Expression, Tissue Localization and Serodiagnostic Potential of Echinococcus granulosus Leucine Aminopeptidase

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
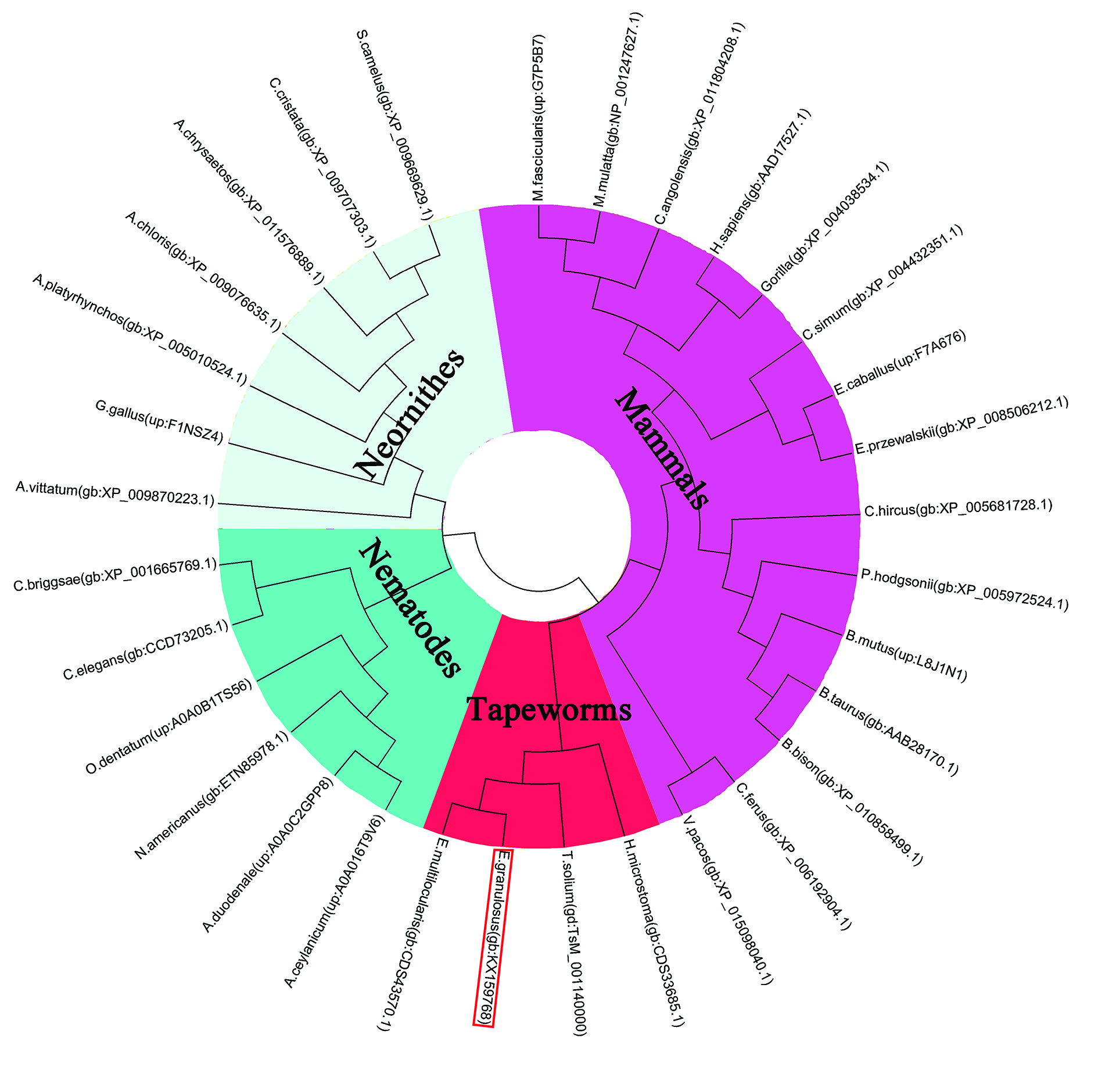
2.1. Sequence Analysis
2.2. Expression and Purification of rEg-LAP
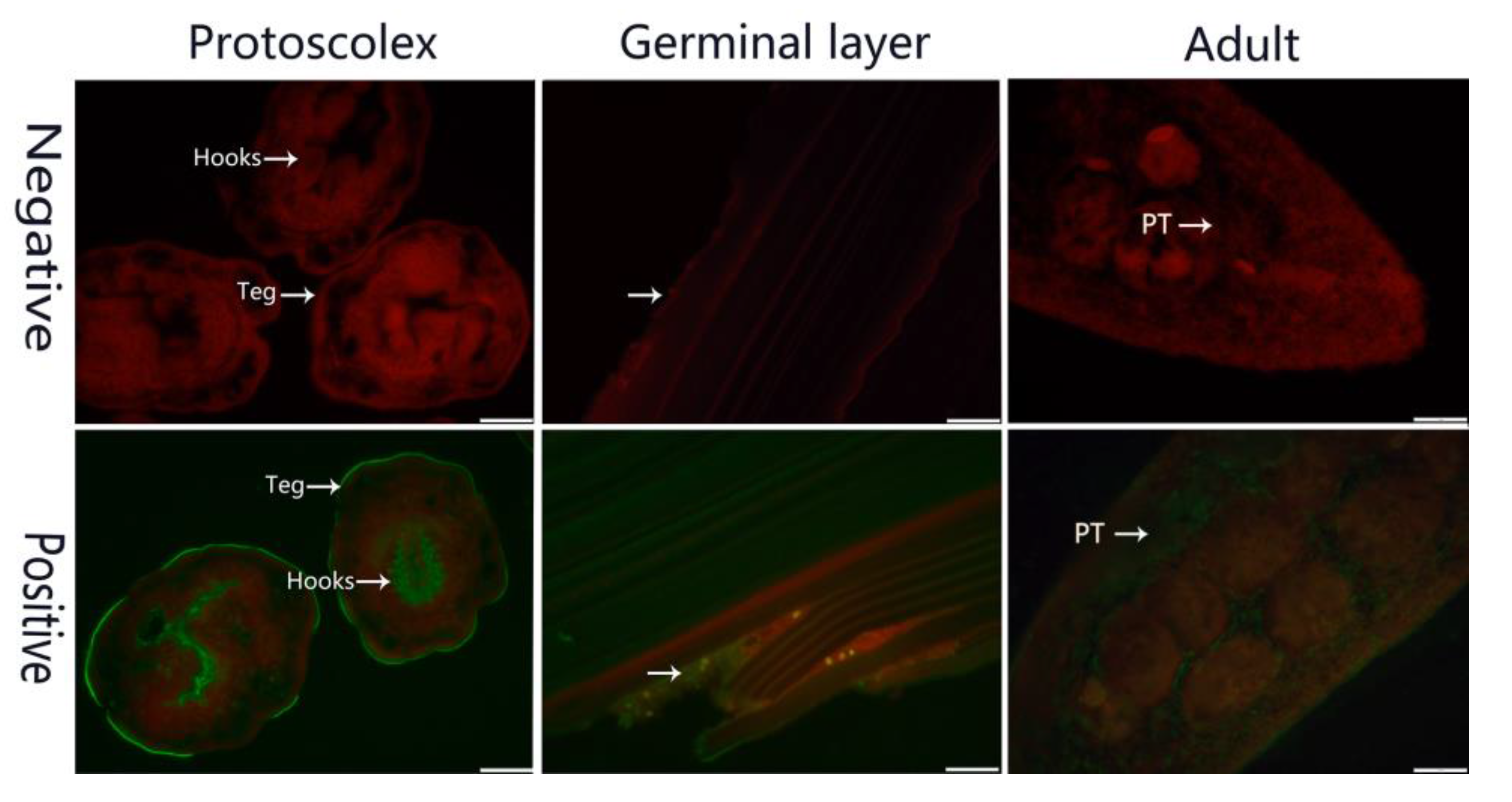
2.3. Identification in Parasites and Fluorescence Immunohistochemistry
2.4. Establishment of Indirect ELISA
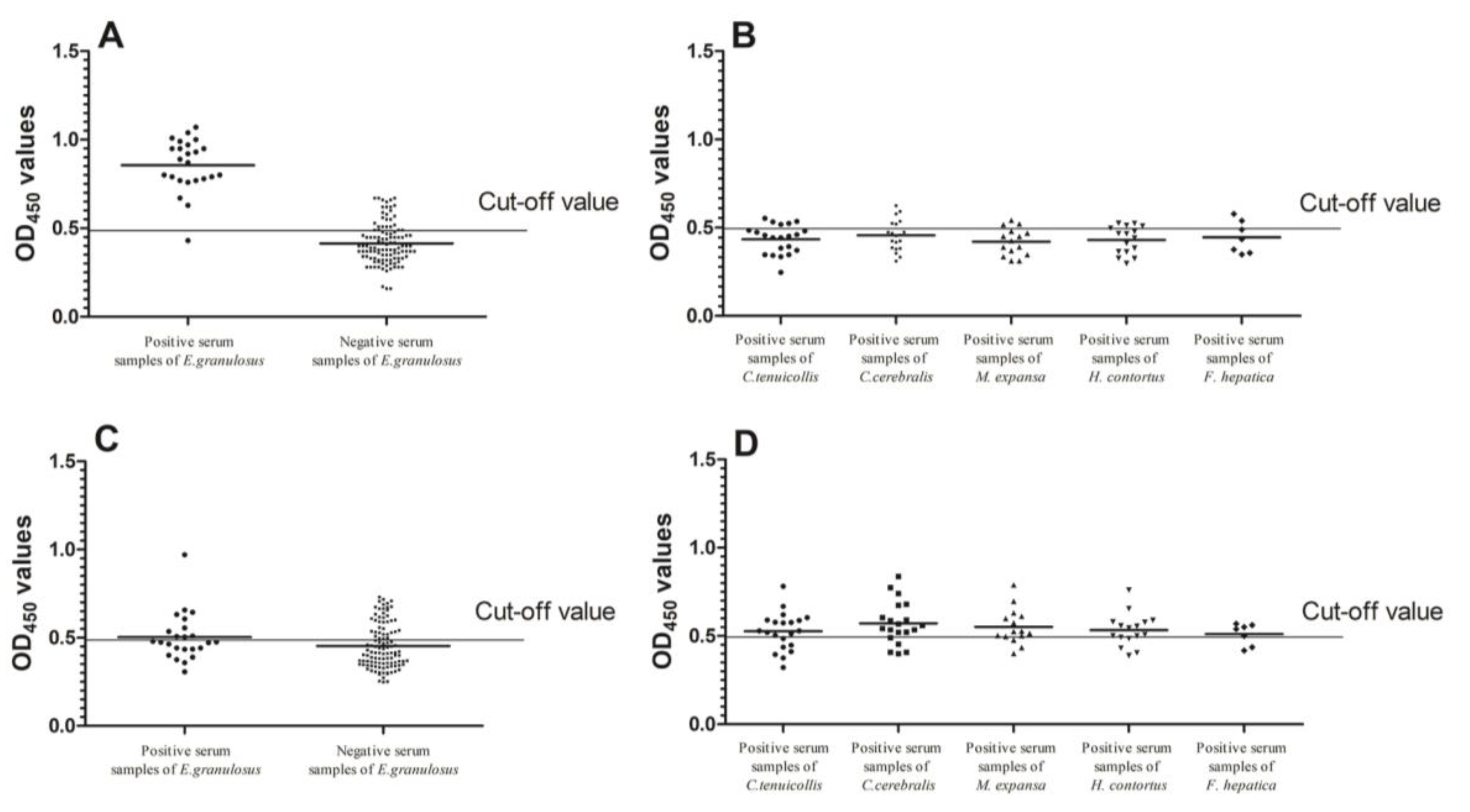
2.5. Sensitivity, Specificity and Cross Reactivity Analysis of Indirect ELISA
2.6. Reproducibility and Repeatability of Indirect ELISA
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Parasites
4.3. Bioinformatics Analysis
4.4. Expression and Purification of Recombinant EgLAP
4.5. Sera
4.6. Western Blotting (WB)
4.7. Immunolocalization
4.8. Indirect ELISA
4.9. Sensitivity, Specificity, and Cross Reactivity of Indirect ELISA
4.10. Repeatability and Reproducibility of Indirect ELISA
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| rEg-LAP | recombinant E. granulosus leucine aminopeptidase |
| CE | Cystic echinococcosis |
| PSCs | Protoscoleces |
| CV | Coefficient of variation |
| EI | E. granulosus-infected |
| CF | cestode-free |
References
- Organization, W.H. Report of the WHO Informal Working Group on Cystic and Alveolar Echninococcosis Surveillance, Prevention and Control, with the Participation of the Food and Agriculture Organization of the United Nations and the World Organisation for Animal Health, 22–23 June 2011, Department of Control of Neglected Tropical Diseases, WHO, Geneva, Switzerland; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Eckert, J.; Deplazes, P. Biological, epidemiological, and clinical aspects of echinococcosis, a zoonosis of increasing concern. Clin. Microbiol. Rev. 2004, 17, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Siracusano, A.; Delunardo, F.; Teggi, A.; Ortona, E. Host-Parasite Relationship in Cystic Echinococcosis: An Evolving Story. Clin. Dev. Immunol. 2012, 2012, 639362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Z.; Wu, W.; Shi, B.; Li, J.; Zhou, X.; Wen, H.; Mcmanus, D.P. Epidemiology and control of echinococcosis in central Asia, with particular reference to the People’s Republic of China. Acta Trop. 2015, 141, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.S.; McManus, D.P.; Lightowlers, M.W.; Chabalgoity, J.A.; Garcia, H.H.; Gavidia, C.M.; Gilman, R.H.; Gonzalez, A.E.; Lorca, M.; Naquira, C. Prevention and control of cystic echinococcosis. Lancet Infect. Dis. 2007, 7, 385–394. [Google Scholar] [CrossRef]
- Moro, P.L.; Gilman, R.H.; Verastegui, M.; Bern, C.; Silva, B.; Bonilla, J.J. Human hydatidosis in the central Andes of Peru: Evolution of the disease over 3 years. Clin. Infect. Dis. 1999, 29, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Ito, A.; Pengcuo, R.; Sako, Y.; Chen, X.; Qiu, D.; Xiao, N.; Craig, P.S. Post-treatment follow-up study of abdominal cystic echinococcosis in tibetan communities of northwest Sichuan Province, China. PLoS Negl. Trop. Dis. 2011, 5, e1364. [Google Scholar] [CrossRef] [PubMed]
- Himsworth, C.G.; Jenkins, E.; Hill, J.E.; Nsungu, M.; Ndao, M.; Thompson, R.A.; Covacin, C.; Ash, A.; Wagner, B.A.; McConnell, A. Emergence of sylvatic Echinococcus granulosus as a parasitic zoonosis of public health concern in an indigenous community in Canada. Am. J. Trop. Med. Hyg. 2010, 82, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Budke, C.M.; Deplazes, P.; Torgerson, P.R. Global socioeconomic impact of cystic echinococcosis. Emerg. Infect. Dis. 2006, 12, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Barnes, T.S.; Deplazes, P.; Gottstein, B.; Jenkins, D.; Mathis, A.; Siles-Lucas, M.; Torgerson, P.; Ziadinov, I.; Heath, D. Challenges for diagnosis and control of cystic hydatid disease. Acta Trop. 2012, 123, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kebede, N.; Mitiku, A.; Tilahun, G. Hydatidosis of slaughtered animals in Bahir Dar abattoir, northwestern Ethiopia. Trop. Anim. Health Prod. 2009, 41, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Gatti, A.; Alvarez, A.R.; Araya, D.; Mancini, S.; Herrero, E.; Santillan, G.; Larrieu, E. Ovine echinococcosis: I. Immunological diagnosis by enzyme immunoassay. Vet. Parasitol. 2007, 143, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Jeyathilakan, N.; Abdul, B.S.; John, L.; Chandran, N.D.; Raj, G.D. Development and evaluation of flow through technique for diagnosis of cystic echinococcosis in sheep. Vet. Parasitol. 2011, 180, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Jeyathilakan, N.; Basith, S.A.; John, L.; Chandran, N.D.J.N.D.; Raj, G.D.; Churchill, R.R. Evaluation of native 8 kDa antigen based three immunoassays for diagnosis of cystic echinococcosis in sheep. Small Rumin. Res. 2014, 116, 199–205. [Google Scholar] [CrossRef]
- Kittelberger, R.; Reichel, M.P.; Jenner, J.; Heath, D.D.; Lightowlers, M.W.; Moro, P.; Ibrahem, M.M.; Craig, P.S.; O’Keefe, J.S. Evaluation of three enzyme-linked immunosorbent assays (ELISAs) for the detection of serum antibodies in sheep infected with Echinococcus granulosus. Vet. Parasitol. 2002, 110, 57–76. [Google Scholar] [CrossRef]
- Simsek, S.; Koroglu, E. Evaluation of enzyme-linked immunosorbent assay (ELISA) and enzyme-linked immunoelectrotransfer blot (EITB) for immunodiagnosis of hydatid diseases in sheep. Acta Trop. 2004, 92, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh, T.; Sako, Y.; Sadjjadi, S.M.; Sarkari, B.; Ito, A. Comparison of the usefulness of hydatid cyst fluid, native antigen B and recombinant antigen B8/1 for serological diagnosis of cystic echinococcosis. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Fathi, S.; Jalousian, F.; Hosseini, S.H.; Parsa, H.; Kordafshari, S. A Study of Cross-Reactivity between Recombinant EPC1 Antigen of Echinococcus granulosus in Serum from Patients with Confirmed Cystic Echinococcosis Infection and Other Parasitic Infections. Am. J. Trop. Med. Hyg. 2016, 94, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, E.; Kern, P.; Vuitton, D.A. Expert consensus for the diagnosis and treatment of cystic and alveolar echinococcosis in humans. Acta Trop. 2010, 114, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ibrahem, M.M.; Rafiei, A.; Dar, F.K.; Azwai, S.M.; Carter, S.D.; Craig, P.S. Serodiagnosis of cystic echinococcosis in naturally infected camels. Parasitology 2002, 125 Pt 3, 245–251. [Google Scholar] [CrossRef]
- Manzanoromán, R.; Sánchezovejero, C.; Hernándezgonzález, A.; Casulli, A.; Sileslucas, M. Serological Diagnosis and Follow-Up of Human Cystic Echinococcosis: A New Hope for the Future? Biomed. Res. Int. 2015, 2015, 428205. [Google Scholar]
- Changklungmoa, N.; Kueakhai, P.; Riengrojpitak, S.; Chaithirayanon, K.; Chaichanasak, P.; Preyavichyapugdee, N.; Chantree, P.; Sansri, V.; Itagaki, T.; Sobhon, P. Immunization with recombinant leucine aminopeptidase showed protection against Fasciola gigantica in mice. Parasitol. Res. 2013, 112, 3653–3659. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Xu, Y.; Sun, H.; Lin, J.; Yu, J.; Tang, Z.; Shen, J.; Liang, C.; Li, S.; Chen, W. Systemic and local mucosal immune responses induced by orally delivered Bacillus subtilis spore expressing leucine aminopeptidase 2 of Clonorchis sinensis. Parasitol. Res. 2014, 113, 3095–3103. [Google Scholar] [CrossRef] [PubMed]
- Faustina, H.L.; Luo, Q.L.; Zhong, Z.R.; Song, X.R.; Chen, Z.W.; Wang, L.; Hu, M.M.; Chu, D.Y.; Shen, J.L. Evaluation of recombinant SjLAP and SjFBPA in detecting antibodies to Schistosoma japonicum. Chin. J. Parasitol. Parasit. Dis. 2011, 29, 339–347. [Google Scholar]
- Mahana, N.; Samy, M.A.; Diab, T.; Rabia, I.; Deeb, S.E. Immunodiagnosis of Egyptian Human Schistosomiasis Using Leucine Aminopeptidase. Glob. Vet. 2015, 14, 729–737. [Google Scholar]
- Pokharel, D.R.; Rathaur, S. Purification and characterization of a leucine aminopeptidase from the bovine filarial parasite Setaria cervi. Acta Trop. 2008, 106, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Nishikawa, Y.; Luo, Y.; Yamagishi, J.; Sugimoto, C.; Xuan, X. Characterization of a leucine aminopeptidase from Toxoplasma gondii. Mol. Biochem. Parasitol. 2010, 170, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Na, B.K.; Moon, E.K.; Song, S.M.; Joo, S.Y.; Kong, H.H.; Goo, Y.K.; Chung, D.I.; Hong, Y. Essential Role for an M17 Leucine Aminopeptidase in Encystation of Acanthamoeba castellanii. PLoS ONE 2015, 10, e0129884. [Google Scholar] [CrossRef] [PubMed]
- Joshua, G.W. Functional analysis of leucine aminopeptidase in Caenorhabditis elegans. Mol. Biochem. Parasitol. 2001, 113, 223–232. [Google Scholar] [CrossRef]
- Taylor, A. Aminopeptidases: Structure and function. FASEB J. 1993, 7, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, R.; Tomar, D.; Pandya, C.D.; Singha, R. The leucine aminopeptidase of Staphylococcus aureus is secreted and contributes to biofilm formation. Int. J. Infect. Dis. 2012, 16, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Fowler, J.H.; Walling, L.L. Leucine aminopeptidases: Diversity in structure and function. Biol. Chem. 2006, 387, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.M.; Ju, H.L.; Sohn, W.M.; Na, B.K. Molecular cloning and characterization of a M17 leucine aminopeptidase of Cryptosporidium parvum. Parasitology 2011, 138, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Sträter, N.; Lipscomb, W.N. Leucyl aminopeptidase (animal). In Handbook of Proteolytic Enzymes; Academic Press: Cambridge, MA, USA, 2004; pp. 896–901. [Google Scholar]
- Colloms, S.D. Chapter 333–Leucyl Aminopeptidase PepA. In Handbook of Proteolytic Enzymes, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1484–1492. [Google Scholar]
- Walling, L.L. Chapter 330–Leucyl Aminopeptidase (Plant). In Handbook of Proteolytic Enzymes, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1471–1476. [Google Scholar]
- Kang, J.M.; Ju, H.L.; Ju, J.W.; Sohn, W.M.; Kim, T.S.; Bahk, Y.Y.; Hong, S.J.; Na, B.K. Comparative biochemical and functional properties of two leucine aminopeptidases of Clonorchis sinensis. Mol. Biochem. Parasitol. 2012, 182, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Sun, J.; Li, X.; Wang, L.; Hu, X.; Wang, X.; Chen, W.; Lv, X.; Liang, C.; Li, W. Molecular identification and characterization of leucine aminopeptidase 2, an excretory-secretory product of Clonorchis sinensis. Mol. Biol. Rep. 2012, 39, 9817–9826. [Google Scholar] [CrossRef] [PubMed]
- Fripp, P.J. The histochemical localization of leucine aminopeptidase in Schis-tosoma rodhaini. Comp. Biochem. Physiol. 1967, 20, 307–309. [Google Scholar] [CrossRef]
- McCarthy, E.; Stack, C.; Donnelly, S.M.; Doyle, S.; Mann, V.H.; Brindley, P.J.; Stewart, M.; Day, T.A.; Maule, A.G.; Dalton, J.P. Leucine aminopeptidase of the human blood flukes, Schistosoma mansoni and Schistosoma japonicum. Int. J. Parasitol. 2004, 34, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Changklungmoa, N.; Chaithirayanon, K.; Kueakhai, P.; Meemon, K.; Riengrojpitak, S.; Sobhon, P. Molecular cloning and characterization of leucine aminopeptidase from Fasciola gigantica. Exp. Parasitol. 2012, 131, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Song, S.-M.; Park, J.-H.; Kim, J.; Kim, S.-I.; Hong, Y.-C.; Kong, H.-H.; Chung, D.-I. Identification and characterization of Paragonimus westermani leucine aminopeptidase. Parasitol. Int. 2008, 57, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Galindo, M.; Schadebrodt, G.; Galanti, N. Echinococcus granulosus: Cellular territories and morphological regions in mature protoscoleces. Exp. Parasitol. 2008, 119, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Galindo, M.; Gonzalez, M.J.; Galanti, N. Echinococcus granulosus protoscolex formation in natural infections. Biol. Res. 2002, 35, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.L.; Trenholme, K.R.; Skinner-Adams, T.S.; Stack, C.M.; Dalton, J.P. Overexpression of Leucyl Aminopeptidase in Plasmodium falciparum Parasites target for the antimalarial activity of bestatin. J. Biol. Chem. 2006, 281, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.R.; Zhou, H.B.; Li, X.Y.; Luo, Q.L.; Song, X.R.; Wang, W.; Wen, H.Q.; Yu, L.; Wei, W.; Shen, J.L. Serological proteome-oriented screening and application of antigens for the diagnosis of Schistosomiasis japonica. Acta Trop. 2010, 116, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, N.; Hu, D.; Zhong, X.; Wang, S.; Gu, X.; Peng, X.; Yang, G. Genetic diversity of Echinococcus granulosus in Southwest China determined by the mitochondrial NADH dehydrogenase subunit 2 gene. Sci. World J. 2014, 2014, 867839. [Google Scholar]
- Zhong, X.; Song, X.; Wang, N.; Hu, D.; Liu, T.; Wang, T.; Gu, X.; Lai, W.; Peng, X.; Yang, G. Molecular identification and characterization of prohibitin from Echinococcus granulosus. Parasitol. Res. 2015, 115, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, L.; Yang, Y.; Gu, X.; Wang, Y.; Lai, W.; Peng, X.; Yang, G. Expression, tissue localization and serodiagnostic potential of Taenia multiceps acidic ribosomal protein P2. Parasites Vectors 2015, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Yan, M.; Hu, D.; Wang, Y.; Wang, N.; Gu, X.; Peng, X.; Yang, G. Molecular characterization and serodiagnostic potential of a novel dithiol glutaredoxin 1 fromEchinococcus granulosus. Parasites Vectors 2016, 9, 456. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Jia, R.; Zhang, Z.; Wang, M.; Xu, Y.; Zhu, D.; Chen, S.; Liu, M.; Yin, Z.; Chen, X. In vitro expression and development of indirect ELISA for Capsid protein of duck circovirus without nuclear localization signal. Int. J. Clin. Exp. Pathol. 2014, 7, 4938–4944. [Google Scholar] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.; Yan, M.; Xu, J.; Liang, Y.; Gu, X.; Xie, Y.; Jing, B.; Lai, W.; Peng, X.; Yang, G. Expression, Tissue Localization and Serodiagnostic Potential of Echinococcus granulosus Leucine Aminopeptidase. Int. J. Mol. Sci. 2018, 19, 1063. https://doi.org/10.3390/ijms19041063
Wu M, Yan M, Xu J, Liang Y, Gu X, Xie Y, Jing B, Lai W, Peng X, Yang G. Expression, Tissue Localization and Serodiagnostic Potential of Echinococcus granulosus Leucine Aminopeptidase. International Journal of Molecular Sciences. 2018; 19(4):1063. https://doi.org/10.3390/ijms19041063
Chicago/Turabian StyleWu, Maodi, Min Yan, Jing Xu, Yuqing Liang, Xiaobin Gu, Yue Xie, Bo Jing, Weimin Lai, Xuerong Peng, and Guangyou Yang. 2018. "Expression, Tissue Localization and Serodiagnostic Potential of Echinococcus granulosus Leucine Aminopeptidase" International Journal of Molecular Sciences 19, no. 4: 1063. https://doi.org/10.3390/ijms19041063
APA StyleWu, M., Yan, M., Xu, J., Liang, Y., Gu, X., Xie, Y., Jing, B., Lai, W., Peng, X., & Yang, G. (2018). Expression, Tissue Localization and Serodiagnostic Potential of Echinococcus granulosus Leucine Aminopeptidase. International Journal of Molecular Sciences, 19(4), 1063. https://doi.org/10.3390/ijms19041063