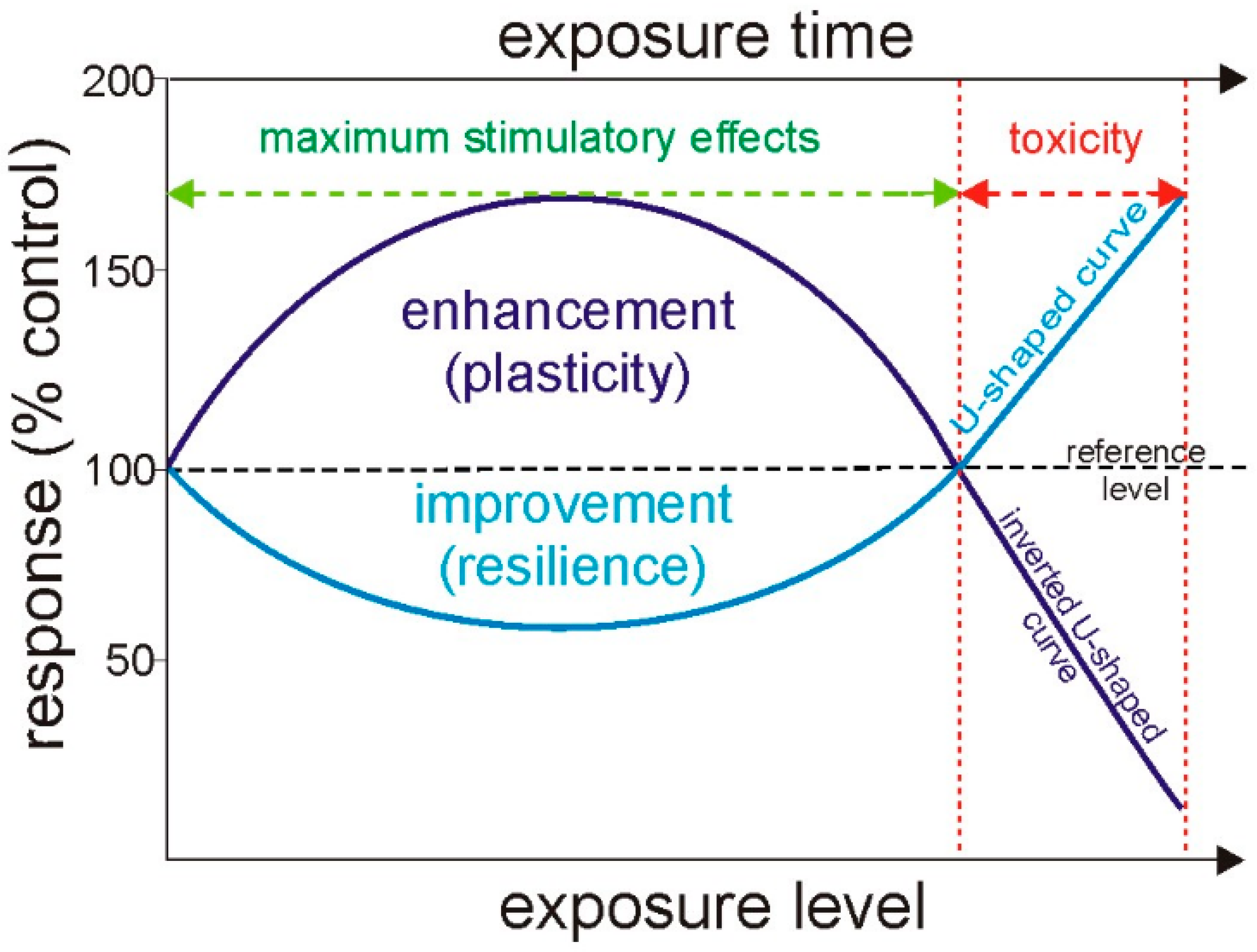
4. Hormesis Effect
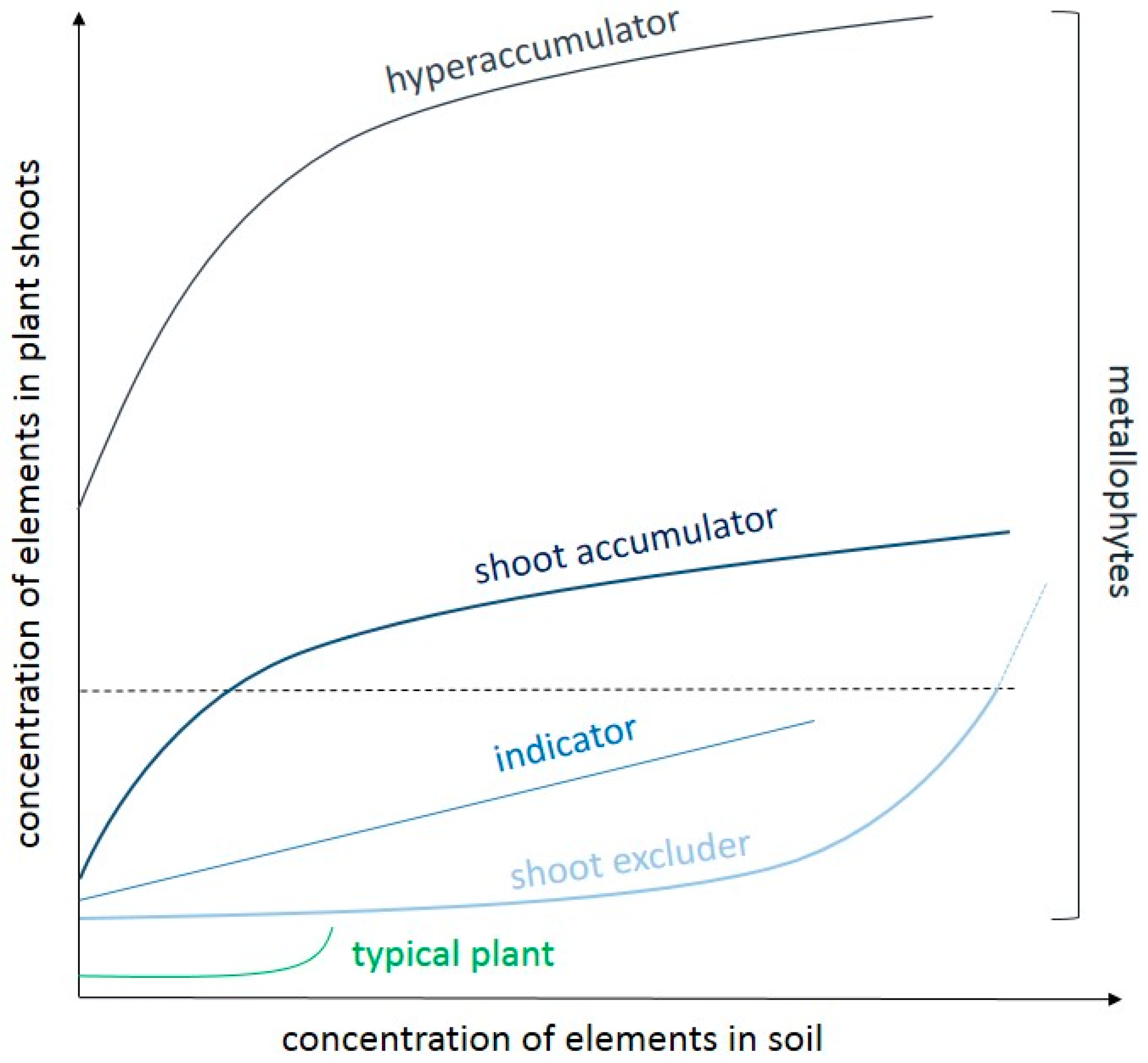
5. Metallophytes as Unique Communities from Metalliferous Sites
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carrasco-Gil, S.; Rios, J.J.; Álvarez-Fernández, A.; Abadía, A.; García-Mina, J.M.; Abadía, J. Effects of individual and combined metal foliar fertilisers on iron- and manganese-deficient Solanum lycopersicum plants. Plant Soil 2016, 402, 27–45. [Google Scholar] [CrossRef]
- Reuscher, S.; Kolter, A.; Hoffmann, A.; Pillen, K.; Krämer, U. Quantitative trait loci and inter-organ partitioning for essential metal and toxic analogue accumulation in barley. PLoS ONE 2016, 11, e0153392. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.Y. Effects of leaf area and transpiration rate on accumulation and compartmentalization of cadmium in Impatiens walleriana. Water Air Soil Pollut. 2015, 226, 2246. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Eqani, S.; Kanwal, A.; Bhowmik, A.K.; Sohail, M.; Ullah, R.; Ali, S.M.; Alamdar, A.; Ali, N.; Fasola, M.; Shen, H. Spatial distribution of dust-bound trace elements in Pakistan and their implications for human exposure. Environ. Pollut. 2016, 213, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, E.; Hanus-Fajerska, E.; Piwowarczyk, B.; Augustynowicz, J.; Ciarkowska, K.; Czech, T. From laboratory to field studies–the assessment of Biscutella laevigata suitability to biological reclamation of areas contaminated with lead and cadmium. Ecotoxicol. Environ. Saf. 2017, 142, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.P.; Human, L.R.D.; Adams, J.B. Wetland plants as indicators of heavy metal contamination. Mar. Pollut. Bull. 2015, 92, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, D.; Padhy, P.K. Metals uptake from particulate matter through foliar transfer and their impact on antioxidant enzymes activity of S. robusta in a tropical forest, West Bengal, India. Arch. Environ. Contam. Toxicol. 2019, 76, 605–616. [Google Scholar] [CrossRef]
- Xiong, T.; Zhang, T.; Dumat, C.; Sobanska, S.; Dappe, V.; Shahid, M.; Xian, Y.; Li, X.; Li, S. Airborne foliar transfer of particular metals in Lactuca sativa L.: Translocation, phytotoxicity, and bioaccessibility. Environ. Sci. Pollut. Res. Int. 2018. [Google Scholar] [CrossRef]
- Chandra, R.; Kumar, V. Phytoextraction of heavy metals by potential native plants and their microscopic observation of root growing on stabilised distillery sludge as a prospective tool for in situ phytoremediation of industrial waste. Environ. Sci. Pollut. Res. 2017, 24, 2605–2619. [Google Scholar] [CrossRef]
- Muszyńska, E.; Hanus-Fajerska, E.; Ciarkowska, K. Studies on lead and cadmium toxicity in Dianthus carthusianorum calamine ecotype cultivated in vitro. Plant Biol. 2018, 20, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Duffus, J.H. “Heavy metals” a meaningless term? (IUPAC Technical Report). Pure Appl. Chem. 2002, 74, 793–807. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Hanus-Fajerska, E.; Ciarkowska, K.; Muszyńska, E. Long-term field study on stabilization of contaminated wastes by growing clonally reproduced Silene vulgaris calamine ecotype. Plant Soil 2019. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Ma, X.; Guo, L.; He, Y.; Ren, Z.; Kuang, Z.; Zhang, X.; Zhang, Z. Analysis of potential strategies for cadmium stress tolerance revealed by transcriptome analysis of upland cotton. Sci. Rep. 2019, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Zhang, J.; Ma, Y.; Li, Y.; Zhang, F.; Zhang, Y.; Liang, Y. Overexpression of a mitogen-activated protein kinase SlMAPK3 positively regulates tomato tolerance to cadmium and drought stress. Molecules 2019, 24, 556. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.S.; Singh, S.; Srivastava, S.; Singh, N.K.; Mani, A. Whole transcriptome expression profiling and biological network analysis of chickpea during heavy metal stress. J. Plant Biochem. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Torabian, S.; Zahedi, M.; Khoshgoftar, A.H. Effects of foliar spray of two kinds of zinc oxide on the growth and ion concentration of sunflower cultivars under salt stress. J. Plant Nutr. 2016, 39, 172–180. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Zhao, F.J.; Sun, C.; Jin, C.; Shi, Y.; Sun, Y.; Li, Y.; Yang, M.; Jing, X.; et al. OsATX1 interacts with heavy metal P1B-type ATPases and affects copper transport and distribution. Plant Physiol. 2018, 178, 329–344. [Google Scholar] [CrossRef]
- Bian, X.; Cui, J.; Tang, B.; Yang, L. Chelant-induced phytoextraction of heavy metals from contaminated soils: A review. Pol. J. Environ. Stud. 2018, 27, 2417–2424. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Tomioka, R.; Takenaka, C. Accumulation of cobalt and nickel in tissues of Clethra barbinervis in a metal dosing trial. Plant Soil 2017, 421, 273–283. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta BBA Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef]
- Luan, M.; Liu, J.; Liu, Y.; Han, X.; Sun, G.; Lan, W.; Luan, S. Vacuolar Phosphate transporter 1 (VPT1) affects arsenate tolerance by regulating phosphate homeostasis in Arabidopsis. Plant Cell Physiol. 2018, 59, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Khare, R.; Trivedi, P.K. Arsenic-responsive high-affinity rice sulphate transporter, OsSultr1;1, provides abiotic stress tolerance under limiting sulphur condition. J. Hazard. Mater. 2019, 373, 753–762. [Google Scholar] [CrossRef]
- Schreck, E.; Dappe, V.; Sarret, G.; Sobanska, S.; Nowak, D.; Nowak, J.; Stefaniak, E.A.; Magnin, V.; Ranieri, V.; Dumat, C. Foliar or root exposures to smelter particles: Consequences for lead compartmentalization and speciation in plant leaves. Sci. Total Environ. 2014, 476–477, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef]
- Arguello, J.M.; Raimunda, D.; Gonzalez-Guerrero, M. Metal transport across biomembranes: Emerging models for a distinct chemistry. J. Biol. Chem. 2012, 287, 13510–13517. [Google Scholar] [CrossRef]
- Kozak, K.; Papierniak, A.; Barabasz, A.; Kendziorek, M.; Palusińska, M.; Williams, L.E.; Antosiewicz, D.M. NtZIP11, a new Zn transporter specifically upregulated in tobacco leaves by toxic Zn level. Environ. Exp. Bot. 2019, 157, 69–78. [Google Scholar] [CrossRef]
- Ammar, W.B.; Zarrouk, M.; Nouairi, I. Zinc alleviates cadmium effects on growth, membrane lipid biosynthesis and peroxidation in Solanum lycopersicum leaves. Biologia 2015, 70, 198–207. [Google Scholar] [CrossRef]
- Souza, V.L.; Almeida, A.A.F.; Mangabeira, P.A.O.; Silva, D.C.; de Jesus, R.M.; Valle, R.R. Morphophysiological, ultrastructural, and nutritional changes induced by Cu toxicity in young Erythrina fusca plants. Int. J. Phytoremediat. 2017, 19, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.T.; Leveque, T.; Austruy, A.; Goix, S.; Schreck, E.; Dappe, V.; Sobanska, S.; Foucault, Y.; Dumat, C. Foliar uptake and metal(loid) bioaccessibility in vegetables exposed to particulate matter. Environ. Geochem. Health 2014, 36, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Dappe, V.; Dumez, S.; Bernard, F.; Hanoune, B.; Cuny, D.; Dumat, C.; Sobanska, S. The role of epicuticular waxes on foliar metal transfer and phytotoxicity in edible vegetables: Case of Brassica oleracea species exposed to manufactured particles. Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sgrigna, G.; Sæbø, A.; Gawronski, S.; Popek, R.; Calfapietra, C. Particulate matter deposition on Quercus ilex leaves in an industrial city of central Italy. Environ. Pollut. 2015, 197, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Gajbhiye, T.; Pandey, S.K.; Kim, K.H.; Szulejko, J.E.; Prasad, S. Airborne foliar transfer of PM bound heavy metals in Cassia siamea: A less common route of heavy metal accumulation. Sci. Total Environ. 2016, 573, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef] [PubMed]
- Kabir, A.H.; Khatun, M.A.; Hossain, M.M.; Haider, S.A.; Alam, M.F.; Paul, N.K. Regulation of phytosiderophore release and antioxidant defense in roots driven by shoot-based auxin signaling confers tolerance to excess iron in wheat. Front. Plant Sci. 2016, 7, 1684. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wang, X.; Guo, Z.; Xiao, X.; Peng, C.; Yang, J.; Zhou, C.; Zeng, P. Chelator-assisted phytoextraction of arsenic, cadmium and lead by Pteris vittata L. and soil microbial community structure response. Int. J. Phytoremediat. 2019, in press. [Google Scholar] [CrossRef]
- Baranowska-Morek, A.; Wierzbicka, M. Localization of lead in root tip of Dianthus carthusianorum. Acta Biol. Crac. Ser. Bot. 2004, 46, 45–56. [Google Scholar]
- Van Belleghem, F.; Cuypers, A.; Semane, B.; Smeets, K.; Vangronsveld, J.; d’Haen, J.; Valcke, R. Subcellular localization of cadmium in roots and leaves of Arabidopsis thaliana. New Phytol. 2007, 173, 495–508. [Google Scholar] [CrossRef]
- Tang, Y.; Cao, Y.; Qiu, J.; Gao, Z.; Ou, Z.; Wang, Y.; Zheng, Y. Expression of a vacuole-localized BURP-domain protein from soybean (SALI3-2) enhances tolerance to cadmium and copper stresses. PLoS ONE 2014, 9, e98830. [Google Scholar] [CrossRef] [PubMed]
- Chamel, A.; Pineri, M.; Escoubes, M. Quantitative determination of water sorption by plant cuticles. Plant Cell Environ. 1991, 14, 87–95. [Google Scholar] [CrossRef]
- Ge, L.; Cang, L.; Yang, J.; Zhou, D. Effects of root morphology and leaf transpiration on Cd uptake and translocation in rice under different growth temperature. Environ. Sci. Pollut. Res. 2016, 23, 24205–24214. [Google Scholar] [CrossRef] [PubMed]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Callahan, D.L.; Hare, D.J.; Bishop, D.P.; Doble, P.A.; Roessner, U. Elemental imaging of leaves from the metal hyperaccumulating plant Noccaea caerulescens shows different spatial distribution of Ni, Zn and Cd. RSC Adv. 2016, 6, 2337–2344. [Google Scholar] [CrossRef]
- Li, T.; Zhang, M.; Lu, Z.; Herman, U.; Mumbengegwi, D.; Crittenden, J. Effects of heavy metals from soil and dust source on DNA damage of the Leymus chinensis leaves in coal-mining area in Northwest China. PLoS ONE 2016, 11, e0166522. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yu, Y.; Wan, Y.; Wang, Q.; Li, H. Effect of humic acid-based amendments with foliar application of Zn and Se on Cd accumulation in tobacco. Ecotoxicol. Environ. Saf. 2017, 138, 286–291. [Google Scholar] [CrossRef]
- Basile, A.; Sorbo, S.; Cardi, M.; Lentini, M.; Castiglia, D.; Cianciullo, P.; Conte, B.; Loppi, S.; Esposito, S. Effects of heavy metals on ultrastructure and Hsp70 induction in Lemna minor L. exposed to water along the Sarno River, Italy. Ecotoxicol. Environ. Saf. 2015, 114, 93–101. [Google Scholar] [CrossRef]
- Ma, J.; Lv, C.; Xu, M.; Chen, G.; Lv, C.; Gao, Z. Photosynthesis performance, antioxidant enzymes, and ultrastructural analyses of rice seedlings under chromium stress. Environ. Sci. Pollut. Res. 2016, 23, 1768–1778. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kamińska, I.; Górecka, M.; Bederska-Błaszczyk, M. Evaluation of heavy metal-induced responses in Silene vulgaris ecotypes. Protoplasma 2019, in press. [Google Scholar]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Piwowarczyk, B.; Tokarz, K.; Muszyńska, E.; Makowski, W.; Jędrzejczyk, R.; Gajewski, Z.; Hanus-Fajerska, E. The acclimatization strategies of kidney vetch (Anthyllis vulneraria L.) to Pb toxicity. Environ. Sci. Pollut. Res. 2018, 25, 19739–19752. [Google Scholar] [CrossRef] [PubMed]
- El-Ashry, Z.M.; Mohamed, F.I. Protective effects of some antioxidant metals against chromosomal damage induced by cadmium in Vicia faba plants. Int. J. Agric. Res. 2012, 7, 376–387. [Google Scholar] [CrossRef][Green Version]
- Song, Y.; Zhou, L.; Yang, S.; Wang, C.; Zhang, T.; Wang, J. Dose-dependent sensitivity of Arabidopsis thaliana seedling root to copper is regulated by auxin homeostasis. Environ. Exp. Bot. 2017, 139, 23–30. [Google Scholar] [CrossRef]
- Chaudhary, E.; Sharma, P. Chromium and cadmium removal from wastewater using duckweed - Lemna gibba L. and ultrastructural deformation due to metal toxicity. Int. J. Phytoremediat. 2019, 21, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Gill, R.A.; Mwamba, T.M.; Zhang, N.; Lv, M.T.; ul Hassan, Z.; Islam, F.; Ali, S.; Zhou, W.J. Differential cobalt-induced effects on plant growth, ultrastructural modifications, and antioxidative response among four Brassica napus (L.) cultivars. Int. J. Environ. Sci. Technol. 2018, 15, 2685–2700. [Google Scholar] [CrossRef]
- Dai, L.P.; Dong, X.J.; Ma, H.H. Molecular mechanism for cadmium-induced anthocyanin accumulation in Azolla imbricata. Chemosphere 2012, 87, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.M.; Singh, A. Metabolic responses of Azolla pinnata to cadmium stress: Photosynthesis, antioxidative system and phytoremediation. Chem. Ecol. 2011, 27, 543–555. [Google Scholar] [CrossRef]
- Lizieri, C.; Aguiar, R.; Kuki, K.N. Manganese accumulation and its effects on three tropical aquatic macrophytes: Azolla caroliniana, Salvinia mínima and Spirodela polyrhiza. Rodriguésia 2011, 62, 909–917. [Google Scholar] [CrossRef]
- Parlak, K.U. Effect of nickel on growth and biochemical characteristics of wheat (Triticum aestivum L.) seedlings. NJAS Wagening. J. Life Sci. 2016, 76, 1–5. [Google Scholar] [CrossRef]
- Medda, S.; Mondal, N.K. Chromium toxicity and ultrastructural deformation of Cicer arietinum with special reference of root elongation and coleoptile growth. Ann. Agrar. Sci. 2017, 15, 396–401. [Google Scholar] [CrossRef]
- Nigam, N.; Khare, P.; Yadav, V.; Mishra, D.; Jain, S.; Karak, T.; Panja, S.; Tandon, S. Biochar-mediated sequestration of Pb and Cd leads to enhanced productivity in Mentha arvensis. Ecotoxicol. Environ. Saf. 2019, 172, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Dresler, S.; Bednarek, W.; Wójcik, M. Effect of cadmium on selected physiological and morphological parameters in metallicolous and non-metallicolous populations of Echium vulgare L. Ecotoxicol. Environ. Saf. 2014, 104, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, P.; Kumar, V.; Joshi, U.N. Effect of chromium(VI) toxicity on enzymes of nitrogen metabolism in clusterbean (Cyamopsis tetragonoloba L.). Enzym. Res. 2014, 2014, 784036. [Google Scholar] [CrossRef] [PubMed]
- Cho, U.H.; Seo, N.H. Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci. 2005, 168, 113–120. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, W.; He, X.; Jia, L.; Yu, S.; Zhao, M. Hormetic responses of Lonicera japonica Thunb. to cadmium stress. Dose Response 2015. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Liu, Z.; Chen, W.; Ye, Y.; Yu, S.; He, X. Hormesis effects induced by cadmium on growth and photosynthetic performance in a hyperaccumulator, Lonicera japonica Thunb. J. Plant Growth Regul. 2015, 34, 13–21. [Google Scholar] [CrossRef]
- Jiang, Q.Y.; Zhuo, F.; Long, S.H.; Zhao, H.D.; Yang, D.J.; Ye, Z.-H.; Li, S.S.; Jing, Y.X. Can arbuscular mycorrhizal fungi reduce Cd uptake and alleviate Cd toxicity of Lonicera japonica grown in Cd-added soils? Sci. Rep. 2016, 6, 21805. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Singh, L.; Zularisam, A.W.; Sakinah, M.; Din, M.F.M. Lead induced oxidative stress and alteration in the activities of antioxidative enzymes in rice shoots. Biol. Plant. 2017, 61, 595–598. [Google Scholar] [CrossRef]
- Babu, T.N.; Varaprasad, D.; Yeduguri, H.; Kumari, K.; Lomada, D.; Reddy, M.; Chandrasekhar, T. Impact of heavy metals (Cr, Pb and Sn) on in vitro seed germination and seedling growth of green gram (Vigna radiata (L.) R. Wilczek). Curr. Trends Biotechnol. Pharm. 2014, 8, 160–165. [Google Scholar]
- Mehta, A.; López-Maury, L.; Florencio, F.J. Proteomic pattern alterations of the cyanobacterium Synechocystis sp. PCC 6803 in response to cadmium, nickel and cobalt. J. Proteom. 2014, 102, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Petraglia, A.; De Benedictis, M.; Degola, F.; Pastore, G.; Calcagno, M.; Ruotolo, R.; Mengoni, A.; Sanità di Toppi, L. The capability to synthesize phytochelatins and the presence of constitutive and functional phytochelatin synthases are ancestral (plesiomorphic) characters for basal land plants. J. Exp. Bot. 2014, 65, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Schor-Fumbarov, T.; Goldsbrough, P.B.; Adam, Z.; Tel-Or, E. Characterization and expression of a metallothionein gene in the aquatic fern Azolla filiculoides under heavy metal stress. Planta 2005, 223, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Shivendra Vikram, S.; JC Favas, P.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2014, 55, 54. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.L.; Han, H.; Meng, F.R.; Cai, L.P.; Liu, A.Q. Intermittent lead-induced stress on antioxidant enzyme activity and subcellular distribution of Pb in Pogonatherum crinitum seedlings. Plant Biol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, J.; Lee, S.; Wen, R. Copper-caused oxidative stress triggers the activation of antioxidant enzymes via ZmMPK3 in maize leaves. PLoS ONE 2018, 13, e0203612. [Google Scholar] [CrossRef] [PubMed]
- Drążkiewicz, M.; Skórzyńska-Polit, E.; Krupa, Z. Copper-induced oxidative stress and antioxidant defence in Arabidopsis thaliana. Biometals 2004, 17, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, W.; Chen, Z.; Gao, Q.; Xu, Q.; Cao, H. A role for APX1 gene in lead tolerance in Arabidopsis thaliana. Plant Sci. 2017, 256, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhao, X.; Fang, J.; Xiao, Y. Physiological responses and metal uptake of Miscanthus under cadmium/arsenic stress. Environ. Sci. Pollut. Res. 2018, 25, 28275–28284. [Google Scholar] [CrossRef] [PubMed]
- Seth, C.S.; Chaturvedi, P.K.; Misra, V. Toxic effect of arsenate and cadmium alone and in combination on giant duckweed (Spirodela polyrrhiza L.) in response to its accumulation. Environ. Toxicol. 2007, 22, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, D.; Li, S.; Dai, Y.; Cao, Y. Evolutionary and functional analysis of the plant-specific NADPH oxidase gene family in Brassica rapa L. R. Soc. Open Sci. 2019, 6, 181727. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef]
- Maleki, M.; Ghorbanpour, M.; Kariman, K. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene 2017, 11, 247–254. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Mishra, B.; Sangwan, R.S.; Mishra, S.; Jadaun, J.S.; Sabir, F.; Sangwan, N.S. Effect of cadmium stress on inductive enzymatic and nonenzymatic responses of ROS and sugar metabolism in multiple shoot cultures of Ashwagandha (Withania somnifera Dunal). Protoplasma 2014, 251, 1031–1045. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Różańska, E.; Hanus-Fajerska, E.; Znojek, E. Heavy metal tolerance in contrasting ecotypes of Alyssum montanum. Ecotoxicol. Environ. Saf. 2018, 161, 305–317. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Gautam, V.; Bali, S.; Sharma, A.; Khanna, K.; Arora, S.; Thukral, A.K.; Ohri, P.; Karpets, Y.V.; et al. ROS Signaling in Plants Under Heavy Metal Stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 185–214. [Google Scholar]
- Rahman, A.; Hossain, M.S.; Mahmud, J.A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced salt stress tolerance in rice seedlings: Regulation of ion homeostasis, antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2016, 22, 291–306. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Różańska, E.; Hanus-Fajerska, E.; Koszelnik-Leszek, A. Structural, physiological and genetic diversification of Silene vulgaris ecotypes from heavy metal-contaminated areas and their synchronous in vitro cultivation. Planta 2019, 249, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Schickler, H.; Caspi, H. Response of antioxidative enzymes to nickel and cadmium stress in hyperaccumulator plants of the genus Alyssum. Physiol. Plant. 1999, 105, 39–44. [Google Scholar] [CrossRef]
- Herald, V.L.; Heazlewood, J.L.; Day, D.A.; Millar, A.H. Proteomic identification of divalent metal cation binding proteins in plant mitochondria. FEBS Lett. 2003, 537, 96–100. [Google Scholar] [CrossRef]
- López-Millán, A.F.; Ellis, D.R.; Grusak, M.A. Effect of zinc and manganese supply on the activities of superoxide dismutase and carbonic anhydrase in Medicago truncatula wild type and raz mutant plants. Plant Sci. 2005, 168, 1015–1022. [Google Scholar] [CrossRef]
- Kaur, C.; Tripathi, A.K.; Nutan, K.K.; Sharma, S.; Ghosh, A.; Tripathi, J.K.; Pareek, A.; Singla-Pareek, S.L.; Sopory, S.K. A nuclear-localized rice glyoxalase I enzyme, OsGLYI-8, functions in the detoxification of methylglyoxal in the nucleus. Plant J. 2017, 89, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Kushwaha, H.R.; Hasan, M.R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Presence of unique glyoxalase III proteins in plants indicates the existence of shorter route for methylglyoxal detoxification. Sci. Rep. 2016, 6, 18358. [Google Scholar] [CrossRef]
- Mustafiz, A.; Ghosh, A.; Tripathi, A.K.; Kaur, C.; Ganguly, A.K.; Bhavesh, N.S.; Tripathi, J.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. A unique Ni2+-dependent and methylglyoxal-inducible rice glyoxalase I possesses a single active site and functions in abiotic stress response. Plant J. 2014, 78, 951–963. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Xu, S.; Sun, X. Nitric oxide mediates molybdenum-induced antioxidant defense in wheat under drought stress. Front. Plant Sci. 2017, 8, 1085. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological responses and yield of wheat plants in zinc-mediated alleviation of drought stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef]
- Li, C.Z.; Wang, D.; Wang, G.X. The protective effects of cobalt on potato seedling leaves during osmotic stress. Bot. Bull. Acad. Sin. 2005, 46, 119–125. [Google Scholar]
- Gómez-Muñoz, B.; Lekfeldt, J.D.S.; Magid, J.; Jensen, L.S.; de Neergaard, A. Seed treatment with Penicillium sp. or Mn/Zn can alleviate the negative effects of cold stress in maize grown in soils dependent on soil fertility. J. Agron. Crop Sci. 2018, 204, 603–612. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: Principles and applications. Homeopathy 2015, 104, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E. Environmental hormesis, a fundamental non-monotonic biological phenomenon with implications in ecotoxicology and environmental safety. Ecotoxicol. Environ. Saf. 2018, 148, 1042–1053. [Google Scholar] [CrossRef]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Chauhan, B.S. Can hormesis of plant-released phytotoxins be used to boost and sustain crop production? Crop Prot. 2017, 93, 69–76. [Google Scholar] [CrossRef]
- Brito, I.P.; Tropaldi, L.; Carbonari, C.A.; Velini, E. Hormetic effects of glyphosate on plants. Pest Manag. Sci. 2018, 74, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Cheng, M.; Sun, Z.; Wang, L.; Zhou, Q.; Huang, X. Combined acid rain and lanthanum pollution and its potential ecological risk for nitrogen assimilation in soybean seedling roots. Environ. Pollut. 2017, 231, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Malea, P.; Haritonidou, K.; Sperdouli, I. Copper bioaccumulation, photosystem II functioning, and oxidative stress in the seagrass Cymodocea nodosa exposed to copper oxide nanoparticles. Environ. Sci. Pollut. Res. 2017, 24, 16007–16018. [Google Scholar] [CrossRef]
- Woźniak, A.; Drzewiecka, K.; Kęsy, J.; Marczak, Ł.; Narożna, D.; Grobela, M.; Motała, R.; Bocianowski, J.; Morkunas, I. The influence of lead on generation of signalling molecules and accumulation of flavonoids in pea seedlings in response to pea aphid infestation. Molecules 2017, 22, 1404. [Google Scholar] [CrossRef]
- Siddhu, G.; Sirohi, D.S.; Kashyap, K.; Khan, I.A.; Khan, M.A.A. Toxicity of cadmium on the growth and yield of Solanum melongena L. J. Environ. Biol. 2008, 29, 853–857. [Google Scholar]
- Durenne, B.; Druart, P.; Blondel, A.; Fauconnier, M.L. How cadmium affects the fitness and the glucosinolate content of oilseed rape plantlets. Environ. Exp. Bot. 2018, 155, 185–194. [Google Scholar] [CrossRef]
- Patnaik, A.R.; Achary, V.M.M.; Panda, B.B. Chromium (VI)-induced hormesis and genotoxicity are mediated through oxidative stress in root cells of Allium cepa L. Plant Growth Regul. 2013, 71, 157–170. [Google Scholar] [CrossRef]
- Bernabé-Antonio, A.; Álvarez, L.; Buendía-González, L.; Maldonado-Magaña, A.; Cruz-Sosa, F. Accumulation and tolerance of Cr and Pb using a cell suspension culture system of Jatropha curcas. Plant Cell Tissue Organ Cult. 2015, 120, 221–228. [Google Scholar] [CrossRef]
- do Nascimento, J.L.; de Almeida, A.A.F.; Barroso, J.P.; Mangabeira, P.A.O.; Ahnert, D.; Sousa, A.G.R.; Silva, J.V.S.; Baligar, V.C. Physiological, ultrastructural, biochemical and molecular responses of young cocoa plants to the toxicity of Cr (III) in soil. Ecotoxicol. Environ. Saf. 2018, 159, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Paisio, C.E.; Fernandez, M.; González, P.S.; Talano, M.A.; Medina, M.I.; Agostini, E. Simultaneous phytoremediation of chromium and phenol by Lemna minuta Kunth: A promising biotechnological tool. Int. J. Environ. Sci. Technol. 2018, 15, 37–48. [Google Scholar] [CrossRef]
- Statwick, J.; Majestic, B.J.; Sher, A.A. Characterization and benefits of selenium uptake by an Astragalus hyperaccumulator and a non-accumulator. Plant Soil 2016, 404, 345–359. [Google Scholar] [CrossRef]
- Schmitt, M.; Watanabe, T.; Jansen, S. The effects of aluminium on plant growth in a temperate and deciduous aluminium accumulating species. AoB Plants 2016, 8, plw065. [Google Scholar] [CrossRef] [PubMed]
- Hajiboland, R.; Rad, S.B.; Barceló, J.; Poschenrieder, C. Mechanisms of aluminum-induced growth stimulation in tea (Camellia sinensis). J. Plant Nutr. Soil Sci. 2013, 176, 616–625. [Google Scholar] [CrossRef]
- Gawrońska, H.; Przybysz, A.; Szalacha, E.; Pawlak, K.; Brama, K.; Miszczak, A.; Stankiewicz-Kosyl, M.; Gawroński, S.W. Platinum uptake, distribution and toxicity in Arabidopsis thaliana L. plants. Ecotoxicol. Environ. Saf. 2018, 147, 982–989. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef]
- Agathokleous, E.; Calabrese, E.J. Hormesis can enhance agricultural sustainability in a changing world. Glob. Food Secur. 2019, 20, 150–155. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. The hormetic dose-response model is more common than the threshold model in toxicology. Toxicol. Sci. 2003, 71, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, C.; Cabot, C.; Martos, S.; Gallego, B.; Barceló, J. Do toxic ions induce hormesis in plants? Plant Sci. 2013, 212, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Preconditioning is hormesis part II: How the conditioning dose mediates protection: Dose optimization within temporal and mechanistic frameworks. Pharmacol. Res. 2016, 110, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Mattson, M.P. How does hormesis impact biology, toxicology, and medicine? Aging Mech. Dis. 2017, 3, 13. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of native plants with phytoremediation potential for highly contaminated Mediterranean soil restoration: Tools for a non-destructive and integrative approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- Wójcik, M.; Gonnelli, C.; Selvi, F.; Dresler, S.; Rostański, A.; Vangronsveld, J. Chapter One: Metallophytes of serpentine and calamine soils—Their unique ecophysiology and potential for phytoremediation. In Advances in Botanical Research; Phytoremediation; Academic Press: Cambridge, MA, USA, 2017; Volume 83, pp. 1–42. [Google Scholar]
- Ernst, W. Evolution of metal tolerance in higher plants. For. Snow Landsc. Res. 2006, 80, 251–274. [Google Scholar]
- Antonovics, J.; Bradshaw, A.D.; Turner, R.G. Heavy metal tolerance in plants. In Advances in Ecological Research; Cragg, J.B., Ed.; Academic Press: Cambridge, MA, USA, 1971; Volume 7, pp. 1–85. [Google Scholar]
- Muszyńska, E.; Hanus-Fajerska, E. In vitro multiplication of Dianthus carthusianorum calamine ecotype with the aim to revegetate and stabilize polluted wastes. Plant Cell Tissue Organ Cult. 2017, 128, 631–640. [Google Scholar] [CrossRef][Green Version]
- Wójcik, M.; Dresler, S.; Plak, A.; Tukiendorf, A. Naturally evolved enhanced Cd tolerance of Dianthus carthusianorum L. is not related to accumulation of thiol peptides and organic acids. Environ. Sci. Pollut. Res. 2015, 22, 7906–7917. [Google Scholar] [CrossRef]
- Olko, A.; Abratowska, A.; Zyłkowska, J.; Wierzbicka, M.; Tukiendorf, A. Armeria maritima from a calamine heap-initial studies on physiologic-metabolic adaptations to metal-enriched soil. Ecotoxicol. Environ. Saf. 2008, 69, 209–218. [Google Scholar] [CrossRef]
- Antosiewicz, D.M. Adaptation of plants to an environment polluted with heavy metals. Acta Soc. Bot. Pol. 1992, 61, 281–299. [Google Scholar] [CrossRef]
- Salt, D.E.; Kato, N.; Kramer, U.; Smith, R.D.; Raskin, I. The role of root exudates in nickel hyperaccumulation and tolerance in accumulator and nonaccumulator species of Thlaspi. In Phytoremediation of Contaminated Soil and Water; Lewis Publishers, Inc.: Boca Raton, FL, USA, 2000; pp. 189–200. [Google Scholar]
- Huang, G.; Guo, G.; Yao, S.; Zhang, N.; Hu, H. Organic acids, amino acids compositions in the root exudates and Cu-accumulation in castor (Ricinus communis L.) under Cu stress. Int. J. Phytoremediat. 2016, 18, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Feng, X.; Hong, C.; Chen, H.; Zeng, F.; Zheng, B.; Jiang, D. Malate secretion from the root system is an important reason for higher resistance of Miscanthus sacchariflorus to cadmium. Physiol. Plant. 2017, 159, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Dietterich, L.H.; Gonneau, C.; Casper, B.B. Arbuscular mycorrhizal colonization has little consequence for plant heavy metal uptake in contaminated field soils. Ecol. Appl. 2017, 27, 1862–1875. [Google Scholar] [CrossRef] [PubMed]
- Mnasri, M.; Janoušková, M.; Rydlová, J.; Abdelly, C.; Ghnaya, T. Comparison of arbuscular mycorrhizal fungal effects on the heavy metal uptake of a host and a non-host plant species in contact with extraradical mycelial network. Chemosphere 2017, 171, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Muschitz, A.; Riou, C.; Mollet, J.C.; Gloaguen, V.; Faugeron, C. Modifications of cell wall pectin in tomato cell suspension in response to cadmium and zinc. Acta Physiol. Plant. 2015, 37, 245. [Google Scholar] [CrossRef]
- Fernández, R.; Fernández-Fuego, D.; Bertrand, A.; González, A. Strategies for Cd accumulation in Dittrichia viscosa (L.) Greuter: Role of the cell wall, non-protein thiols and organic acids. Plant Physiol. Biochem. 2014, 78, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Samardjieva, K.A.; Tavares, F.; Pissarra, J. Histological and ultrastructural evidence for zinc sequestration in Solanum nigrum L. Protoplasma 2015, 252, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Krzesłowska, M.; Rabęda, I.; Basińska, A.; Lewandowski, M.; Mellerowicz, E.J.; Napieralska, A.; Samardakiewicz, S.; Woźny, A. Pectinous cell wall thickenings formation—A common defense strategy of plants to cope with Pb. Environ. Pollut. 2016, 214, 354–361. [Google Scholar] [CrossRef]
- Seregin, I.V.; Erlikh, N.T.; Kozhevnikova, A.D. Nickel and zinc accumulation capacities and tolerance to these metals in the excluder Thlaspi arvense and the hyperaccumulator Noccaea caerulescens. Russ. J. Plant Physiol. 2014, 61, 204–214. [Google Scholar] [CrossRef]
- Wójcik, M.; Dresler, S.; Tukiendorf, A. Physiological mechanisms of adaptation of Dianthus carthusianorum L. to growth on a Zn-Pb waste deposit—The case of chronic multi-metal and acute Zn stress. Plant Soil 2015, 390, 237–250. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Stanek, M.; Woch, M.W.; Kapusta, P. The accumulation of elements in plants growing spontaneously on small heaps left by the historical Zn-Pb ore mining. Environ. Sci. Pollut. Res. 2016, 23, 6524–6534. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, E.; Hanus-Fajerska, E. Evaluation of Scabiosa ochroleuca L. vitality after introduction on post-flotation wastes. Ochr. Srodowiska Zasobów Nat. 2016, 27, 37–41. [Google Scholar] [CrossRef]
- Lu, L.; Tian, S.; Zhang, J.; Yang, X.; Labavitch, J.M.; Webb, S.M.; Latimer, M.; Brown, P.H. Efficient xylem transport and phloem remobilization of Zn in the hyperaccumulator plant species Sedum alfredii. New Phytol. 2013, 198, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Halimaa, P.; Lin, Y.F.; Ahonen, V.H.; Blande, D.; Clemens, S.; Gyenesei, A.; Häikiö, E.; Kärenlampi, S.O.; Laiho, A.; Aarts, M.G.M.; et al. Gene expression differences between Noccaea caerulescens ecotypes help to identify candidate genes for metal phytoremediation. Environ. Sci. Technol. 2014, 48, 3344–3353. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Fernández, A.; Díaz-Benito, P.; Abadía, A.; López-Millán, A.F.; Abadía, J. Metal species involved in long distance metal transport in plants. Front. Plant Sci. 2014, 5, 105. [Google Scholar] [CrossRef]
- Agrawal, B.; Czymmek, K.J.; Sparks, D.L.; Bais, H.P. Transient Influx of nickel in root mitochondria modulates organic acid and reactive oxygen species production in nickel hyperaccumulator Alyssum murale. J. Biol. Chem. 2013, 288, 7351–7362. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, C.L.; Chaney, R.L. Growth and metal accumulation of an Alyssum murale nickel hyperaccumulator ecotype Co-cropped with Alyssum montanum and perennial ryegrass in serpentine soil. Front. Plant Sci. 2016, 7, 451. [Google Scholar] [CrossRef]
- Huguet, S.; Bert, V.; Laboudigue, A.; Barthès, V.; Isaure, M.P.; Llorens, I.; Schat, H.; Sarret, G. Cd speciation and localization in the hyperaccumulator Arabidopsis halleri. Environ. Exp. Bot. 2012, 82, 54–65. [Google Scholar] [CrossRef]
- Cappa, J.J.; Pilon-Smits, E.A.H. Evolutionary aspects of elemental hyperaccumulation. Planta 2014, 239, 267–275. [Google Scholar] [CrossRef]
- Baker, A.J.M.; Brooks, R.R. Terrestial higher plants which hyperaccumulate metallic elements. A review of their distribution, ecology and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Baliardini, C.; Meyer, C.L.; Salis, P.; Saumitou-Laprade, P.; Verbruggen, N. Cation exchanger 1 cosegregates with cadmium tolerance in the metal hyperaccumulator Arabidopsis halleri and plays a role in limiting oxidative stress in Arabidopsis spp. Plant Physiol. 2015, 169, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Dinh, N.T.; Vu, D.T.; Mulligan, D.; Nguyen, A.V. Accumulation and distribution of zinc in the leaves and roots of the hyperaccumulator Noccaea caerulescens. Environ. Exp. Bot. 2015, 110, 85–95. [Google Scholar] [CrossRef]
- Escande, V.; Olszewski, T.K.; Grison, C. From biodiversity to catalytic diversity: How to control the reaction mechanism by the nature of metallophytes. Environ. Sci. Pollut. Res. Int. 2015, 22, 5653–5666. [Google Scholar] [CrossRef]
- Roosens, N.H.C.J.; Willems, G.; Godé, C.; Courseaux, A.; Saumitou-Laprade, P. The use of comparative genome analysis and syntenic relationships allows extrapolating the position of Zn tolerance QTL regions from Arabidopsis halleri into Arabidopsis thaliana. Plant Soil 2008, 306, 105–116. [Google Scholar] [CrossRef]
- Grison, C.M.; Mazel, M.; Sellini, A.; Escande, V.; Biton, J.; Grison, C. The leguminous species Anthyllis vulneraria as a Zn-hyperaccumulator and eco-Zn catalyst resources. Environ. Sci. Pollut. Res. Int. 2015, 22, 5667–5676. [Google Scholar] [CrossRef]
- Pavoni, E.; Petranich, E.; Adami, G.; Baracchini, E.; Crosera, M.; Emili, A.; Lenaz, D.; Higueras, P.; Covelli, S. Bioaccumulation of thallium and other trace metals in Biscutella laevigata nearby a decommissioned zinc-lead mine (Northeastern Italian Alps). J. Environ. Manag. 2017, 186, 214–224. [Google Scholar] [CrossRef]
- Pošćić, F.; Fellet, G.; Vischi, M.; Casolo, V.; Schat, H.; Marchiol, L. Variation in heavy metal accumulation and genetic diversity at a regional scale among metallicolous and non-metallicolous populations of the facultative metallophyte Biscutella laevigata ssp. laevigata. Int. J. Phytoremediat. 2015, 17, 464–475. [Google Scholar] [CrossRef]
- Aksoy, A.; Zeliha, L.; Majeti, P. Metal-accumulating plants from serpentine habitats of KIzIldag, Konya Province, Turkey. Aust. J. Bot. 2015, 63, 372–378. [Google Scholar] [CrossRef]
- Antosiewicz, D.M.; Barabasz, A.; Siemianowski, O. Phenotypic and molecular consequences of overexpression of metal-homeostasis genes. Front. Plant Sci. 2014, 5, 80. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bothe, H.; Słomka, A. Divergent biology of facultative heavy metal plants. J. Plant Physiol. 2017, 219, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomic Group | Taxa Examples | Ion(s) | Response | Ref. | |
|---|---|---|---|---|---|
| Decrease | Increase | ||||
| Cyanobacteria | Synechocystis sp. PCC6803 | Ni, Cd | phycocyanin α-subunit | - | [72] |
| Ni, Co | glucose-1-phosphate adenylyltransferase | - | |||
| Ni, Co, Cd | ribulose1,5-bisphosphate carboxylase, periplasmic iron-binding protein | - | |||
| Ni | - | aspartyl/glutamyl-tRNA (Asn/Gln) amidotransferase subunit B, ferredoxin-NADP oxidoreductase | |||
| Co | aspartyl/glutamyl-tRNA (Asn/Gln) amidotransferase subunit B | - | |||
| Cd | ferredoxin-NADP oxidoreductase | - | |||
| Bryophyta | Fontinalis antipyretica Polytrichastrum formosum Sphagnum palustre | Cd | - | GSH | [73] |
| Marchantiophyta | Conocephalum conicum | Cd | - | GSH | |
| Lycopodiophyta | Selaginella denticulata | Cd | - | GSH | |
| Anthocerotophyta | Phaeoceros laevis | Cd | GSH | - | |
| Pteridophyta | Azolla imbricata, Azolla pinnata | Cd | photosynthetic rate, chlorophyll a, b | anthocyanins, carotenoids, chalcone synthase, dihydroflavonol reductase, SOD, CAT, POD, lipid peroxidation | [58,59] |
| Azolla caroliniana, Salvinia minima | Mn | chlorophylls | lipid peroxidation | [60] | |
| Azolla filiculoides | Mn | - | metallothionein gene expression | [74] | |
| Magnoliophyta -Monocots | Eichhornia crassipes | Pb | - | lipid peroxidation, SOD, CAT, POD, APX | [75] |
| Oryza sativa | Pb | CAT | lipid peroxidation, SOD, APX, GR, α-tocopherol | [70] | |
| Pogonatherum crinitum | Pb | - | lipid peroxidation, SOD, POD | [76] | |
| Triticum aestivum | Ni | chlorophylls | lipid peroxidation, proline, SOD | [61] | |
| Zea mays | Cu | - | SOD, CAT, APX, MAP- kinase activity | [77] | |
| Magnoliophyta - Dicots | Arabidopsis thaliana | Cd | SOD, CAT, GR | lipid peroxidation, APX | [66] |
| Cu | auxin level, CAT, shoot biomass | SOD, POD, chlorosis, necrosis and violet colouring of leaves | [55,78] | ||
| Pb | - | CAT, POD, GPX, GSH | [79] | ||
| Cicer arietinum | Cr | chlorophylls | - | [62] | |
| Echium vulgare | Cd | chlorophylls, carotenoids, proline, anthocyanins, phytochelatins, tartrate and succinate acids | - | [64] | |
| Lonicera japonica | Cd | GSH, APX, GR | net photosynthesis, PSII quantum efficiency, photochemical quenching, chlorophylls, carotenoids, CAT, lipid peroxidation, phytochelatins | [67,68,69] | |
| Mentha arvensis | Pb, Cd | chlorophylls | SOD, CAT, POD | [63] | |
| Miscanthus sacchariflorus | Cd, As | lipid peroxidation, POD | net photosynthesis, chlorophylls, stomatal conductance | [80] | |
| Silene vulgaris | Ni | growth rate, root and shoot development | - | [51] | |
| Pb | chlorophyll a, chlorophyll b | lipid peroxidation, phenols | |||
| Spirodela polyrrhiza | Cd, As | - | chlorophylls, carotenoids, non-protein thiols, ascorbic acid, cysteine and protein contents, biomass | [81] | |
| Element | Minimal Concentration in Leaves (mg kg−1 DW) | Main Families and Their Total Number (in Bracket) | Genera Number | Examples of Species |
|---|---|---|---|---|
| Arsenic | 1000 | Pteridaceae (1) | 2 | Pteris vittata, Pityrogramma calomelanos |
| Cadmium | 100 | Brassicaceae, Crassulaceae (6) | 7 | Arabidopsis halleri, Thlaspi (Noccaea) praecox, Solanum nigrum, Viola boachanensis |
| Copper | 300 | Asteraceae, Commelinaceae, Fabaceae, Lamiaceae, Linderniaceae, Malvaceae, Orobanchaceae, Polygonaceae (20) | 43 | Aeolanthus biformifolius, Anisopappus chinensis, Commoelina communis, Haumaniastrum katangense |
| Cobalt | 300 | Asteraceae, Lamiaceae, Linderniaceae, Orobanchaceae, Phyllanthaceae (18) | 34 | Crotalaria cobalticola, Haumaniastrum katangense, H. robertii |
| Manganese | 10,000 | Celastraceae, Myrtaceae, Proteaceae (16) | 24 | Gossia bidwillii, Virotia neurophylla |
| Nickel | 1000 | Asteraceae, Brassicaceae, Buxaceae, Cunoniaceae, Phyllanthaceae, Salicaceae, Violaceae (52) | 130 | Alyssum bertolonii, A. murale, A. lesbiacum, Berkhleya coddii, Phyllanthus cataractarum, Sebertia accuminata, Noccaea goesingense |
| Lead | 1000 | Brassicaceae, Caryophyllaceae (6) | 8 | Armeria maritima ssp. halleri, Minuartia verna, Noccaea rotondifolia subsp. cepaeifolia |
| Selenium | 100 | Brassicaceae, Fabaceae (7) | 15 | Astragalus bisulcatus, Hesperis persica, Stanleya pinnata |
| Thallium | 100 | Brassicaceae (1) | 2 | Biscutella laevigata, Iberis intermedia |
| Zinc | 3000 | Brassicaceae, Crassulaceae (9) | 12 | Anthyllis vulneraria, Arabidopsis halleri, Noccaea caerulescens, Sedum alfredii |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muszyńska, E.; Labudda, M. Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants. Int. J. Mol. Sci. 2019, 20, 3117. https://doi.org/10.3390/ijms20133117
Muszyńska E, Labudda M. Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants. International Journal of Molecular Sciences. 2019; 20(13):3117. https://doi.org/10.3390/ijms20133117
Chicago/Turabian StyleMuszyńska, Ewa, and Mateusz Labudda. 2019. "Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants" International Journal of Molecular Sciences 20, no. 13: 3117. https://doi.org/10.3390/ijms20133117
APA StyleMuszyńska, E., & Labudda, M. (2019). Dual Role of Metallic Trace Elements in Stress Biology—From Negative to Beneficial Impact on Plants. International Journal of Molecular Sciences, 20(13), 3117. https://doi.org/10.3390/ijms20133117