
Biohydrogen and Bioethanol Production from Biodiesel-Based Glycerol by Enterobacter aerogenes in a Continuous Stir Tank Reactor

Abstract
:1. Introduction
2. Results and Discussion
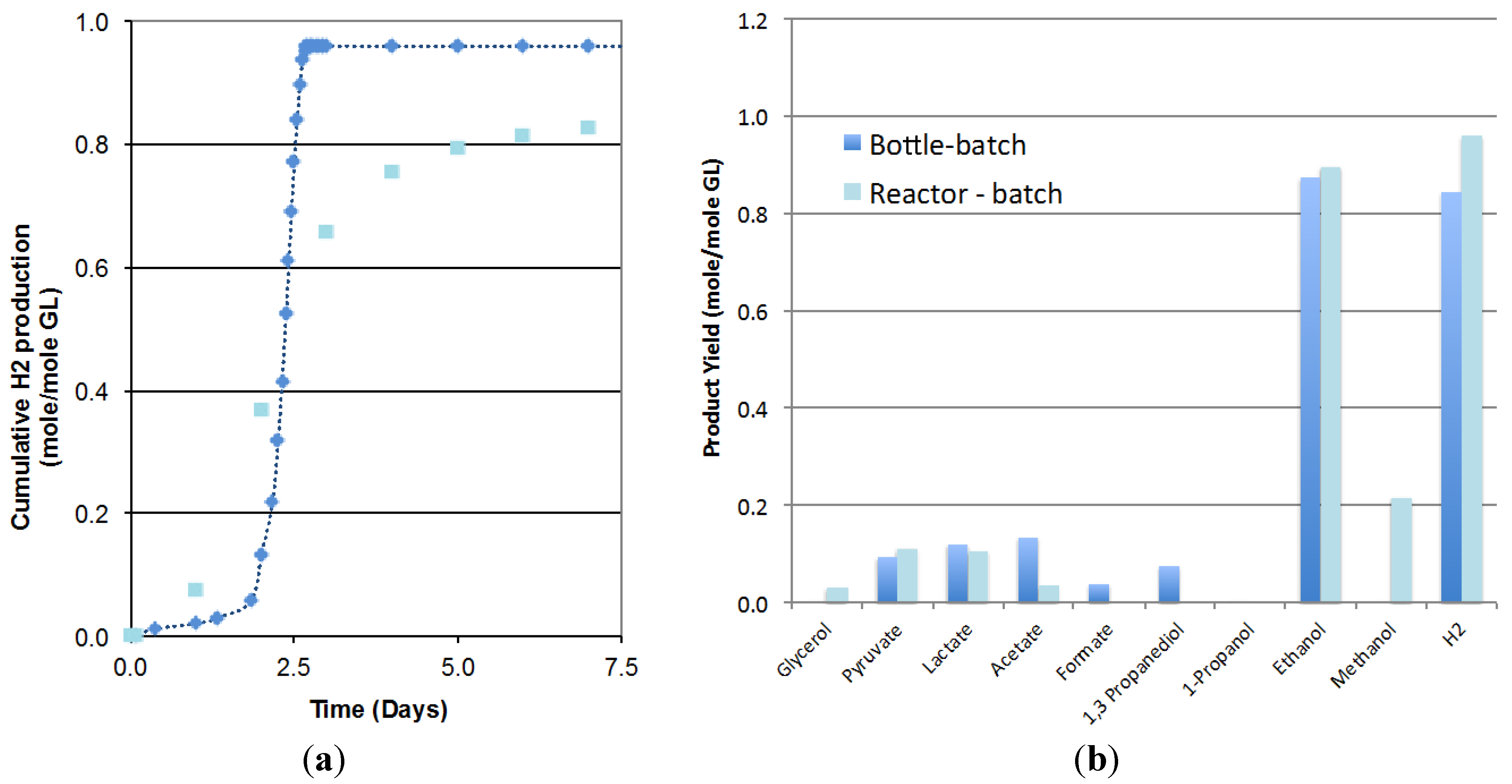
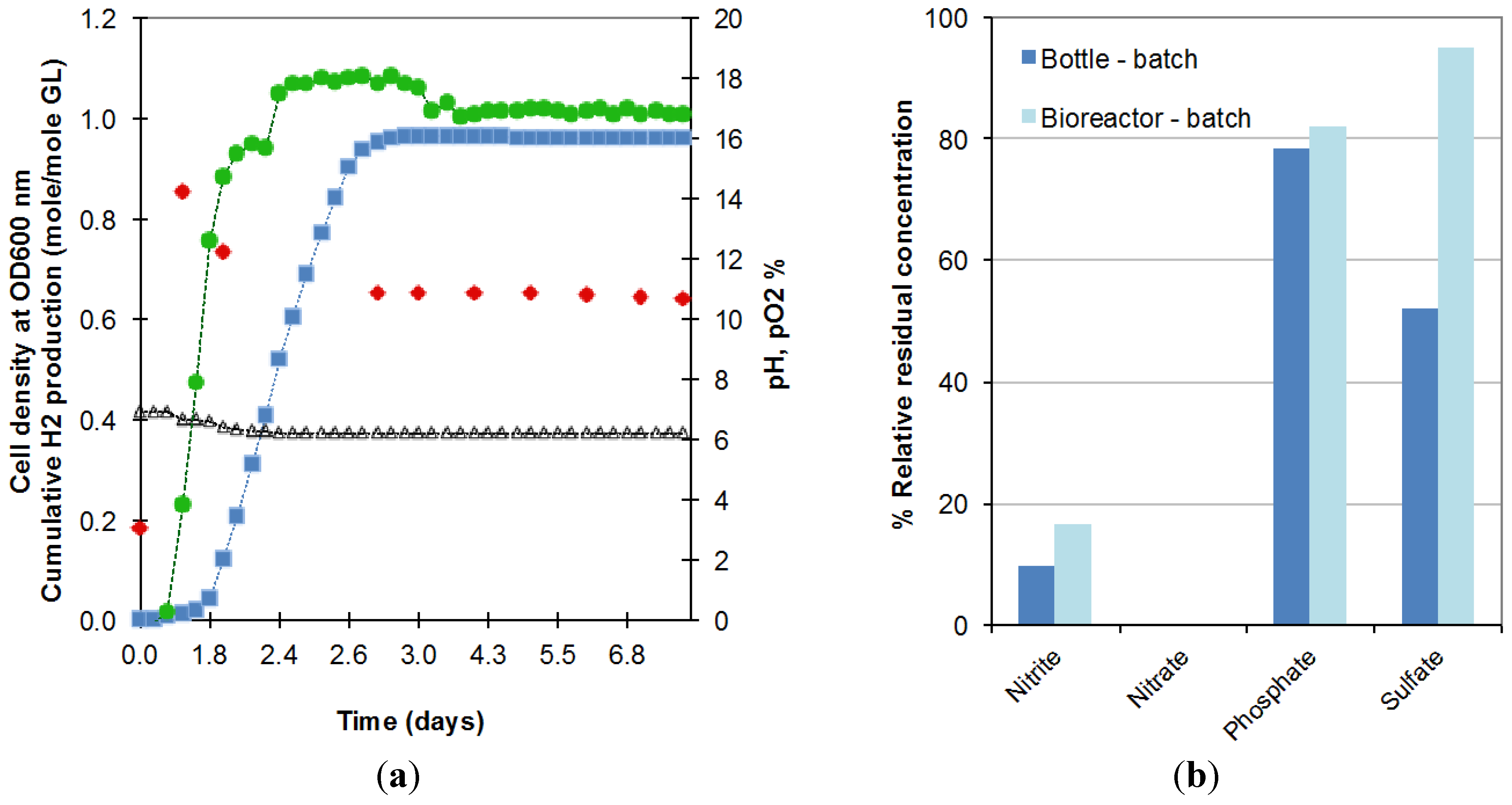
2.1. Scale up of Batch Fermentation
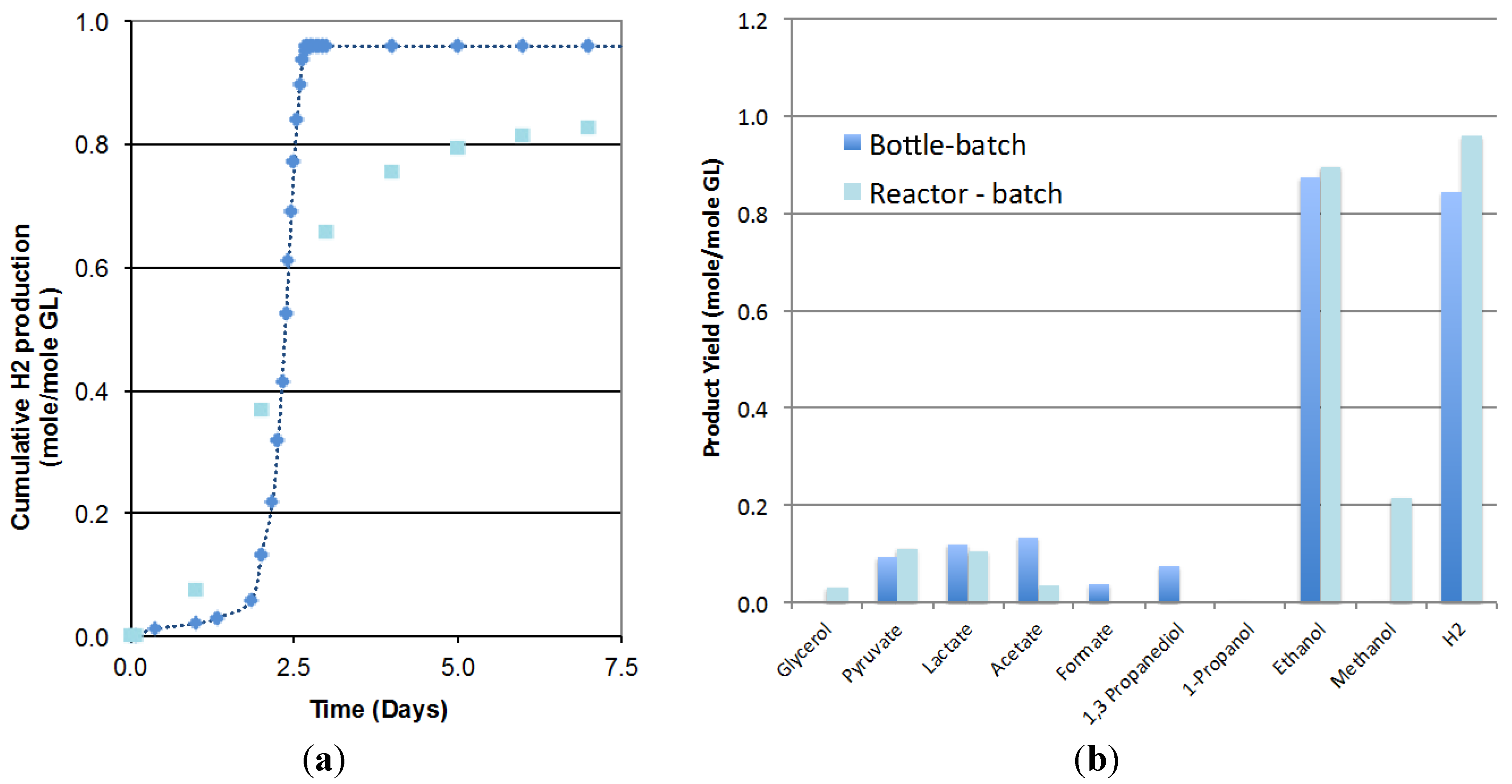
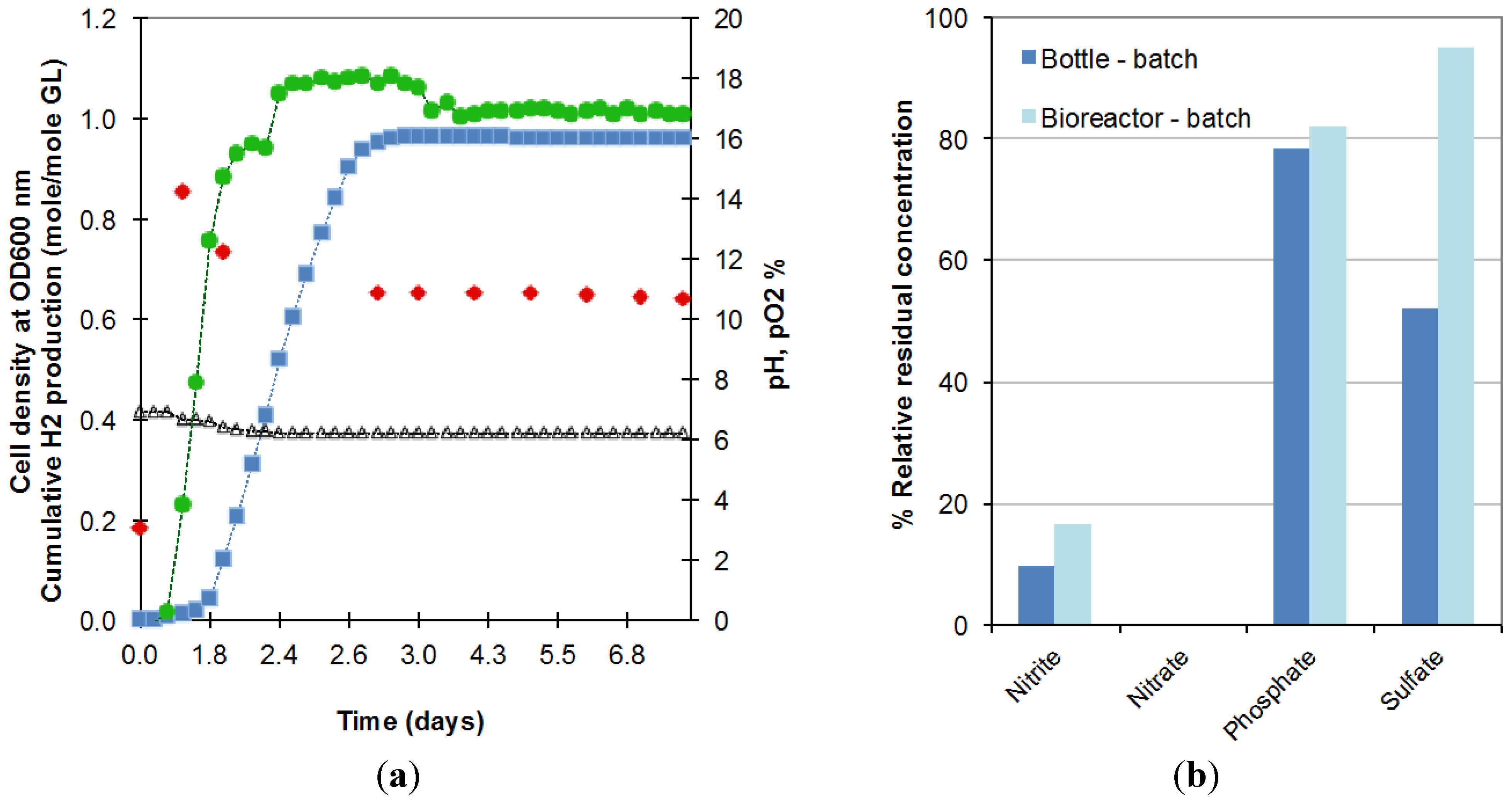
2.2. Optimization of the CSTR
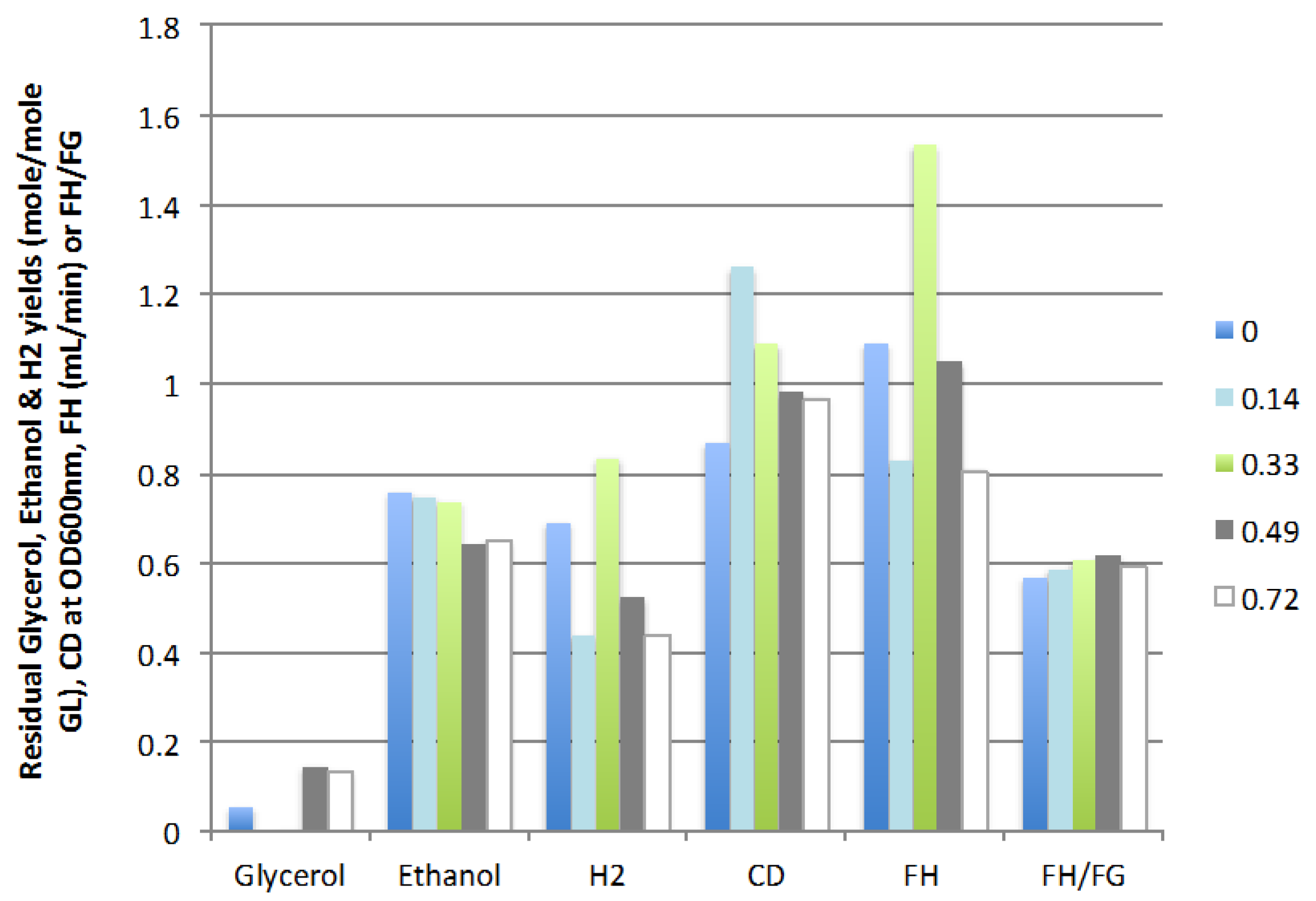
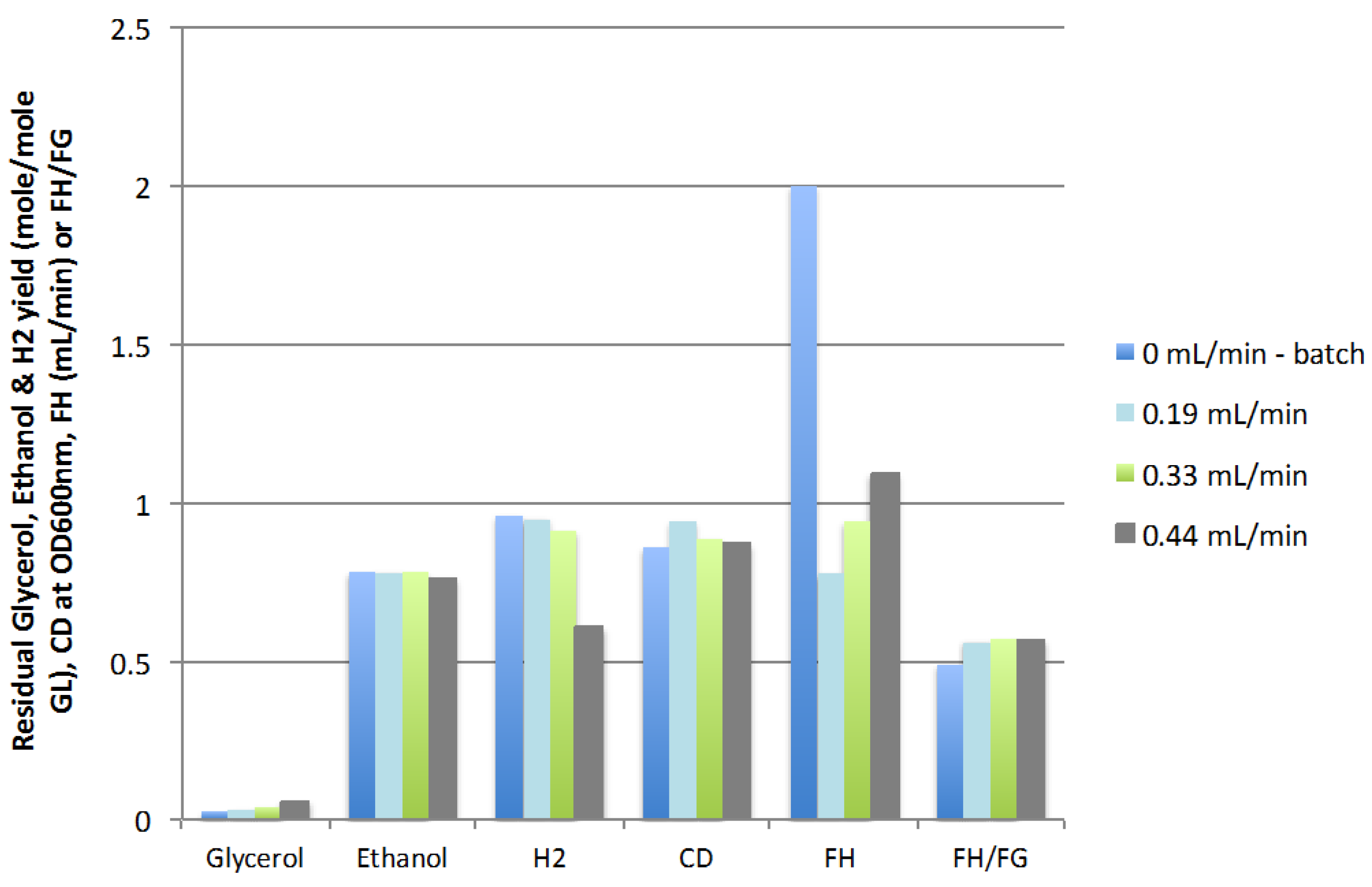
2.2.1. Feed Rate

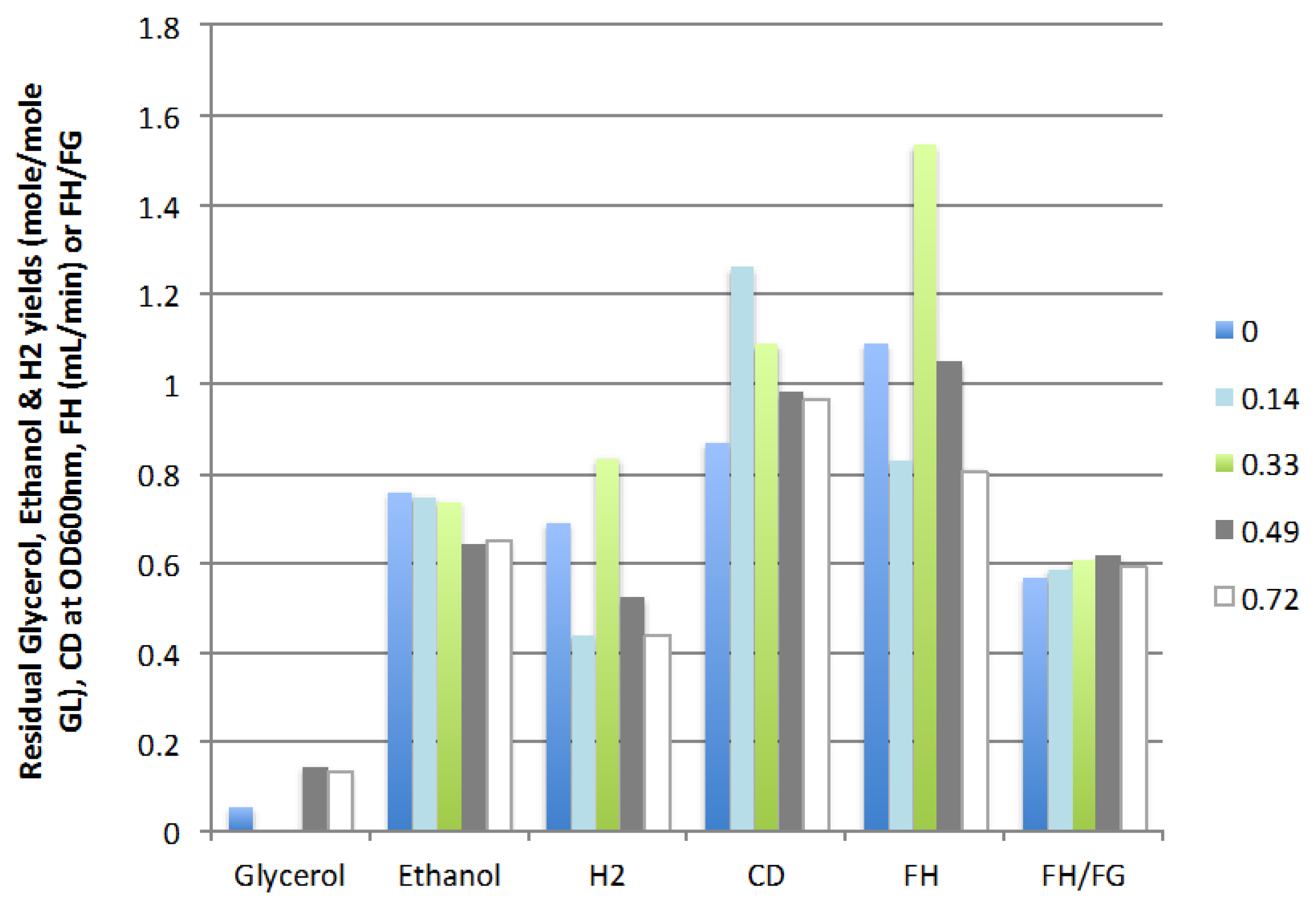
2.2.2. Liquid Recycle Ratio (LR)
2.2.3. pH
2.2.4. Mixing Speed
2.2.5. Glycerol Concentration
2.2.6. Waste Recycling
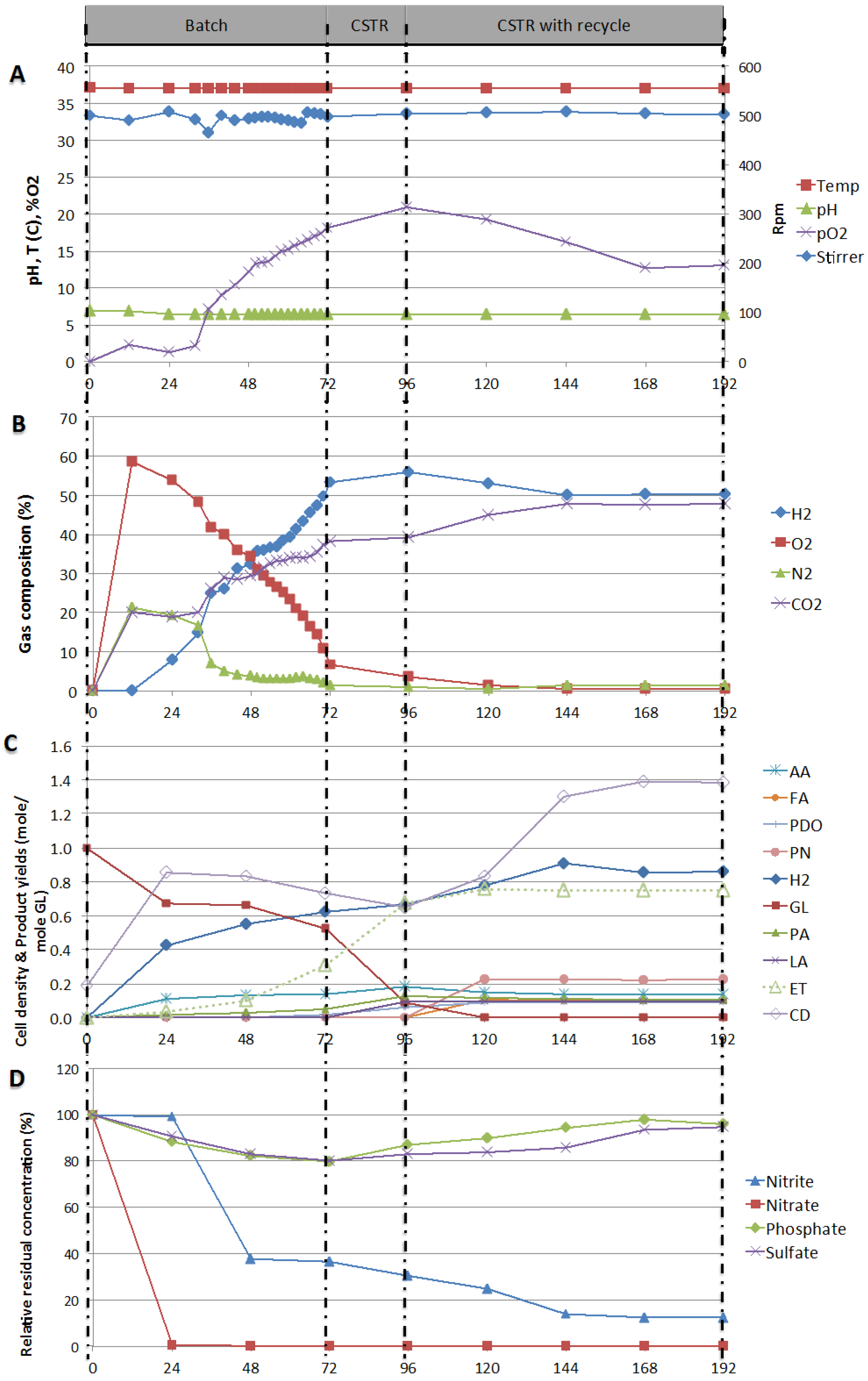
2.3. Testing of Stability of Production in Continuous Mode—Process Stability
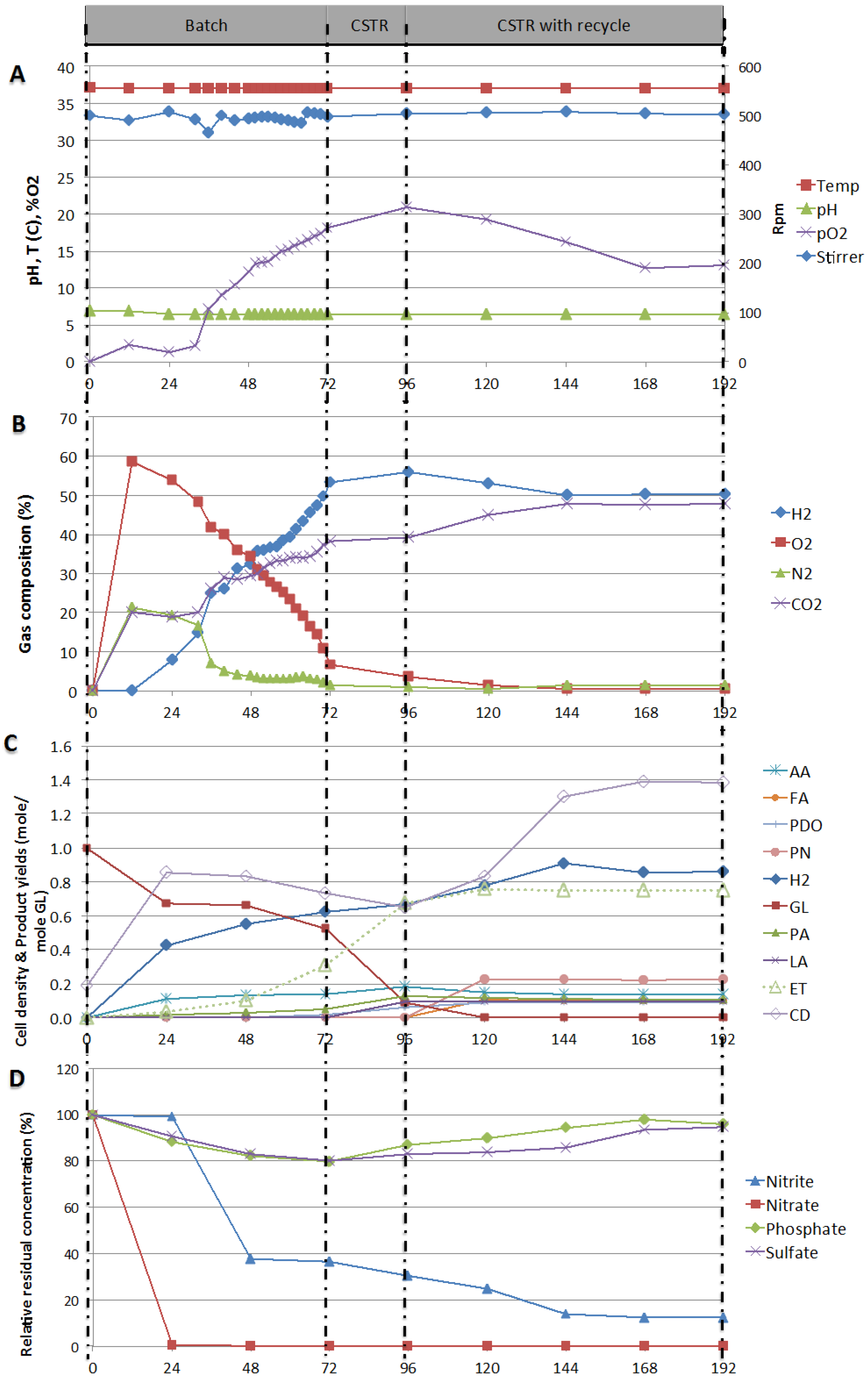
3. Experimental Section
3.1. Microorganism and Inoculum Preparation
3.2. Minimum Mineral Synthetic Media (MMSM) and Glycerol Concentration
3.3. Biohydrogen Production Experiments
3.3.1. Biohydrogen Production in the Experiment Bottles
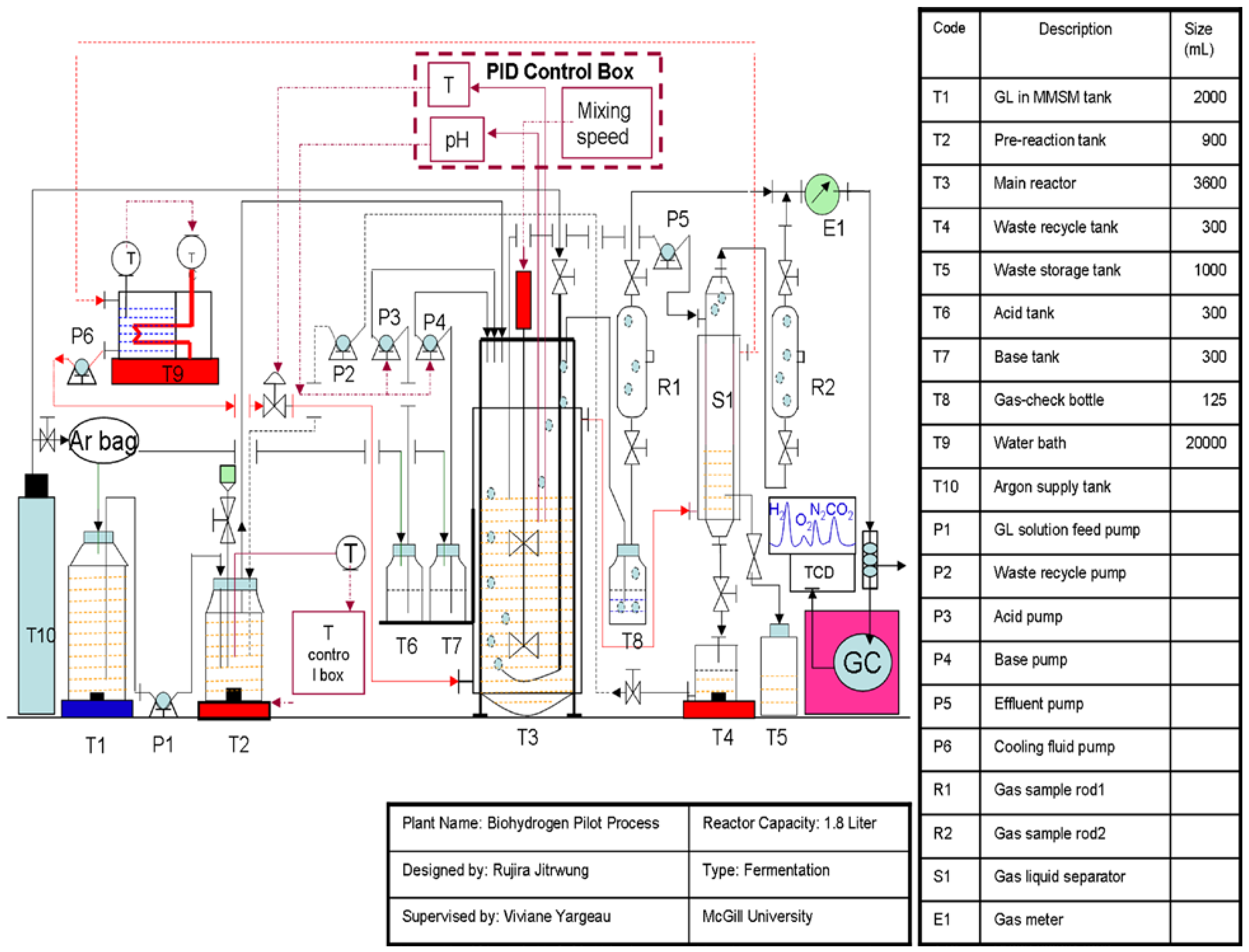
3.3.2. Biohydrogen Production in the Bioreactor
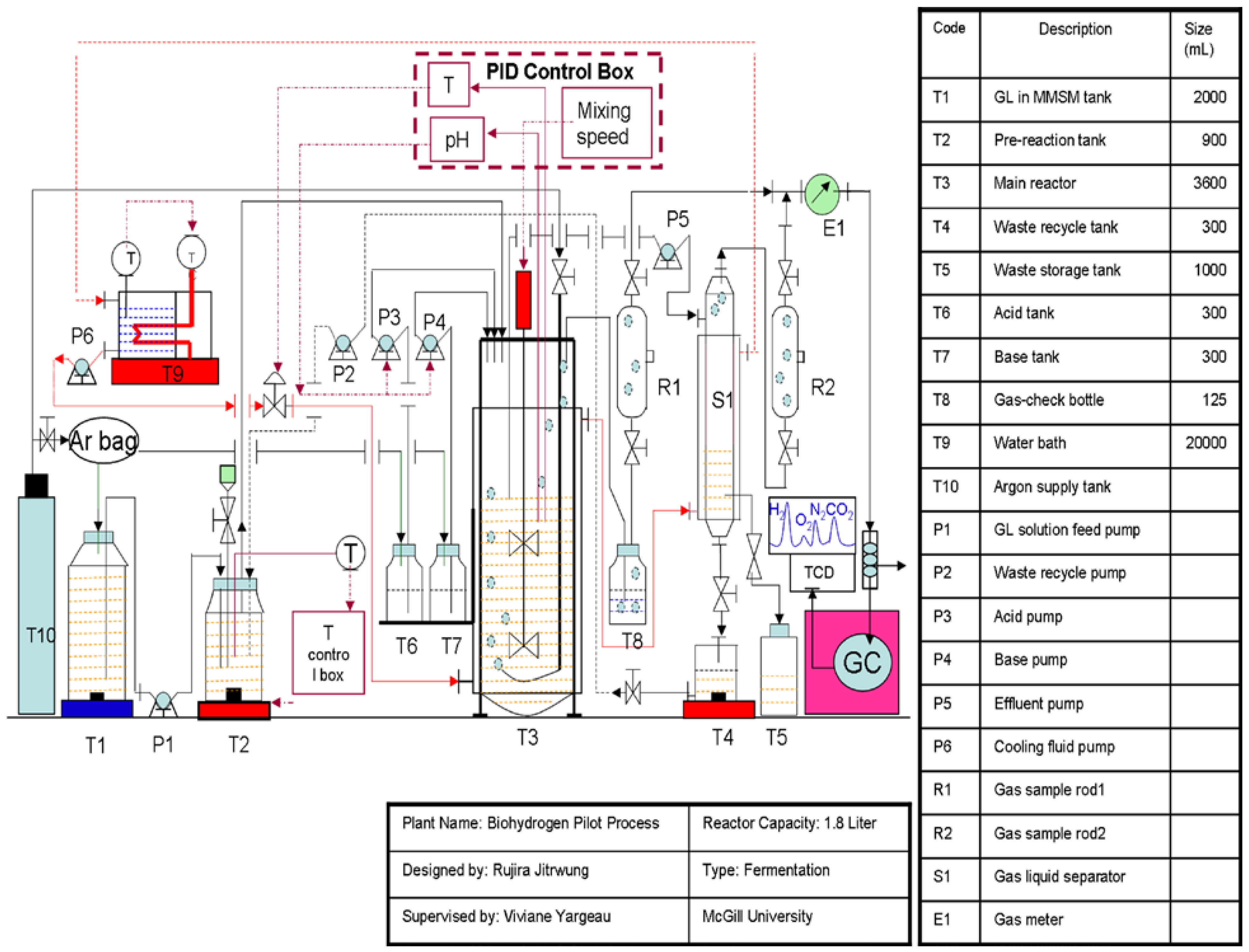
3.4. Analytical Methods
3.4.1. Gas Analysis
3.4.2. Liquid Analysis
3.4.3. Monitoring of Cell Growth
4. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yields (mole/mole GL) | Initial Glycerol Concentration (g/L) | Reactor Type | $CAD/L of Media | Reference | |
|---|---|---|---|---|---|
| H2 | Ethanol | ||||
| 1.12 | 0.96 | 1.7 | Batch | 3.68 | Ito et al. 2005 [20] |
| 0.96 | 0.90 | 15 | Batch–3.6L | 0.91 | This study |
| 0.86 | 0.75 | 15 | CSTR | 0.91 | This study |
| 0.74 | 0.92 | 10 | Bioelectrochemical | Not reported | Sakai and Yagishita 2007 [21] |
| 0.12 | 0.83 | 31 | Batch | Not reported | Reungsang et al. 2013 [38] |
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- (S&T)2 Consultants Inc. Meyers Norris Penny LLP. Economic, Financial, Social Analysis and Public Policies for Biodiesel Phase 1. Avaliable online: http://www.ghgenius.ca/reports/nrcanbiodieselphase1.pdf22 (accessed on 22 November 2004).
- Bonnardeaux, J. Glycerin Overview; Department of Agriculture and Food: Western Australia, Australia, 2006. [Google Scholar]
- Adhikari, S.; Fernando, S.D.; Haryanto, A. Kinetics and reactor modeling of hydrogen production from glycerol via steam reforming process over Ni/Ceo2 catalysts. Chem. Eng. Technol. 2009, 32, 541–547. [Google Scholar] [CrossRef]
- Hirai, T.; Ikenaga, N.-O.; Miyake, T.; Suzuki, T. Production of hydrogen by steam reforming of glycerin on ruthenium catalyst. Energy Fuels 2005, 19, 1761–1762. [Google Scholar] [CrossRef]
- Nandi, R.; Sengupta, S. Microbial production of hydrogen: An overview. Crit. Rev. Microbiol. 1998, 24, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Willke, T.; Vorlop, K.V. Biotransformation of glycerol into 1,3-propanediol. Eur. J. Lipid Sci. Technol. 2008, 110, 831–840. [Google Scholar] [CrossRef]
- Solomon, B.O.; Zeng, A.P.; Biebl, H.; Ejiofor, A.O.; Posten, C.; Deckwer, W.D. Effects of substrate limitation on product distribution and H2/CO2 ratio in klebsiella pnuemoniae during anaerobic fermentation of glycerol. J. Appl. Microbiol. Biotechnol. 1994, 42, 222–226. [Google Scholar]
- Liu, F.; Fang, B. Optimization of bio-hydrogen production from biodiesel wastes by Klebsiella pneumoniae. Biotechnol. J. 2007, 2, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Sabourin-Provost, G.; Hallenbeck, P.C. High yield conversion of a crude glycerol fraction from biodiesel production to hydrogen by photofermentation. Bioresour. Technol. 2009, 100, 3513–3517. [Google Scholar] [CrossRef] [PubMed]
- González-Pajuelo, M.; Meynial-Salles, I.; Mendes, F.; Andrade, J.C.; Vasconcelos, I.; Soucaille, P. Metabolic engineering of clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol. Metab. Eng. 2005, 7, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Biebl, H. Glycerol fermentation of 1,3-propanediol by clostridium butyricum. Measurement of product inhibition by use of a pH-auxostat. Appl. Microbiol. Biotechnol. 1991, 35, 701–705. [Google Scholar]
- Saint-Amans, S.; Perlot, P.; Goma, G.; Soucaille, P. High production of 1,3-propanediol from glycerol by clostridium butyricum vpi 3266 in a simply controlled fed-batch system. Biotechol. Lett. 1994, 16, 831–836. [Google Scholar] [CrossRef]
- Biebl, H. Fermentation of glycerol by clostridium pasteurianum-batch and continuous culture studies. J. Ind. Mocrobiol. Biotechnol. 2001, 27, 18–26. [Google Scholar] [CrossRef]
- Taconi, K.A.; Venkataramanan, K.P.V.; Johnson, D.T. Growth and solvent production by clostridium pasteurianum ATCC®6013™ utilizing biodiesel-derived crude glycerol as the sole carbon source. Environ. Progress Sustain. Energy 2009, 28, 100–110. [Google Scholar] [CrossRef]
- Gonzalez, R.; Murarka, A.; Dharmadi, Y.; Yazdani, S.S. A new model for the anaerobic fermentation of glycerol in enteric bacteria: Trunk and auxiliary pathways in Escherichia coli. Metab. Eng. 2008, 10, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Barbirato, F.; Himmi, E.H.; Conte, T.; Bories, A. 1,3-propanediol production by fermentation: An interesting way to valorize glycerin from the ester and ethanol industries. Ind. Crops Prod. 1998, 7, 281–289. [Google Scholar] [CrossRef]
- Zhang, Z.-P.; Tay, J.-H.; Show, K.-Y.; Yan, R.; Tee Liang, D.; Lee, D.-J.; Jiang, W.-J. Biohydrogen production in a granular activated carbon anaerobic fluidized bed reactor. Int. J. Hydrog. Energy 2007, 32, 185–191. [Google Scholar] [CrossRef]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Jitrwung, R.; Yargeau, V. Optimization of media composition for the production of biohydrogen from waste glycerol. Int. Hydrog. Energy 2011, 36, 9602–9611. [Google Scholar] [CrossRef]
- Ito, T.; Nakashimada, Y.; Senba, K.; Matsui, T.; Nishio, N. Hydrogen and ethanol production from glycerol-containing wastes discharged after biodiesel manufacturing process. J. Biosci. Bioeng. 2005, 100, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Yagishita, T. Microbial production of hydrogen and ethanol from glycerol-containing wastes discharged from a biodiesel fuel production plant in a bioelectrochemical reactor with thionine. Biotechnol. Bioeng. 2007, 98, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.; Waligorska, M.; Wojtowski, M.; Laniecki, M. Hydrogen generation from glycerol in batch fermentation process. Int. J. Hydrog. Energy 2009, 34, 3671–3678. [Google Scholar] [CrossRef]
- Jitrwung, R.; Verrett, J.; Yargeau, V. Improved biohydrogen production from biodiesel-based glycerol using enterobacter aerogenes optimized media salts composition. Int. Hydrog. Energy 2010, 50, 222–226. [Google Scholar]
- Reungsang, A.; Sittijunda, S.; O-thong, S. Bio-hydrogen production from glycerol by immobilized enterobacter aerogenes ATCC 13048 on heat-treated uasb granules as affected by organic loading rate. Int. J. Hydrog. Energy 2013, 38, 6970–6979. [Google Scholar]
- Nwachukwu, R.E.S.; Shahbazi, A.; Wnag, L.; Worku, M.; Ibrahim, S.; Schimmel, K. Optimization of cultural conditions for conversion of glycerol to ethanol by enterobcater aerogenes s012. AMS Express 2013, 2, 12–20. [Google Scholar] [CrossRef]
- Ren, N.; Li, J.; Li, B.; Wang, Y.; Liu, S. Biohydrogen production from molasses by anaerobic fermentation with a pilot-scale bioreactor system. Int. J. Hydrog. Energy 2006, 31, 2147–2157. [Google Scholar] [CrossRef]
- Ren, N.Q.; Chua, H.; Chan, S.Y.; Tsang, Y.F.; Wang, Y.J.; Sin, N. Assessing optimal fermentation type for bio-hydrogen production in continuous-flow acidogenic reactors. Bioresour. Technol. 2007, 98, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.K.; Chen, W.-H.; Li, L.; Sung, S. Biohydrogen production in continuous-flow reactor using mixed microbial culture. Water Environ. Res. 2006, 78, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bruns, M.A.; Logan, B.E. Biological hydrogen production by clostridium acetobutylicum in an unsaturated flow reactor. Water Res. 2006, 40, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Rachman, M.A.; Furutani, Y.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Enhanced hydrogen production in altered mixed acid fermentation of glucose by enterobacter aerogenes. J. Ferment. Bioeng. 1997, 83, 358–363. [Google Scholar] [CrossRef]
- Yokoi, H.; Maki, R.; Hirose, J.; Hayashi, S. Microbial production of hydrogen from starch-manufacturing wastes. Biomass Bioenergy 2002, 22, 389–395. [Google Scholar] [CrossRef]
- Fabiano, B.; Perego, P. Thermodynamic study and optimization of hydrogen production by enterobacter aerogenes. Int. J. Hydrog. Energy 2002, 27, 149–156. [Google Scholar] [CrossRef]
- Jo, J.H.; Lee, D.S.; Park, D.; Choe, W.-S.; Park, J.M. Optimization of key process variables for enhanced hydrogen production by enterobacter aerogenes using statistical methods. Bioresour. Technol. 2008, 99, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Rachman, M.A.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Hydrogen production with high yield and high evolution rate by self-flocculated cells of enterobacter aerogenes in a packed-bed reactor. Appl. Microbiol. Biotechnol. 1998, 49, 450–454. [Google Scholar] [CrossRef]
- Temudo, M.F.; Poldermans, R.; Kleerebezem, R.; van Loosdrecht, M.C.M. Glycerol fermentation by (open) mixed cultures: A chemostat study. Biotechnol. Bioeng. 2008, 100, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Murarka, A.; Dharmadi, Y.; Yazdani, S.S.; Gonzalez, R. Fermentativeutilization of glycerol by Escherichia coli and its implications for the production of fuels and chemicals. Appl. Environ. Microbiol. 2008, 74, 1124–1135. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L.; Wolin, M.J. A serum bottle modification of the hungate technique for cultivating obligate anaerobes. Appl. Microbiol. 1974, 27, 985–987. [Google Scholar] [PubMed]
- Reungsang, A.; Sittijunda, S.; Angelidaki, I. Simultaneous production of hydrogen and ethanol from waste glycerol by enterobacter aerogenes KKU-S1. Int. J. Hydrog. Energy 2013, 38, 1813–1825. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jitrwung, R.; Yargeau, V. Biohydrogen and Bioethanol Production from Biodiesel-Based Glycerol by Enterobacter aerogenes in a Continuous Stir Tank Reactor. Int. J. Mol. Sci. 2015, 16, 10650-10664. https://doi.org/10.3390/ijms160510650
Jitrwung R, Yargeau V. Biohydrogen and Bioethanol Production from Biodiesel-Based Glycerol by Enterobacter aerogenes in a Continuous Stir Tank Reactor. International Journal of Molecular Sciences. 2015; 16(5):10650-10664. https://doi.org/10.3390/ijms160510650
Chicago/Turabian StyleJitrwung, Rujira, and Viviane Yargeau. 2015. "Biohydrogen and Bioethanol Production from Biodiesel-Based Glycerol by Enterobacter aerogenes in a Continuous Stir Tank Reactor" International Journal of Molecular Sciences 16, no. 5: 10650-10664. https://doi.org/10.3390/ijms160510650
APA StyleJitrwung, R., & Yargeau, V. (2015). Biohydrogen and Bioethanol Production from Biodiesel-Based Glycerol by Enterobacter aerogenes in a Continuous Stir Tank Reactor. International Journal of Molecular Sciences, 16(5), 10650-10664. https://doi.org/10.3390/ijms160510650