
Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus chilensis Hemocytes

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Primary Culture of Hemocytes from M. chilensis
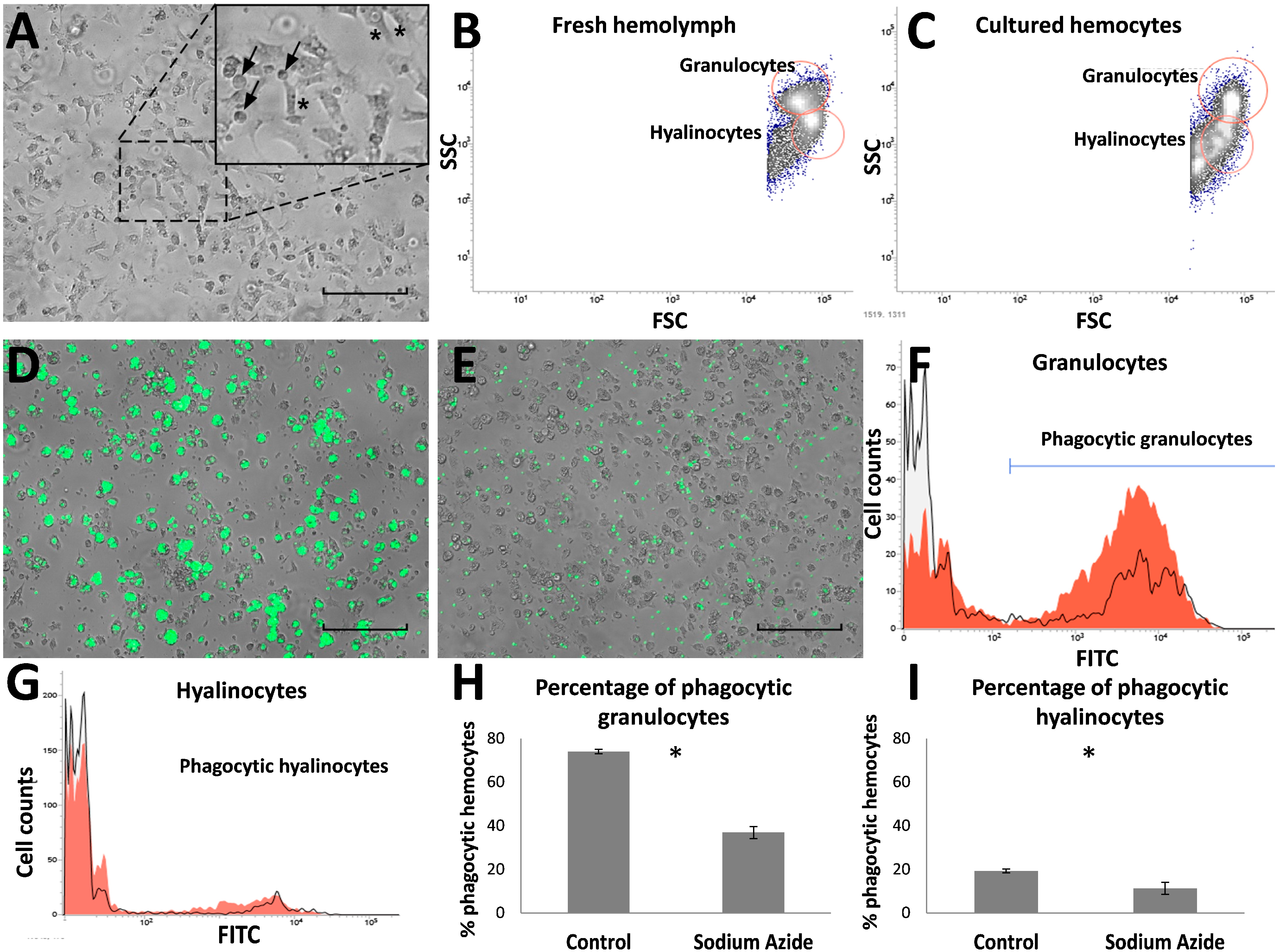
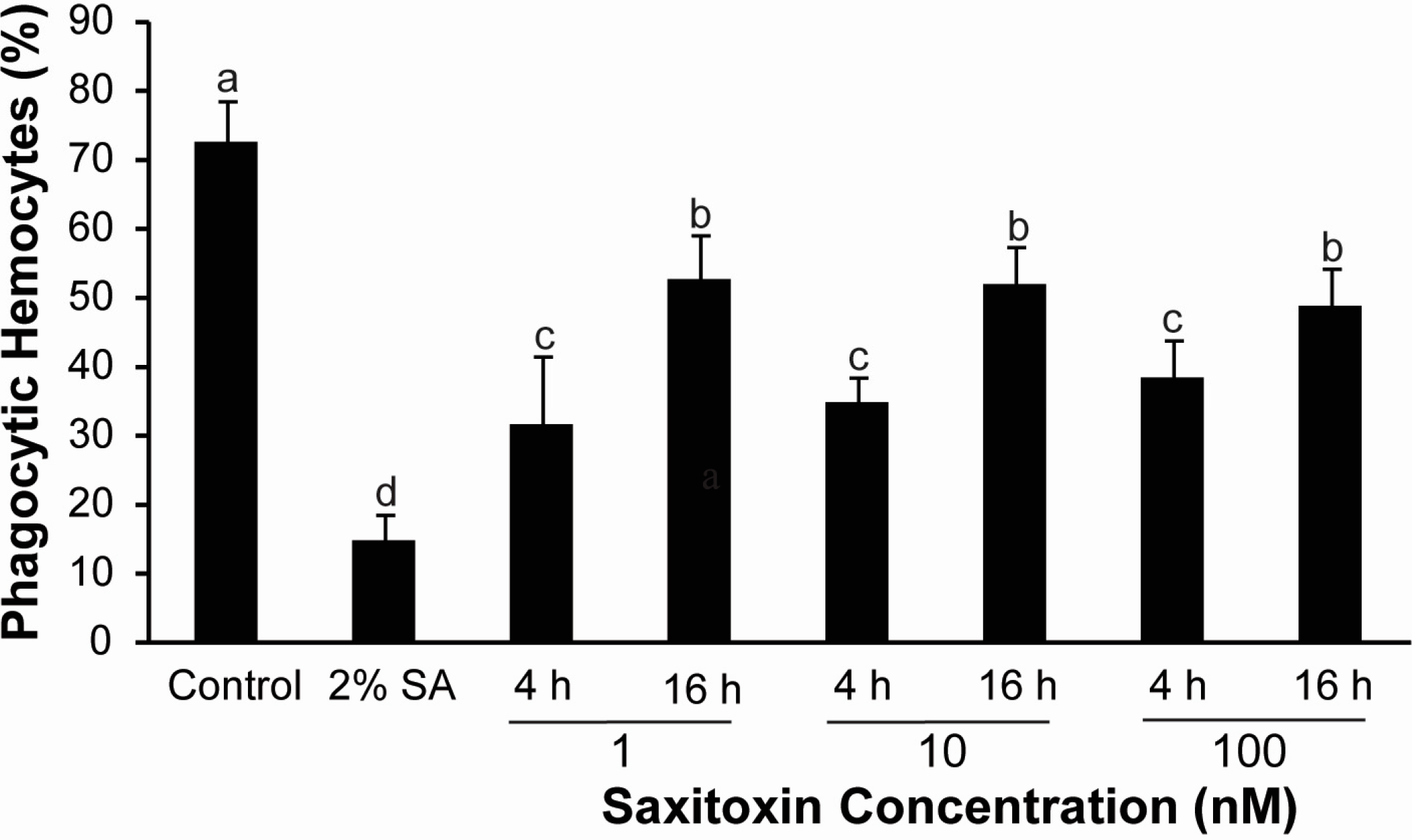
2.2. Effects of Saxitoxin (STX) on Cellular Parameters
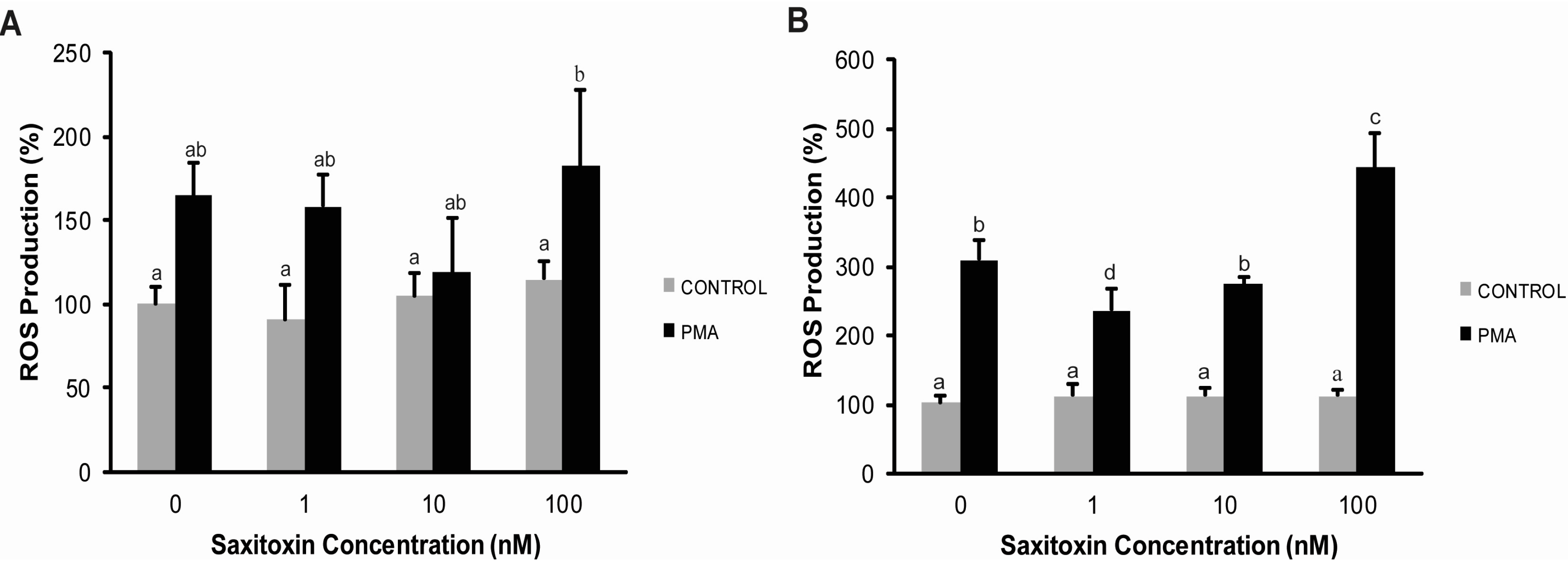
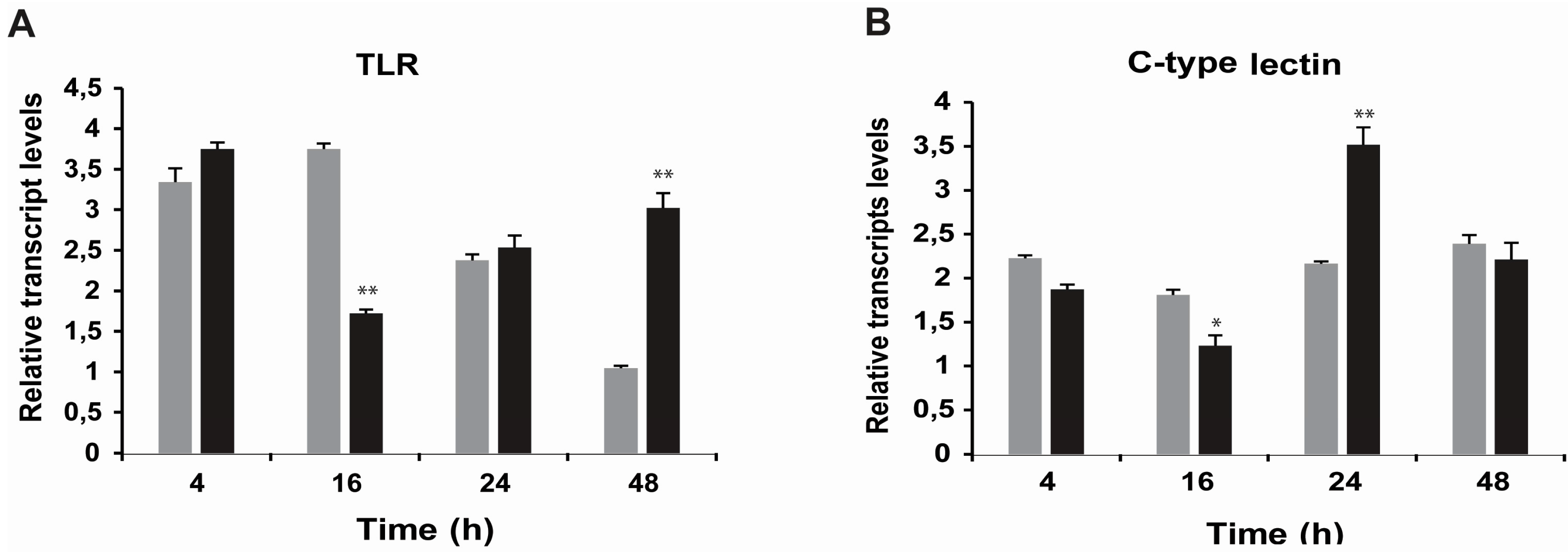
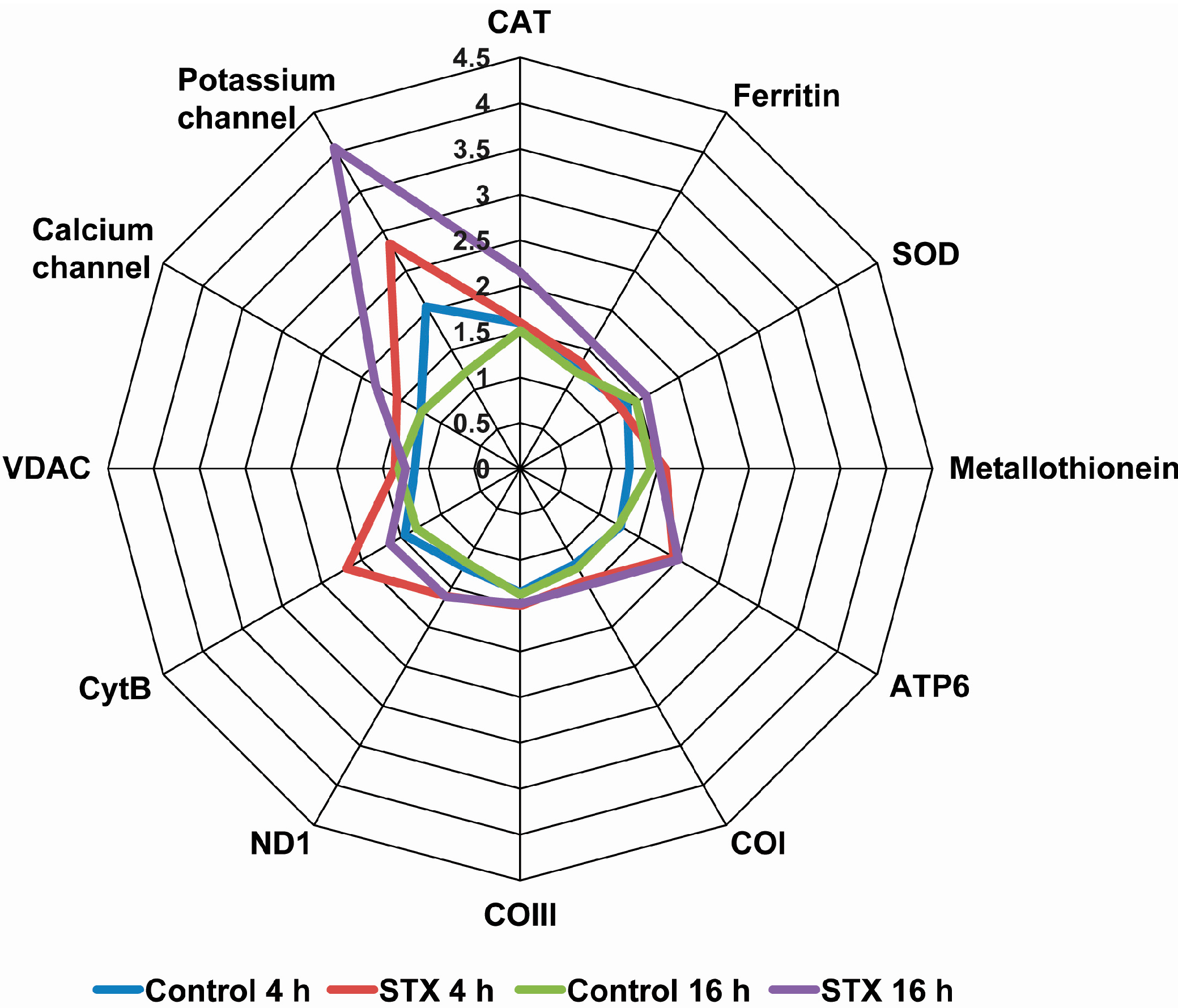
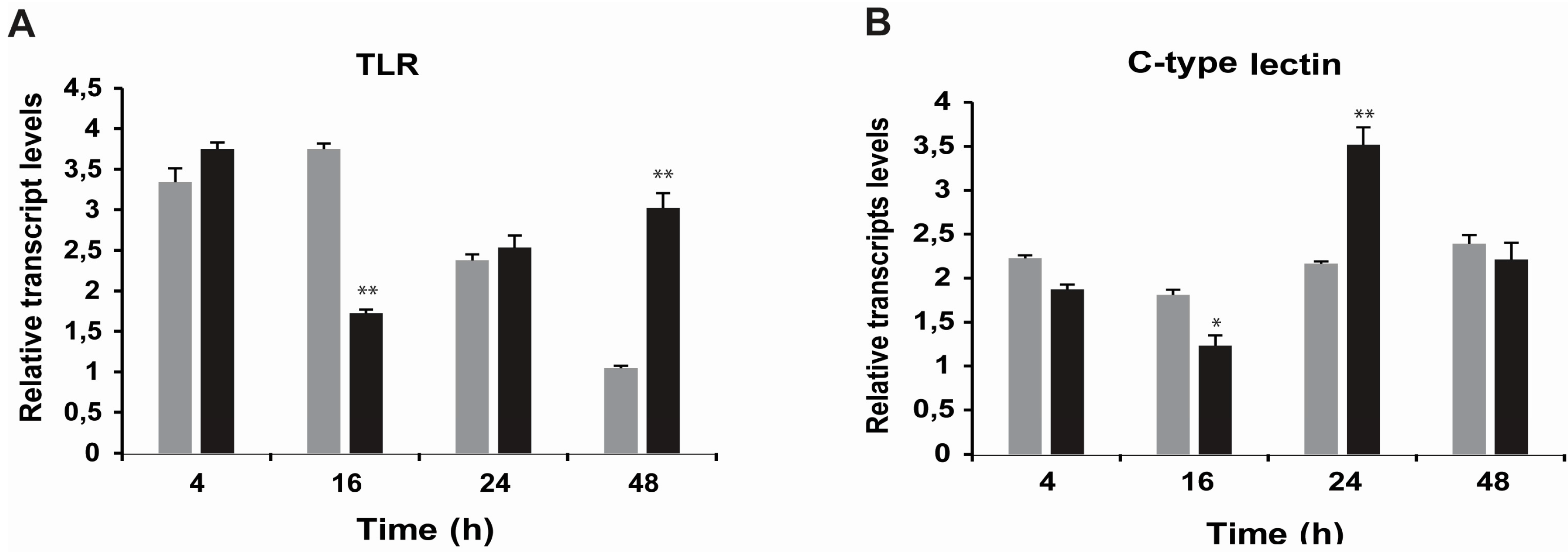
2.3. Effects of STX on the Gene Expression of Antioxidant Enzymes, Oxidative Phosphorylation Transcripts, Ion Channels and Pattern Recognition Receptors
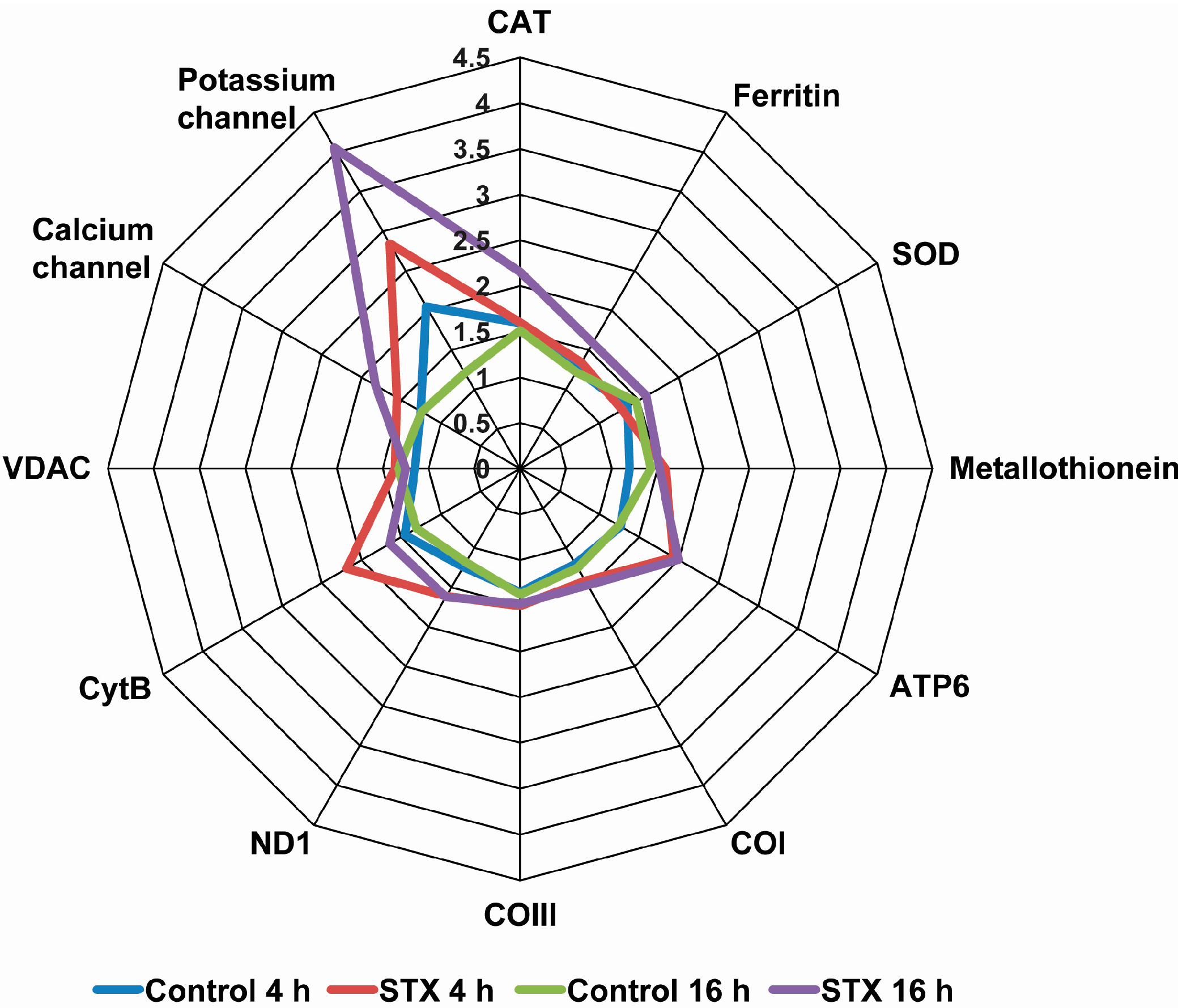
3. Discussion
3.1. Characterization of Hemocyte Primary Cell Cultures
3.2. Cellular Response of Hemocytes to STX Exposure
3.3. Effects of STX on Gene Transcription in M. chilensis Hemocytes
3.4. Use of Primary Hemocyte Cultures in Toxicological Studies
4. Experimental Section
4.1. Primary Culture of Hemocytes
4.2. Experimental Design
4.3. Viability and Phagocytosis Assays Using Light and Fluorescent Microscopy
4.4. Hemocyte Phagocytosis Using Flow Cytometry
4.5. Intracellular ROS Production Using Fluorescent Microscopy
4.6. Transcription Expression of Candidate Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers (5ʹ–3ʹ) | Reference |
|---|---|---|
| Catalase | CCAGGTGTCCTTCCTGTTTTCT | Núñez-Acuña et al., 2012 [60] |
| TGTCCATCCTTGTTGACTGTCTTAA | ||
| Ec-SOD Cu/Zn | TCGCTTTCAGTCAACAGAATGG | Núñez-Acuña et al., 2012 [60] |
| CCAAACTCGTGAACGTGGAA | ||
| Ferritin | CAAGTGAACGCCATCAAGGA | Núñez-Acuña et al., 2012 [60] |
| CGTTGATGCTCTCTTTGTCATACA | ||
| Metallothionein | CTGTGGTGACGCCTGCAAGT | Núñez-Acuña et al., 2013 [20] |
| CTGTGGTGACGCCTGCAAGT | ||
| NADH dehydrogenase I | CTTACTAGGTGCAGTCCGTG | Primers designed in this study |
| ACATAAACACGCCTGAAGATAGT | ||
| ATP6 | TGGGCATACCGCTGTGACTTACT | Primers designed in this study |
| GTGCAGGGGCACCCATAGGA | ||
| Cytochrome oxidase subunit I | TCAGCTGCTGTAGAGAGTGGGG | Primers designed in this study |
| TCACGCCGGTGGTTCAGTTG | ||
| Cytochrome oxidase subunit III | TATGTACCAGGCCCAAGTCC | Primers designed in this study |
| AAAATCTGGGGGTTCGATGC | ||
| Cytochrome B | GGAAGATGCCCGTTGGAGGC | Primers designed in this study |
| CCCTGCGCTCCAGAAATTATGGG | ||
| Voltage-dependent anion channel | CAGACAGGAAAGAAGCAGGG | Primers designed in this study |
| CTAAGGCACCATTCACAGCT | ||
| Calcium channel | TGTAAGCAAGTCGTGTGGGGCAC | Primers designed in this study |
| ACGAGTTTGGGCAGACTATGATCCTG | ||
| Potassium channel | CGGCAATTTGGACCGGGCTTGTG | Primers designed in this study |
| GCGCCCTCTGTCCATTACTCGGC | ||
| C-type lectin | GATATCGCTGTGCAGAACCA | Primers designed in this study |
| AAACACAAGGCCAGTGGAAA | ||
| TLR | CTTTCTTAGCCACGCCAGAC | Primers designed in this study |
| TGTCATTGTCATTGTCAGTTTCGTCA | ||
| α-Tubulin | GAGCCGTCTGCATGTTGAGC | Núñez-Acuña et al., 2012 [60] |
| TGGACGAAAGCACGTTTGGC |
4.7. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Perovic, S.; Tretter, L.; Brummer, F.; Wetzler, C.; Brenner, J.; Donner, G.; Schroder, H.C.; Muller, W.E. Dinoflagellates from marine algal blooms produce neurotoxic compounds: Effects on free calcium levels in neuronal cells and synaptosomes. Environ. Toxicol. Pharmacol. 2000, 8, 83–94. [Google Scholar] [CrossRef]
- Hegaret, H.; da Silva, P.M.; Sunila, I.; Shumway, S.E.; Dixon, M.S.; Alix, J.; Wikfors, G.H.; Soudant, P. Perkinsosis in the Manila clam Ruditapes philippinarum affects responses to the harmful-alga, Prorocentrum minimum. J. Exp. Mar. Biol. Ecol. 2009, 371, 112–120. [Google Scholar] [CrossRef]
- Estrada, N.; Rodriguez-Jaramillo, C.; Contreras, G.; Ascencio, F. Effects of induced paralysis on hemocytes and tissues of the giant lions-paw scallop by paralyzing shellfish poison. Mar. Biol. 2010, 157, 1401–1415. [Google Scholar] [CrossRef]
- Navarro, J.M.; Contreras, A.M. An integrative response by Mytilus chilensis to the toxic dinoflagellate Alexandrium catenella. Mar. Biol. 2010, 157, 1967–1974. [Google Scholar] [CrossRef]
- Hegaret, H.; da Silva, P.M.; Wikfors, G.H.; Lambert, C.; de Bettignies, T.; Shumway, S.E.; Soudant, P. Hemocyte responses of Manila clams, Ruditapes philippinarum, with varying parasite, Perkinsus olseni, severity to toxic-algal exposures. Aquat. Toxicol. 2007, 84, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Shumway, S.E. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Twarog, B.M.; Hidaka, T.; Yamaguchi, H. Resistance to tetrodotoxin and saxitoxin in nerves of bivalve molluscs. A possible correlation with paralytic shellfish poisoning. Toxicon 1972, 10, 273–278. [Google Scholar] [CrossRef]
- MacQuarrie, S.P.; Bricelj, V.M. Behavioral and physiological responses to PSP toxins in Mya arenaria populations in relation to previous exposure to red tides. Mar. Ecol. Prog. Ser. 2008, 366, 59–74. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D.; Anderson, D.M. Uptake kinetics of paralytic shellfish toxins from the dinoflagellate Alexandrium fundyense in the mussel Mytilus edulis. Mar. Ecol. Prog. Ser. 1990, 63, 177–188. [Google Scholar] [CrossRef]
- Shumway, S.E.; Cucci, T.L. The effects of the toxic dinoflagellate Protogonyaulax tamarensis on the feeding and behaviour of bivalve molluscs. Aquat. Toxicol. 1987, 10, 9–27. [Google Scholar] [CrossRef]
- Shumway, S.E.; Pierce, F.C.; Knowlton, K. The effect of Protogonyaulax tamarensis on byssus production in Mytilus edulis L., Modiolus modiolus linnaeus, 1758 and Geukensia demissa dillwyn. Comp. Biochem. Physiol. A Physiol. 1987, 10, 1021–1023. [Google Scholar] [CrossRef]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramón, M.; Wikfors, G.H. Experimental exposure of the blue mussel (Mytilus edulis, L.) to the toxic dinoflagellate Alexandrium fundyense: Histopathology, immune responses, and recovery. Harmful Algae 2008, 7, 702–711. [Google Scholar] [CrossRef]
- Hegaret, H.; Smolowitz, R.M.; Sunila, I.; Shumway, S.E.; Alix, J.; Dixon, M.; Wikfors, G.H. Combined effects of a parasite, QPX, and the harmful-alga, Prorocentrum minimum on northern quahogs, Mercenaria mercenaria. Mar. Environ. Res. 2010, 69, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Haberkorn, H.; Lambert, C.; le Goic, N.; Gueguen, M.; Moal, J.; Palacios, E.; Lassus, P.; Soudant, P. Effects of Alexandrium minutum exposure upon physiological and hematological variables of diploid and triploid oysters, Crassostrea gigas. Aquat. Toxicol. 2010, 97, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, T.C. Hemocytes: Forms and functions. In The Eastern Oyster: Crassostrea Virginica; Kennedy, V.S., Newell, R.I., Eble, A.F., Eds.; University of Maryland Sea Grant Publications: College Park, MD, USA, 1996; pp. 299–333. [Google Scholar]
- Mello, D.F.; da Silva, P.M.; Barracco, M.A.; Soudant, P.; Hegaret, H. Effects of the dinoflagellate Alexandrium minutum and its toxin (saxitoxin) on the functional activity and gene expression of Crassostrea gigas hemocytes. Harmful Algae 2013, 26, 45–51. [Google Scholar] [CrossRef]
- Mello, D.F.; de Oliveira, E.S.; Vieira, R.C.; Simoes, E.; Trevisan, R.; Dafre, A.L.; Barracco, M.A. Cellular and transcriptional responses of Crassostrea gigas hemocytes exposed in vitro to brevetoxin (PbTx-2). Mar. Drugs 2012, 10, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Prado-Alvarez, M.; Florez-Barros, F.; Mendez, J.; Fernandez-Tajes, J. Effect of okadaic acid on carpet shell clam (Ruditapes decussatus) haemocytes by in vitro exposure and harmful algal bloom simulation assays. Cell Biol. Toxicol. 2013, 29, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Mat, A.M.; Haberkorn, H.; Bourdineaud, J.P.; Massabuau, J.C.; Tran, D. Genetic and genotoxic impacts in the oyster Crassostrea gigas exposed to the harmful alga Alexandrium minutum. Aquat. Toxicol. 2013, 140–141, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Acuña, G.; Aballay, A.E.; Hégaret, H.; Astuya, A.P.; Gallardo-Escárate, C. Transcriptional responses of Mytilus chilensis exposed in vivo to saxitoxin (STX). J. Molluscan Stud. 2013, 79, 323–331. [Google Scholar] [CrossRef]
- Manfrin, C.; de Moro, G.; Torboli, V.; Venier, P.; Pallavicini, A.; Gerdol, M. Physiological and molecular responses of bivalves to toxic dinoflagellates. Invertebr. Surival J. 2012, 9, 184–199. [Google Scholar]
- Suárez-Ulloa, V.; Fernández-Tajes, J.; Manfrin, C.; Gerdol, M.; Venier, P.; Eirín-López, J.M. Bivalve omics: State of the art and potential applications for the biomonitoring of harmful marine compounds. Mar. Drugs 2013, 11, 4370–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera-Belmonte, A.; Inostroza, I.; Franco, J.M.; Riobo, P.; Gomez, P.I. The growth, toxicity and genetic characterization of seven strains of Alexandrium catenella (Whedon and Kofoid) Balech 1985 (Dinophyceae) isolated during the 2009 summer outbreak in southern Chile. Harmful Algae 2011, 12, 105–112. [Google Scholar] [CrossRef]
- Varela, D.; Paredes, J.; Alves-de-Souza, C.; Seguel, M.; Sfeir, A.; Frangópulos, M. Intraregional variation among Alexandrium catenella (Dinophyceae) strains from southern Chile: Morphological, toxicological and genetic diversity. Harmful Algae 2012, 15, 8–18. [Google Scholar] [CrossRef]
- Jedlicki, A.; Fernandez, G.; Astorga, M.; Oyarzun, P.; Toro, J.E.; Navarro, J.M.; Martinez, V. Molecular detection and species identification of Alexandrium (Dinophyceae) causing harmful algal blooms along the Chilean coastline. AoB Plants 2012, 2012, pls033. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y. Postcolumn derivatization liquid-chromatographic method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar]
- Navarro, J.M.; Aguila, B.L.; Machmar, F.; Chaparro, O.R.; Contreras, A.M. Dynamic of intoxication and detoxification in juveniles of Mytilus chilensis (Bivalvia: Mytilidae) exposed to paralytic shellfish toxins. Aquat. Living Resour. 2011, 24, 93–98. [Google Scholar] [CrossRef]
- Brousseau, P.; Pellerin, J.; Morin, Y.; Cyr, D.; Blakley, B.; Boermans, H.; Fournier, M. Flow cytometry as a tool to monitor the disturbance of phagocytosis in the clam Mya arenaria hemocytes following in vitro exposure to heavy metals. Toxicology 2000, 142, 145–156. [Google Scholar] [CrossRef]
- Cao, A.; Mercado, L.; Ramos-Martinez, J.I.; Barcia, R. Primary cultures of hemocytes from Mytilus galloprovincialis Lmk.: Expression of IL-2Rα subunit. Aquaculture 2003, 216, 1–8. [Google Scholar] [CrossRef]
- Lebel, J.M.; Giard, W.; Favrel, P.; BoucaudCamou, E. Effects of different vertebrate growth factors on primary cultures of hemocytes from the gastropod mollusc, Haliotis tuberculata. Biol. Cell 1996, 86, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Cajaraville, M.P.; Pal, S.G. Morphofunctional study of the haemocytes of the bivalve mollusc Mytilus galloprovincialis with emphasis on the endolysosomal compartment. Cell Struct. Funct. 1995, 20, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Hine, P.M. The inter-relationships of bivalve haemocytes. Fish Shellfish Immunol. 1999, 9, 367–385. [Google Scholar] [CrossRef]
- Le Foll, F.; Rioult, D.; Boussa, S.; Pasquier, J.; Dagher, Z.; Leboulenger, F. Characterisation of Mytilus edulis hemocyte subpopulations by single cell time-lapse motility imaging. Fish Shellfish Immunol. 2010, 28, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, M.; Chiang, M.W.; Shin, P.K.; Cheung, S.G. Characterization of subpopulations and immune-related parameters of hemocytes in the green-lipped mussel Perna viridis. Fish Shellfish Immunol. 2012, 32, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Ford, S.E.; Bricelj, V.M.; Lambert, C.; Paillard, C. Deleterious effects of a nonPST bioactive compound(s) from Alexandrium tamarense on bivalve hemocytes. Mar. Biol. 2008, 154, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Romestand, B.; Torreilles, J.; Roch, P. In vitro production of superoxide and nitric oxide (as nitrite and nitrate) by Mytilus galloprovincialis haemocytes upon incubation with PMA or laminarin or during yeast phagocytosis. Eur. J. Cell Biol. 2000, 79, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Hegaret, H.; Wikfors, G.H.; Soudant, P. Flow cytometric analysis of haemocytes from eastern oysters, Crassostrea virginica, subjected to a sudden temperature elevation II. Haemocyte functions: Aggregation, viability, phagocytosis, and respiratory burst. J. Exp. Mar. Biol. Ecol. 2003, 293, 249–265. [Google Scholar] [CrossRef]
- Lambert, C.; Soudant, P.; Choquet, G.; Paillard, C. Measurement of Crassostrea gigas hemocyte oxidative metabolism by flow cytometry and the inhibiting capacity of pathogenic vibrios. Fish Shellfish Immunol. 2003, 15, 225–240. [Google Scholar] [CrossRef]
- Lemasters, J.J.; Holmuhamedov, E. Voltage-dependent anion channel (VDAC) as mitochondrial governator—Thinking outside the box. Biochim. Biophys. Acta 2006, 1762, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Schild, L.; Moczydlowski, E.G. Use-dependent block of the voltage-gated Na+ channel by tetrodotoxin and saxitoxin: Effect of pore mutations that change ionic selectivity. J. Gen. Physiol. 2012, 140, 435–454. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Song, X.; Zhao, J.; Mu, C.; Wang, L.; Zhang, H.; Zhou, Z.; Liu, X.; Song, L. A C-type lectin (AiCTL-3) from bay scallop Argopecten irradians with mannose/galactose binding ability to bind various bacteria. Gene 2013, 531, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, C.; Ma, C.; Li, H.; Zuo, H.; Weng, S.; Chen, X.; Zeng, D.; He, J.; Xu, X. Identification of a C-type lectin with antiviral and antibacterial activity from pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2014, 46, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, M.; McKenna, P.; Gauthier-Clerc, S.; Pellerin, J.; Danger, J.M.; Siah, A. Sequence analysis of a normalized cDNA library of Mytilus edulis hemocytes exposed to Vibrio splendidus LGP32 strain. Results Immunol. 2013, 3, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Toubiana, M.; Gerdol, M.; Rosani, U.; Pallavicini, A.; Venier, P.; Roch, P. Toll-like receptors and MyD88 adaptors in Mytilus: Complete cds and gene expression levels. Dev. Comp. Immunol. 2013, 40, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Duchemin, M.B.; Auffret, M.; Wessel, N.; Fortier, M.; Morin, Y.; Pellerin, J.; Fournier, M. Multiple experimental approaches of immunotoxic effects of mercury chloride in the blue mussel, Mytilus edulis, through in vivo, in tubo and in vitro exposures. Environ. Pollut. 2008, 153, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, B.; Thomas-Guyon, H.; Renault, T. In vitro effects of cadmium and mercury on Pacific oyster, Crassostrea gigas (Thunberg), haemocytes. Fish Shellfish Immunol. 2004, 16, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Latire, T.; le Pabic, C.; Mottin, E.; Mottier, A.; Costil, K.; Koueta, N.; Lebel, J.M.; Serpentini, A. Responses of primary cultured haemocytes from the marine gastropod Haliotis tuberculata under 10-day exposure to cadmium chloride. Aquat. Toxicol. 2012, 109, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Mottin, E.; Caplat, C.; Mahaut, M.L.; Costil, K.; Barillier, D.; Lebel, J.M.; Serpentini, A. Effect of in vitro exposure to zinc on immunological parameters of haemocytes from the marine gastropod Haliotis tuberculata. Fish Shellfish Immunol. 2010, 29, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Olabarrieta, I.; L’Azou, B.; Yuric, S.; Cambar, J.; Cajaraville, M.P. In vitro effects of cadmium on two different animal cell models. Toxicol. in Vitro 2001, 15, 511–517. [Google Scholar] [CrossRef]
- Anderson, R.S.; Brubacher, L.L.; Calvo, L.M.R.; Burreson, E.M.; Unger, M.A. Effect of in vitro exposure to tributyltin on generation of oxygen metabolites by oyster hemocytes. Environ. Res. 1997, 74, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, B.; Thomas-Guyon, H.; Burgeot, T.; Renault, T. Pollutant effects on Pacific oyster, Crassostrea gigas (Thunberg), hemocytes: Screening of 23 molecules using flow cytometry. Cell Biol. Toxicol. 2006, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Mendikute, A.; Etxeberria, A.; Olabarrieta, I.; Cajaraville, M.P. Oxygen radicals production and actin filament disruption in bivalve haemocytes treated with benzo(a)pyrene. Mar. Environ. Res. 2002, 54, 431–436. [Google Scholar] [CrossRef]
- Blasco, D.; Levasseur, M.; Bonneau, E.; Gelinas, R.; Packard, T.T. Patterns of paralytic shellfish toxicity in the St. Lawrence region in relationship with the abundance and distribution of Alexandrium tamarense. Sci. Mar. 2003, 67, 261–278. [Google Scholar]
- Cao, A.; Ramos-Martinez, J.I.; Barcia, R. In hemocytes from Mytilus galloprovincialis Lmk., treatment with corticotropin or growth factors conditions catecholamine release. Int. Immunopharmacol. 2007, 7, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Sauvé, S.; Brousseau, P.; Pellerin, J.; Morin, Y.; Senécal, L.; Goudreau, P.; Fournier, M. Phagocytic activity of marine and freshwater bivalves: In vitro exposure of hemocytes to metals (Ag, Cd, Hg and Zn). Aquat. Toxicol. 2002, 58, 189–200. [Google Scholar] [CrossRef]
- Donaghy, L.; Hong, H.K.; Lambert, C.; Park, H.S.; Shim, W.J.; Choi, K.S. The known and unknown sources of reactive oxygen and nitrogen species in haemocytes of marine bivalve molluscs. Fish Shellfish Immunol. 2010, 28, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1–research0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Núñez-Acuña, G.; Gallardo-Escárate, C. Identification of immune-related SNPs in the transcriptome of Mytilus chilensis through high-throughput sequencing. Fish Shellfish Immunol. 2013, 35, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Acuña, G.; Tapia, F.J.; Haye, P.A.; Gallardo-Escárate, C. Gene expression analysis in Mytilus chilensis populations reveals local patterns associated with ocean environmental conditions. J. Exp. Mar. Biol. Ecol. 2012, 420–421, 56–64. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Astuya, A.; Carrera, C.; Ulloa, V.; Aballay, A.; Núñez-Acuña, G.; Hégaret, H.; Gallardo-Escárate, C. Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus chilensis Hemocytes. Int. J. Mol. Sci. 2015, 16, 15235-15250. https://doi.org/10.3390/ijms160715235
Astuya A, Carrera C, Ulloa V, Aballay A, Núñez-Acuña G, Hégaret H, Gallardo-Escárate C. Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus chilensis Hemocytes. International Journal of Molecular Sciences. 2015; 16(7):15235-15250. https://doi.org/10.3390/ijms160715235
Chicago/Turabian StyleAstuya, Allisson, Crisleri Carrera, Viviana Ulloa, Ambbar Aballay, Gustavo Núñez-Acuña, Hélène Hégaret, and Cristian Gallardo-Escárate. 2015. "Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus chilensis Hemocytes" International Journal of Molecular Sciences 16, no. 7: 15235-15250. https://doi.org/10.3390/ijms160715235
APA StyleAstuya, A., Carrera, C., Ulloa, V., Aballay, A., Núñez-Acuña, G., Hégaret, H., & Gallardo-Escárate, C. (2015). Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus chilensis Hemocytes. International Journal of Molecular Sciences, 16(7), 15235-15250. https://doi.org/10.3390/ijms160715235