
The Role of Phytochemicals in the Inflammatory Phase of Wound Healing


Abstract
:1. Introduction
2. The Role of Inflammation in Wound Healing
2.1. Acute Wounds
2.2. Excessive Healing
2.3. Deficient Healing: Non-Healing Wounds
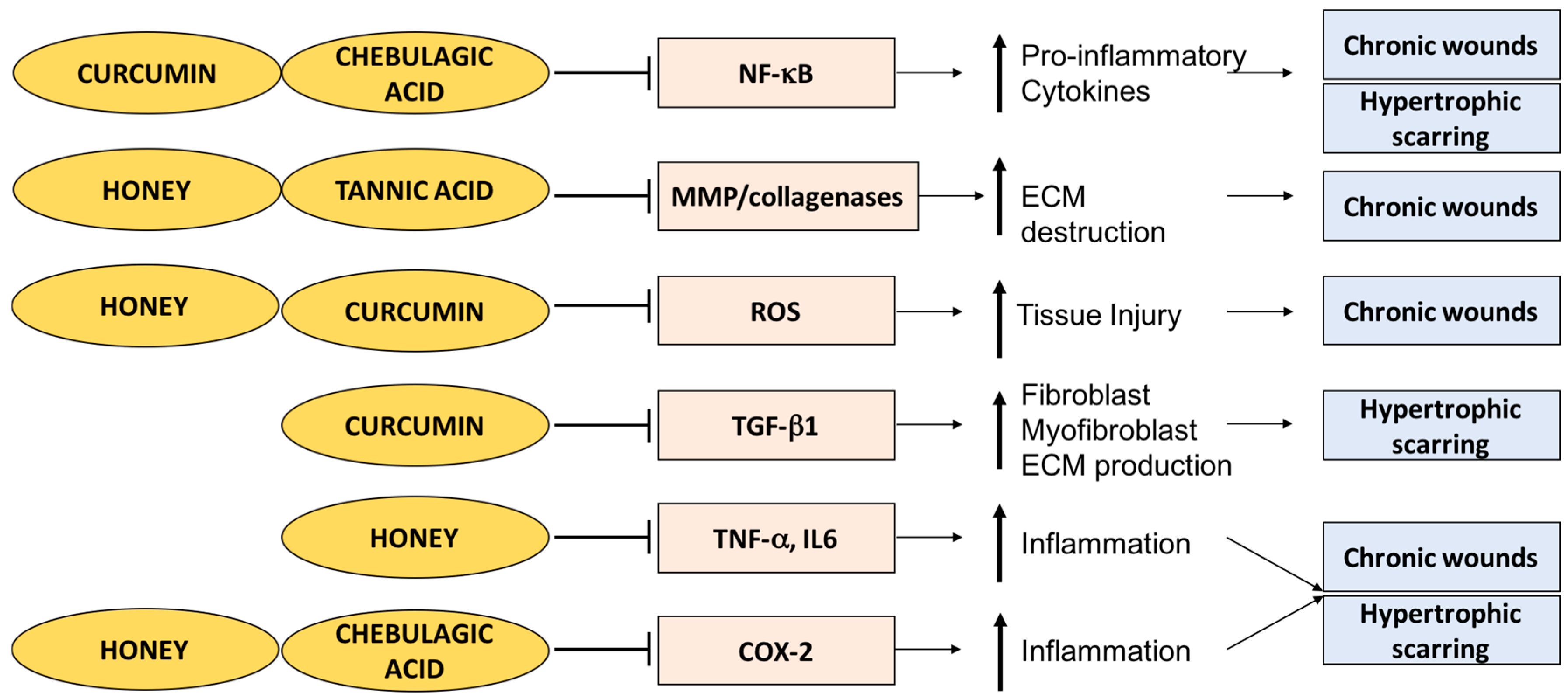
3. Phytochemicals and Traditional Therapies in Wound Healing
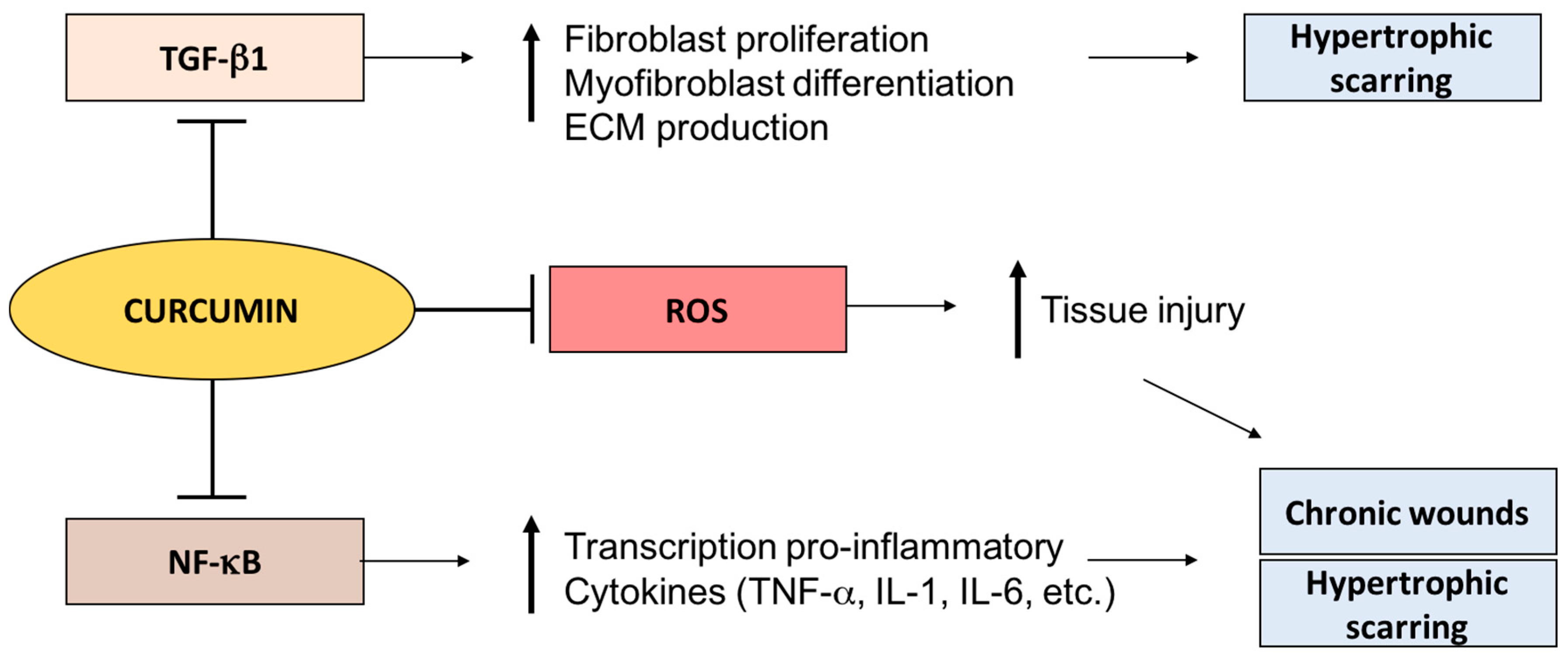
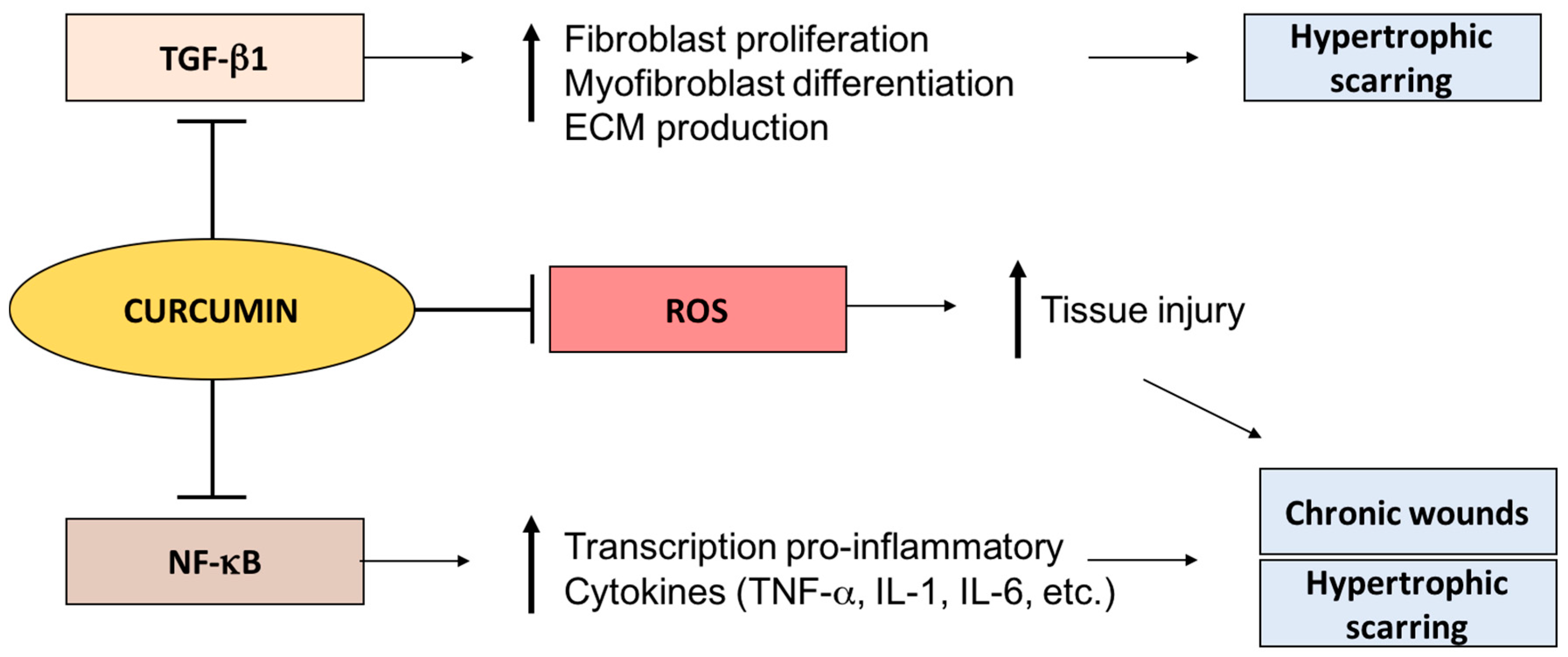
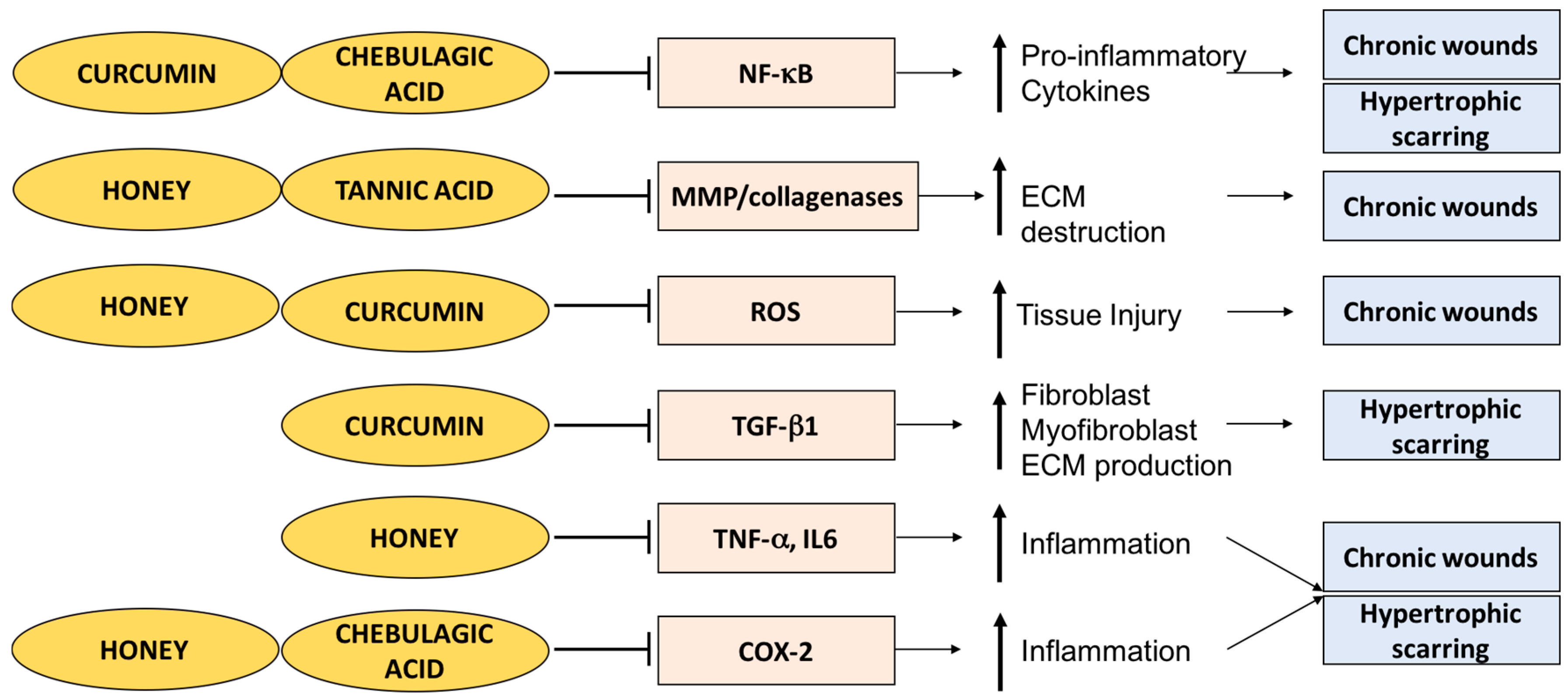
3.1. Curcumin
3.1.1. Curcumin and Chronic Wounds
3.1.2. Curcumin and Hypertrophic Scarring
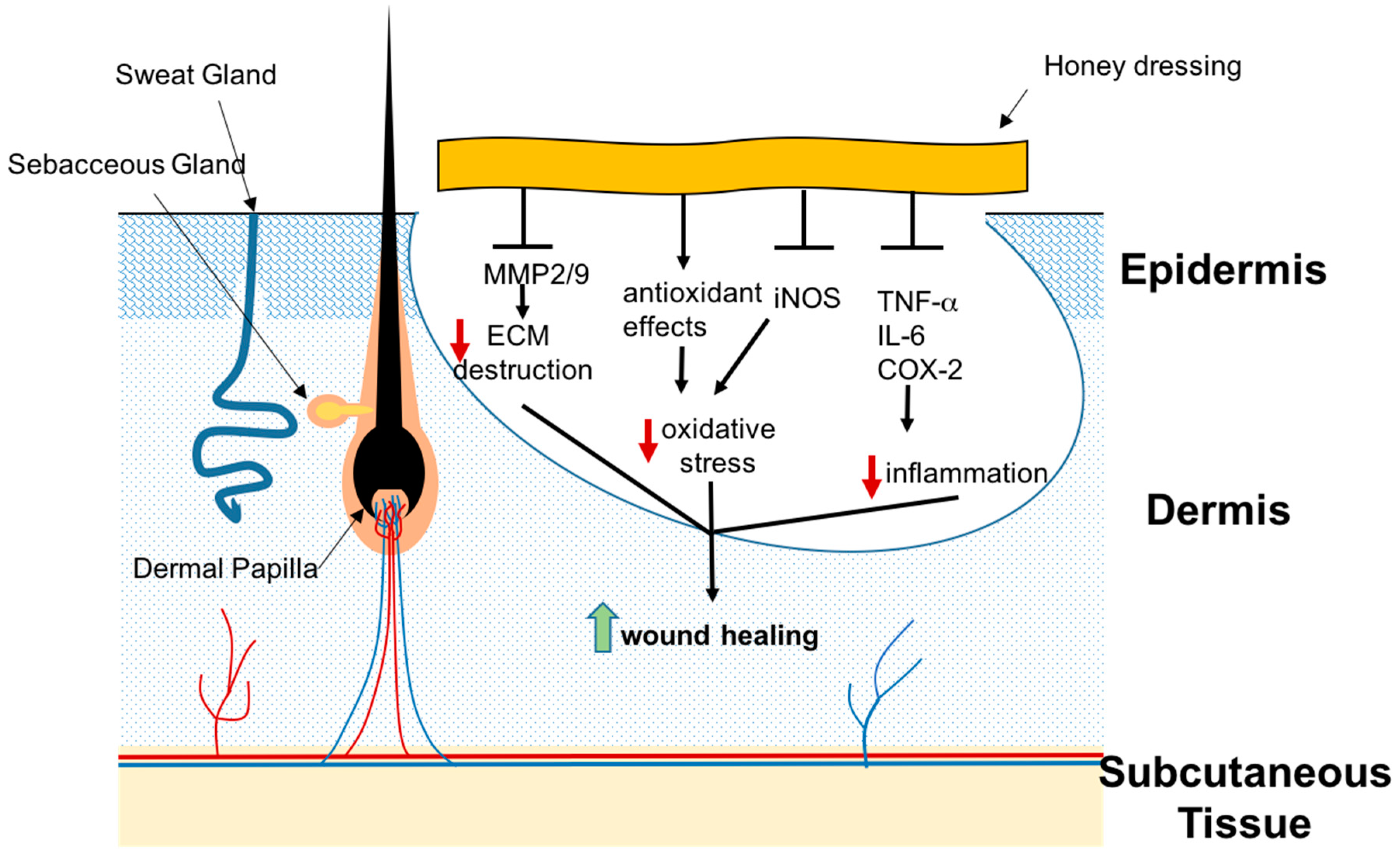
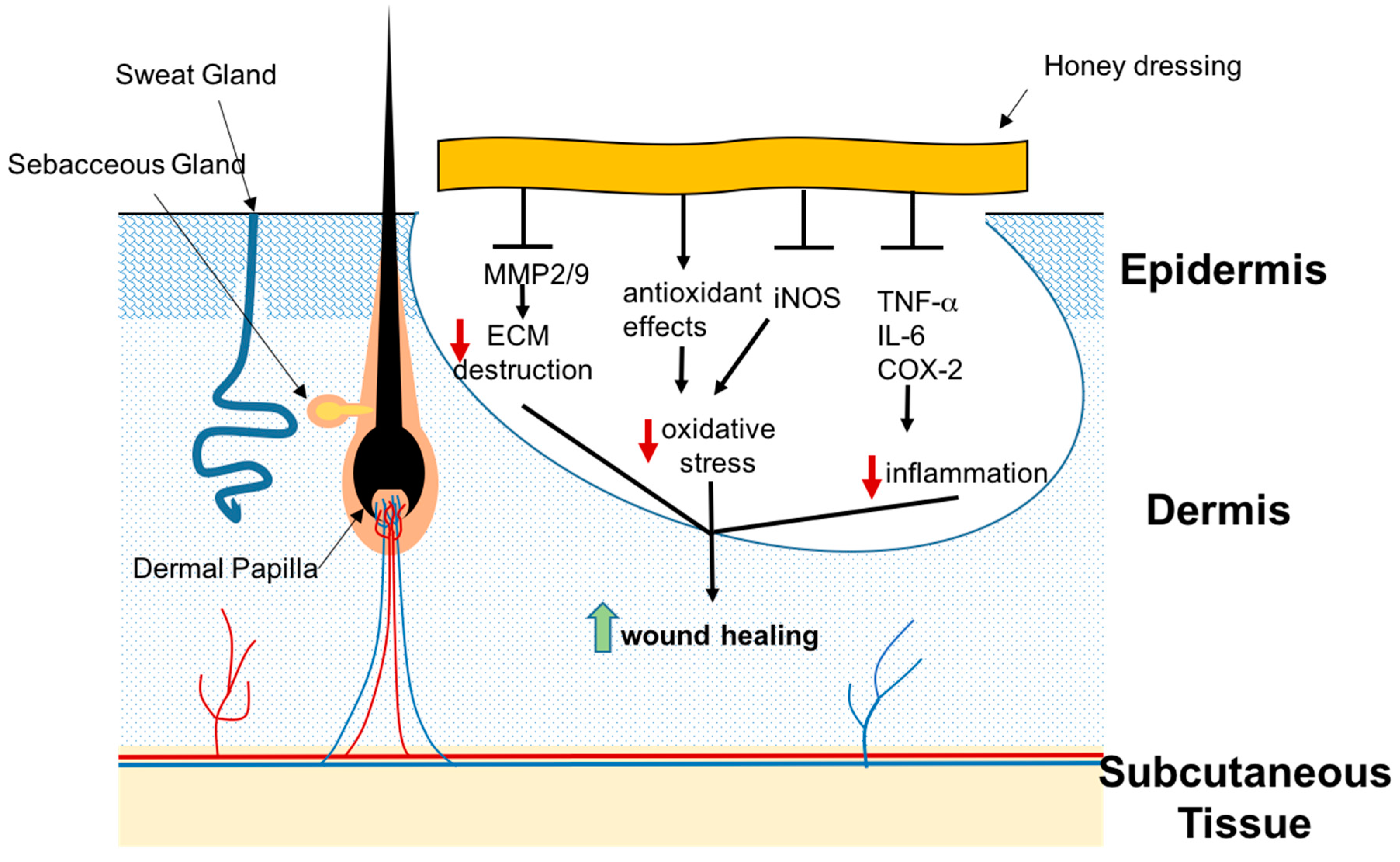
3.2. Honey
3.2.1. Honey and Chronic Wounds
3.2.2. Honey and Burns
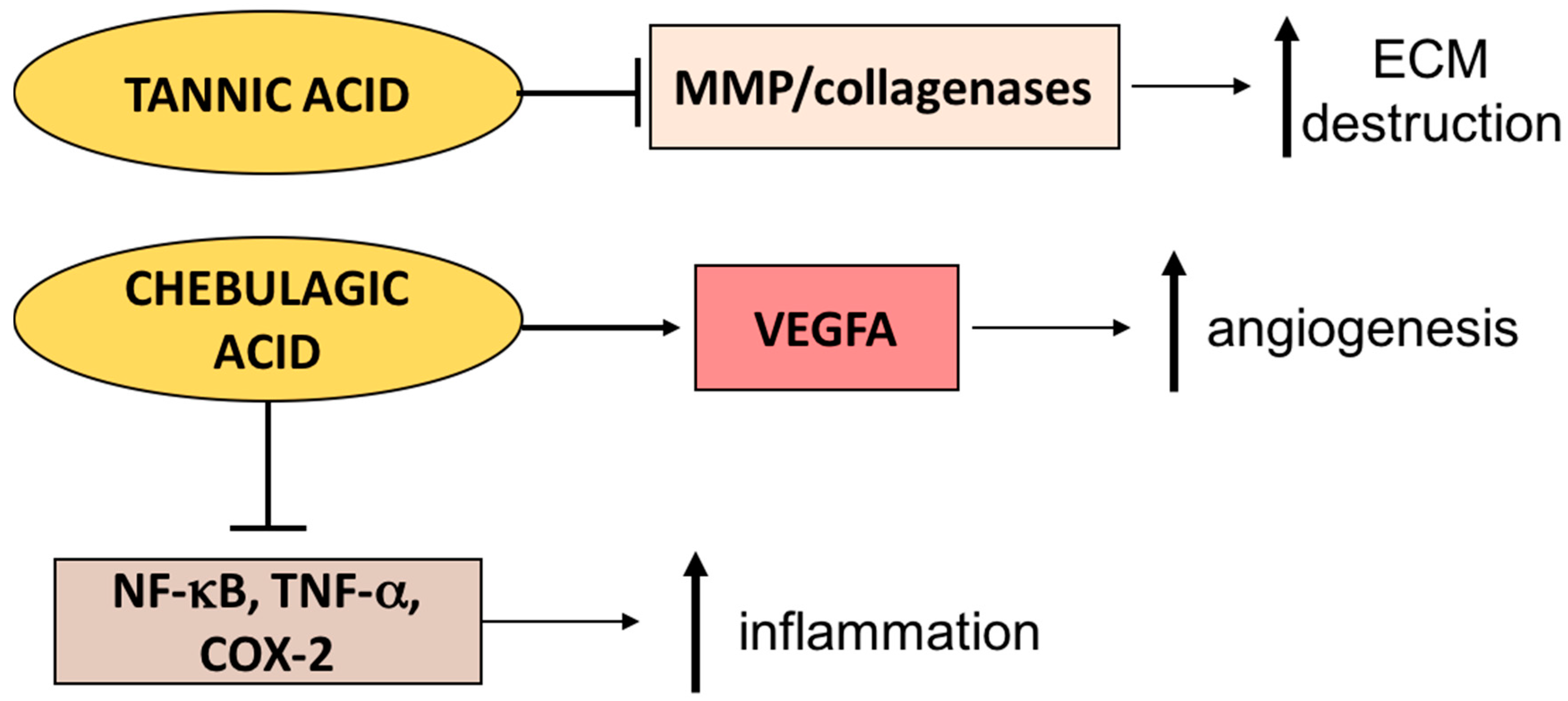
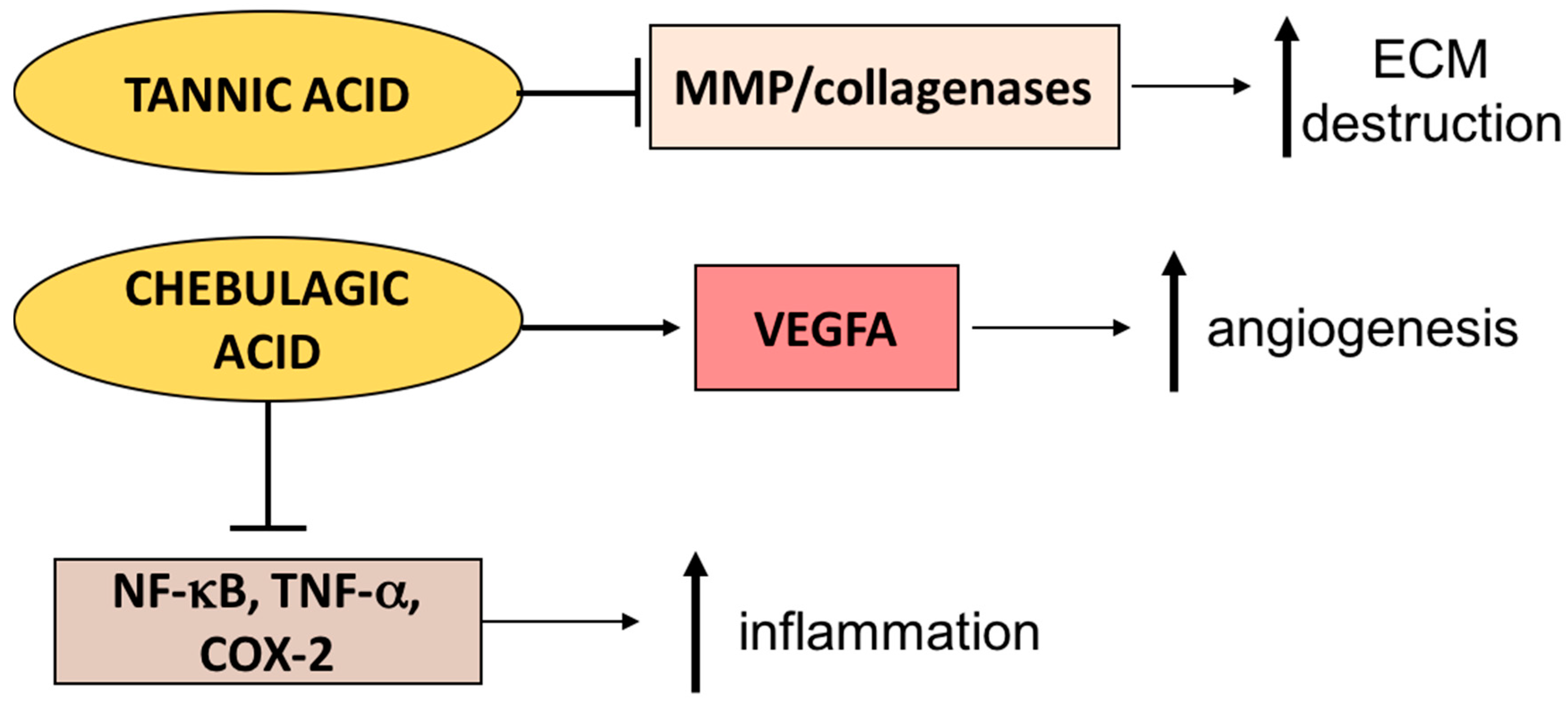
3.3. Terminalia Genus
3.4. Other Plant-Based Sources of Wound Care
3.5. Therapies Combining Phytochemicals with Modern Wound Dressings
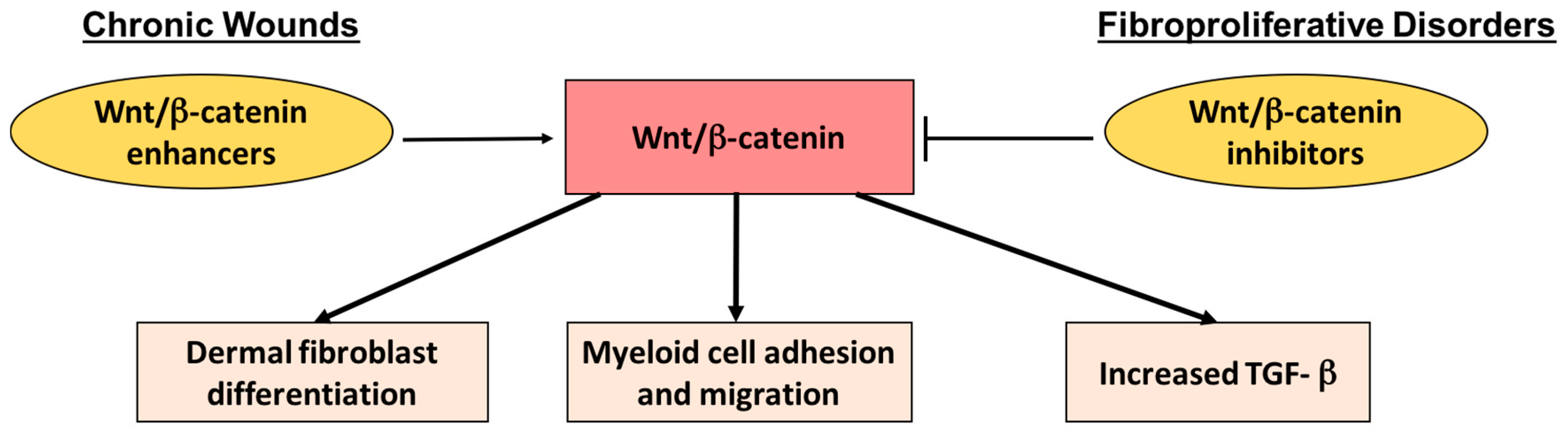
3.6. Effects of Phytochemicals on Stem Cells
4. Discussion and Conclusions
Acknowledgments
Conflicts of Interest
References
- Portou, M.J.; Baker, D.; Abraham, D.; Tsui, J. The innate immune system, toll-like receptors and dermal wound healing: A review. Vascul. Pharmacol. 2015, 71, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, C.C.; Jeschke, M.G.; Branski, L.K.; Barret, J.P.; Dziewulski, P.; Herndon, D.N. Hypertrophic scarring: The greatest unmet challenge after burn injury. Lancet 2016, 388, 1427–1436. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [PubMed]
- Schneider, D.F.; Palmer, J.L.; Tulley, J.M.; Speicher, J.T.; Kovacs, E.J.; Gamelli, R.L.; Faunce, D.E. A novel role for NKT cells in cutaneous wound repair. J. Surg. Res. 2011, 168, 325–333. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, L.A. Wound healing: The role of the macrophage and other immune cells. Shock 1995, 4, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Rodero, M.P.; Khosrotehrani, K. Skin wound healing modulation by macrophages. Int. J. Clin. Exp. Pathol. 2010, 3, 643–653. [Google Scholar] [PubMed]
- Han, Y.P.; Tuan, T.L.; Wu, H.; Hughes, M.; Garner, W.L. TNF-α stimulates activation of pro-MMP2 in human skin through NF-κB mediated induction of MT1-MMP. J. Cell Sci. 2001, 114, 131–139. [Google Scholar] [PubMed]
- Park, J.E.; Barbul, A. Understanding the role of immune regulation in wound healing. Am. J. Surg. 2004, 187, S11–S16. [Google Scholar] [CrossRef]
- Boyce, D.E.; Jones, W.D.; Ruge, F.; Harding, K.G.; Moore, K. The role of lymphocytes in human dermal wound healing. Br. J. Dermatol. 2000, 143, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Stout, R.D.; Suttles, J. T cell signaling of macrophage function in inflammatory disease. Front. Biosci. 1997, 2, d197–d206. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Hammerschmidt, M.; Krieg, T.; Roers, A. Interrelation of immunity and tissue repair or regeneration. Semin. Cell Dev. Biol. 2009, 20, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Iwata, Y.; Yoshizaki, A.; Komura, K.; Shimizu, K.; Ogawa, F.; Hara, T.; Muroi, E.; Bae, S.; Takenaka, M.; Yukami, T.; et al. CD19, a response regulator of B lymphocytes, regulates wound healing through hyaluronan-induced TLR4 signaling. Am. J. Pathol. 2009, 175, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hori, K.; Ding, J.; Huang, Y.; Kwan, P.; Ladak, A.; Tredget, E.E. Toll-like receptors expressed by dermal fibroblasts contribute to hypertrophic scarring. J. Cell. Physiol. 2011, 226, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Butzelaar, L.; Ulrich, M.M.; Mink van der Molen, A.B.; Niessen, F.B.; Beelen, R.H. Currently known risk factors for hypertrophic skin scarring: A review. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiao, H.; Stewart, T.L.; Shankowsky, H.A.; Scott, P.G.; Tredget, E.E. Increased TGF-β-producing CD4+ T lymphocytes in postburn patients and their potential interaction with dermal fibroblasts in hypertrophic scarring. Wound Repair Regen. 2007, 15, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.V.; Hawkins, H.K.; Chinkes, D.; Burke, A.; Tavares, A.L.; Ramos-e-Silva, M.; Albrecht, T.B.; Kitten, G.T.; Herndon, D.N. Hypertrophic versus non hypertrophic scars compared by immunohistochemistry and laser confocal microscopy: Type I and III collagens. Int. Wound J. 2009, 6, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Leventhal, D.; Furr, M.; Reiter, D. Treatment of keloids and hypertrophic scars: A meta-analysis and review of the literature. Arch. Facial Plast. Surg. 2006, 8, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Rabello, F.B.; Souza, C.D.; Farina Junior, J.A. Update on hypertrophic scar treatment. Clinics 2014, 69, 565–573. [Google Scholar] [CrossRef]
- Wetzler, C.; Kampfer, H.; Stallmeyer, B.; Pfeilschifter, J.; Frank, S. Large and sustained induction of chemokines during impaired wound healing in the genetically diabetic mouse: Prolonged persistence of neutrophils and macrophages during the late phase of repair. J. Investig. Dermatol. 2000, 115, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Dovi, J.V.; He, L.K.; DiPietro, L.A. Accelerated wound closure in neutrophil-depleted mice. J. Leukoc. Biol. 2003, 73, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Menke, N.B.; Ward, K.R.; Witten, T.M.; Bonchev, D.G.; Diegelmann, R.F. Impaired wound healing. Clin. Dermatol. 2007, 25, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Nwomeh, B.C.; Liang, H.X.; Cohen, I.K.; Yager, D.R. MMP-8 is the predominant collagenase in healing wounds and nonhealing ulcers. J. Surg. Res. 1999, 81, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, A.B.; Staiano-Coico, L.; Grinnell, F. Wound fluid from chronic leg ulcers contains elevated levels of metalloproteinases MMP-2 and MMP-9. J. Investig. Dermatol. 1993, 101, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Blakytny, R.; Jude, E. The molecular biology of chronic wounds and delayed healing in diabetes. Diabet. Med. 2006, 23, 594–608. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M. Phagocytic leukocytes and reactive oxygen species. Histochem. Cell Biol. 2009, 131, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Dissemond, J.; Goos, M.; Wagner, S.N. The role of oxidative stress in the pathogenesis and therapy of chronic wounds. Hautarzt 2002, 53, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Mirza, R.; Koh, T.J. Dysregulation of monocyte/macrophage phenotype in wounds of diabetic mice. Cytokine 2011, 56, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Suntar, I.; Akkol, E.; Nahar, L.; Sarker, S. Wound healing and antioxidant properties: Do they coexist in plants? Free Rad. Antiox. 2012, 2, 1–7. [Google Scholar] [CrossRef]
- Biswas, T.K.; Mukherjee, B. Plant medicines of indian origin for wound healing activity: A review. Int. J. Low Extrem. Wounds 2003, 2, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M. Inhibition of tumor necrosis factor by curcumin, a phytochemical. Biochem. Pharmacol. 1995, 49, 1551–1556. [Google Scholar] [CrossRef]
- Bierhaus, A.; Zhang, Y.; Quehenberger, P.; Luther, T.; Haase, M.; Muller, M.; Mackman, N.; Ziegler, R.; Nawroth, P.P. The dietary pigment curcumin reduces endothelial tissue factor gene expression by inhibiting binding of AP-1 to the DNA and activation of NF-κB. Thromb. Haemost. 1997, 77, 772–782. [Google Scholar] [PubMed]
- Singh, S.; Aggarwal, B.B. Activation of transcription factor NF-κB is suppressed by curcumin (diferuloylmethane). J. Biol. Chem. 1995, 270, 24995–25000. [Google Scholar] [CrossRef] [PubMed]
- El-Refaie, W.M.; Elnaggar, Y.S.; El-Massik, M.A.; Abdallah, O.Y. Novel curcumin-loaded gel-core hyaluosomes with promising burn-wound healing potential: Development, in vitro appraisal and in vivo studies. Int. J. Pharm. 2015, 486, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Heng, M.C. Wound healing in adult skin: Aiming for perfect regeneration. Int. J. Dermatol. 2011, 50, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, C.; Das, M.; Sahoo, S.K. Sustained wound healing activity of curcumin loaded oleic acid based polymeric bandage in a rat model. Mol. Pharm. 2012, 9, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, D.; Ahmed, M.R.; Gomathi, K.; Chitra, K.; Sehgal, P.K.; Jayakumar, R. Dermal wound healing processes with curcumin incorporated collagen films. Biomaterials 2004, 25, 1911–1917. [Google Scholar] [CrossRef]
- Hegge, A.B.; Andersen, T.; Melvik, J.E.; Bruzell, E.; Kristensen, S.; Tonnesen, H.H. Formulation and bacterial phototoxicity of curcumin loaded alginate foams for wound treatment applications: Studies on curcumin and curcuminoides xlii. J. Pharm. Sci. 2011, 100, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, R.B.; Kann, B.; Coradini, K.; Offerhaus, H.L.; Beck, R.C.; Windbergs, M. Skin penetration behavior of lipid-core nanocapsules for simultaneous delivery of resveratrol and curcumin. Eur. J. Pharm. Sci. 2015, 78, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Karri, V.V.; Kuppusamy, G.; Talluri, S.V.; Mannemala, S.S.; Kollipara, R.; Wadhwani, A.D.; Mulukutla, S.; Raju, K.R.; Malayandi, R. Curcumin loaded chitosan nanoparticles impregnated into collagen-alginate scaffolds for diabetic wound healing. Int. J. Biol. Macromol. 2016, 93, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Feng, X.; Xiang, X.; Jin, Y. Wound healing effect of an in situ forming hydrogel loading curcumin-phospholipid complex. Curr. Drug Deliv. 2016, 13, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, G.S.; Mani, H.; Gaddipati, J.P.; Singh, A.K.; Seth, P.; Banaudha, K.K.; Patnaik, G.K.; Maheshwari, R.K. Curcumin enhances wound healing in streptozotocin induced diabetic rats and genetically diabetic mice. Wound Repair Regen. 1999, 7, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Komada, M.R.; Sane, D.C. Abnormal angiogenesis in diabetes mellitus. Med. Res. Rev. 2003, 23, 117–145. [Google Scholar] [CrossRef] [PubMed]
- Kant, V.; Gopal, A.; Kumar, D.; Pathak, N.N.; Ram, M.; Jangir, B.L.; Tandan, S.K.; Kumar, D. Curcumin-induced angiogenesis hastens wound healing in diabetic rats. J. Surg. Res. 2015, 193, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Gadekar, R.; Saurabh, M.; Thakur, G.; Saurabh, A. Study of formulation, characterisation and wound healing potential of transdermal patches of curcumin. Asian J. Pharm. Clin. Res. 2012, 5, 225–230. [Google Scholar]
- Panchatcharam, M.; Miriyala, S.; Gayathri, V.S.; Suguna, L. Curcumin improves wound healing by modulating collagen and decreasing reactive oxygen species. Mol. Cell. Biochem. 2006, 290, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Guntas, G.; Engin, B.; Ekmekci, O.B.; Kutlubay, Z.; Ekmekci, H.; Songur, A.; Uzuncakmak, T.K.; Vehid, H.E.; Serdaroglu, S.; Tuzun, Y.; et al. Evaluation of advanced oxidation protein products, prooxidant-antioxidant balance, and total antioxidant capacity in untreated vitiligo patients. Ann. Dermatol. 2015, 27, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Asawanonda, P.; Klahan, S.O. Tetrahydrocurcuminoid cream plus targeted narrowband UVB phototherapy for vitiligo: A preliminary randomized controlled study. Photomed. Laser Surg. 2010, 28, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Arno, A.I.; Amini-Nik, S.; Blit, P.H.; Al-Shehab, M.; Belo, C.; Herer, E.; Jeschke, M.G. Effect of human wharton’s jelly mesenchymal stem cell paracrine signaling on keloid fibroblasts. Stem Cells Transl. Med. 2014, 3, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Chen, M.J.; Yu, Y.M.; Ko, S.Y.; Chang, C.C. Suppression of TGF-β1/smad pathway and extracellular matrix production in primary keloid fibroblasts by curcuminoids: Its potential therapeutic use in the chemoprevention of keloid. Arch. Dermatol. Res. 2010, 302, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Peng, S.; Sun, Z.; Li, H.; Yang, R. Curcumin suppresses TGF-β signaling by inhibition of TGIF degradation in scleroderma fibroblasts. Biochem. Biophys. Res. Commun. 2011, 411, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.T.; Sun, L.; Bay, B.H.; Chan, S.Y.; Lee, S.T. Dietary compounds inhibit proliferation and contraction of keloid and hypertrophic scar-derived fibroblasts in vitro: Therapeutic implication for excessive scarring. J. Trauma 2003, 54, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J. Honey: An immunomodulator in wound healing. Wound Repair Regen. 2014, 22, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Suarez, J.M.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Mazzoni, L.; Giampieri, F. The composition and biological activity of honey: A focus on manuka honey. Foods 2014, 3, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Jerkovic, I.; Kus, P.M.; Tuberoso, C.I.; Sarolic, M. Phytochemical and physical-chemical analysis of polish willow (Salix spp.) honey: Identification of the marker compounds. Food Chem. 2014, 145, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.E.; Cokcetin, N.N.; Harry, E.J.; Carter, D.A. The unusual antibacterial activity of medical-grade leptospermum honey: Antibacterial spectrum, resistance and transcriptome analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Church, D.; Elsayed, S.; Reid, O.; Winston, B.; Lindsay, R. Burn wound infections. Clin. Microbiol. Rev. 2006, 19, 403–434. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. Potential of honey in the treatment of wounds and burns. Am. J. Clin. Dermatol. 2001, 2, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Alam, F.; Islam, M.A.; Gan, S.H.; Khalil, M.I. Honey: A potential therapeutic agent for managing diabetic wounds. Evid. Based Complement. Altern. Med. 2014, 2014, 169130. [Google Scholar] [CrossRef] [PubMed]
- Hussein, S.Z.; Mohd Yusoff, K.; Makpol, S.; Mohd Yusof, Y.A. Gelam honey inhibits the production of proinflammatory, mediators NO, PGE2, TNF-α, and IL-6 in carrageenan-induced acute paw edema in rats. Evid. Based Complement. Altern. Med. 2012, 2012, 109636. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Bohova, J.; Garcia-Villalba, R.; Tomas-Barberan, F.A.; Madakova, Z.; Majtan, T.; Majtan, V.; Klaudiny, J. Fir honeydew honey flavonoids inhibit TNF-β-induced MMP-9 expression in human keratinocytes: A new action of honey in wound healing. Arch. Dermatol. Res. 2013, 305, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Park, H.J.; Chung, H.J.; Min, H.Y.; Park, E.J.; Hong, J.Y.; Lee, S.K. Inhibitory effects of caffeic acid phenethyl ester on cancer cell metastasis mediated by the down-regulation of matrix metalloproteinase expression in human ht1080 fibrosarcoma cells. J. Nutr. Biochem. 2006, 17, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Moskwa, J.; Borawska, M.H.; Markiewicz-Zukowska, R.; Puscion-Jakubik, A.; Naliwajko, S.K.; Socha, K.; Soroczynska, J. Polish natural bee honeys are anti-proliferative and anti-metastatic agents in human glioblastoma multiforme U87MG cell line. PLoS ONE 2014, 9, e90533. [Google Scholar] [CrossRef] [PubMed]
- Henriques, A.; Jackson, S.; Cooper, R.; Burton, N. Free radical production and quenching in honeys with wound healing potential. J. Antimicrob. Chemother. 2006, 58, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Gheldof, N.; Wang, X.H.; Engeseth, N.J. Identification and quantification of antioxidant components of honeys from various floral sources. J. Agric. Food Chem. 2002, 50, 5870–5877. [Google Scholar] [CrossRef] [PubMed]
- Efem, S.E. Clinical observations on the wound healing properties of honey. Br. J. Surg. 1988, 75, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Moghazy, A.M.; Shams, M.E.; Adly, O.A.; Abbas, A.H.; El-Badawy, M.A.; Elsakka, D.M.; Hassan, S.A.; Abdelmohsen, W.S.; Ali, O.S.; Mohamed, B.A. The clinical and cost effectiveness of bee honey dressing in the treatment of diabetic foot ulcers. Diabetes Res. Clin. Pract. 2010, 89, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. The evidence supporting the use of honey as a wound dressing. Int. J. Low Extrem. Wounds 2006, 5, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Pecanac, M.; Janjic, Z.; Komarcevic, A.; Pajic, M.; Dobanovacki, D.; Miskovic, S.S. Burns treatment in ancient times. Med. Pregl. 2013, 66, 263–267. [Google Scholar] [PubMed]
- Iftikhar, F.; Arshad, M.; Rasheed, F.; Amraiz, D.; Anwar, P.; Gulfraz, M. Effects of acacia honey on wound healing in various rat models. Phytother. Res. 2010, 24, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, M.; Weatherall, M.; Perrin, K.; Beasley, R. Honey in the treatment of burns: A systematic review and meta-analysis of its efficacy. N. Z. Med. J. 2009, 122, 47–60. [Google Scholar] [PubMed]
- Subrahmanyam, M. Topical application of honey in treatment of burns. Br. J. Surg. 1991, 78, 497–498. [Google Scholar] [CrossRef] [PubMed]
- Cock, I.E. The medicinal properties and phytochemistry of plants of the genus terminalia (Combretaceae). Inflammopharmacology 2015, 23, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Diao, Y.; Zhang, H.; Wang, S.; Zhang, Z.; Yu, B.; Huang, S.; Yang, H. Tannin extracts from immature fruits of terminalia chebula fructus retz. Promote cutaneous wound healing in rats. BMC Complement. Altern. Med. 2011, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Rane, M.M.; Mengi, S.A. Comparative effect of oral administration and topical application of alcoholic extract of terminalia arjuna bark on incision and excision wounds in rats. Fitoterapia 2003, 74, 553–558. [Google Scholar] [CrossRef]
- Mongalo, N.I.; McGaw, L.J.; Segapelo, T.V.; Finnie, J.F.; van Staden, J. Ethnobotany, phytochemistry, toxicology and pharmacological properties of Terminalia sericea Burch. Ex DC. (Combretaceae)—A review. J. Ethnopharmacol. 2016, 194, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, S.J.; Wang, D.; Wei, Y.J.; Hu, S.S. Intramyocardial injection of tannic acid attenuates postinfarction remodeling: A novel approach to stabilize the breaking extracellular matrix. J. Thorac. Cardiovasc. Surg. 2009, 137, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, V.; Krithica, N.; Madhan, B.; Sehgal, P.K. Preparation and properties of tannic acid cross-linked collagen scaffold and its application in wound healing. J. Biomed. Mater. Res. B Appl. Biomater. 2013, 101, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Suguna, L.; Singh, S.; Sivakumar, P.; Sampath, P.; Chandrakasan, G. Influence of terminalia chebula on dermal wound healing in rats. Phytother. Res. 2002, 16, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Singh, D.; Choi, S.M.; Zo, S.M.; Painuli, R.M.; Kwon, S.W.; Han, S.S. Effect of extracts of terminalia chebula on proliferation of keratinocytes and fibroblasts cells: An alternative approach for wound healing. Evid. Based Complement. Altern. Med. 2014, 2014, 701656. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.B.; Reddanna, P. Chebulagic acid (CA) attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in raw 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 381, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Mehta, N.M.; Srivastava, D.N. Healing potential of ethyl acetate soluble fraction of ethanolic extract of terminalia chebula on experimental cutaneous wounds in streptozotocin induced diabetic rats. Asian J. Biomed. Pharm. Sci. 2013, 3, 32–36. [Google Scholar]
- Nasiri, E.; Hosseinimehr, S.J.; Azadbakht, M.; Akbari, J.; Enayati-Fard, R.; Azizi, S. The effect of terminalia chebula extract vs. Silver sulfadiazine on burn wounds in rats. J. Complement. Integr. Med. 2015, 12, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Steenkamp, V.; Mathivha, E.; Gouws, M.C.; van Rensburg, C.E. Studies on antibacterial, antioxidant and fibroblast growth stimulation of wound healing remedies from south africa. J. Ethnopharmacol. 2004, 95, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, P.; Mwasumbi, L.; Haeggstrom, C.A.; Vuorela, H.; Hiltunen, R.; Vuorela, P. Ethnobotanical and antimicrobial investigation on some species of terminalia and combretum (Combretaceae) growing in tanzania. J. Ethnopharmacol. 2002, 79, 169–177. [Google Scholar] [CrossRef]
- Moshi, M.J.; Mbwambo, Z.H. Some pharmacological properties of extracts of terminalia sericea roots. J. Ethnopharmacol. 2005, 97, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Stulberg, D.L.; Penrod, M.A.; Blatny, R.A. Common bacterial skin infections. Am. Fam. Phys. 2002, 66, 119–124. [Google Scholar]
- Phan, T.T.; Hughes, M.A.; Cherry, G.W.; Le, T.T.; Pham, H.M. An aqueous extract of the leaves of Chromolaena odorata (formerly eupatorium odoratum) (eupolin) inhibits hydrated collagen lattice contraction by normal human dermal fibroblasts. J. Altern. Complement. Med. 1996, 2, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.T.; Hughes, M.A.; Cherry, G.W. Enhanced proliferation of fibroblasts and endothelial cells treated with an extract of the leaves of Chromolaena odorata (eupolin), an herbal remedy for treating wounds. Plast. Reconstr. Surg. 1998, 101, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Thang, P.T.; Patrick, S.; Teik, L.S.; Yung, C.S. Anti-oxidant effects of the extracts from the leaves of Chromolaena odorata on human dermal fibroblasts and epidermal keratinocytes against hydrogen peroxide and hypoxanthine-xanthine oxidase induced damage. Burns 2001, 27, 319–327. [Google Scholar] [CrossRef]
- Adetutu, A.; Morgan, W.A.; Corcoran, O. Antibacterial, antioxidant and fibroblast growth stimulation activity of crude extracts of bridelia ferruginea leaf, a wound-healing plant of Nigeria. J. Ethnopharmacol. 2011, 133, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Annan, K.; Houghton, P.J. Antibacterial, antioxidant and fibroblast growth stimulation of aqueous extracts of Ficus asperifolia Miq. And Gossypium arboreum L., wound-healing plants of Ghana. J. Ethnopharmacol. 2008, 119, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Bahramsoltani, R.; Farzaei, M.H.; Rahimi, R. Medicinal plants and their natural components as future drugs for the treatment of burn wounds: An integrative review. Arch. Dermatol. Res. 2014, 306, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Duansak, D.; Somboonwong, J.; Patumraj, S. Effects of aloe vera on leukocyte adhesion and TNF-α and IL-6 levels in burn wounded rats. Clin. Hemorheol. Microcirc. 2003, 29, 239–246. [Google Scholar] [PubMed]
- Hosseinimehr, S.J.; Khorasani, G.; Azadbakht, M.; Zamani, P.; Ghasemi, M.; Ahmadi, A. Effect of aloe cream versus silver sulfadiazine for healing burn wounds in rats. Acta Dermatovenerol. Croat. 2010, 18, 2–7. [Google Scholar] [PubMed]
- Somboonwong, J.; Thanamittramanee, S.; Jariyapongskul, A.; Patumraj, S. Therapeutic effects of Aloe vera on cutaneous microcirculation and wound healing in second degree burn model in rats. J. Med. Assoc. Thai. 2000, 83, 417–425. [Google Scholar] [PubMed]
- Rodriguez-Bigas, M.; Cruz, N.I.; Suarez, A. Comparative evaluation of Aloe vera in the management of burn wounds in guinea pigs. Plast. Reconstr. Surg. 1988, 81, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.N.; Ahmed, N. Effectiveness of aloe vera gel compared with 1% silver sulphadiazine cream as burn wound dressing in second degree burns. JPMA J. Pak. Med. Assoc. 2013, 63, 225–230. [Google Scholar] [PubMed]
- Visuthikosol, V.; Chowchuen, B.; Sukwanarat, Y.; Sriurairatana, S.; Boonpucknavig, V. Effect of Aloe vera gel to healing of burn wound a clinical and histologic study. J. Med. Assoc. Thai. 1995, 78, 403–409. [Google Scholar] [PubMed]
- Jayakumar, R.; Prabaharan, M.; Sudheesh Kumar, P.T.; Nair, S.V.; Tamura, H. Biomaterials based on chitin and chitosan in wound dressing applications. Biotechnol. Adv. 2011, 29, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, E.A.; Kenawy, E.S.; Chen, X. A review on polymeric hydrogel membranes for wound dressing applications: PVA-based hydrogel dressings. J. Adv. Res. 2017, 8, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramirez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Boonkaew, B.; Kempf, M.; Kimble, R.; Supaphol, P.; Cuttle, L. Antimicrobial efficacy of a novel silver hydrogel dressing compared to two common silver burn wound dressings: Acticoat™ and polymem silver®. Burns 2014, 40, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Varaprasad, K.; Vimala, K.; Ravindra, S.; Narayana Reddy, N.; Venkata Subba Reddy, G.; Mohana Raju, K. Fabrication of silver nanocomposite films impregnated with curcumin for superior antibacterial applications. J. Mater. Sci. Mater. Med. 2011, 22, 1863–1872. [Google Scholar] [CrossRef] [PubMed]
- Merrell, J.G.; McLaughlin, S.W.; Tie, L.; Laurencin, C.T.; Chen, A.F.; Nair, L.S. Curcumin-loaded poly(epsilon-caprolactone) nanofibres: Diabetic wound dressing with anti-oxidant and anti-inflammatory properties. Clin. Exp. Pharmacol. Physiol. 2009, 36, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zohdi, R.; Abu Bakar Zakaria, Z.; Yusof, N.; Mohamed Mustapha, N.; Abdullah, M.N. Gelam (Melaleuca spp.) honey-based hydrogel as burn wound dressing. Evid. Based Complement. Altern. Med. 2012, 2012, 843025. [Google Scholar] [CrossRef] [PubMed]
- Nho, Y.-C.; Park, J.-S.; Lim, Y.-M. Preparation of hydrogel by radiation for the healing of diabetic ulcer. Radiat. Phys. Chem. 2014, 94, 176–180. [Google Scholar] [CrossRef]
- Kawasaki, B.T.; Hurt, E.M.; Mistree, T.; Farrar, W.L. Targeting cancer stem cells with phytochemicals. Mol. Interv. 2008, 8, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.; Padhye, S.; Ahmad, A.; Sarkar, F.H. Novel strategies targeting cancer stem cells through phytochemicals and their analogs. Drug Deliv. Transl. Res. 2013, 3, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Choi, K.Y.; Surh, Y.J. β-catenin-mediated signaling: A novel molecular target for chemoprevention with anti-inflammatory substances. Biochim. Biophys. Acta 2006, 1765, 14–24. [Google Scholar] [CrossRef] [PubMed]
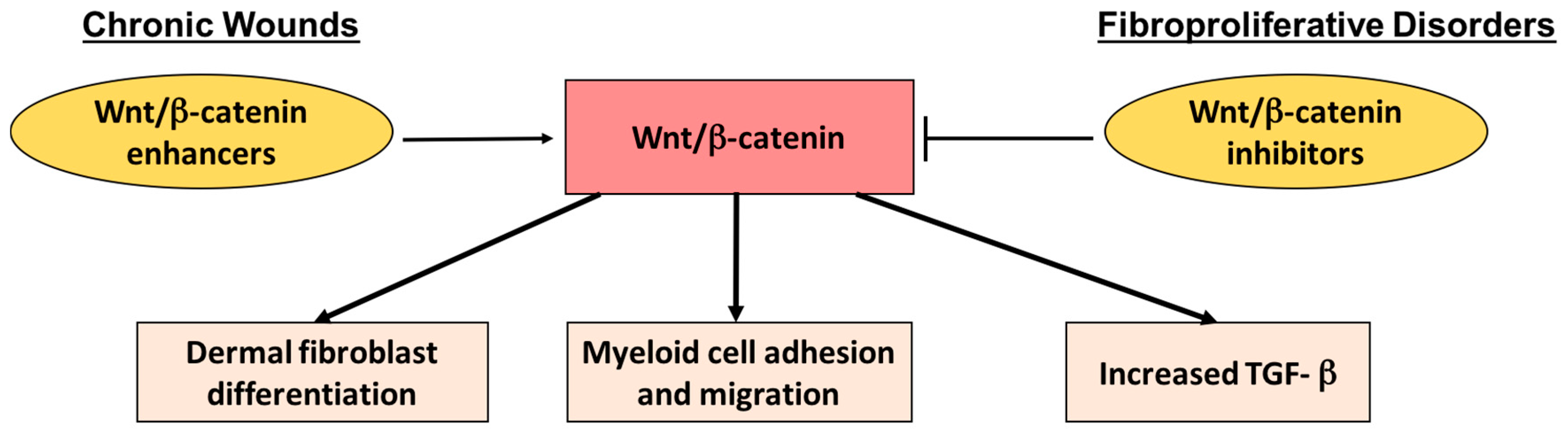
- Bielefeld, K.A.; Amini-Nik, S.; Alman, B.A. Cutaneous wound healing: Recruiting developmental pathways for regeneration. Cell. Mol. Life Sci. 2013, 70, 2059–2081. [Google Scholar] [CrossRef] [PubMed]
- Bielefeld, K.A.; Amini-Nik, S.; Whetstone, H.; Poon, R.; Youn, A.; Wang, J.; Alman, B.A. Fibronectin and β-catenin act in a regulatory loop in dermal fibroblasts to modulate cutaneous healing. J. Biol. Chem. 2011, 286, 27687–27697. [Google Scholar] [CrossRef] [PubMed]
- Amini Nik, S.; Ebrahim, R.P.; van Dam, K.; Cassiman, J.J.; Tejpar, S. TGF-β modulates β-catenin stability and signaling in mesenchymal proliferations. Exp. Cell. Res. 2007, 313, 2887–2895. [Google Scholar] [CrossRef] [PubMed]
- Amini-Nik, S.; Cambridge, E.; Yu, W.; Guo, A.; Whetstone, H.; Nadesan, P.; Poon, R.; Hinz, B.; Alman, B.A. β-catenin-regulated myeloid cell adhesion and migration determine wound healing. J. Clin. Investig. 2014, 124, 2599–2610. [Google Scholar] [CrossRef] [PubMed]
- Amini-Nik, S.; Glancy, D.; Boimer, C.; Whetstone, H.; Keller, C.; Alman, B.A. Pax7 expressing cells contribute to dermal wound repair, regulating scar size through a β-catenin mediated process. Stem Cells 2011, 29, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Peters, T.; Sindrilaru, A.; Hinz, B.; Hinrichs, R.; Menke, A.; Al-Azzeh, E.A.; Holzwarth, K.; Oreshkova, T.; Wang, H.; Kess, D.; et al. Wound-healing defect of CD18−/− mice due to a decrease in TGF-β1 and myofibroblast differentiation. EMBO J. 2005, 24, 3400–3410. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.S.; Wei, Q.; Gurung, A.; Youn, A.; Bright, T.; Poon, R.; Whetstone, H.; Guha, A.; Alman, B.A. β-catenin regulates wound size and mediates the effect of TGF-β in cutaneous healing. FASEB J. 2006, 20, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.S.; Cheah, A.Y.; Turley, S.; Nadesan, P.; Poon, R.; Clevers, H.; Alman, B.A. β-catenin stabilization dysregulates mesenchymal cell proliferation, motility, and invasiveness and causes aggressive fibromatosis and hyperplastic cutaneous wounds. Proc. Natl. Acad. Sci. USA 2002, 99, 6973–6978. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Ramakrishnan, S.M.; Bagler, G. Phytochemica: A platform to explore phytochemicals of medicinal plants. Database 2015, 2015. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal | Excessive | Deficient |
|---|---|---|
| Adequate inflammatory response | Prolonged inflammatory phase | Dysregulated hyperinflammatory response |
| Adequate collagenase expression | Decreased collagenase expression | Overexpression of collagenases and gelatinases |
| Increased type I collagen | Increased type III collagen | Matrix metalloproteinases (MMP)-mediated destruction of extra-cellular matrix (ECM) |
| Minimal scarring | Hypertrophic scarring/fibrosis | Inadequate wound closure |
| Phytochemical | Phase of Wound Healing | In Vitro | In Vivo (Animal) | In Vivo (Human) |
|---|---|---|---|---|
| Curcumin | Inflammatory phase | [33,34,35] | Rat [47,48] | Clinical Trials [50] |
| Proliferation phase | [52] | Rat [46] | - | |
| Remodeling phase | - | - | - | |
| Honey | Inflammatory phase | [64] | Rat [70] | RCT [66,67,68,71,72] |
| Proliferation phase | - | - | RCT [72] | |
| Remodeling phase | - | - | - | |
| Terminalia | Inflammatory phase | [76,77,78,81] | Rat [79,82,83] | - |
| Proliferation phase | [80] | Rat [74] | - | |
| Remodeling phase | - | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, A.; Amini-Nik, S. The Role of Phytochemicals in the Inflammatory Phase of Wound Healing. Int. J. Mol. Sci. 2017, 18, 1068. https://doi.org/10.3390/ijms18051068
Shah A, Amini-Nik S. The Role of Phytochemicals in the Inflammatory Phase of Wound Healing. International Journal of Molecular Sciences. 2017; 18(5):1068. https://doi.org/10.3390/ijms18051068
Chicago/Turabian StyleShah, Ahmed, and Saeid Amini-Nik. 2017. "The Role of Phytochemicals in the Inflammatory Phase of Wound Healing" International Journal of Molecular Sciences 18, no. 5: 1068. https://doi.org/10.3390/ijms18051068