Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation

Abstract
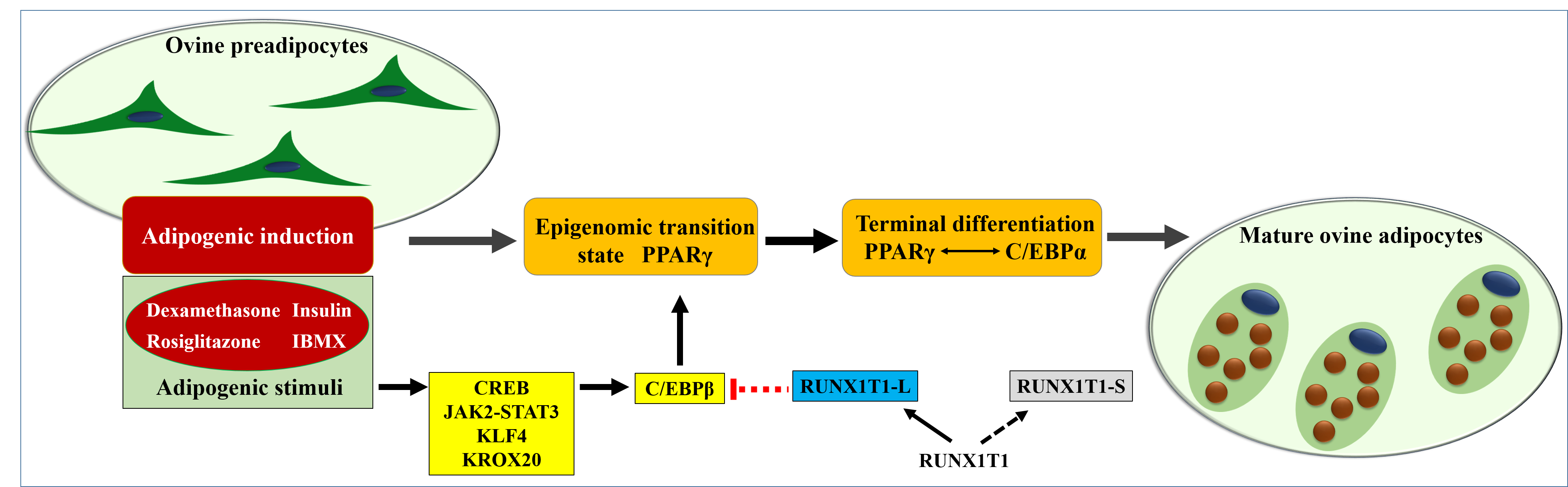
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. cDNA Cloning and Sequence Analysis
2.2. Expression Pattern of RUNX1T1 Isoforms in Different Tissues of Hu Sheep
2.3. Expression Pattern of RUNX1T1 Isoforms and Adipogenic Marker Genes in Subcutaneous Fat Tissue at Different Developmental Stages
2.4. Expression Pattern of RUNX1T1 Isoforms during Ovine Preadipocyte Differentiation
2.5. RUNX1T1-L Knockdown Promoted Lipid Accumulation and Ovine Preadipocyte Differentiation
3. Discussion
4. Material and Methods
4.1. Sample Collections
4.2. Primary Adipocyte Culture
4.3. Small Interfering RNAs (siRNAs)
4.4. Oil Red-O Staining and Extraction Assay
4.5. RNA Isolation and cDNA Synthesis
4.6. Cloning of RUNX1T1
4.7. Real-time PCR Analysis for mRNA Expression
4.8. Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (Semi-qRT-PCR)
4.9. Western Blot Analysis for Protein Expression
4.10. Immunofluorescence
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADIPOQ | Adiponectin |
| C/EBP | CCAAT/enhancer-binding protein |
| FTO | Fat mass and obesity-associated |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| LPL | Lipoprotein lipase |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| RUNX1T1 | Runt-related transcription factor 1 translocation partner 1 |
References
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Cristancho, A.G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 12, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Veiga-Lopez, A. PPAR gamma agonist through the terminal differentiation phase is essential for adipogenic differentiation of fetal ovine preadipocytes. Cell. Mol. Biol. Lett. 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Rochford, J.J.; Semple, R.K.; Laudes, M.; Boyle, K.B.; Christodoulides, C.; Mulligan, C.; Lelliott, C.J.; Schinner, S.; Hadaschik, D.; Mahadevan, M.; et al. ETO/MTG8 is an inhibitor of C/EBPβ activity and a regulator of early adipogenesis. Mol. Cell. Biol. 2004, 24, 9863–9872. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Laber, S.; McMurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.D.; Usher, S.; Sellayah, D.; Ashcroft, F.M.; et al. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Amann, J.M.; Nip, J.; Strom, D.K.; Lutterbach, B.; Harada, H.; Lenny, N.; Downing, J.R.; Meyers, S.; Hiebert, S.W. ETO, a target of t(8;21) in acute leukemia, makes distinct contacts with multiple histone deacetylases and binds mSin3A through its oligomerization domain. Mol. Cell. Biol. 2001, 21, 6470–6483. [Google Scholar] [CrossRef] [PubMed]
- Okumura, A.J.; Peterson, L.F.; Lo, M.C.; Zhang, D.E. Expression of AML/RUNX and ETO/MTG family members during hematopoietic differentiation of embryonic stem cells. Exp. Hematol. 2007, 35, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Hug, B.A.; Huang, E.Y.; Chen, C.W.; Gelmetti, V.; Maccarana, M.; Minucci, S.; Pelicci, P.G.; Lazar, M.A. Oligomerization of ETO is obligatory for corepressor interaction. Mol. Cell. Biol. 2001, 21, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.T.; Tumer, Z.; Mollgard, K.; Barbi, G.; Rossier, E.; Bendsen, E.; Moller, R.S.; Ullmann, R.; He, J.; Papadopoulos, N.; et al. Characterization of a t(5;8)(q31;q21) translocation in a patient with mental retardation and congenital heart disease: Implications for involvement of RUNX1T1 in human brain and heart development. Eur. J. Hum. Genet. 2009, 17, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Kim, M.S.; Lee, S.H.; Yoo, N.J. Mutational analysis of RUNX1T1 gene in acute leukemias, breast and lung carcinomas. Leuk. Res. 2011, 35, E157–E158. [Google Scholar] [CrossRef] [PubMed]
- Migasa, A.A.; Mishkova, O.A.; Ramanouskaya, T.V.; Ilyushonak, I.M.; Aleinikova, O.V.; Grinev, V.V. RUNX1T1/MTG8/ETO gene expression status in human t(8;21)(q22;q22)-positive acute myeloid leukemia cells. Leuk. Res. 2014, 38, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.Q.; Jin, G.H.; Li, H.M.; Tao, X.L.; Qin, J.B.; Tian, M.L. RUNX1T1 regulates the neuronal differentiation of radial glial cells from the rat hippocampus. Stem Cell Transl. Med. 2015, 4, 110–116. [Google Scholar]
- Zhao, X.; Yang, Y.; Sun, B.F.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.J.; Ping, X.L.; Chen, Y.S.; Wang, W.J.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.C.; Veiga-Lopez, A.; Moeller, J.; Beckett, E.; Pease, A.; Keller, E.; Madrigal, V.; Chazenbalk, G.; Dumesic, D.; Padmanabhan, V. Developmental programming: Impact of gestational steroid and metabolic milieus on adiposity and insulin sensitivity in prenatal testosterone-treated female sheep. Endocrinology 2016, 157, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Veiga-Lopez, A.; Moeller, J.; Sreedharan, R.; Singer, K.; Lumeng, C.; Ye, W.; Pease, A.; Padmanabhan, V. Developmental programming: Interaction between prenatal BPA exposure and postnatal adiposity on metabolic variables in female sheep. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E238–E247. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Kanbe, E.; Peterson, L.F.; Boyapati, A.; Miao, Y.; Wang, Y.; Chen, I.M.; Chen, Z.X.; Rowley, J.D.; Willman, C.L.; et al. A previously unidentified alternatively spliced isoform of t(8;21) transcript promotes leukemogenesis. Nat. Med. 2006, 12, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Erickson, P.F.; Robinson, M.; Owens, G.; Drabkin, H.A. The ETO portion of acute myeloid-leukemia t(8-21) fusion transcript encodes a highly evolutionarily conserved, putative transcription factor. Cancer Res. 1994, 54, 1782–1786. [Google Scholar] [PubMed]
- Benhaim, M.S.; Moshitchmoshkovitz, S.; Rechavi, G. FTO: Linking m6A demethylation to adipogenesis. Cell Res. 2015, 25, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Wolford, J.K.; Prochazka, M. Structure and expression of the human MTG8/ETO gene. Gene 1998, 212, 103–109. [Google Scholar] [CrossRef]
- Galic, S.; Oakhill, J.S.; Steinberg, G.R. Adipose tissue as an endocrine organ. Mol. Cell. Endocrinol. 2010, 316, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Yamamoto, Y.; Gesta, S.; Kahn, C.R. Beneficial effects of subcutaneous fat transplantation on metabolism. Cell Metab. 2008, 7, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Rodeheffer, M.S.; Birsoy, K.; Friedman, J.M. Identification of white adipocyte progenitor cells in vivo. Cell 2008, 135, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Zeve, D.; Suh, J.M.; Bosnakovski, D.; Kyba, M.; Hammer, R.E.; Tallquist, M.D.; Graff, J.M. White fat progenitor cells reside in the adipose vasculature. Science 2008, 322, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Takakuwa, R.; Marchand, S.; Dentz, E.; Bornert, J.M.; Messaddeq, N.; Wendling, O.; Mark, M.; Desvergne, B.; Wahli, W.; et al. Peroxisome proliferator-activated receptor gamma is required in mature white and brown adipocytes for their survival in the mouse. Proc. Natl. Acad. Sci. USA 2004, 101, 4543–4547. [Google Scholar] [CrossRef] [PubMed]
- Schupp, M.; Cristancho, A.G.; Lefterova, M.I.; Hanniman, E.A.; Briggs, E.R.; Steger, D.J.; Qatanani, M.; Curtin, J.C.; Schug, J.; Ochsner, S.A.; et al. Re-expression of GATA2 cooperates with peroxisome proliferator-activated receptor-gamma depletion to revert the adipocyte phenotype. J. Biol. Chem. 2009, 284, 9458–9464. [Google Scholar] [CrossRef] [PubMed]
- Lt, V.D.L.; Smetsers, T.F.; Wittebol, S.; Raymakers, R.A.; Mensink, E.J. Molecular diversity in AML1/ETO fusion transcripts in patients with t(8;21) positive acute myeloid leukaemia. Leukemia 1994, 8, 1780–1784. [Google Scholar]
- Miyoshi, H.; Kozu, T.; Shimizu, K.; Enomoto, K.; Maseki, N.; Kaneko, Y.; Kamada, N.; Ohki, M. The t(8-21) translocation in acute myeloid-Leuk. Results in production of an AML1-MTG8 fusion transcript. EMBO J. 1993, 12, 2715–2721. [Google Scholar] [PubMed]
- Liao, K.H.; Chang, S.J.; Chang, H.C.; Chien, C.L.; Huang, T.S.; Feng, T.C.; Lin, W.W.; Shih, C.C.; Yang, M.H.; Yang, S.H.; et al. Endothelial angiogenesis is directed by RUNX1T1-regulated VEGFA, BMP4 and TGF-β2 expression. PLoS ONE 2017, 12, e0179758. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Sellayah, D. Role of FTO in adipocyte development and function: Recent insights. Int. J. Endocrinol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.F.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.Q.; Yang, Y.; Yi, C.Q.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Savaskan, N.E.; Heckel, A.; Hahnen, E.; Engelhorn, T.; Doerfler, A.; Ganslandt, O.; Nimsky, C.; Buchfelder, M.; Eyupoglu, I.Y. Small interfering RNA-mediated xCT silencing in gliomas inhibits neurodegeneration and alleviates brain edema. Nat. Med. 2008, 14, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.P.; Fan, Y.X.; Ma, T.W.; Wang, Z.; TanTai, W.J.; Nie, H.T.; Guo, Y.X.; Yu, X.Q.; Sun, L.W.; Wang, F. Carcass traits, meat quality, antioxidant status and antioxidant gene expression in muscle and liver of Hu lambs fed perilla seed. J. Anim. Physiol. Anim. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, K.; Ren, C.; Liu, Z.; Gao, X.; Fan, Y.; Zhang, G.; Zhang, Y.; MA, E.-S.; Wang, F.; You, P. Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation. Int. J. Mol. Sci. 2018, 19, 1300. https://doi.org/10.3390/ijms19051300
Deng K, Ren C, Liu Z, Gao X, Fan Y, Zhang G, Zhang Y, MA E-S, Wang F, You P. Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation. International Journal of Molecular Sciences. 2018; 19(5):1300. https://doi.org/10.3390/ijms19051300
Chicago/Turabian StyleDeng, Kaiping, Caifang Ren, Zifei Liu, Xiaoxiao Gao, Yixuan Fan, Guomin Zhang, Yanli Zhang, Ei-Samahy MA, Feng Wang, and Peihua You. 2018. "Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation" International Journal of Molecular Sciences 19, no. 5: 1300. https://doi.org/10.3390/ijms19051300