Characterization of the Copper Transporters from Lotus spp. and Their Involvement under Flooding Conditions

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Lotus Species Differentially Respond to Partial Submergence Conditions
2.2. Partial Submergence Conditions Affected Plant Metal Content in Lotus Species
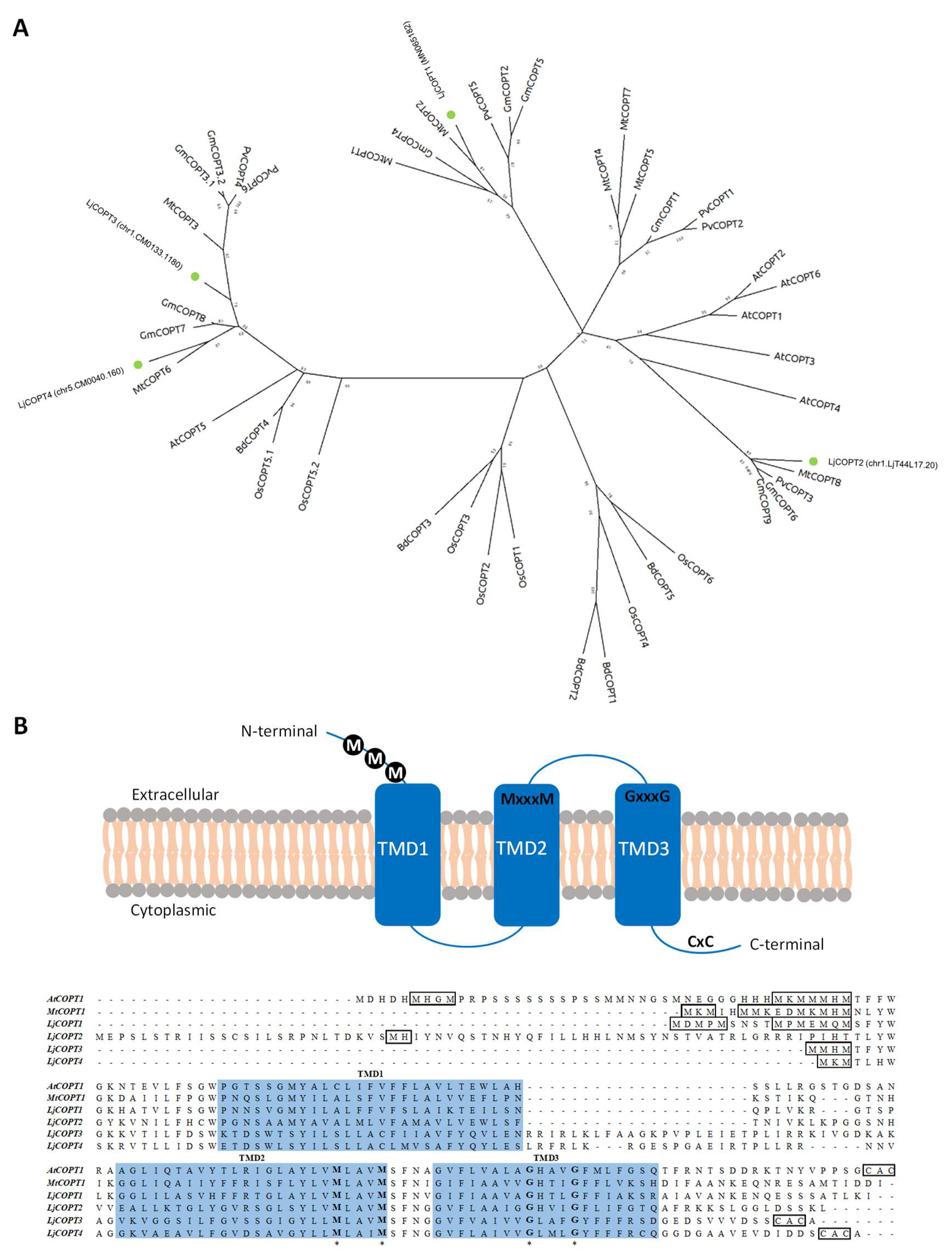
2.3. The Predicted Family of Copper Transporters from Lotus japonicus
2.4. Functional Complementation of Lotus japonicus COPTs
2.5. COPTs Gene Expression in Lotus japonicus
2.6. Regulation of Metal Homeostasis Genes under Partial Submergence Conditions
3. Discussion
4. Material and Methods
4.1. Plant Material and Treatments
4.2. Fluorescence and Gas Exchange Measurements
4.3. Copper, Iron, and Molybdenum Determination
4.4. Plasmid Constructs
4.5. Functional Complementation Experiments in Yeast
4.6. RNA Isolation and Gene Expression by Real-Time qPCR
4.7. Bioinformatics and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press Limited: London, UK, 2012; ISBN 978-0-12-384905-2. [Google Scholar]
- Halliwell, B.; Gutteridget, J.M.C. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- McDowell, L.R.; Conrad, J.H. Trace mineral nutrition in Latin America. In Ruminant Nutrition: Selected Articles from the World Animal Review; Food and Agriculture Organization of the United Estates: Rome, Italy, 1978; p. 160. ISBN 92-5-100650-4. [Google Scholar]
- Ramírez, C.E.; Mattioli, G.A.; Tittarelli, C.M.; Giuliodori, M.J.; Yano, H. Cattle hypocuprosis in Argentina associated with periodically flooded soils. Livest. Prod. Sci. 1998, 55, 47–52. [Google Scholar] [CrossRef]
- Picco, S.J.; Rosa, D.E.; Mattioli, G.A.; Fazzio, L.E. Nutrición Mineral y Vitamínica de Bovinos; CCB Academic Press: Cambridge, MA, USA, 2013; ISBN 978-987-29573-1-5. [Google Scholar]
- Soriano, A. Rio de la Plata Grasslands. In Natural Grasslands. Introduction and Western Hemisphere; Coupland, R.T., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 367–407. [Google Scholar]
- Chaneton, E.J.; Perelman, S.B.; León, R.J.C. Floristic heterogeneity of Flooding Pampa grasslands: A multi-scale analysis. Plant Biosyst. 2005, 139, 245–254. [Google Scholar] [CrossRef]
- Vignolio, O.R.; Fernández, O.N. Bioecología de Lotus glaber Mill. (Fabaceae) en la Pampa Deprimida (provincia de Buenos Aires, Argentina). Rev. Argentina Prod. Anim. 2006, 26, 113–130. [Google Scholar]
- Antonelli, C.J.; Calzadilla, P.I.; Vilas, J.M.; Campestre, M.P.; Escaray, F.J.; Ruiz, O.A. Physiological and anatomical traits associated with tolerance to long-term partial submergence stress in the Lotus genus: Responses of forage species, a model and an interspecific hybrid. J. Agron. Crop Sci. 2019, 205, 65–76. [Google Scholar] [CrossRef]
- Teakle, N.L.; Amtmann, A.; Real, D.; Colmer, T.D. Lotus tenuis tolerates combined salinity and waterlogging: Maintaining O2 transport to roots and expression of an NHX1-like gene contribute to regulation of Na+ transport. Physiol. Plant 2010, 139, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G.; Insausti, P.; Grimoldi, A.; Ploschuk, E.; Vasellati, V. Physiological and anatomical basis of differential tolerance to soil flooding of Lotus corniculatus L. and Lotus glaber Mill. Plant Soil 2005, 276, 301–311. [Google Scholar] [CrossRef]
- Colmer, T.D.; Voesenek, L. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Armstrong, W. Aeration in higher plants. Adv. Bot. Res. 1980, 7, 225–332. [Google Scholar]
- Torri, S.I.; Urricariet, S.; Lavado, R.S. Micronutrient availability in crop soils of the Pampas region, Argentina. In Soil Nutrients; Miransari, M., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2011; pp. 1–19. [Google Scholar]
- Alloway, B.J.; Tills, A.R. Copper deficiency in world crops. Outlook Agric. 1984, 13, 32–42. [Google Scholar] [CrossRef]
- Gupta, U.C.; Lipsett, J. Molybdenum in soils, plants, and animals. Adv. Agron. 1981, 34, 73–115. [Google Scholar]
- Spears, J.W. Trace mineral bioavaliability in ruminants. J. Nutr. 2003, 133, 1506S–1509S. [Google Scholar] [CrossRef] [PubMed]
- Picco, S.J.; Ponzzinibio, M.V.; Mattioli, G.A.; Rosa, D.E.; Minatel, L.; Fazzio, L.E.; Seoane, A. Physiological and genotoxic effects of molybdenum-induced copper deficiency in cattle. Agrociencia 2012, 46, 107–117. [Google Scholar]
- Mason, J. Thiomolybdates: Mediators of molybdenum toxicity and enzyme inhibitors. Toxicology 1986, 42, 99–109. [Google Scholar] [CrossRef]
- Arredondo, M.; Muñoz, P.; Mura, C.V.; Núñez, M.T. DMT1, a physiologically relevant apical Cu+1 transporter of intestinal cells. Am. J. Physiol. Physiol. 2013, 284, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Gengelbach, G.P.; Ward, J.D.; Spears, J.W. Effect of dietary copper, iron, and molybdenum on growth and copper status of beef cows and calves. J. Anim. Sci. 1994, 72, 2722–2727. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mira, H.; Thiele, D.J.; Peñarrubia, L. Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol. Biol. 2003, 51, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Lee, J.; Lau, M.; Thiele, D.J. Biochemical and genetic analyses of yeast and human high affinity copper transporters suggest a conserved mechanism for copper uptake. J. Biol. Chem. 2002, 277, 26021–26030. [Google Scholar] [CrossRef] [PubMed]
- Kampfenkel, K.; Kushnir, S.; Inzé, D.; Van Montagu, M.; Babiychuk, E.; Inze, D. Membranes and bioenergetics: Molecular characterization of a putative Arabidopsis thaliana copper transporter and its yeast homologue. J. Biol. Chem. 1995, 270, 28479–28486. [Google Scholar] [PubMed]
- Perea-García, A.; Garcia-Molina, A.; Andrés-Colás, N.; Vera-Sirera, F.; Pérez-Amador, M.A.; Puig, S.; Peñarrubia, L. Arabidopsis copper transport protein COPT2 participates in the cross talk between iron deficiency responses and low-phosphate signaling. Plant Physiol. 2013, 162, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Molina, A.; Andrés-Colás, N.; Perea-García, A.; Neumann, U.; Dodani, S.C.; Huijser, P.; Peñarrubia, L.; Puig, S. The arabidopsis COPT6 transport protein functions in copper distribution under copper-deficient conditions. Plant Cell Physiol. 2013, 54, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Hayashi, M.; Fukazawa, M.; Kobayashi, Y.; Shikanai, T. SQUAMOSA romoter binding protein-like7 is a central regulator for copper homeostasis in Arabidopsis. Plant Cell 2009, 21, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Casero, D.; Singh, V.; Wilson, G.T.; Grande, A.; Yang, H.; Dodani, S.C.; Pellegrini, M.; Huijser, P.; Connolly, E.L.; et al. Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis. Plant Cell 2012, 24, 738–761. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Colás, N.; Perea-García, A.; Puig, S.; Peñarrubia, L. Deregulated copper transport affects Arabidopsis development especially in the absence of environmental cycles. Plant Physiol. 2010, 153, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Molina, A.; Andrés-Colás, N.; Perea-García, A.; Del Valle-Tascón, S.; Peñarrubia, L.; Puig, S. The intracellular Arabidopsis COPT5 transport protein is required for photosynthetic electron transport under severe copper deficiency. Plant J. Cell Mol. Biol. 2011, 65, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mateu-Andrés, I.; Dorcey, E.; Thiele, D.J.; Peñarrubia, L. The Arabidopsis copper transporter COPT1 functions in root elongation and pollen development. J. Biol. Chem. 2004, 279, 15348–15355. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Colás, N.; Carrió-Seguí, A.; Abdel-Ghany, S.E.; Pilon, M.; Peñarrubia, L. Expression of the intracellular COPT3-mediated Cu transport is temporally regulated by the TCP16 transcription factor. Front. Plant Sci. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Gayomba, S.R.; Rutzke, M.A.; Craft, E.; Kochian, L.V.; Vatamaniuk, O.K. COPT6 is a plasma membrane transporter that functions in copper homeostasis in Arabidopsis and is a novel target of SQUAMOSA promoter-binding protein-like 7. J. Biol. Chem. 2012, 287, 33252–33267. [Google Scholar] [CrossRef] [PubMed]
- Carrió-Seguí, À.; Romero, P.; Curie, C.; Mari, S.; Peñarrubia, L. Copper transporter COPT5 participates in the crosstalk between vacuolar copper and iron pools mobilisation. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Yuan, M.; Li, X.; Xiao, J.; Wang, S. Molecular and functional analyses of COPT/Ctr-type copper transporter-like gene family in rice. BMC Plant Biol. 2011, 11, 69. [Google Scholar] [CrossRef]
- Martins, V.; Hanana, M.; Blumwald, E.; Gerós, H. Copper transport and compartmentation in grape cells. Plant Cell Physiol. 2012, 53, 1866–1880. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Du, H.; Li, H.; Huang, Y.; Ding, J.; Liu, C.; Wang, N.; Lan, H.; Zhang, S. Identification and functional characterization of the ZmCOPT copper transporter family in maize. PLoS ONE 2018, 13, e0199081. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Wedd, A.G. A C-terminal domain of the membrane copper pump Ctr1 exchanges copper(I) with the copper chaperone Atx1. Chem. Commun. 2002, 2, 588–589. [Google Scholar] [CrossRef]
- Wu, X.; Sinani, D.; Kim, H.; Lee, J. Copper transport activity of yeast Ctr1 is down-regulated via its C terminus in response to excess copper. J. Biol. Chem. 2009, 284, 4112–4122. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Andrés-Colás, N.; García-Molina, A.; Peñarrubia, L. Copper and iron homeostasis in Arabidopsis: Responses to metal deficiencies, interactions and biotechnological applications. Plant Cell Environ. 2007, 30, 271–290. [Google Scholar] [CrossRef] [PubMed]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann Bot 2009, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-takahashi, A.; Shibagaki, N. An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar] [CrossRef] [PubMed]
- Billard, V.; Ourry, A.; Maillard, A.; Garnica, M.; Coquet, L.; Jouenne, T.; Cruz, F.; Garcia-Mina, J.-M.; Yvin, J.-C.; Etienne, P. Copper-Deficiency in Brassica napus Induces Copper Remobilization, Molybdenum Accumulation and Modification of the Expression of Chloroplastic Proteins. PLoS ONE 2014, 9, e109889. [Google Scholar] [CrossRef] [PubMed]
- Gooneratne, S.R.; Buckley, W.T.; Christensen, D.A. Review of copper deficiency and metabolism in ruminants. Can. J. Anim. Sci. 1989, 69, 819–845. [Google Scholar] [CrossRef]
- Miltimore, J.E.; Mason, J. Copper to molybdenum ratio and molybdenum and copper concentrations in ruminant feeds. Can. J. Anim. Sci. 1971, 51, 193–200. [Google Scholar] [CrossRef]
- Migocka, M.; Malas, K. Plant responses to copper: Molecular and regulatory mechanisms of copper uptake, distribution and accumulation in plants. In Plant Micronutrient Use efficiency: Molecular and Genomic Perspectives in Crop Plants; Elsevier: Amsterdam, The Netherlands, 2018; pp. 71–86. ISBN 9780128122433. [Google Scholar]
- Manuel-Navarrete, D.; Gallopín, G.C.; Blanco, M.; Díaz-Zorita, M.; Ferraro, D.O.; Herzer, H.; Laterra, P.; Murmis, M.R.; Podestá, G.P.; Rabinovich, J.; et al. Multi-causal and integrated assessment of sustainability: The case of agriculturization in the Argentine Pampas. Environ. Dev. Sustain. 2007, 11, 621–638. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Lee, S.C.; Brinton, E. Waterproofing crops: Effective flooding survival strategies. Plant Physiol. 2012, 160, 1698–1709. [Google Scholar] [CrossRef] [PubMed]
- Escaray, F.J.; Menéndez, A.B.; Gárriz, A.; Pieckenstain, F.L.; Estrella, M.J.; Castagno, L.N.; Carrasco, P.; Sanjuán, J.; Ruiz, O.A. Ecological and agronomic importance of the plant genus Lotus. Its application in grassland sustainability and the amelioration of constrained and contaminated soils. Plant Sci. 2012, 182, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G.; Colmer, T.D. Flooding tolerance of forage legumes. J. Exp. Bot. 2017, 68, 1851–1872. [Google Scholar] [CrossRef] [PubMed]
- Verboven, P.; Pedersen, O.; Herremans, E.; Ho, Q.T.; Nicolaï, B.M.; Colmer, T.D.; Teakle, N. Root aeration via aerenchymatous phellem: Three-dimensional micro-imaging and radial O2 profiles in Melilotus siculus. New Phytol. 2012, 193, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, S.; Shabala, S. Waterlogging Signalling and Tolerance in Plants, 1st ed.; Mancuso, S., Shabala, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 9783642103049. [Google Scholar]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Szczyglowski, K.; Stougaard, J. Lotus genome: Pod of gold for legume research. Trends Plant Sci. 2008, 13, 515–517. [Google Scholar] [CrossRef]
- Martins, V.; Bassil, E.; Hanana, M.; Blumwald, E.; Gerós, H. Copper homeostasis in grapevine: Functional characterization of the Vitis vinifera copper transporter 1. Planta 2014, 240, 91–101. [Google Scholar] [CrossRef]
- Hirsch, A.M. Developmental biology of legume nodulation. New Phytol. 1992, 122, 211–237. [Google Scholar] [CrossRef]
- Klaumann, S.; Nickolaus, S.D.; Furst, S.H.; Starck, S.; Schneider, S.; Ekkehard, H.; Trentmann, O. The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana. New Phytol. 2011, 393–404. [Google Scholar] [CrossRef]
- Waters, B.M.; Sankaran, R.P. Moving micronutrients from the soil to the seeds: Genes and physiological processes from a biofortification perspective. Plant Sci. 2011, 180, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Kropat, J.; Tottey, S.; Birkenbihl, R.P.; Depege, N.; Huijser, P.; Merchant, S. A regulator of nutritional copper signaling in Chlamydomonas is an SBP domain protein that recognizes the GTAC core of copper response element. Proc. Natl. Acad. Sci. USA 2005, 102, 18730–18735. [Google Scholar] [CrossRef] [PubMed]
- Mustroph, A.; Lee, S.C.; Oosumi, T.; Zanetti, M.E.; Yang, H.; Ma, K.; Yaghoubi-Masihi, A.; Fukao, T.; Bailey-Serres, J. Cross-kingdom comparison of transcriptomic adjustments to low-oxygen stress highlights conserved and plant-specific responses. Plant Physiol. 2010, 152, 1484–1500. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.; Krishnan, S.P.T.; Swarup, S.; Bajic, V. Detection and preliminary analysis of motifs in promoters of anaerobically induced genes of different plant species. Ann. Appl. Biol. 2005, 96, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Gasch, P.; Fundinger, M.; Müller, J.T.; Lee, T.; Bailey-Serres, J.; Mustroph, A. Redundant ERF-VII transcription factors bind to an evolutionarily conserved cis-motif to regulate hypoxia-responsive gene expression in Arabidopsis. Plant Cell 2016, 28, 160–180. [Google Scholar] [CrossRef] [PubMed]
- Hoeren, F.U.; Dolferus, R.; Wu, Y.; Peacock, W.J.; Dennis, E.S. Evidence for a role for AtMYB2 in the induction of the Arabidopsis alcohol. Genetics 1998, 149, 479–490. [Google Scholar]
- Olive, M.R.; Peacock, W.J.; Dennis, E.S. The anaerobic responsive element contains two GC-rich sequences essential for binding a nuclear protien and hypoxic activation of the maize Adh1 promoter. Nucleic Acids Res. 1991, 19, 7053–7060. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, W.; Wu, Z.; Yang, Y.; Kang, Y.J. Copper levels affect targeting of hypoxia-inducible factor 1 to the promoters of hypoxia-regulated genes. J. Biol. Chem. 2018, 293, 14669–14677. [Google Scholar] [CrossRef] [PubMed]
- Hell, R.; Mendel, R.-R. Cell Biology of Metals and Nutrients; Springer: Berlin/Heidelberg, Germany, 2010; pp. 75–94. [Google Scholar]
- Lee, S.; Jeon, U.S.; Lee, S.J.; Kim, Y.-K.; Persson, D.P.; Husted, S.; Schjorring, J.K.; Kakei, Y.; Masuda, H.; Nishizawa, N.K.; et al. Iron fortification of rice seeds through activation of the nicotianamine synthase gene. Proc. Natl. Acad. Sci. USA 2009, 106, 22014–22019. [Google Scholar] [CrossRef]
- Alam, S.M. Nutrient uptake by plants under stress conditions. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 1999; pp. 285–313. [Google Scholar]
- Kuper, J.; Llamas, A.; Hecht, H.-J.; Mendel, R.R.; Schwarz, G. Structure of the molybdopterin-bound Cnx1G domain links molybdenum and copper metabolism. Nature 2004, 430, 803–806. [Google Scholar] [CrossRef]
- Mysore, K.S.; Wen, J.; Imperial, J.; Gil-Díez, P.; León-Mediavilla, J.; González-Guerrero, M.; Tejada-Jiménez, M. Medicago truncatula Molybdate Transporter type 1 (MtMOT1.3) is a plasma membrane molybdenum transporter required for nitrogenase activity in root nodules under molybdenum deficiency. New Phytol. 2017, 216, 1223–1235. [Google Scholar]
- Escaray, F.J.; Passeri, V.; Babuin, M.F.; Marco, F.; Carrasco, P.; Damiani, F.; Pieckenstain, F.L.; Paolocci, F.; Ruiz, O.A. Lotus tenuis x L. corniculatus interspecific hybridization as a means to breed bloat-safe pastures and gain insight into the genetic control of proanthocyanidin biosynthesis in legumes. BMC Plant Biol. 2014, 14, 14–40. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. In California Agricultural Experiment Station Circular; Hoagland, D.R., Arnon, D.I., Eds.; The College of Agriculture University of California: Berkeley, CA, USA, 1950; Volume 347, pp. 4–32. [Google Scholar]
- Strasser, R.J.; Srivastava, A. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Mumberg, D.; Müller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 1–10. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2011. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: https://www.infostat.com.ar/ (accessed on 26 June 2019).






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escaray, F.J.; Antonelli, C.J.; Copello, G.J.; Puig, S.; Peñarrubia, L.; Ruiz, O.A.; Perea-García, A. Characterization of the Copper Transporters from Lotus spp. and Their Involvement under Flooding Conditions. Int. J. Mol. Sci. 2019, 20, 3136. https://doi.org/10.3390/ijms20133136
Escaray FJ, Antonelli CJ, Copello GJ, Puig S, Peñarrubia L, Ruiz OA, Perea-García A. Characterization of the Copper Transporters from Lotus spp. and Their Involvement under Flooding Conditions. International Journal of Molecular Sciences. 2019; 20(13):3136. https://doi.org/10.3390/ijms20133136
Chicago/Turabian StyleEscaray, Francisco J., Cristian J. Antonelli, Guillermo J. Copello, Sergi Puig, Lola Peñarrubia, Oscar A. Ruiz, and Ana Perea-García. 2019. "Characterization of the Copper Transporters from Lotus spp. and Their Involvement under Flooding Conditions" International Journal of Molecular Sciences 20, no. 13: 3136. https://doi.org/10.3390/ijms20133136
APA StyleEscaray, F. J., Antonelli, C. J., Copello, G. J., Puig, S., Peñarrubia, L., Ruiz, O. A., & Perea-García, A. (2019). Characterization of the Copper Transporters from Lotus spp. and Their Involvement under Flooding Conditions. International Journal of Molecular Sciences, 20(13), 3136. https://doi.org/10.3390/ijms20133136