Selective Isolation of Trypsin Inhibitor and Lectin from Soybean Whey by Chitosan/Tripolyphosphate/Genipin Co-Crosslinked Beads


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
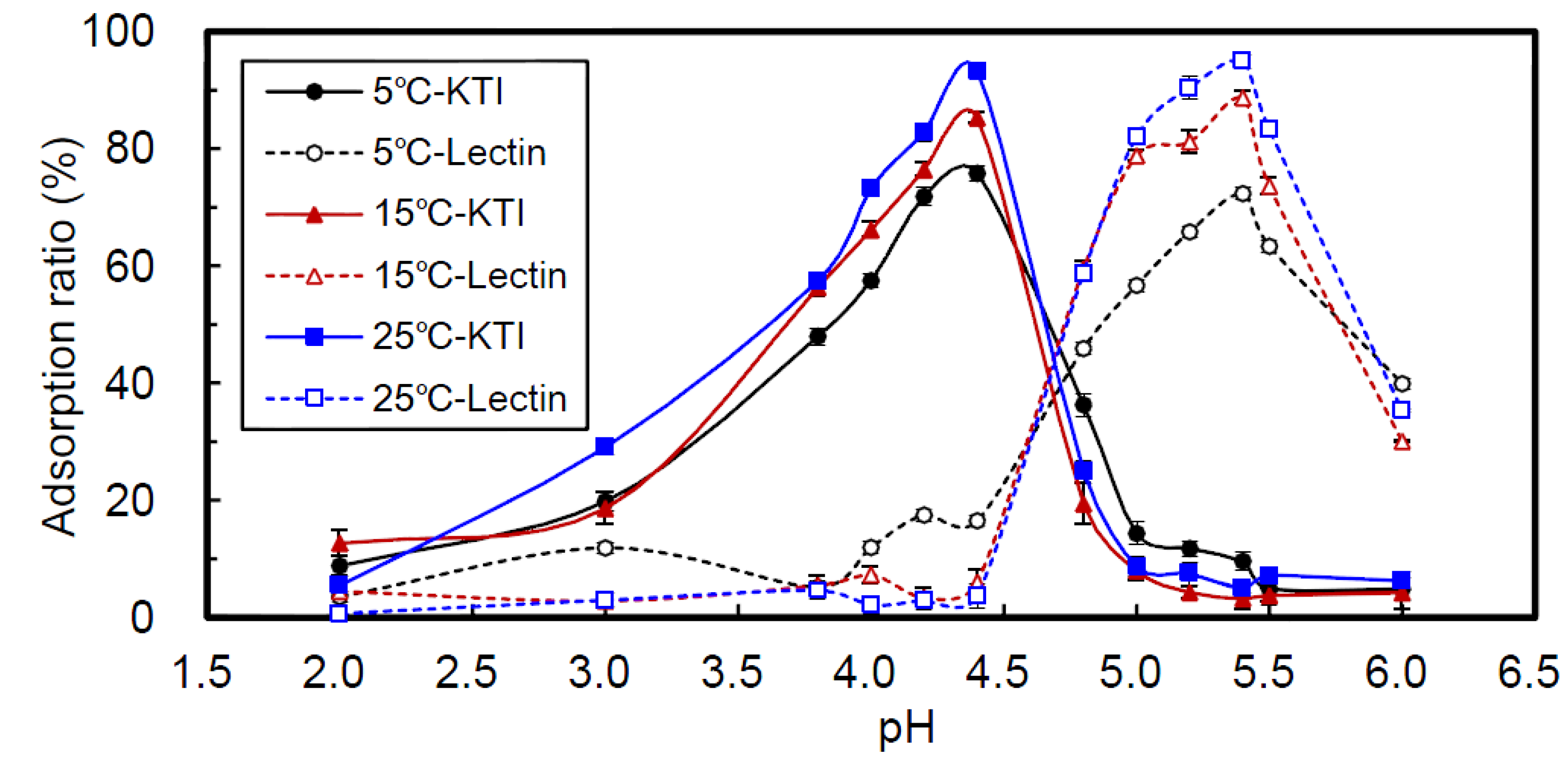
2.1. Effect of pH
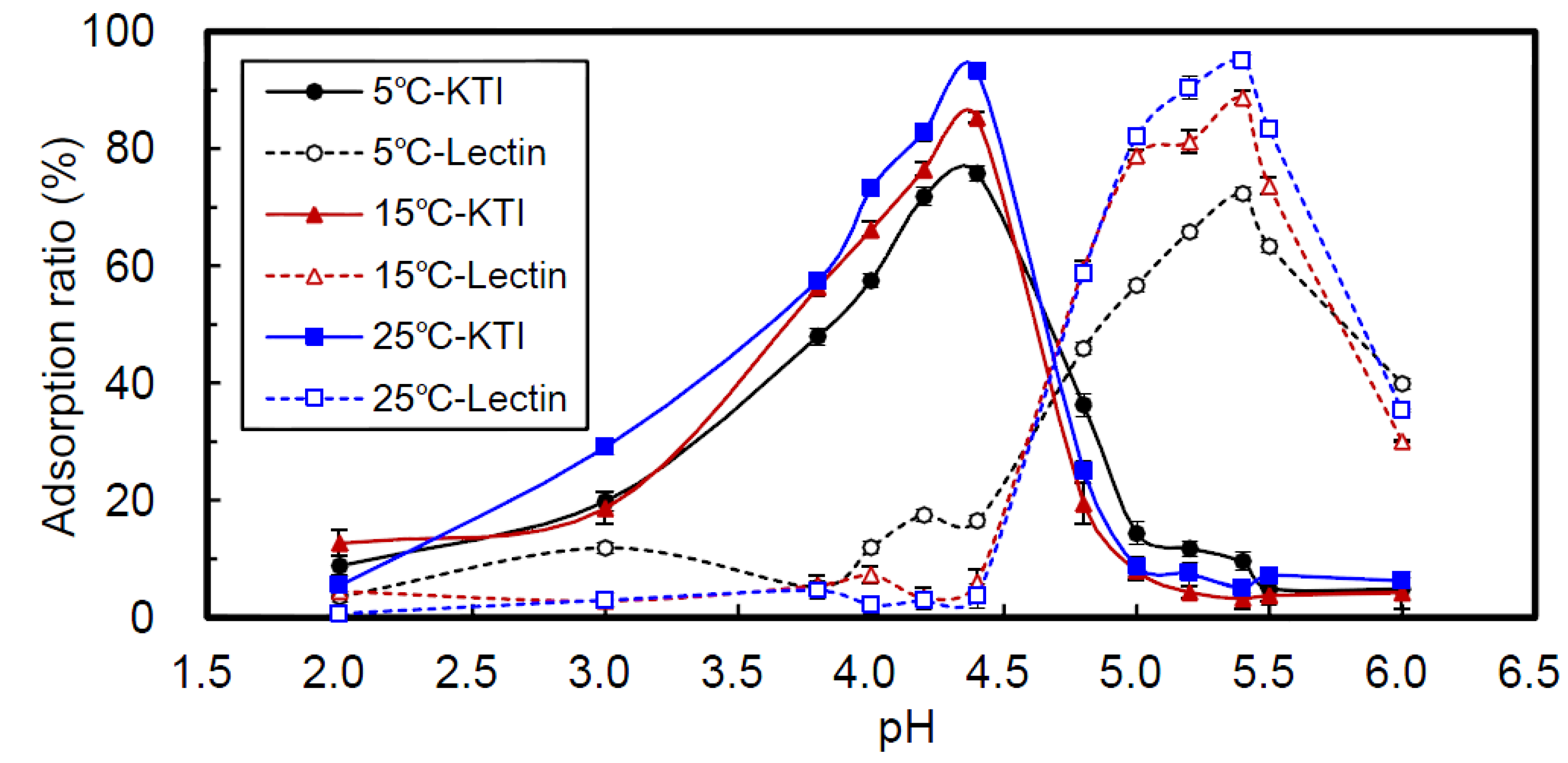
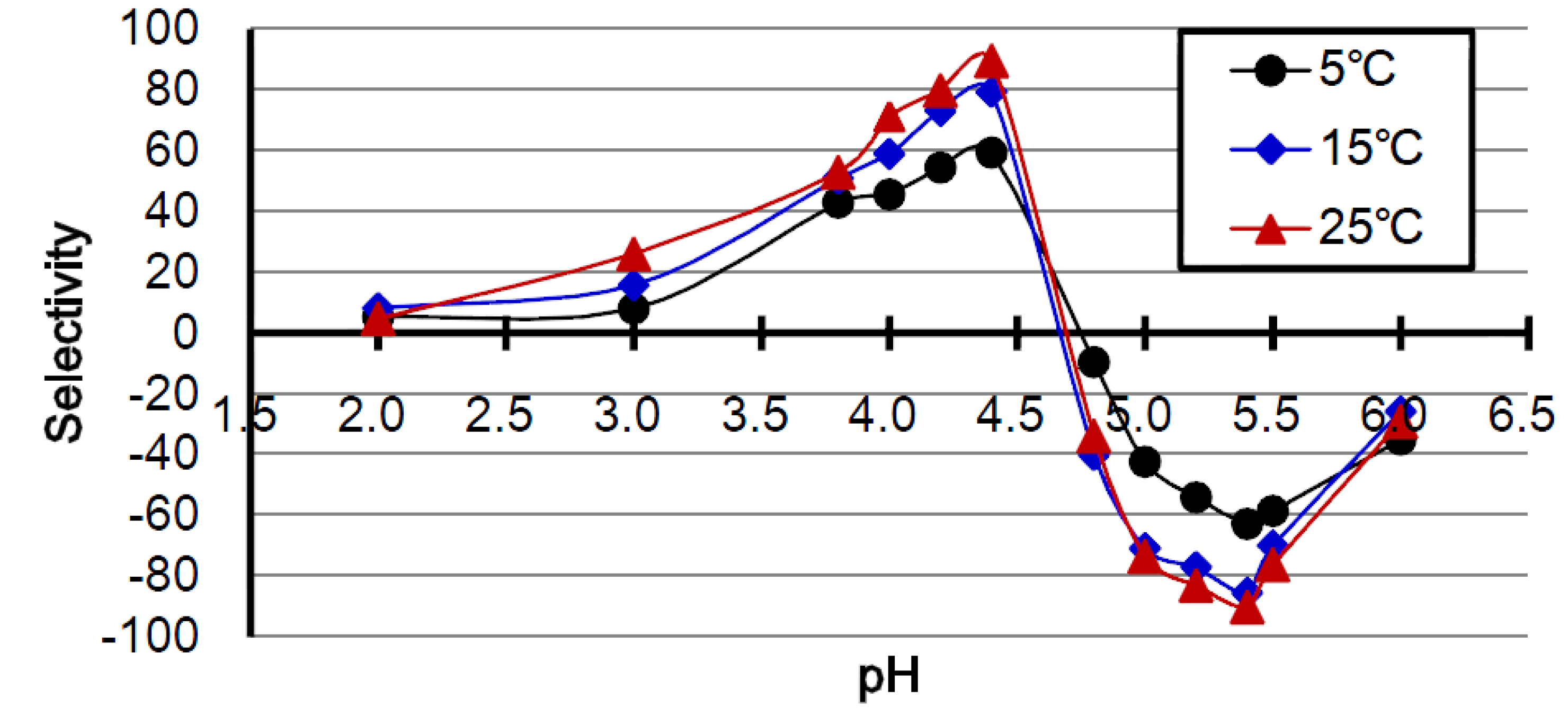
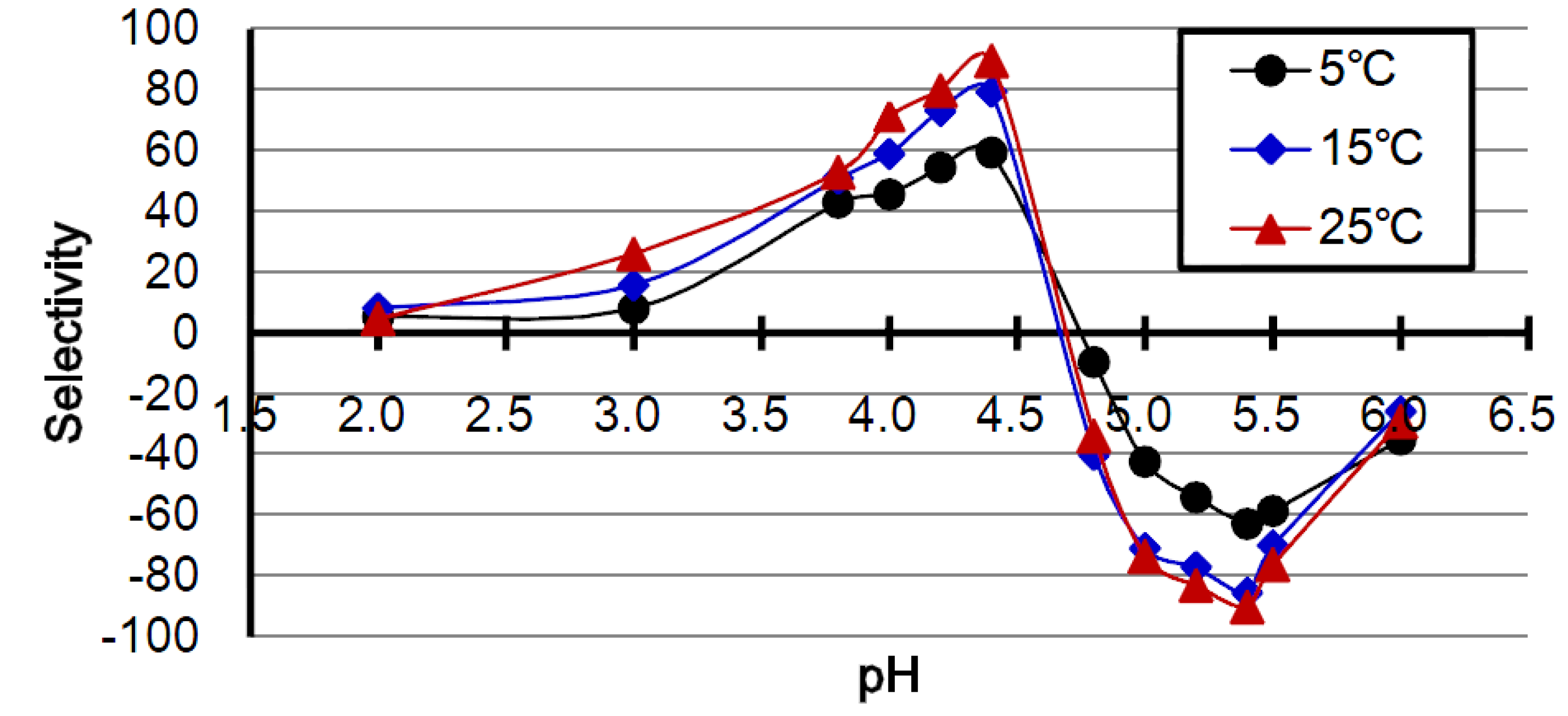
2.2. Effect of Temperature

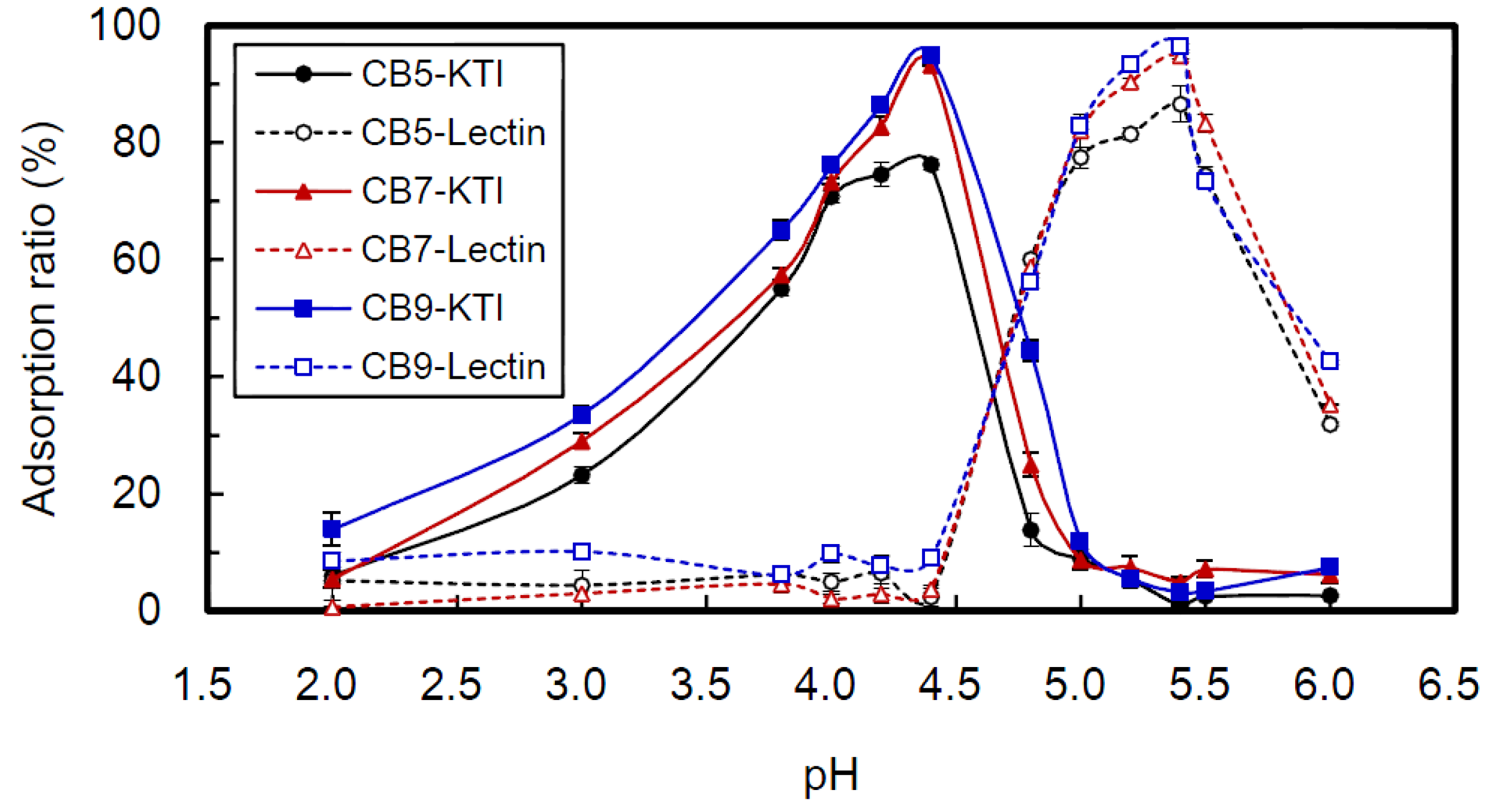
2.3. Effect of Kinds of CB

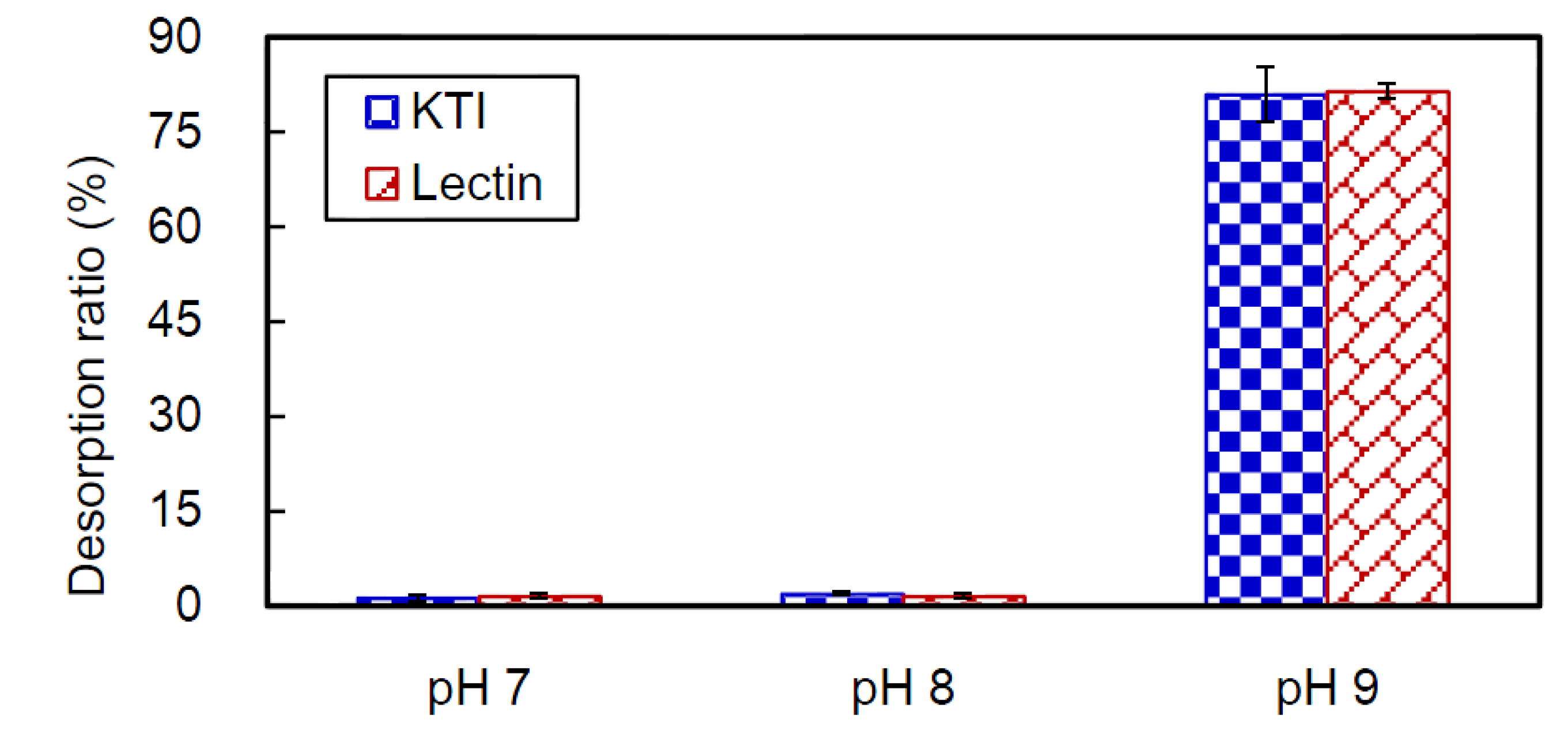
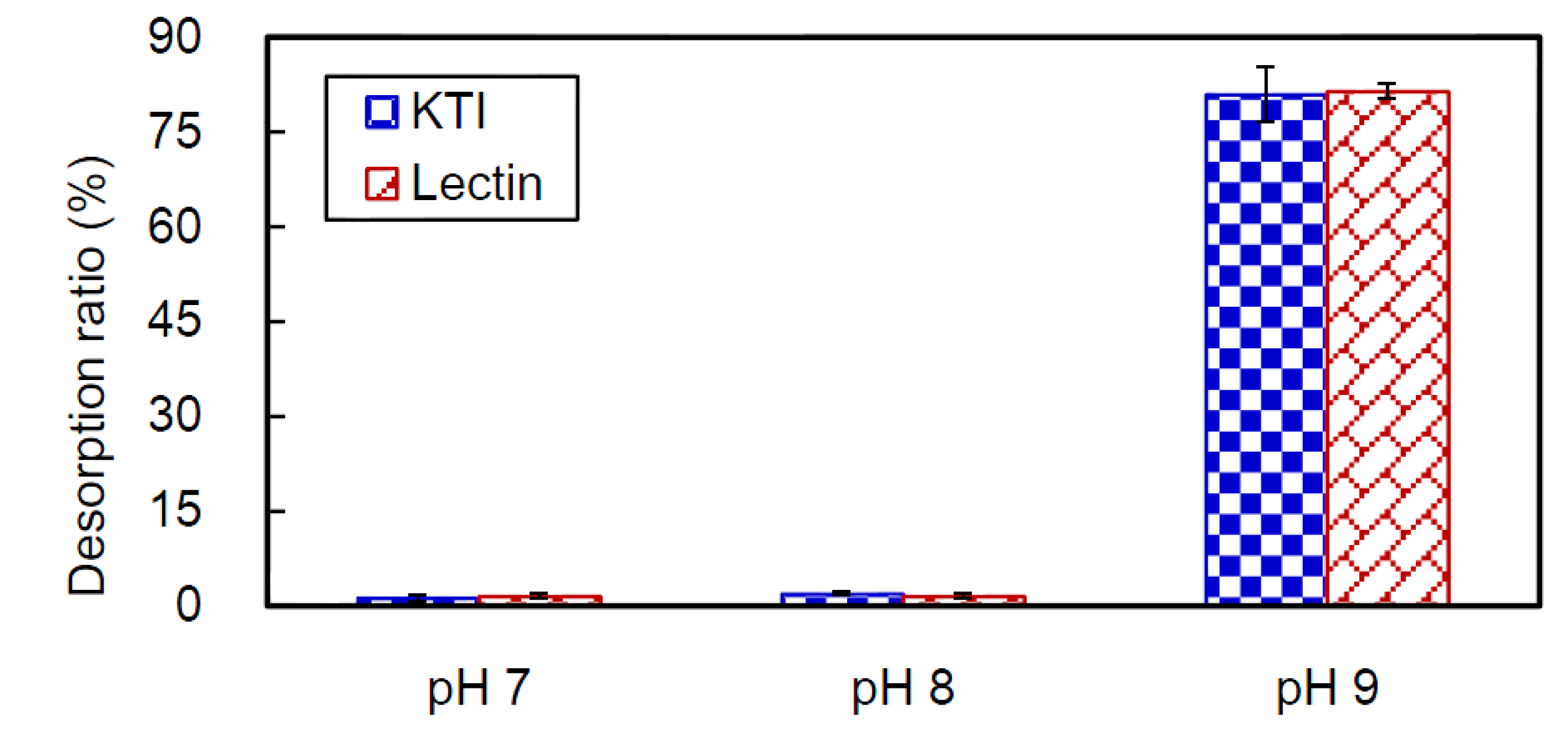
2.4. Desorption of CB
3. Experimental Section
3.1. Materials
3.2. Preparation of TPP/Genipin Co-Crosslinked Chitosan Beads
3.3. Determination of Crosslinking Degree of Chitosan Bead
3.4. Preparation of Soybean Whey
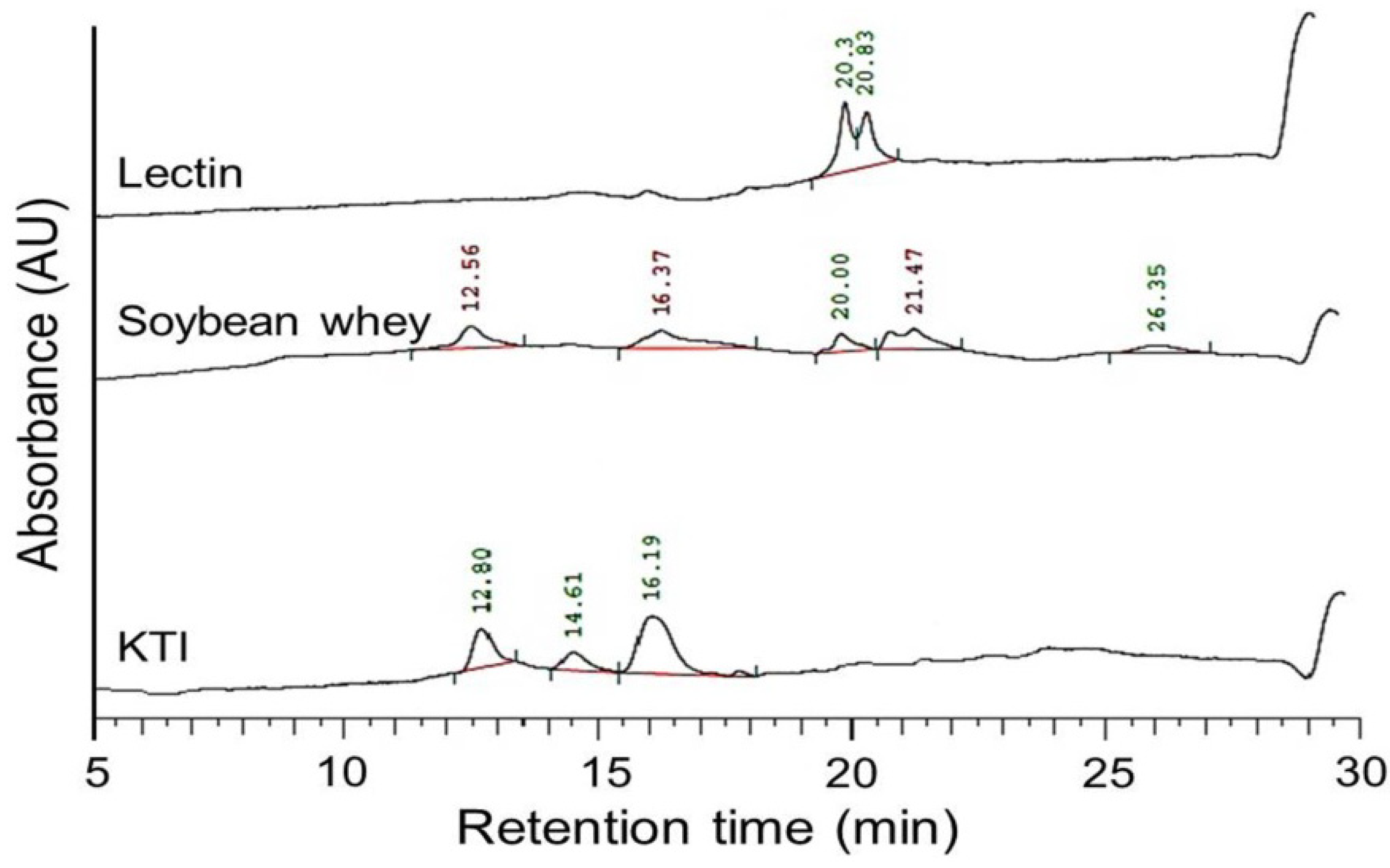
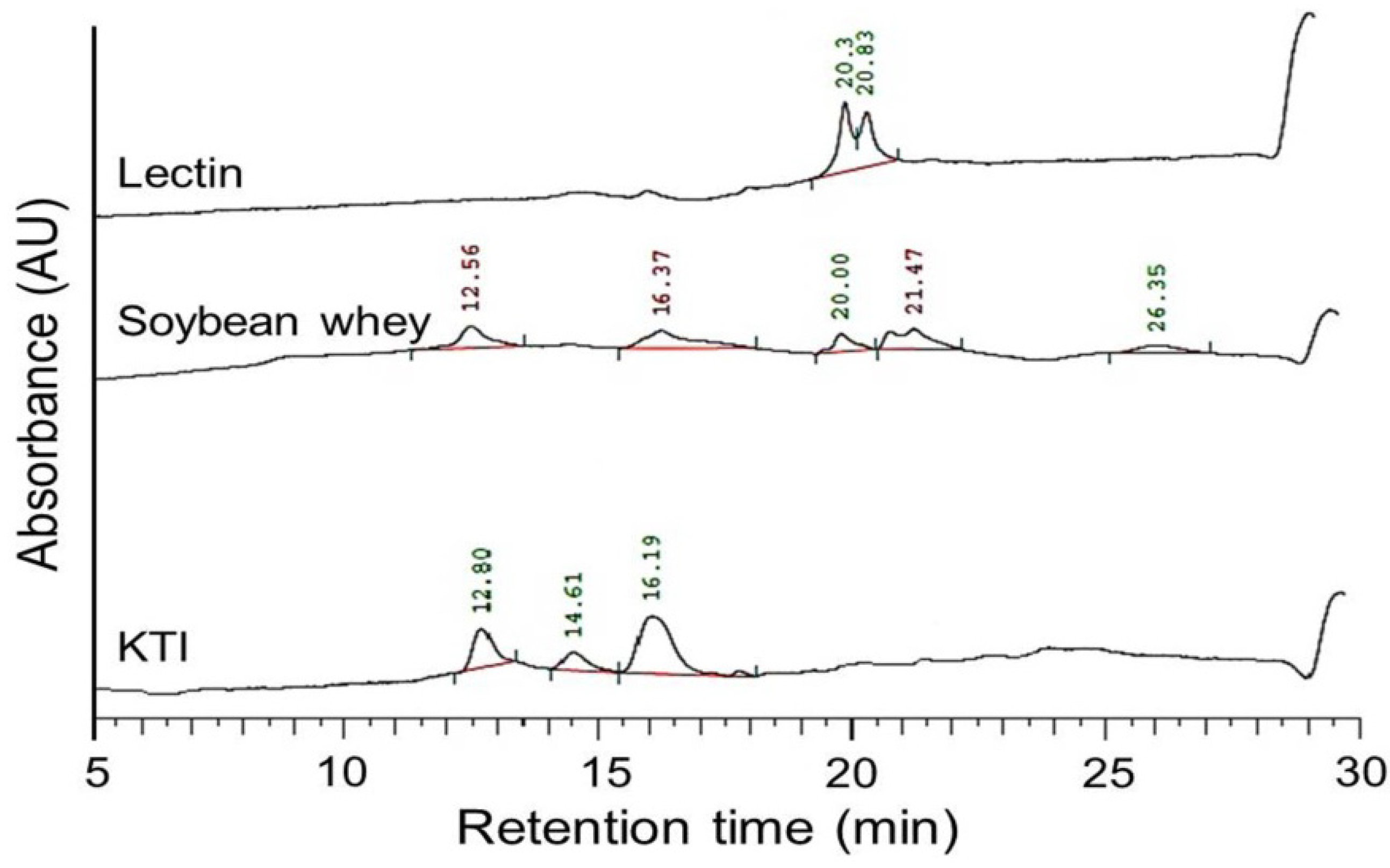
3.5. Determination of Trypsin Inhibitor and Lectin
3.6. Adsorption and Desorption of KTI and Lectin
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Préstamo, G.; Arroyo, G. Biological response of rats fed with tofu treated with high hydrostatic pressure. J. Agric. Food Chem. 2000, 48, 4641–4646. [Google Scholar] [CrossRef]
- Smith, A.K.; Nash, A.M.; Eldrige, A.C.; Wolf, W.J. Recovery of soybean whey proteins with edible gums and detergents. J. Agric. Food Chem. 1962, 10, 302–304. [Google Scholar] [CrossRef]
- Sorgentini, D.A.; Wanger, J.R. Comparative study of structural characteristics and thermal behavior of whey and isolate soybean proteins. J. Food Biochem. 1999, 23, 489–507. [Google Scholar] [CrossRef]
- Koide, T.; Ikenaka, T. Studies on soybean trypsin inhibitors. 3. Amino acid sequence of the carboxyl terminal region and the complete amino acid sequence of soybean trypsin inhibitor (Kunitz). Eur. J. Biochem. 1973, 32, 417–431. [Google Scholar] [CrossRef]
- Koshiyama, Y.; Kikuchi, M.; Fucusshima, D. 2S Globulins of soybean seeds. 2. Physicochemical and biological properties of protease inhibitors in 2S globulins. J. Agric. Food Chem. 1981, 29, 340–343. [Google Scholar] [CrossRef]
- Clawson, G.A. Protease inhibitors and carcinogenesis: A review. Cancer Investig. 1996, 14, 597–608. [Google Scholar] [CrossRef]
- Lotan, R.; Siegelman, H.W.; Lis, H.; Sharon, N. Subunit structure of soybean agglutinin. J. Biol. Chem. 1974, 249, 1219–1224. [Google Scholar]
- Khan, J.M.; Qadeer, A.; Ahmad, E.; Ashraf, R.; Bhushan, B.; Chaturvedi, S.K.; Rabbani, G.; Khan, R.H. Monomeric banana lectin at acidic pH overrules conformational stability of its native dimeric form. PLoS One 2013, 8, e62428. [Google Scholar]
- Kassell, B. Trypsin and chymotrypsin inhibitors from soybeans. Methods Enzymol. 1970, 19, 853–862. [Google Scholar] [CrossRef]
- Duranti, M.; Barbiroli, A.; Scarafoni, A.; Tedeschi, G.; Morazzonic, P. One-step purification of Kunitz soybean trypsin inhibitor. Protein Expr. Purif. 2003, 30, 167–170. [Google Scholar]
- Gupta, M.N.; Jain, S.; Roy, I. Immobilized metal affinity chromatography without chelating ligands: Purification of soybean trypsin inhibitor on zinc alginate beads. Biotechnol. Prog. 2002, 18, 78–81. [Google Scholar]
- Singh, K.; Kaur, M.; Rup, P.J.; Singh, J. Exploration for anti-insect properties of lectin from seeds of soybean (Glycine max) using Bactrocera cucurbitae as a model. Phytoparasitica 2006, 34, 463–473. [Google Scholar] [CrossRef]
- Takahashi, K.G.; Kuroda, T.; Muroga, K. Purification and antibacterial characterization of a novel isoform of the Manila clam lectin (MCL-4) from the plasma of the Manila clam Ruditapes philippinarum. Comp. Biochem. Phys. B 2008, 150, 45–52. [Google Scholar] [CrossRef]
- Watanabe, Y.; Shiina, N.; Shinozaki, F.; Yokoyama, H.; Kominami, J.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Sugahara, K.; Kamiya, H.; Matsubara, H.; et al. Isolation and characterization of l-rhamnose-binding lectin, which binds to microsporidian Glugea plecoglossi, from ayu (Plecoglossus altivelis) eggs. Dev. Comp. Immunol. 2008, 32, 487–499. [Google Scholar]
- Nascimento, C.O.; Costa, R.M.P.B.; Araujo, R.M.S.; Chaves, M.E.C.; Coelho, L.C.B.B.; Paiva, P.M.G.; Teixeira, J.A.; Correia, M.T.S.; Carneiro-da-Cunha, M.G. Optimized extraction of a lectin from Crataeva tapia bark using AOT in isooctane reversed micelles. Process Biochem. 2008, 43, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, C.O.; Soares, P.A.G.; Porto, T.S.; Costa, R.M.P.B.; Lima, C.A.; Lima Filho, J.L.; Coelho, L.C.B.B.; Santos Correia, M.T.; Carneiro Cunha, M.G.; Porto, A.L.F. Aqueous two-phase systems: New strategies for separation and purification of lectin from crude extract of Cratylia mollis seeds. Sep. Purif. Technol. 2013, 116, 154–161. [Google Scholar]
- Teotia, S.; Mondal, K.; Gupta, M.N. Integration of affinity precipitation with partitioning methods for bioseparation of chitin binding lectins. Food Bioprod. Process. 2006, 84, 37–43. [Google Scholar] [CrossRef]
- Fields, C.; Mallee, P.; Muzard, J.; Lee, G.U. Isolation of Bowman-Birk-Inhibitor from soybean extracts using novel peptide probes and high gradient magnetic separation. Food Chem. 2012, 134, 1831–1838. [Google Scholar]
- Muzard, J.; Fields, C.; O’Mahony, J.J.; Lee, G.U. Probing the soybean Bowman-Birk Inhibitor using recombinant antibody fragments. J. Agric. Food Chem. 2012, 60, 6164–6172. [Google Scholar] [CrossRef]
- Casal, E.; Montilla, A.; Moreno, F.J.; Olano, A.; Corzo, N. Use of chitosan for selective removal of β-lactoglobulin from whey. J. Dairy Sci. 2006, 89, 1384–1389. [Google Scholar] [CrossRef]
- Feng, Z.; Shao, Z.; Yao, J.; Huang, Y.; Chen, X. Protein adsorption and separation with chitosan-based amphoteric membranes. Polymer 2009, 50, 1257–1263. [Google Scholar] [CrossRef]
- Montilla, A.; Casal, E.; Moreno, F.J.; Belloque, J.; Olano, A.; Corzo, N. Isolation of bovine beta-lactoglobulin from complexes with chitosan. Int. Dairy J. 2007, 17, 459–464. [Google Scholar]
- Sepehran, M.; Nasernejad, B.; Edrissi, M. Selective removal of copper(II) and nickel(II) from aqueous solution using the chemically treated chitosan: Factorial design evaluation. Metall. Mater. Trans. B 2008, 39B, 905–910. [Google Scholar]
- Vold, I.M.N.; Vårum, K.M.; Guibal, E.; Smidsrød, O. Binding of ions to chitosan-selectivity studies. Carbohydr. Polym. 2003, 54, 471–477. [Google Scholar] [CrossRef]
- Yang, C.Y.; Hsu, C.H.; Tsai, M.L. Effect of crosslinked condition on characteristics of chitosan/tripolyphosphate/genipin beads and their application in the selective adsorption of phytic acid from soybean whey. Carbohydr. Polym. 2011, 86, 659–665. [Google Scholar]
- Mi, F.L.; Sung, H.W.; Shyu, S.S.; Su, C.C.; Peng, C.K. Synthesis and characterization of biodegradable TPP/genipin co-crosslinked chitosan gel beads. Polymer 2003, 24, 6521–6530. [Google Scholar]
- Tsai, M.L.; Chang, H.W.; Yu, H.C.; Lin, Y.S.; Tsai, Y.D. Effect of chitosan characteristics and solution conditions on gelation temperatures of chitosan/2-glycerophosphate/nanosilver hydrogels. Carbohydr. Polym. 2011, 84, 1337–1343. [Google Scholar]
- Mi, F.L.; Tan, Y.C.; Liang, H.C.; Huang, R.N.; Sung, H.W. In vitro evaluation of a chitosan membrane cross-linked with genipin. J. Biomater. Sci. Polym. Ed. 2001, 12, 835–850. [Google Scholar] [CrossRef]
- Castro-Rubio, F.; Marina, M.L.; García, M.C. Perfusion reversed-phase high-performance liquid chromatography/mass spectrometry analysis of intact soybean proteins for the characterization of soybean cultivars. J. Chromatogr. A 2007, 1170, 34–43. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, Y.-L.; Liu, T.C.; Tsai, M.-L. Selective Isolation of Trypsin Inhibitor and Lectin from Soybean Whey by Chitosan/Tripolyphosphate/Genipin Co-Crosslinked Beads. Int. J. Mol. Sci. 2014, 15, 9979-9990. https://doi.org/10.3390/ijms15069979
Chang Y-L, Liu TC, Tsai M-L. Selective Isolation of Trypsin Inhibitor and Lectin from Soybean Whey by Chitosan/Tripolyphosphate/Genipin Co-Crosslinked Beads. International Journal of Molecular Sciences. 2014; 15(6):9979-9990. https://doi.org/10.3390/ijms15069979
Chicago/Turabian StyleChang, Yu-Lung, Tristan C. Liu, and Min-Lang Tsai. 2014. "Selective Isolation of Trypsin Inhibitor and Lectin from Soybean Whey by Chitosan/Tripolyphosphate/Genipin Co-Crosslinked Beads" International Journal of Molecular Sciences 15, no. 6: 9979-9990. https://doi.org/10.3390/ijms15069979