1. Introduction
Thrombotic disorders, including ischemic stroke, unstable angina, pulmonary embolism (PE), deep venous thrombosis (DVT) and myocardial infarction, are major causes of morbidity and mortality worldwide [
1]. A number of antithrombotic agents, which include anticoagulant, antiplatelet, and fibrinolytic agents, are available for the treatment and prevention of thrombosis. Warfarin is the first widely used anticoagulant shortly followed by heparin. Heparin, which is the second-most used naturally occurring drug in medicine, is widely applied in clinical for a long term. But, the long-standing use of anticoagulant can cause a number of side effects, such as bleeding, thrombocytopenia, changes in lipid metabolism, and osteoporosis [
2,
3,
4]. Subsequently, the antithrombotic agents, such as nattokinase, streptokinase, urokinase, and t-PA are exploited for treatment of thrombosis and they show significant therapeutic effects [
5,
6]. However, these antithrombotic agents are very expensive and still present some inescapable side effects [
5,
6,
7]. Hence, the existing antithrombotic agents cannot satisfy the requirements of patients. Due to the value of antithrombotic agents, the search for other antithrombotic agents has been implemented from various sources without interruption since the 1990s. Increasing studies indicate that many new thrombolytic agents are identified and characterized from natural resources, such as
Eisenia foetida, snake venoms,
Neanthes japonica,
Urechis unicinctus, and so on [
8,
9,
10].
Sipunculus nudus, an old marine species, approximately emerged in Middle Paleozoic Era and even Cambrian Period.
S. nudus belongs to the
Sipuncula phylum and it has a wide distribution along the coast of Indian Ocean and Western Pacific [
11].
S. nudus is an edible marine organism that is popular for its delicious taste in southeast China for many years. With the help of modern technology, it is found that
S. nudus has great potential for use as functional seafood due to its various bioactivities, such as anti-thrombus [
12], anti-hypoxic, anti-fatigue [
13], immune modulatory [
14], antimicrobial, and antioxidant activities [
15,
16]. Recently, the potential antithrombotic activity of
S. nudus has attracted considerable attention, as it is considered to be rich in free amino acids, polypeptides, and proteins. By the inspiration of obtaining multiple fibrinoclase fromnatural resources as well as consideration of close affinity among
Sipuncula,
Annelida, and
Echiura [
17], the antithrombotic activity of aqueous extract from
S. nudus was evaluated in our previous work, and the results indicated that the crude protein extract of
S. nudus had strong activities of fibrinolysis, anti-coagulating, inhibiting platelet aggregation, as well as scavenging oxygen free radical, and so on [
12]. Accordingly, to find out the bio-active ingredients, the present study reported the isolation, purification, and characterization of a novel protease named SK (the kinase obtained from
S. nudus) from
S. nudus and its antithrombotic effects, as well as the related mechanism.
3. Discussion
Authors should discuss the results and how they can be interpreted in perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
In our study, after an efficient extract method, a fast and gentle (NH4)2SO4 graded precipitation method was applied to remove fat-soluble and other water-soluble constituents, including some protein, which could not only be beneficial to concentrate the extract volume, but also observably maintain the activity of SK.
Then in the following process of purification, a successional method involving a hydrophobic interaction chromatography, an ion exchange chromatography, and a gel filtration chromatography was applied and optimized in our study. Normally, according to Hofmeister series, (NH
4)
2SO
4 was always chosen as a high salt phase in hydrophobic interaction separation. At high concentrations of (NH
4)
2SO
4, hydrophobic ligands combined most of protein in column and removed other constituents of non-protein. Meanwhile, when the ratio of (NH
4)
2SO
4 went down, the protein was eluted successively due to degressive interaction [
19]. In this experiment, a Phenyl Sepharose High Performance column showed strong interaction with protein in condition of 1 mol/L (NH
4)
2SO
4. Besides, we were fortunate enough to attain high recovery of SK since SK could be easily eluted from this column with deionized water, which also provided the possibility of high efficient recycling use of this column.
As a special biological macromolecule, protein is charged and is differently depended on its own isoelectric point (pI) and its surrounding pH. Based on this property, ion exchange chromatography is commonly used in protein purification based on the interaction of ion exchange ligands and target protein. According to law of chromatographic action, ion exchanger loads more protein under lower concentrate of saltiness, as the salinity rises, the binding protein is also eluted with the binding sites decreases [
20,
21]. The column of ion exchanger is employed to purify target protein on Q Sepharose High Performance. Proper pH is important parameter for purifying the charge of protein. Different pH values between 6.5 and 9.5 were investigated in preliminary tests. Consequently, the closest combination of protein and anion exchanger occurred at pH 8.5. Unfortunately, some chromoprotein in loading sample was always absorbed while using Q Sepharose High Performance as strong anion exchanger, which was not easily cleaned up in ion exchange chromatography. To overcome these defects, a flushing combination of 2 mol/L NaCl and 1 mol/L NaOH was used for making empty sites at one sample injection interval.
In the purification of protein, gel filtration chromatography exhibited irreplaceable value based on molecular size as well as mild separation conditions [
22]. In this study, a Superdex G-75 gel filtration column with molecular range of 3000–70,000 Da was applied in the last step, which was used to further purify the protein. Besides during the purification, 3500 Da dialysis bags, Zeba spin desalting columns, and Millipore amicon ultra 3K ultrafiltration were also used for desalting, purification, and concentration of samples.
The protein contents and total enzyme activities of obtained fractions were determined for calculation of specific activity, yield and purity. The yield of fractions (total enzyme activity after each step/total enzyme activity after extraction × 100%) collected after all the steps utilized in our experiments was 14.47%, and the purification fold (specific activity after each step/specific activity after extraction × 100%) was 2466.62 (
Table S9). Despite that we got relative high purity of SK in this study, we still have to admit that as process goes in preparation procedure, total activity of SK inevitably decreases. Thus, we still need to seek much more simplified and quick methods in future study if we aim at studying the activity and its mechanism of SK.
After the identification of purity by the methods of Native PAGE, SDS-PAGE and gel filtration chromatography, SK was further detected by MALDI-TOF for molecular weight determination. The molecular weight of SK was 28.0kDa, which was similar to many other fibrinolytic proteases, including nattokinase (27.3 kDa), urokinase (34.0 kDa), and the lumbrukinase in Eisenia foetida (24.0 kDa and 34.0 kDa) and Neanthes japonica (33.5 kDa) with anticoagulative activity [
23]. It also came to reasonable agreement with the results determined by SDS–PAGE.
Because the biosynthesis of protein was normally started from N terminal, the posttranslational modification of N terminal usually possessed close relationship with its biological functions and stabilities [
24]. Therefore, N terminal determination analysis was helpful for further analysis of higher spatial structures and for revealing functional mechanism of protein. Edman degradation method is a powerful method that is widely used in N-terminal amino acids sequencing [
25]. With the help of Edman degradation, the 15 N-terminal amino acids sequence was PFPVPDPFVWDTSFQ, which showed 84% of identities with hemerythrin in
S. nudus. When compared with many other peptides with antithrombotic activity or fibrinolytic activity, it was found that SK was a novel active peptide from the aspect of the N terminal amino acids including urokinase, lumbrukinase, nattokinase, streptokinase and other fibrinolytic enzymes (
Table S10).
It was crucial to develop the research on the enzymatic properties for further intensive study and reasonable utilization on SK. The properties of SK showed the activity difference of SK had no significant differences and pH value, which meant that SK was a thermostable protease that is resistant to acids and alkalis in some degree. The effects on activities of SK by different metal ions showed not significantly different as well, however, according to obvious inhibitory effects by PMSF and aprotinin when compared with leupeptin and pepstatin, we could come to a conclusion that SK typically belonged to serine protease category.
The studies of in vitro fibrinolytic activities of SK showed that SK not only presented direct effects on fibrinogen, but also had indirect effects on fibrinogen by activating plasminogen. According to the result of SDS–PAGE, it was found that the fibrinolytic effects of SK were established by degrading chain α, chain β, and chain γ in fibrinogen to inhibit the form of fibrin. Hence, SK was a plasminogen activator with the capacity of direct fibrinolysis.
The thrombosis was usually formed from coagulation disorders that were induced by vascular endothelial cells injury or platelet aggregation as well as changes on the hemorheology [
26]. To simulate the properties of thrombosis as much as possible, the FeCl
3-induced common carotid artery thrombosis model with easy operation and highly successful rate was applied in our experiment. This method was widely used in many researches in regard to the occurrence and development of thrombosis as well as exploitation of therapy methods and drugs [
27,
28]. The idea was that when FeCl
3 stimulated the vascular wall, the oxidative stress reacted by free radicals led to a series of reactions, including the lipid peroxidation, endothelial injury, platelet adhesion and aggregation, and finally the occluded thrombus [
29]. During the formation of thrombus, a series of endogenous regulatory factors were also involved in the regulation. TXA
2 and PGI
2 were two important regulation substances from arachidonic acid (AA) metabolism with contrary function: TXA
2 presented certain effects of contracting the vascular and of promoting the platelet to aggregate, adhere and release, while PGI
2 was regarded as a powerful active substance with an antithrombotic effect, which had the ability of dilating the blood vessels, inhibiting the platelet aggregation, promoting fibrinolytic activity, and so on. These two substances were unstable in vivo and could be easily metabolized into TXB
2 and 6-keto-PGF
1α, respectively. Hence, these two metabolites were detected to assess the contents of TXA
2 and PGI
2 in most cases [
30]. Tissue type plasminogen activator (t-PA) and urokinase type plasminogen activator (u-PA) could activate PLG into plasmin, which inhibited thrombus by catalyzing fibrin to degrade into fibrin degradation products (FDP). PAI-1 was the substances physiologically inhibit t-PA and u-PA [
31]. Besides endothelin (ET-1) lying in vascular endothelial cells also had vital function in regulating cardiovascular efficiency and in maintaining basic vascular tension and cardiovascular homeostasis. As an endogenously long-acting vasoconstrictor, it kept long action without antagonism by α receptor antagonists, H1 receptor antagonists, and 5-HT receptor antagonists [
32]. Finally, the CGRP was proved to be a bioactive polypeptide that regulated the cardiovascular system by relaxing blood vessel, strengthening myocardiac contraction, accelerating heart rate, dilating coronary artery, and so on [
33].
By the measurement of weight of thrombus and observation of histomorphology, we found that SK had a significant therapeutic effect on carotid arterial thrombosis in rats, and SK got a distinctly better effect than that of urokinase in most cases. The exploration of underlying mechanisms revealed that the antithrombotic effect of SK, which was represented by inhibition of FeCl3-induced thrombosis in the common carotid arterial was related to the regulation of vascular, fibrinolytic, and coagulation systems.
Based on the mechanism study by in vivo experiments, the underlying mechanisms of SK on FeCl3-induced common carotid artery thrombosis in rats were further studied by untargeted mass spectrometry-based metabolomics profiling. Three classical metabolism pathways were discovered and responsible to illustrate the effect mechanisms.
AA metabolism pathway was an important metabolic pathway in occurrence and development of thrombotic disease. It acted as a metabolic substrate and also an important precursor of several bioactive substances [
34]. During the process of thrombus development, the vascular endothelial cells promoted the conjunction type of AA to break into free AA by phospholipase A
2 (PLA
2) and released them out. Then, under the reducing action of the cyclo-oxygenase (CO), some free AA turned into prostaglandin G
2 (PGG
2) and further turned into prostaglandin H
2 (PGH
2). PGG
2 and PGH
2 were particularly unstable intermediate products, and they will further turn into other prostaglandins and thromboxanes, such as TXA
2 and PGI
2 [
35]. Some free AAs turned into 5-HPETE by double oxidation of 5-lipoxygenase (5-LOX), and some of 5-HPETE further turned into 5-HETE by 5-LOX, while the other turned into leukotriene A
4 (LTA
4). LTA4 was an unstable intermediate of monooxygenase, and some LTA
4 turned into leukotriene B
4 (LTB
4) by adding H
2O or turned into leukotriene C
4 (LTC
4) by glutathione S-transferase. LTC
4 possessed the certain effects of a contracting blood vessel, which was equal to angiotensin and even stronger than LTD
4 and LTE
4 [
36,
37]. In the current study, the contents of AA, LTC
4, TXA
2 and TXB
2 were enhanced in FeCl
3-induced common carotid arterial thrombosis rats. The results indicated that thrombus was easily excitated when the contents of AA, LTC
4, TXA
2, and TXB
2 were raised in organism. However, these endogenous substances (such as AA, LTC
4, TXA
2, and TXB
2) were significantly improved in model rats with the administration of SK. Based on the method of metabolism, SK exerted a key role in inhibiting vessel contraction, platelet aggregation, adhesion, and release by adjusting the AA metabolism pathway (
Figure S6).
Recently, more and more researches focused on the disorders of sphingolipid metabolism, which leads to a disorder of signal transduction and even a series of metabolic diseases. In sphingolipid metabolism pathway, ceramides were the center substances of sphingolipid biosynthesis, degradation, and metabolism, as well as the general precursors of further sphingolipid metabolism [
38]. Some studies have found that the contents of ceramide rose significantly in the endothelial cell dysfunction, and ceramides might mediate the metabolic dysfunction of endothelial cells [
39]. There were also some studies discovered in the case of vascular injury, a vascular endothelial growth factor (VEGF) that is specific to vascular endothelial cells would mediate angiogenesis, and it also leads to a sharp rise in glycosphingolipid (a combination of ceramides and carbohydrate chain). In the latter studies, although the specific mechanism is not fully clear, some hypotheses suggested that sphingolipid metabolism might be involved in the regulation of VEGF [
40,
41]. All of the above results indicated that ceramides accumulation was closely related to endothelial cell dysfunction. In our research, the marked increase of dihydroceramide, which was an important precursor of ceramides, indicated that SK might play positive regulation on endothelial cell dysfunction by compensating the contents of dihydroceramide (
Figure S7).
In the process of thrombosis, as oxidization-reduction coenzymes, nicotinamide adenine dinucleotides (NAD) and nicotinamide adenine dinucleotide phosphate (NADP) were usually served as electron acceptors and hydrogen donor in cells. Since they were synthesized by nicotinamides and ribose-adenine RNA, this reaction also caused the compensatory increases of nicotinic acids and nicotinamides [
42]. When compared with the common carotid arterial thrombosis model group, the contents of niacinamide, which could be metabolized by NAD, NADP, nicotinamide ribotide, and nicotinamide riboside notably decreased in the SK group, which could imply that the process of thrombosis could be markedly slowed down with the function of SK (
Figure S8).
The above results suggested that AA, sphingolipid, nicotinate, and nicotinamide mechanism pathways have close relationship with thrombus development. SK could significantly improve the metabolic abnormalities in common carotid arterial thrombosis of rats. The untargeted mass spectrometry-based metabolomics profiling of the thrombosis feature and underlying mechanisms of SK on thrombosis were firstly constructed. This study provided a systematic view of the development and progress of common carotid arterial thrombosis, and it also offered the application of evaluating the mechanisms of SK on common carotid arterial thrombosis induced by FeCl3 and other thrombotic diseases.
In summary, a set of methods was first established in the study, including freeze-thaw coupled with homogenate, (NH4)2SO4 graded precipitation, and combination of hydrophobic interaction chromatography, anion-exchange chromatography, and gel filtration chromatography for extraction, isolation, and purification of the thrombolytic protease in S. nudus based on the fibrinolytic activities tracking by fibrin plate assay. This chromatographically and electrophoretically pure peptide with a molecular weight of 28,003.67 Da and a 15 N-terminal amino acids sequence of PFPVPDPFVWDTSFQ was found to be a novel active peptide when comparing with other antithrombotic and fibrinolytic peptides.
4. Materials and Methods
4.1. Sources of S. Nudus Samples
S. nudus samples were collected and identified by Shao-Xiong Ding (Xiamen University, Xiamen, China). The samples were kept in −80 °C refrigerators after collection.
4.2. Animals
Forty-eight male Sprague-Dawley (SD) rats (220–250 g) were obtained from Experimental Animal Center of Zhejiang Province (SCXK (2016-0001), 10 January 2016). The rats were kept in plastic cages with fresh water and pellet food. The environmental temperature and humidity were respectively maintained at 24 ± 2 °C and 40–60%. Animal welfare and experimental procedures were carried out in accordance with the guide for care and use of laboratory animals. The ethic approval number was ACU-25 (20161229).and the approval date was 29 December 2016.
4.3. Sample Preparation
Fresh samples (800 g) were carefully dissected and removed excreta and 1600 mL of PBS was added for homogenization (1000 r/min, 30 s). Then, the homogenate was extracted by freezing and thawing twice. Finally, extract of S. nudus was obtained by collecting and mixing two-step supernatant.
4.4. Protease Isolation and Purification
The best range of (NH
4)
2SO
4 percent saturation was chosen for preparing SK according to fibrinolytic activities that were determined by fibrin plate assay. The details of choosing best range of (NH
4)
2SO
4 was described in “1. Option of best best range of (NH
4)
2SO
4 in precipitation” in
Supplementary Materials.
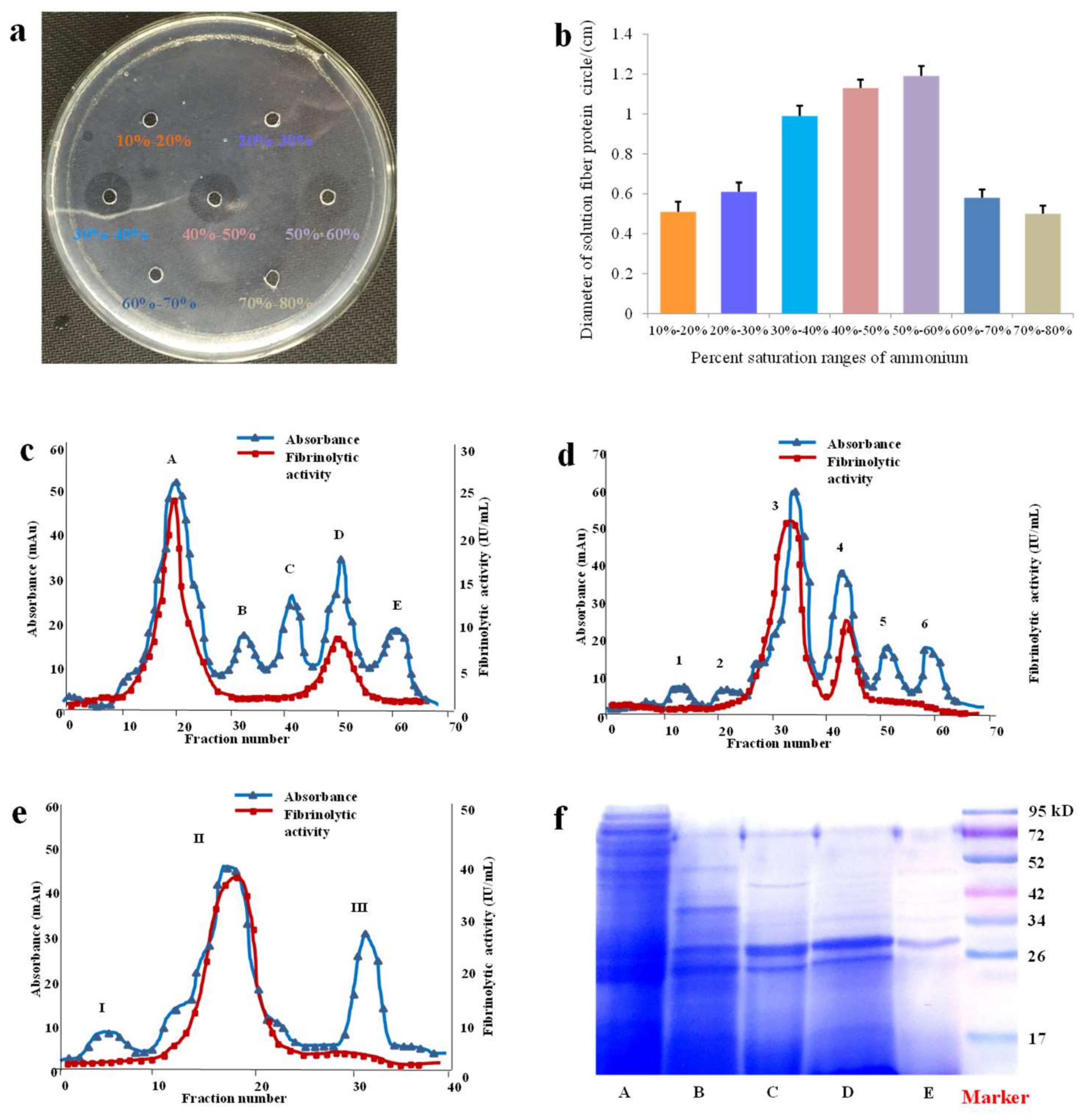
ÄKTA™ pure system (GE Health, Chicago, MA, USA) was employed to purify SK. SK was first isolated with hydrophobic interaction chromatography by fractionating extract through Phenyl Sepharose High Performance column (26 mm × 170 mm, GE Healthcare, Chicago, MA, USA). The mobile phase A and mobile phase B were PBS buffer and 1 mol/L (NH4)2SO4 solution. The flow rate was 3 mL/min. The linear gradient conditions were: 100% B, 3CV; 100–60% B, 0.5 CV; 60% B, 0.75 CV; 60–40% B, 0.25 CV; 40% B, 0.75 CV; 40–20% B, 0.25 CV; 20% B, 0.75 CV; 20–0% B, 0.25 CV; 0% B, 2 CV. The collected volume was set at 9 mL per fraction and the fibrinolytic activity to protein content ratio (FA/PC) of each fraction was monitored by UV detector at the wavelength of 280 nm. The collected fractions with high value of FA/PC were pooled and then desalted.
The sample purified with above method was loaded onto a Q Sepharose High Performance ion-exchange column (26 mm × 180 mm, GE Healthcare) with a gradient elution by phase A (PBS buffer) and phase B (0.8 mol/L NaCl). The flow rate was set at 3 mL/min and the system was eluted as following: 0% B, 2 CV; 0−20% B, 0.25 CV; 20% B, 2 CV; 20−30% B, 0.25 CV; 30% B, 1 CV; 30−50% B, 0.25 CV; 50% B, 1 CV; 50−70% B, 0.25 CV; 70% B, 0.75 CV; 70−100% B, 0.25 CV; 100% B, 2 CV. The samples were collected with the same method described above.
After the above process of isolation and purification, the sample was ultimately loaded onto the gel filtration column of Superdex prep grade G-75 (16 mm × 500 mm, GE Healthcare), equilibrated with PBS buffer of an isocratic elution at a flow rate of 0.8 mL/min. Finally, the protease SK was obtained followed by the collection method described above.
4.5. Identification of Purity, Determination of Molecular Weight and Analysis of N-terminal Amino Acids Sequence
SDS–PAGE, Native–PAGE and gel filtration chromatography were employed to identify the purity of SK and the methods were displayed in items “SDS-PAGE and Native PAGE of SK” and “Gel filtration chromatography of SK” in supporting information. Besides, molecular weight and N-terminal amino acids sequence of SK was respectively determined and analysed as the details in items “Determination of molecular weight” and “Analysis of N-terminal amino acids”.
4.6. Enzymatic Properties
In this experiment, the enzymatic properties of SK were studied from four aspects, including temperatures, pH values, metal ions, and inhibitors. The effects of temperatures on SK were studied by determining the residual activities by fibrin plate method after SK (20 μg/mL) pre-incubated at different temperatures (10, 20, 30, 40, 50, and 60 °C) for 24 h. The effects of pH on SK were researched by determining the residual activities after SK was being dissolved in different buffer solution with a series of pH values (pH = 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, and 11.0). In order to explore the effects of metal ions (K+, Na+, Mg2+, Ca2+, Cu2+, Fe2+, Zn2+, Al3+, Fe3+, and Mn2+) and protease inhibitors (ethylenediamine-tetraacetic acid (EDTA), phenyl methyl sulfonyl fluoride (PMSF), aprotinin, leupeptin, and pepstatin A) on SK, it was pre-incubated with different metal ions and different protease inhibitors respectively, and then the residual activities were determined for evaluating SK properties.
4.7. Fibrinolytic Activity and Fibrinogenolytic Activity
To study the fibrinolytic activity of SK on the degradation of fibrin, the fibrinolytic activities of SK in different concentrations (100.00, 50.00, 25.00, 12.50, 6.25, 3.06, and 1.53 μg/mL) were respectively determined by fibrin plate assay.
For further exploration of the fibrinolytic mechanism of SK, one fibrin plate was heated at 85 °C for 30 min in advance to inactivate plasminogen, and another plate without any treatment was served as control. Aliquots (10 μL) of 2000 IU/mL urokinase and 10 μg/mL SK were added into two plates, respectively.
Fibrinogenolytic mechanism of SK was further studied based on the above researches. Ten aliquots (200 μL) of 1% fibrinogen were mixed with 50 μL of 10 μg/mL SK and then the reaction solutions were respectively incubated at 37 °C for 0, 5, 10, 30 min, 1, 2, 4, 6, 8, and 16 h. After cooling down, aliquots of the reaction solutions (20 μL) were loaded onto SDS-PAGE consisting of 12% separation gel and 5% stacking gel for analysis.
4.8. Antithrombotic Effect on FeCl3-Induced Common Carotid Arterial Thrombosis in Rats
After one week of acclimatization, 48 rats were randomly divided into six groups: sham-operated group (C), FeCl3-induced common carotid arterial thrombosis model group (M), urokinase group (UK, 2000 IU/kg), high-dose SK group (SKH, 100 μg/kg), middle-dose SK group (SKM, 10 μg/kg), and low-dose SK group (SKL, 1 μg/kg). The rats in groups UK, SKH, SKM and SKL were injected with corresponding medication by caudal vein continuously for seven days; meanwhile, the groups C and M were injected with sterile water. On the seventh day, the FeCl3-induced common carotid arterial thrombosis was modeled in rats in groups UK, SK, and M (see item “Establishment of FeCl3-induced common carotid arterial thrombosis” in supporting information) and rats in group C was set as control. Then the blood samples (approximate 8 mL) were collected by abdominal aorta intubation approach in rats and divided into two parts. One part of 6 mL blood (3.8% sodium citrate as anticoagulant) was centrifuged (3000 r/min, 10 min) to get plasma for four coagulation tests (activation part thrombin time (APTT), thrombin time (TT), prothrombin time (PT), fibrinogen (FIB)), and four Elisa kit tests (thromboxane B2 (TXB2), plasminogen activator inhibitor-1 (PAI-1), plasminogen (PLG), and tissue t-PA. Another part of 2 mL blood (1.5% EDTA as anticoagulant) was centrifuged (3000 r/min, 10 min) to get plasma for five Elisa kit tests (calcitonin gene-related peptide (CGRP), endothelin-1 (ET-1), fibrin degradation products (FDP), 6-keto-prostaglandin F1α (6-keto-PGF1α), and prostacyclin I2 (PGI2). The arterial thrombus on the right artery (A) as well as segments with equal length (B) on the left artery of all rats executed were quickly cut off and respectively weighed for calculating thrombus weight (A–B). After weighing, the arterial thrombus was fixed in formalin and was embedded by paraffin for cutting sections and HE staining.
4.9. Mechanism of Effect on FeCl3-Induced Common Carotid Arterial Thrombosis Based on Metabonomics
To further investigate the mechanism of effect, an untargeted metabonomics method was established. The detail method was described in
Supplementary Materials.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





