Genetic Variation of the Endangered Gentiana lutea L. var. aurantiaca (Gentianaceae) in Populations from the Northwest Iberian Peninsula


Abstract
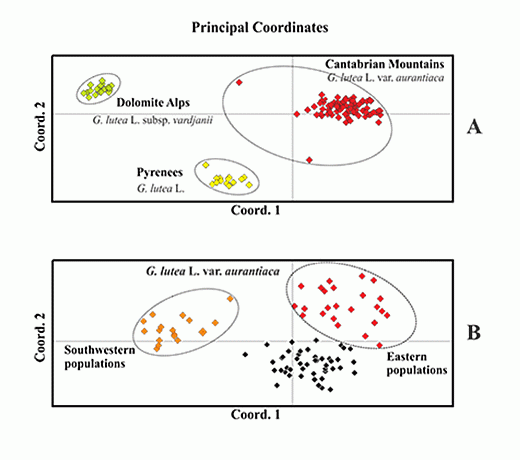
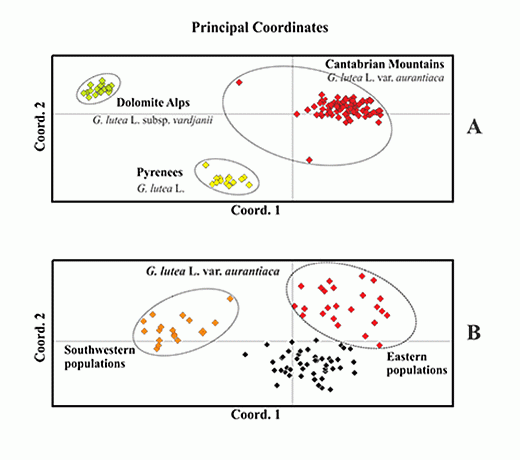
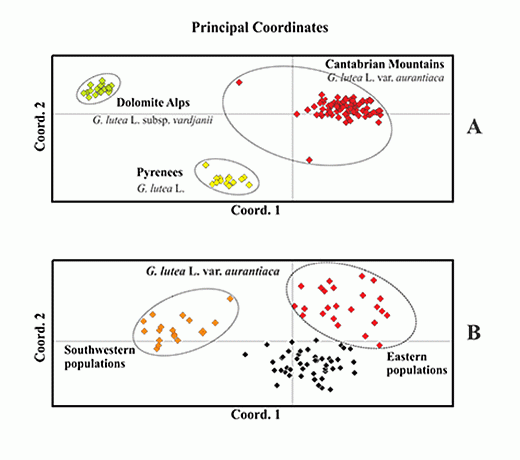
:
1. Introduction
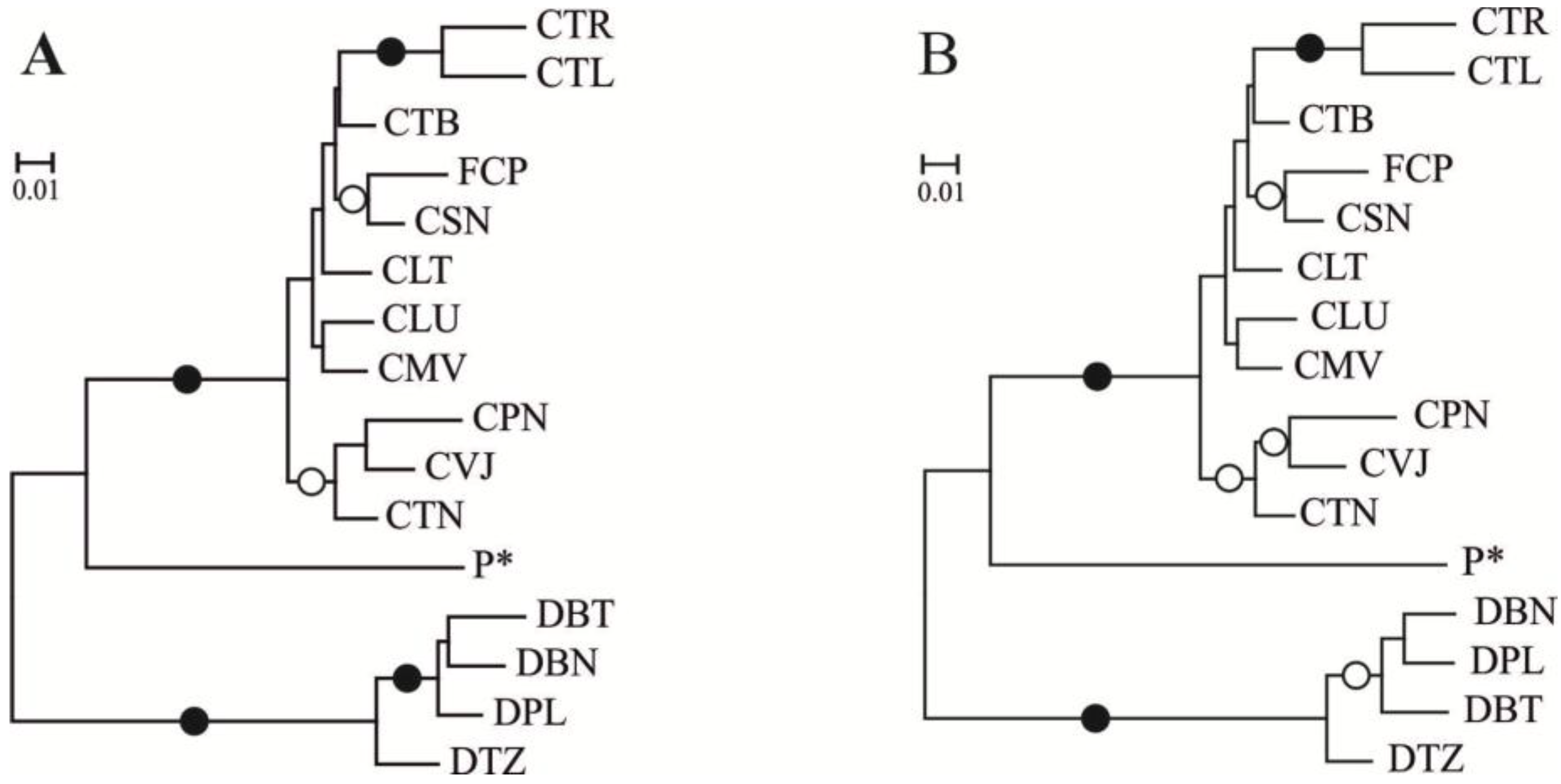
2. Results

{kind=link}
{kind=link}
{kind=link}
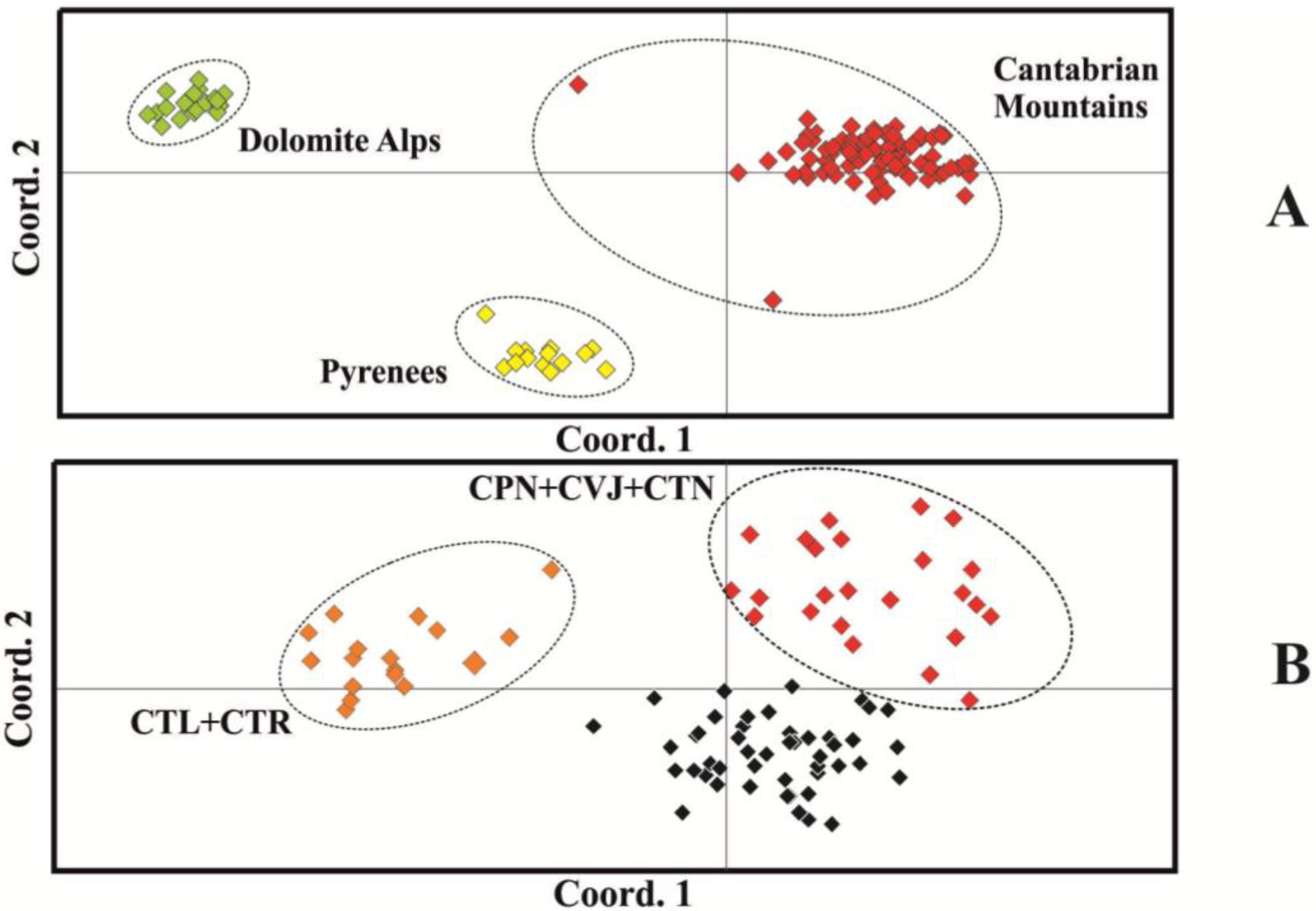{kind=link}
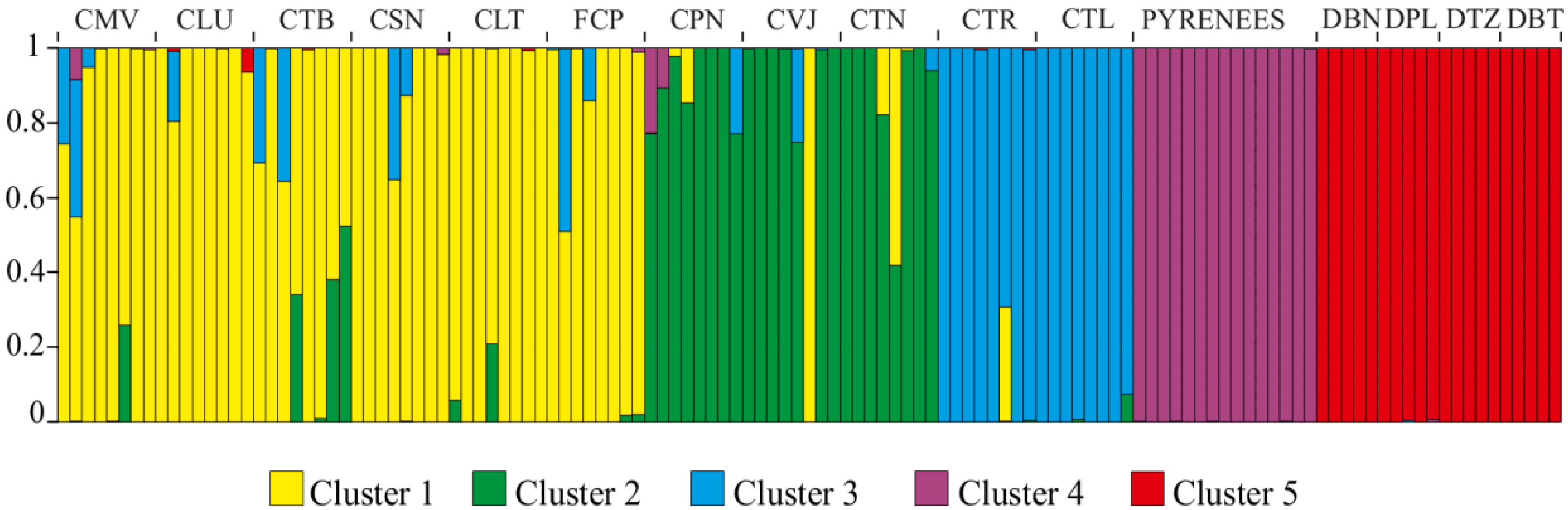{kind=link}
{kind=link}
| Geographic Region (Taxon) | Population Names | N | n | PPL | Na | Ne | I | He (S.E.) |
|---|---|---|---|---|---|---|---|---|
| Cantabrian Mountains (G. lutea var. aurantiaca) | CMV | 8 | 33 | 31.13 | 1.3113 | 1.1743 | 0.1579 | 0.1042 (0.0167) |
| CLU | 8 | 26 | 24.53 | 1.2453 | 1.1297 | 0.1192 | 0.0780 (0.0149) | |
| CTB | 8 | 32 | 30.19 | 1.3019 | 1.1800 | 0.1568 | 0.1046 (0.0172) | |
| CSN | 8 | 27 | 25.47 | 1.2547 | 1.1361 | 0.1218 | 0.0801 (0.0154) | |
| CLT | 8 | 29 | 27.36 | 1.2736 | 1.1530 | 0.1398 | 0.0922 (0.0159) | |
| CPN | 8 | 35 | 33.02 | 1.3302 | 1.2033 | 0.1716 | 0.1152 (0.0181) | |
| CVJ | 8 | 41 | 38.68 | 1.3868 | 1.2266 | 0.1993 | 0.1329 (0.0184) | |
| CTN | 8 | 32 | 30.19 | 1.3019 | 1.1878 | 0.1608 | 0.1081 (0.0176) | |
| CTR | 8 | 21 | 19.81 | 1.1981 | 1.1264 | 0.1066 | 0.0719 (0.0153) | |
| CTL | 8 | 20 | 18.87 | 1.1887 | 1.1290 | 0.1082 | 0.0736 (0.0155) | |
| FCP | 8 | 28 | 26.42 | 1.2642 | 1.1583 | 0.1363 | 0.0911 (0.0165) | |
| Mean values | 29.45 | 27.79 | 1.2779 | 1.1640 | 0.1435 | 0.0956 (0.0058) | ||
| Group | 88 | 70 | 66.04 | 1.6604 | 1.2346 | 0.2361 | 0.1480 (0.0028) | |
| Pyrenees (G. lutea), Dolomite Alps (G. lutea L. sub sp. vardjanii) | Group | 15 | 30 | 28.30 | 1.283 | 1.1883 | 0.1581 | 0.1074 (0.0179) |
| DBN | 5 | 18 | 16.98 | 1.1698 | 1.1138 | 0.0964 | 0.0654 (0.0147) | |
| DPL | 5 | 24 | 22.64 | 1.2264 | 1.1502 | 0.1263 | 0.0856 (0.0164) | |
| DTZ | 5 | 20 | 18.87 | 1.1887 | 1.1304 | 0.1078 | 0.0735 (0.0157) | |
| DBT | 5 | 15 | 14.15 | 1.1415 | 1.0978 | 0.08 | 0.0546 (0.0139) | |
| Mean values | 19.25 | 18.16 | 1.1816 | 1.1231 | 0.1026 | 0.0698 (0.0065) | ||
| All | Group | 20 | 31 | 29.20 | 1.2925 | 1.1735 | 0.1499 | 0.1002 (0.0017) |
| Global mean | 26.9 | 25.40 | 1.2541 | 1.1553 | 0.1342 | 0.0900 (0.0052) | ||
| Total | 123 | 95 | 89.60 | 1.8962 | 1.3446 | 0.3415 | 0.2168 (0.0166) |
| Analysis | Source of Variation | d.f. | Sum of Squares | Estimated Variance | % Total Variance | Φ | Significance (p) |
|---|---|---|---|---|---|---|---|
| G. lutea | Among taxons | 2 | 552.721 | 9.786 | 57 | 0.568 | <0.001 |
| Within taxons | 120 | 891.962 | 7.433 | 43 | |||
| G. lutea var. aurantiaca | Among clusters * | 2 | 123.608 | 1.765 | 20 | 0.198 | <0.001 |
| Among populations | 8 | 124.688 | 1.208 | 14 | 0.169 | <0.001 | |
| Within populations | 77 | 456.250 | 5.925 | 67 | 0.334 | <0.001 |
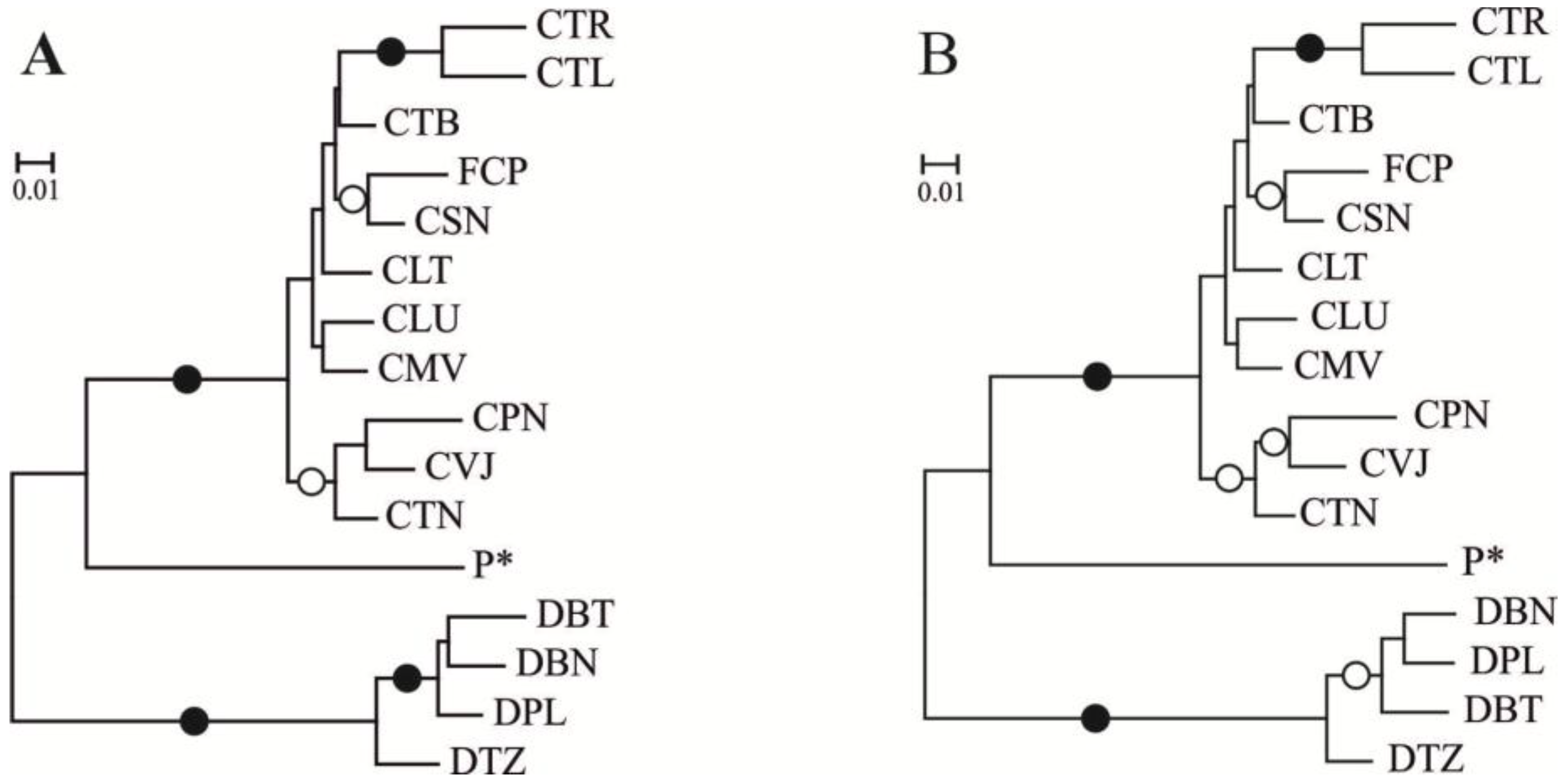

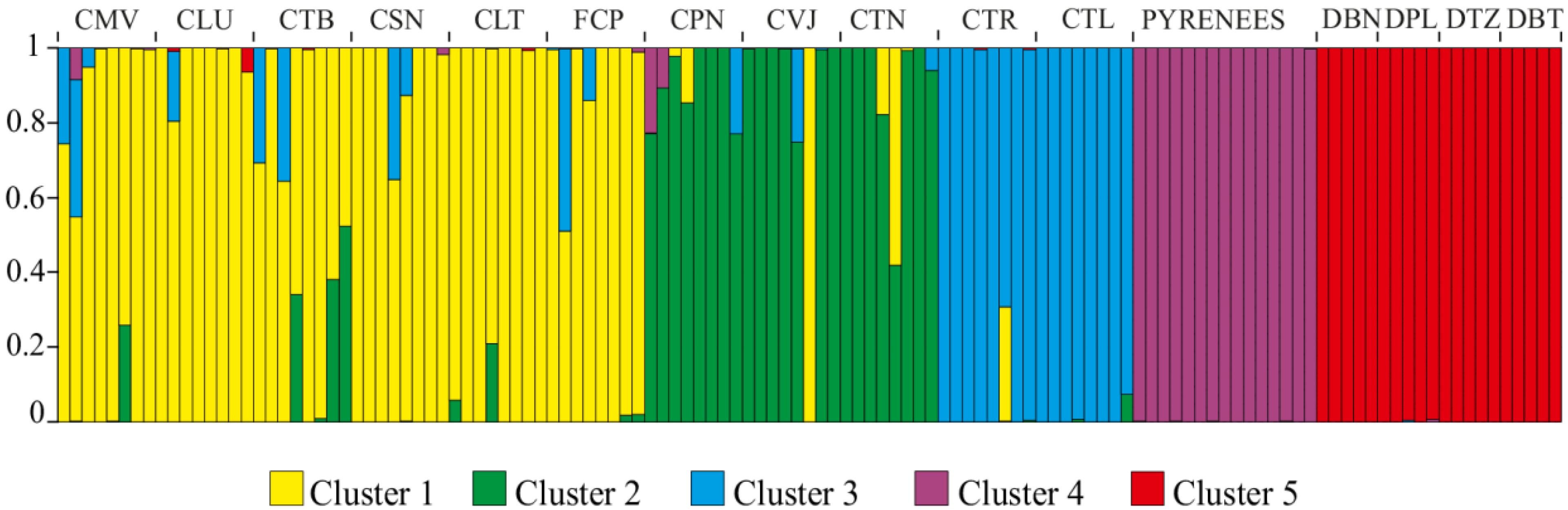
3. Discussion
4. Experimental Section
4.1. Study Species and Population Sampling
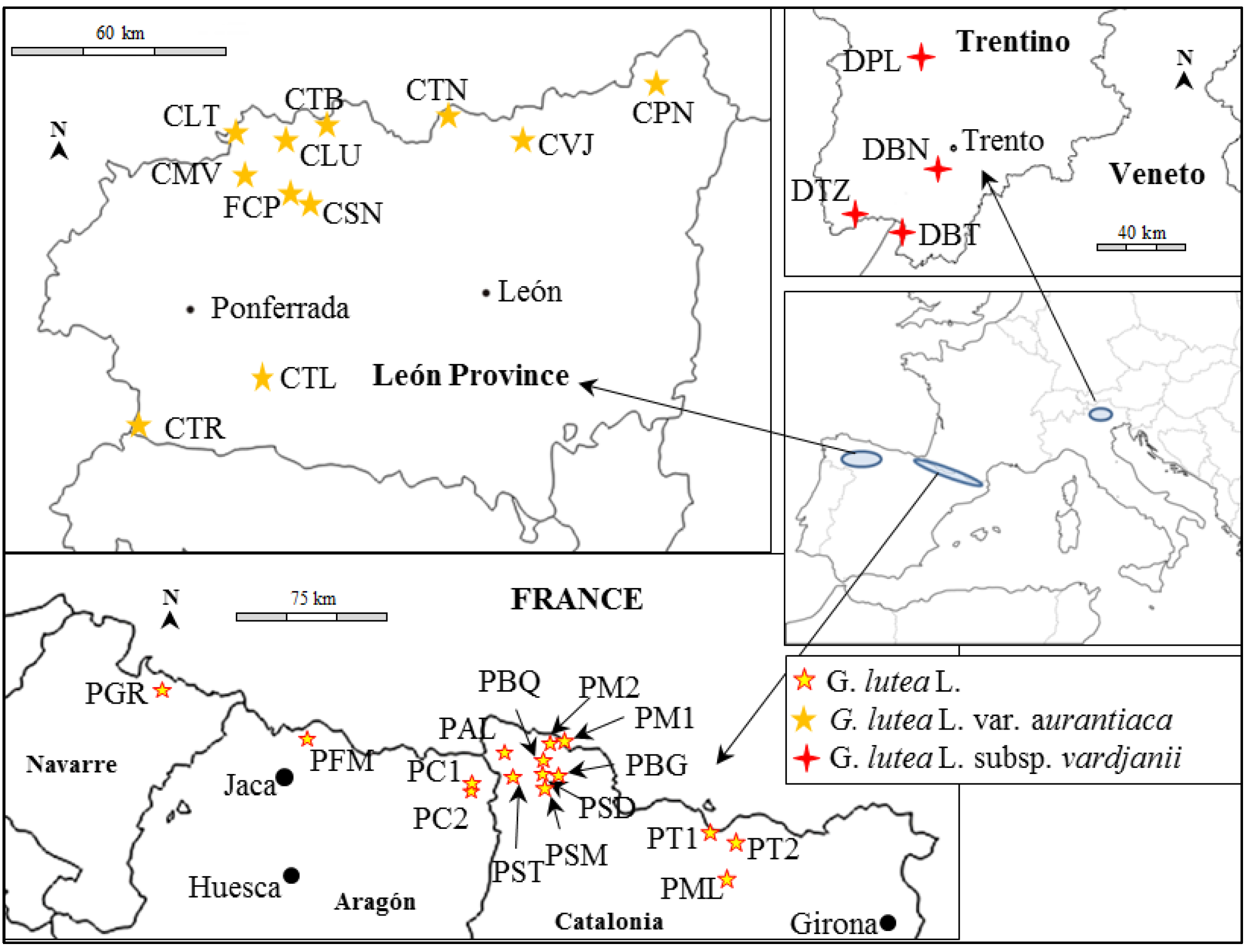
| Location | Pop name (Figure 1) | Latitude (North) | Longitude | Altitude (m) | Sample Size |
|---|---|---|---|---|---|
| Matalavilla | CMV | 42°52'13'' | 06°23'06''W | 1794 | 8 |
| Lumajo | CLU | 42°59'14'' | 06°15'33''W | 1396 | 8 |
| Torre de Babia | CTB | 43°00'47'' | 06°05'59''W | 1801 | 8 |
| Senra | CSN | 42°50'07'' | 06°11'25''W | 1724 | 8 |
| Leitariegos | CLT | 42°59'18'' | 06°25'26''W | 1880 | 8 |
| Pontón | CPN | 43°06'02'' | 05°00'29''W | 1278 | 8 |
| Valdeteja | CVJ | 42°56'28'' | 05°27'34''W | 1419 | 8 |
| Tonin | CTN | 43°00'15'' | 05°41'00''W | 1294 | 8 |
| Trevinca | CTR | 42°09'30'' | 06°44'29''W | 1727 | 8 |
| Teleno | CTL | 42°21'20'' | 06°25'22''W | 1867 | 8 |
| Cultivated field | FCP | 42°52'09'' | 06°12'49''W | 1439 | 8 |
| Collada de Tosas 1 | PT1 | 42°20'53'' | 01°59'08''E | 1738 | 1 |
| Collada de Tosas 2 | PT2 | 42°20'29'' | 02°01'49''E | 1654 | 1 |
| La Molina | PML | 42°17'13'' | 02°02'07''E | 1578 | 1 |
| La Bonaigua | PBG | 42°40'10'' | 00°58'11''E | 1813 | 1 |
| Salardu | PSD | 42°40'20'' | 00°54'58''E | 1516 | 1 |
| Montgarri | PMG | 42°45'27'' | 01°01'58''E | 1602 | 1 |
| Montgarri 2 | PMA | 42°45'33'' | 01°01'14''E | 1588 | 1 |
| Baqueira | PBQ | 42°43'31'' | 00°55'59''E | 1764 | 1 |
| Sant Maurici | PSM | 42°39'28'' | 00°55'24''E | 1516 | 1 |
| Artiga de Lin | PAL | 42°40'50'' | 00°42'24''E | 1458 | 1 |
| Senet | PST | 42°36'23'' | 00°46'01''E | 1487 | 1 |
| Cerler 1 | PC1 | 42°33'55'' | 00°34'01''E | 1902 | 1 |
| Cerler 2 | PC2 | 42°33'43'' | 00°34'03''E | 1892 | 1 |
| Garaioa | PGR | 42°53'23'' | 01°13'51''W | 1170 | 1 |
| Formigal | PFM | 42°47'45'' | 00°24'11''W | 1701 | 1 |
| Bondone | DBN | 46°00'34'' | 11°02'50''E | 1557 | 5 |
| Peller | DPL | 46°19'04'' | 10°57'34''E | 1904 | 5 |
| Tremalzo | DTZ | 45°50'15'' | 10°41'23''E | 1694 | 5 |
| Brentonico | DBT | 45°47'16'' | 10°54'04''E | 1482 | 5 |
4.2. DNA Extraction and Polymerase Chain Reaction (PCR) Amplification of Inter-Simple Sequence Repeat Polymorphism (ISSR) Markers
4.3. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aiello, N.; Bontempo, R.; Vender, C. Use of morphological features and amarogentin content for characterization of wild yellow gentian (Gentiana lutea L.) populations in north-east Italy. Acta Bot. Gallica 2013, 160, 33–41. [Google Scholar]
- Struwe, L. Gentian Research Network. Available online: http://gentian.rutgers.edu (accessed on 10 March 2014).
- Ando, H.; Hirai, Y.; Fujii, M.; Hori, Y.; Fukumura, M.; Niiho, Y.; Nakajima, Y.; Shibata, T.; Toriizuka, K.; Ida, Y. The chemical constituents of fresh gentian root. J. Nat. Med. 2007, 61, 269–279. [Google Scholar] [CrossRef]
- Renobales, G. Notas acerca del tratamiento de las gentianaceae para flora ibérica. Anales. Jard. Bot. Madrid 2003, 60, 461–469. (In Spanish) [Google Scholar]
- Ho, T.; Liu, S. A Worldwide Monograph of Gentiana; Science Press: Beijing, China, 2001. [Google Scholar]
- Struwe, L.; Kadereit, J.W.; Klackenberg, J.; Nilsson, S.; Thiv, M.; Hagen, K.B.V.; Albert, V.A. Systematics, character evolution, and biogeography of gentianaceae, including a new tribal and subtribal classification. In Gentianaceae: Systematics and Natural History; Struwe, L., Albert, V.A., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 21–309. [Google Scholar]
- Carnat, A.; Fraisse, D.; Carnat, A.P.; Felgines, C.; Chaud, D.; Lamaison, J.L. Influence of drying mode on iridoid bitter constituent levels in gentian root. J. Sci. Food Agric. 2005, 85, 598–602. [Google Scholar] [CrossRef]
- Sa, O.; Pereira, J.A.; Baptista, P. Optimization of DNA extraction for RAPD and ISSR analysis of Arbutus unedo L. leaves. Int. J. Mol. Sci. 2011, 12, 4156–4164. [Google Scholar] [CrossRef]
- Ge, X.J.; Zhang, L.B.; Yuan, Y.M.; Hao, G.; Chiang, T.Y. Strong genetic differentiation of the east-Himalayan Megacodon stylophorus (Gentianaceae) detected by inter-simple sequence repeats (ISSR). Biodivers. Conserv. 2005, 14, 849–861. [Google Scholar] [CrossRef]
- Zhang, X.L.; Yuan, Y.M.; Ge, X.J. Genetic structure and differentiation of Gentiana atuntsiensis WW Smith and G. striolata TN Ho (Gentianaceae) as revealed by ISSR markers. Bot. J. Linn. Soc. 2007, 154, 225–232. [Google Scholar] [CrossRef]
- Yang, L.C.; Zhou, G.Y.; Chen, G.C. Genetic diversity and population structure of Swertia tetraptera (Gentianaceae), an endemic species of Qinghai-Tibetan plateau. Biochem. Syst. Ecol. 2011, 39, 302–308. [Google Scholar] [CrossRef]
- Zheng, P.; Zhang, K.; Wang, Z. Genetic diversity and gentiopicroside content of four gentiana species in China revealed by ISSR and HPLC methods. Biochem. Syst. Ecol. 2011, 39, 704–710. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Struwe, L.; Albert, V.A. Gentianaceae: Systematics and Natural History; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Gielly, L.; Taberlet, P. A phylogeny of the European gentians inferred from chloroplast trnl (uaa) intron sequences. Bot. J. Linn. Soc. 1996, 120, 57–75. [Google Scholar]
- Lainz, M. Mis Contribuciones al Conocimiento de la Flora de Asturias; (In Spanish). Diputación Provincial de Asturias, Instituto de Estudios Asturianos (del C.S.I.C): Oviedo, Spain, 1982. [Google Scholar]
- Silva, F.J.; Gómez, F.; García, X.R.; Blanco-Dios, J.B. Aportacions a flora de galicia, VI. Nova Acta Cient. Compostel. 2000, 10, 21–33. (In Spanish) [Google Scholar]
- Anchisi, E. Gentiana lutea L., la variabilità morfologica e le sue ibridazioni interspecifiche. Aigba Notes 2008, 30–39. [Google Scholar]
- Bucher, T. Biosystematische Untersuchungen an Gentiana lutea L., Gentiana purpurea L. und deren Hybriden (Biosystematic Investigations on Gentiana lutea L., Gentiana purpurea L. and Their Hybrids). Master’s Thesis, University of Zürich, Zürich, Switzerland, 1987. [Google Scholar]
- Hegi, G. Illustrierte Flora von Mitteleuropa; (In German). Julius Friedrich Lehmann: München, Germany, 1927. [Google Scholar]
- Kozuharova, E. Wild bees as pollinators of four Gentiana species on mount Vitosa (Bulgaria). Bocconea 1997, 5, 619–623. [Google Scholar]
- Kozuharova, E.K.; Anchev, M.E.; Popov, P. The pollination ecology of Gentiana cruciata (Gentianaceae)-specifics of a Bulgarian population in comparison to Dutch populations. Nord. J. Bot. 2003, 23, 365–372. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef]
- Leimu, R.; Mutikainen, P.I.A.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Kery, M.; Matthies, D.; Spillmann, H.H. Reduced fecundity and offspring performance in small populations of the declining grassland plants Primula veris and Gentiana lutea. J. Ecol. 2000, 88, 17–30. [Google Scholar] [CrossRef]
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Shaffer, M.L. Minimum population sizes for species conservation. BioScience 1981, 31, 131–134. [Google Scholar] [CrossRef]
- Schaal, B.A.; Hayworth, D.A.; Olsen, K.M.; Rauscher, J.T.; Smith, W.A. Phylogeographic studies in plants: Problems and prospects. Mol. Ecol. 1998, 7, 465–474. [Google Scholar] [CrossRef]
- Hogbin, P.M.; Peakall, R. Evaluation of the contribution of genetic research to the management of the endangered plant Zieria prostrata. Conserv. Biol. 1999, 13, 514–522. [Google Scholar] [CrossRef]
- Qian, X.; Wang, C.X.; Tian, M. Genetic diversity and population differentiation of Calanthe tsoongiana, a rare and endemic orchid in China. Int. J. Mol. Sci. 2013, 14, 20399–20413. [Google Scholar] [CrossRef]
- Pirttilä, A.; Hirsikorpi, M.; Kämäräinen, T.; Jaakola, L.; Hohtola, A. DNA isolation methods for medicinal and aromatic plants. Plant Mol. Biol. Rep. 2001, 19, 273–273. [Google Scholar] [CrossRef]
- He, T.; Yang, L.; Zhao, Z. Embryogenesis of Gentiana straminea and assessment of genetic stability of regenerated plants using inter simple sequence repeat (ISSR) marker. Afr. J. Biotechol. 2011, 10, 7604–7610. [Google Scholar]
- Gomez-Gomez, L.; Ahrazem, O.; Herranz, J.M.; Ferrandis, P. Genetic characterization and variation within and among populations of Anthyllis rupestris Coss., and endangered endemism of southern Spain. Biochem. Syst. Ecol. 2012, 45, 138–147. [Google Scholar] [CrossRef]
- Yeh, F.C.; Boyle, T.J.B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 157–163. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Lewontin, R.C. The apportionment of human diversity. Evol. Biol. 1972, 6, 381–398. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar]
- PHYLIP (Phylogeny Inference Package). Available online: http://evolution.genetics.washington.edu/phylip.html (accessed on 11 November 2013).
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Peakall, R.; Smouse, P.E. Genalex 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Earl, D.; von Holdt, B. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
González-López, O.; Polanco, C.; György, Z.; Pedryc, A.; Casquero, P.A. Genetic Variation of the Endangered Gentiana lutea L. var. aurantiaca (Gentianaceae) in Populations from the Northwest Iberian Peninsula. Int. J. Mol. Sci. 2014, 15, 10052-10066. https://doi.org/10.3390/ijms150610052
González-López O, Polanco C, György Z, Pedryc A, Casquero PA. Genetic Variation of the Endangered Gentiana lutea L. var. aurantiaca (Gentianaceae) in Populations from the Northwest Iberian Peninsula. International Journal of Molecular Sciences. 2014; 15(6):10052-10066. https://doi.org/10.3390/ijms150610052
Chicago/Turabian StyleGonzález-López, Oscar, Carlos Polanco, Zsuzsanna György, Andrzej Pedryc, and Pedro A. Casquero. 2014. "Genetic Variation of the Endangered Gentiana lutea L. var. aurantiaca (Gentianaceae) in Populations from the Northwest Iberian Peninsula" International Journal of Molecular Sciences 15, no. 6: 10052-10066. https://doi.org/10.3390/ijms150610052