
Variation and Genetic Structure in Platanus mexicana (Platanaceae) along Riparian Altitudinal Gradient

Abstract
:1. Introduction
2. Results
2.1. Genetic Diversity

{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Size Range | No. of Bands | No. of Polymorphic Bands |
|---|---|---|---|---|
| 818 | (CA)8G | 350–1000 | 12 | 9 |
| 824 | (TC)8G | 350–1500 | 9 | 8 |
| 827 | (AC)8G | 400–1400 | 9 | 8 |
| 829 | (TG)8C | 480–1700 | 7 | 4 |
| 835 | (AG)8YC | 180–1400 | 9 | 7 |
| 841 | (GA)8YC | 300–1200 | 9 | 7 |
| 845 | (CT)8RG | 250–1500 | 12 | 10 |
| 848 | (CA)8RG | 300–1500 | 10 | 9 |
| 850 | (GT)8TYC | 300–1500 | 13 | 11 |
| 857 | (AC)8YG | 180–1500 | 15 | 14 |
| Elevation (m a.s.l.) | He | % p | A | Ne | I |
|---|---|---|---|---|---|
| 70 | 0.228 a | 81.905 a | 1.352 a | 1.378 a | 0.352 a |
| 200 | 0.257 a,b | 88.571 a | 1.543 b | 1.430 a,b | 0.396 a,b |
| 600 | 0.265 a,b | 88.571 a | 1.448 a,b | 1.449 a,b | 0.402 a,b |
| 1700 | 0.279 b | 87.619 a | 1.514 a,b | 1.475 b | 0.424 b |
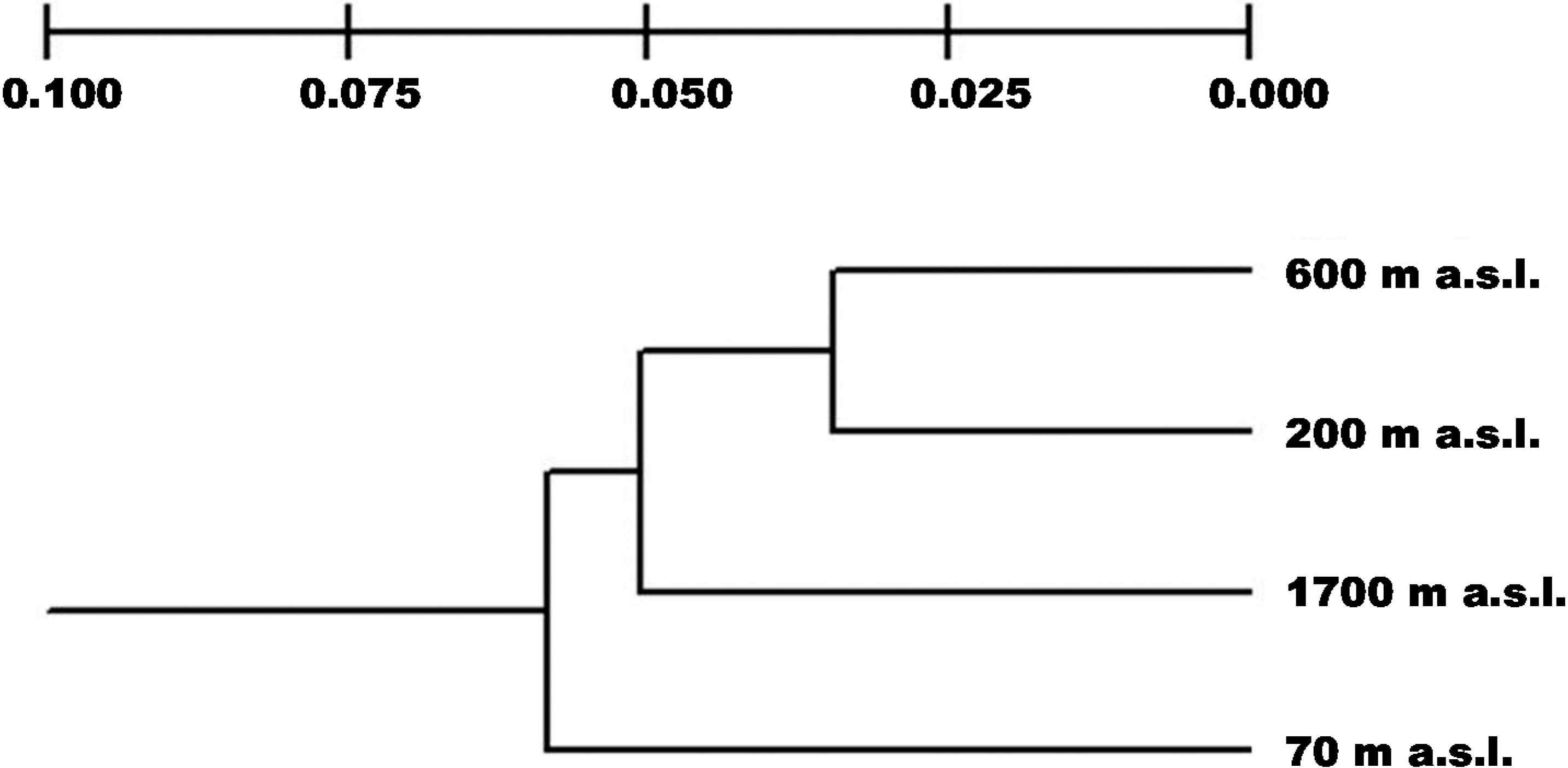
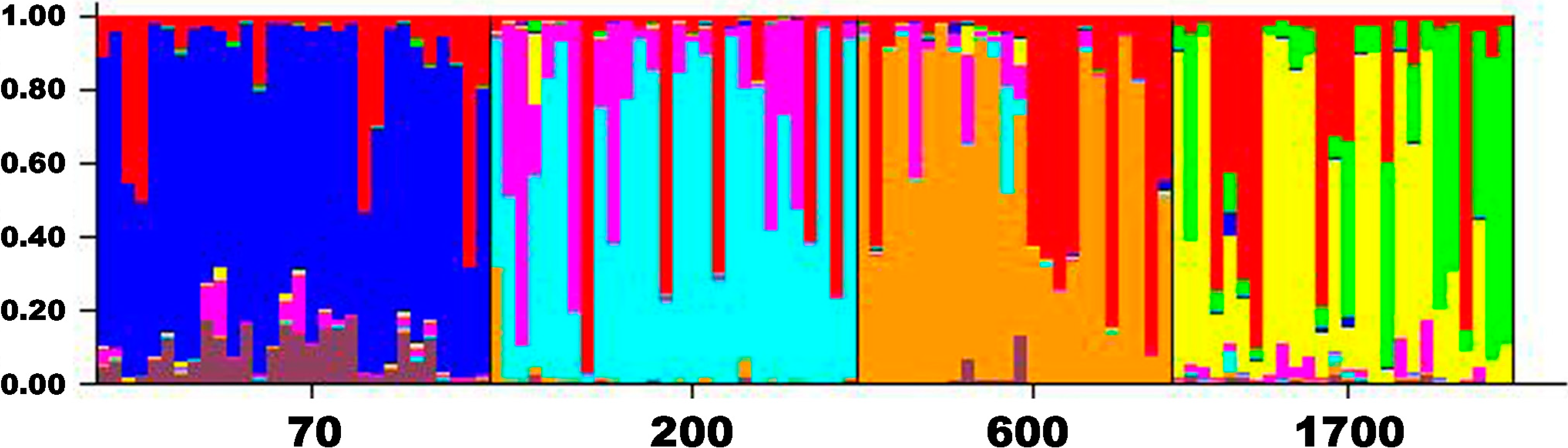
2.2. Genetic Differentiation and Population Structure
| Sites (m a.s.l.) | ΦST |
|---|---|
| 70 & 200 | 0.238 * |
| 70 & 600 | 0.216 * |
| 70 & 1700 | 0.198 * |
| 200 & 600 | 0.166 * |
| 200 & 1700 | 0.176 * |
| 600 & 1700 | 0.169 * |
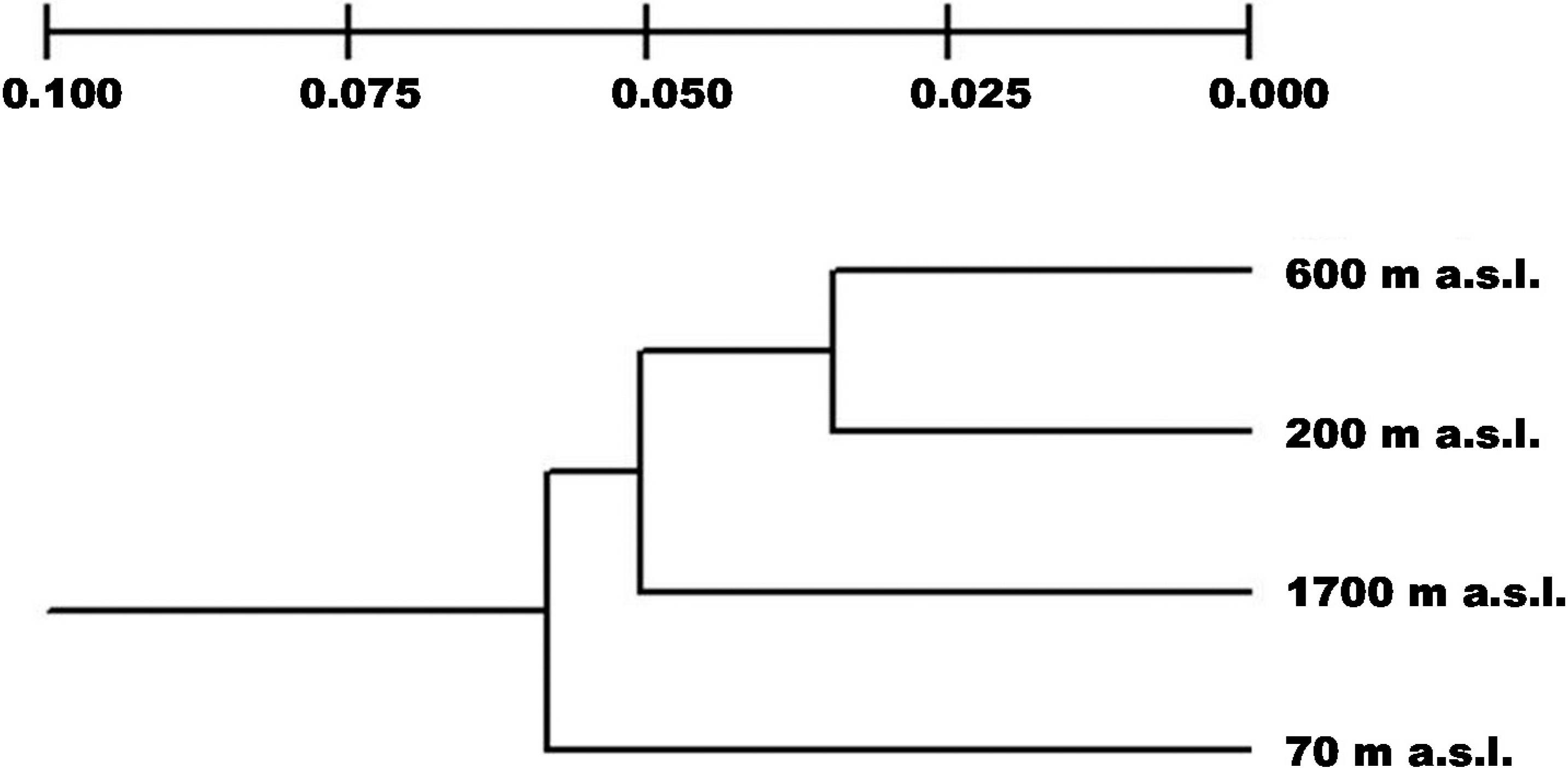
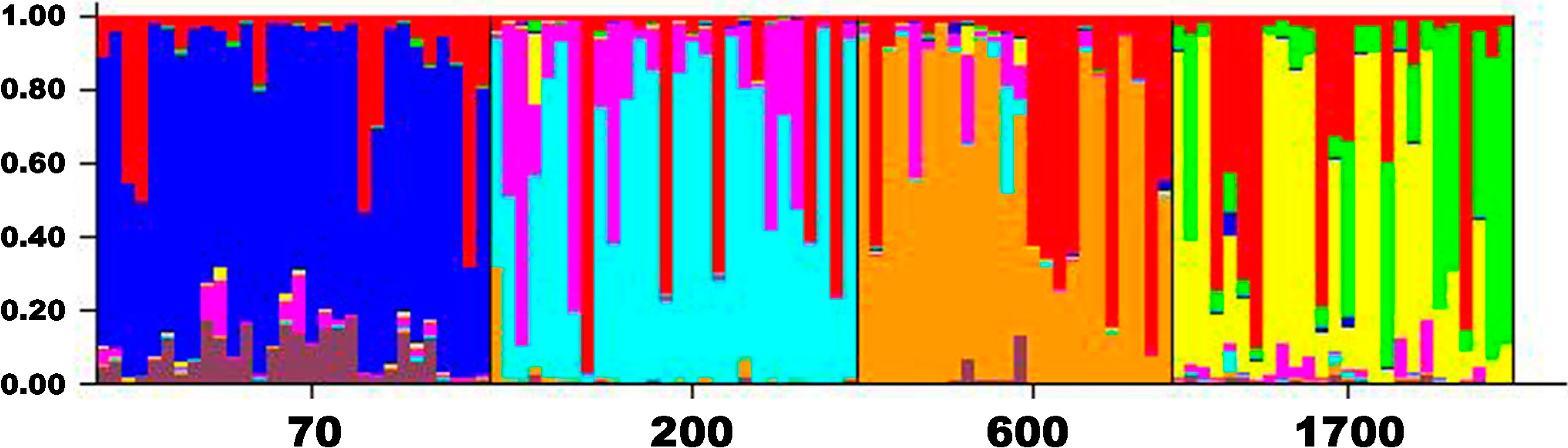
2.3. Genetic Drift
| Sites Compared (m a.s.l.) | Valid N | Z | p Level |
|---|---|---|---|
| 70 & 200 | 54 | 2.292338 | 0.021887 * |
| 70 & 600 | 54 | 1.357806 | 0.174526 |
| 70 & 1700 | 54 | 2.973587 | 0.002944 * |
| 200 & 600 | 54 | 1.158860 | 0.246514 |
| 200 & 1700 | 54 | 0.909226 | 0.363231 |
| 600 & 1700 | 54 | 0.909226 | 0.363231 |
3. Discussion
Genetic Diversity and Differentiation
4. Materials and Methods
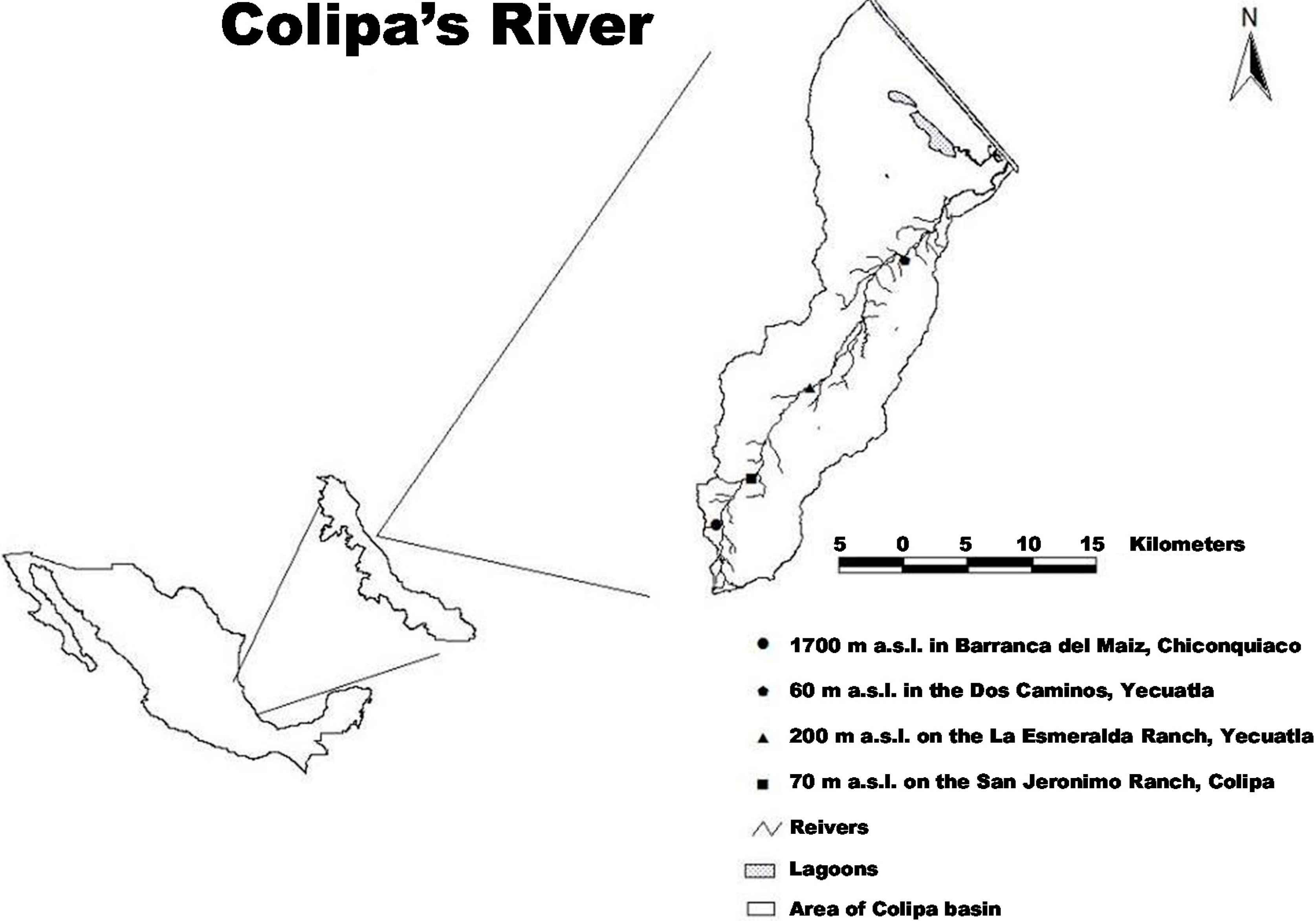
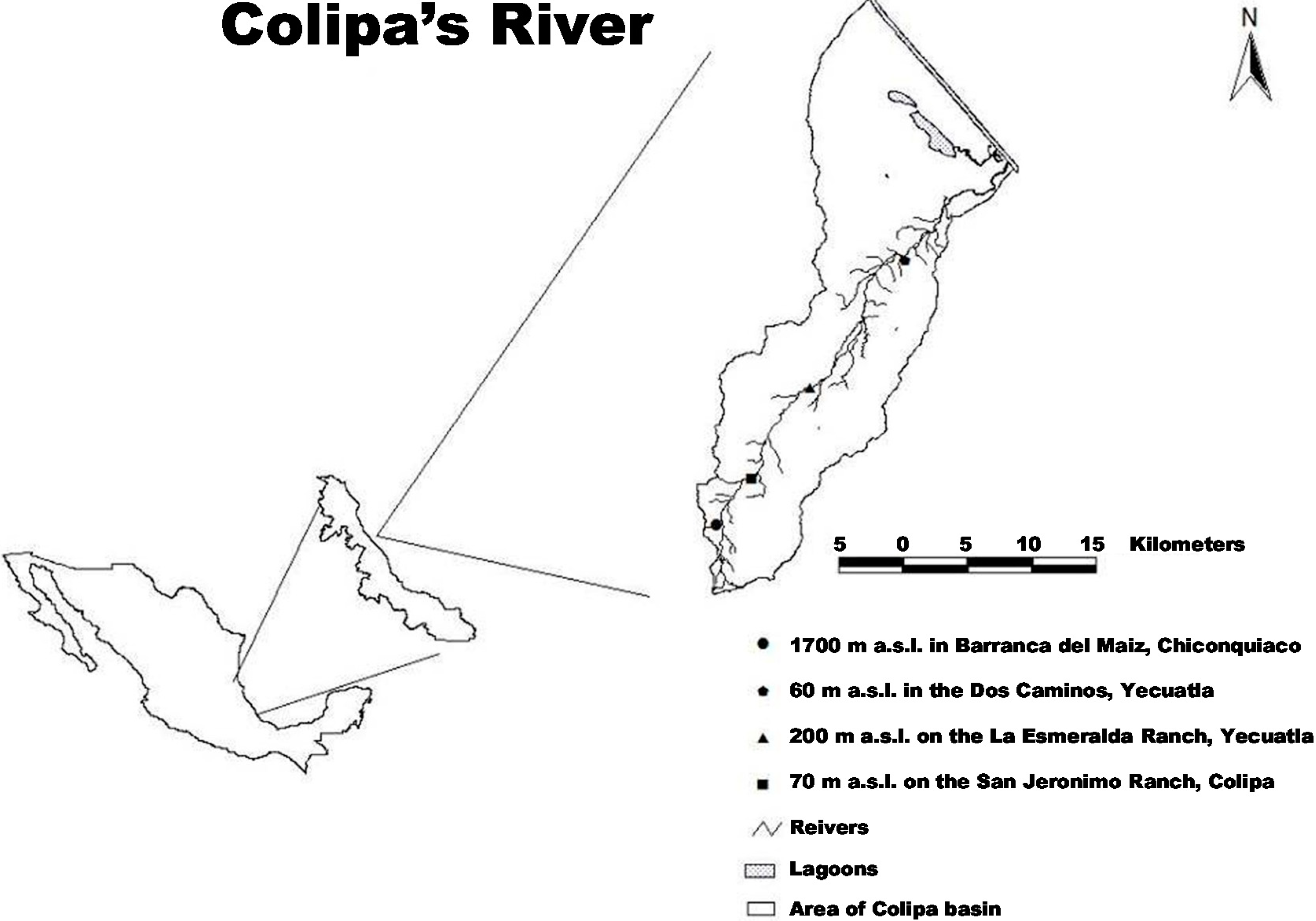
4.1. Study Area
4.2. Collection and Preservation of Plant Material
4.3. DNA Extraction and ISSR Amplification
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gabarral, E.; Belmonte, J.; Canela, M. Aerobiological behavior of Platanus L. pollen in Catalonia (North-East Spain). Aerobiologia 2002, 18, 185–193. [Google Scholar] [CrossRef]
- Treviño, E.J.; Cavazos, C.; Aguirre, O.A. Distribución y estructura de los bosques de galería en dos ríos del centro sur de Nuevo León. Madera Bosques 2001, 7, 13–25. [Google Scholar]
- Hu, L.J.; Uchiyama, K.; Shen, H.L.; Ide, Y. Multiple-scaled spatial genetic structures of Fraxinus mandshurica over a riparian-mountain landscape in Northeast China. Conserv. Genet. 2010, 11, 77–87. [Google Scholar] [CrossRef]
- Premoli, A.C. Isozyme polymorphisms provide evidence of clinal variation with elevation in Nothofagus pumilo. J. Hered. 2003, 94, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Meng, H.; Jiang, M. Landscape genetic structure of a streamside tree species Euptelea pleiospermum (Eupteleaceae): Contrasting roles or River Valley and Mountain Ridge. PLoS One 2013, 8, e66928. [Google Scholar] [CrossRef] [PubMed]
- Piñero, D. La diversidad genética como instrumento para la conservación y el aprovechamiento de la biodiversidad: Estudios en especies mexicanas. In Capital Natural de México; Soberón, J., Halffter, G., Lorente-Bousquets, J., Eds.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2008; Volume 1, pp. 415–435. [Google Scholar]
- Lesica, P.; Allendorf, F.W. When are peripheral populations valuable for conservation? Conserv. Biol. 1995, 9, 753–760. [Google Scholar] [CrossRef]
- Hedrick, P. Population genetics and ecology. In The Princeton Guide to Ecology; Lewin, S.A., Carpenter, S.R., Eds.; Princeton University Press: Princeton, NJ, USA, 2009; p. 737. [Google Scholar]
- Liu, Y.; Wang, Y.; Huang, H. High interpopulation genetic differentiation and unidirectional linear migration patterns in Myricaria laxiflora (Tamaricaceae), an endemic riparian plant in the three Gorges Valley of the Yangtze River. Am. J. Bot. 2006, 93, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Honnay, O.; Jacquemyn, H.; Nackaerts, K.; Breyne, P.; van Looy, K. Patterns of population genetic diversity in riparian and aquatic plant species along rivers. J. Biogeogr. 2010, 37, 1730–1739. [Google Scholar] [CrossRef]
- Viveros-Viveros, H.; Tapia-Olivares, B.L.; Sáenz-Romero, C.; Vargas-Hernández, J.J.; López-Upton, J.; Santacruz-Varela, A.; Ramírez-Valverde, G. Variación isoenzimática de Pinus hartwegii Lindl. en un gradiente altitudinal en Michoacán, México. Agrociencia 2010, 44, 723–733. [Google Scholar]
- Nee, M. Platanaceae. In Flora de Veracruz Fascículo 19; Instituto Nacional de Investigaciones Sobre Recursos Bióticos: Xalapa, México, 1981. [Google Scholar]
- Bornet, B.; Branchard, M. Nonanchored inter simple sequence repeat (ISSR) markers: Reproducible and specific tools for genome fingerprinting. Plant Mol. Biol. Rep. 2001, 19, 209–215. [Google Scholar] [CrossRef]
- Ohsawa, T.; Ide, Y. Global patterns of genetic variation in plant species along vertical and horizontal gradients on mountains. Glob. Ecol. Biogeogr. 2008, 17, 152–163. [Google Scholar] [CrossRef]
- Li, H.; Xie, G.; Blum, M.J.; Zhen, Y.; Lin, M.; Guo, P. Genetic diversity of the endangered Chinese endemic plant Monimopetalum Chinense revealed by amplified fragment length polymorphism (AFLP). Biochem. Syst. Ecol. 2011, 39, 384–391. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Tapia-Olivares, B.L. Pinus oocarpa isoenzymatic variation along an altitudinal gradient in Michoacán, México. Silvae Genet. 2003, 52, 237–240. [Google Scholar]
- Ohsawa, T.; Saito, Y.; Sawada, H.; Ide, Y. Impact of altitude and topography on the genetic diversity of Quercus serrata populations in the Chichibu mountains, central Japan. Flora 2008, 203, 187–196. [Google Scholar] [CrossRef]
- Nishimura, M.; Setoguchi, H. Homogeneous genetic structure and variation in tree architecture of Larix kaempferi along altitudinal gradients on Mt. Fuji. J. Plant Res. 2011, 124, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Unger, G.M.; Konrad, H.; Geburek, T. Does spatial genetic structure increase with altitude? An answer from Picea abies in Tyrol, Austria. Plant Syst. Evol. 2011, 292, 133–141. [Google Scholar] [CrossRef]
- Bricchi, E.; Frenguelli, G.; Mincigrucci, G. Experimental results about Platanus pollen deposition. Aerobiologia 2000, 16, 347–352. [Google Scholar] [CrossRef]
- Jordano, P. Pollen, seeds and genes: The movement ecology of plants. Heredity 2010, 105, 329–330. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Peñuelas, J. Genetic effects of chronic habitat fragmentation in a wind-pollinated tree. Proc. Natl. Acad. Sci. USA 2006, 103, 8096–8100. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Tsuda, Y.; Koike, Y.; Lian, C.; Ide, Y. Effects of landscape and demographic history on genetic variation in Picea glehnii at the regional scale. Ecol. Res. 2009, 24, 1267–1277. [Google Scholar] [CrossRef]
- Hahn, T.; Kettle, C.J.; Ghazoul, J.; Frei, E.R.; Matter, P.; Pluess, A.R. Patterns of genetic variation across altitude in three plant species of semi-dry grasslands. PLoS One 2012, 7, e41608. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, T.; Tsuda, Y.; Saito, Y.; Sawada, H.; Ide, Y. Altitudinal genetic diversity and differentiation of Quercus crispula in the Chichibu mountains, central Japan. Int. J. Plant Sci. 2007, 168, 333–340. [Google Scholar] [CrossRef]
- Chase, M.W.; Hills, H.H. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Xiao, L.Q.; Gong, X. Genetic differentiation and relationships of populations in the Cycas balansae complex (Cycadaceae) and its conservation implications. Ann. Bot. 2006, 97, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Milligan, B.G. Analysis of population genetic structure with RAPD markers. Mol. Ecol. 1994, 3, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.P. Tools for Population Genetic Analyses (TFPGA) Version 1.3. A Windows Program for the Analysis of Allozyme and Molecular Population Genetic Data; Utah State University: Logan, UT, USA, 1997. [Google Scholar]
- Data Analysis Software System, (STATISTICA) Version 8.0. Available online: http://www.statsoft.com (accessed on 20 November 2014).
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galván-Hernández, D.M.; Lozada-García, J.A.; Flores-Estévez, N.; Galindo-González, J.; Vázquez-Torres, S.M. Variation and Genetic Structure in Platanus mexicana (Platanaceae) along Riparian Altitudinal Gradient. Int. J. Mol. Sci. 2015, 16, 2066-2077. https://doi.org/10.3390/ijms16012066
Galván-Hernández DM, Lozada-García JA, Flores-Estévez N, Galindo-González J, Vázquez-Torres SM. Variation and Genetic Structure in Platanus mexicana (Platanaceae) along Riparian Altitudinal Gradient. International Journal of Molecular Sciences. 2015; 16(1):2066-2077. https://doi.org/10.3390/ijms16012066
Chicago/Turabian StyleGalván-Hernández, Dulce M., J. Armando Lozada-García, Norma Flores-Estévez, Jorge Galindo-González, and S. Mario Vázquez-Torres. 2015. "Variation and Genetic Structure in Platanus mexicana (Platanaceae) along Riparian Altitudinal Gradient" International Journal of Molecular Sciences 16, no. 1: 2066-2077. https://doi.org/10.3390/ijms16012066
APA StyleGalván-Hernández, D. M., Lozada-García, J. A., Flores-Estévez, N., Galindo-González, J., & Vázquez-Torres, S. M. (2015). Variation and Genetic Structure in Platanus mexicana (Platanaceae) along Riparian Altitudinal Gradient. International Journal of Molecular Sciences, 16(1), 2066-2077. https://doi.org/10.3390/ijms16012066