Structural and Molecular Modeling Features of P2X Receptors

Abstract
:1. Introduction
2. General Principles of Ion Channels
3. Molecular Modeling as an Essential Tool for Investigating Ion Channels
3.1. Ligand-Gated Ion Channels
3.2. Pentameric Receptors
3.3. Tetrameric Receptors
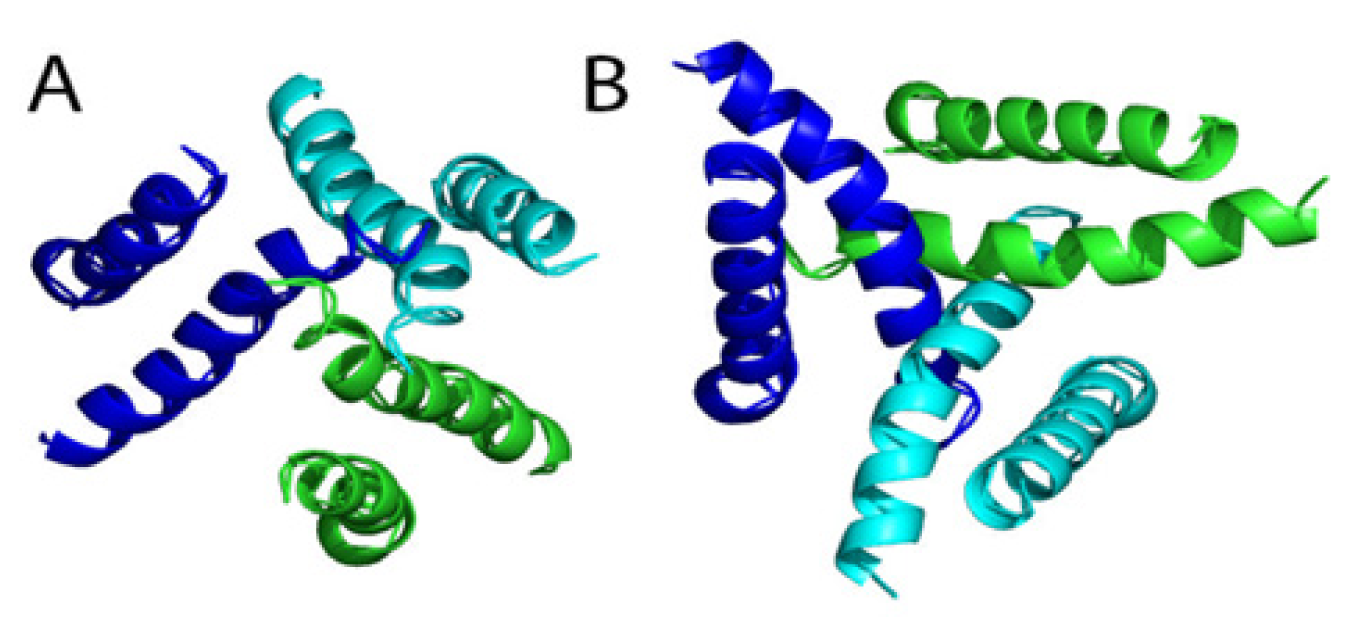
3.4. Trimeric Receptors
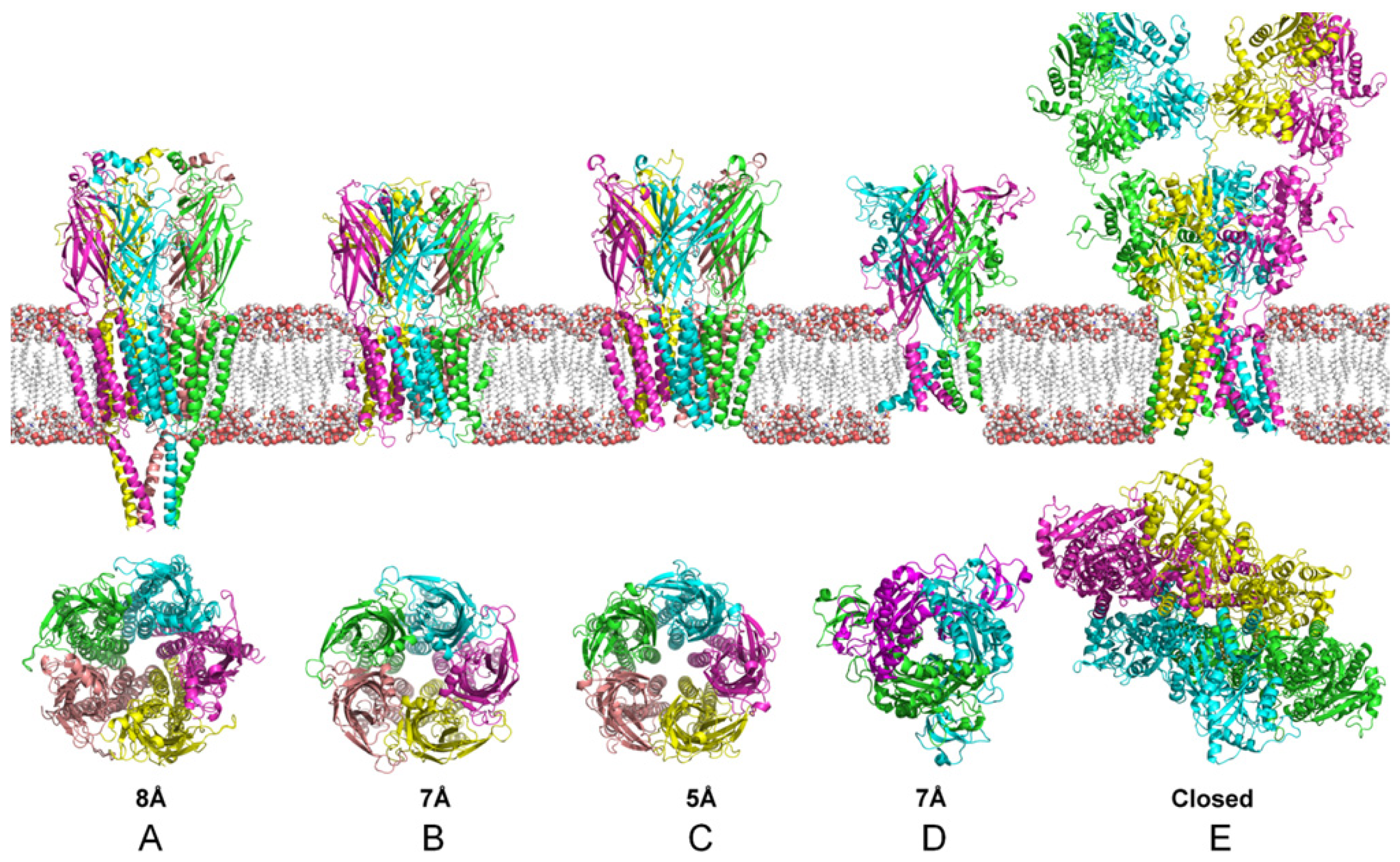
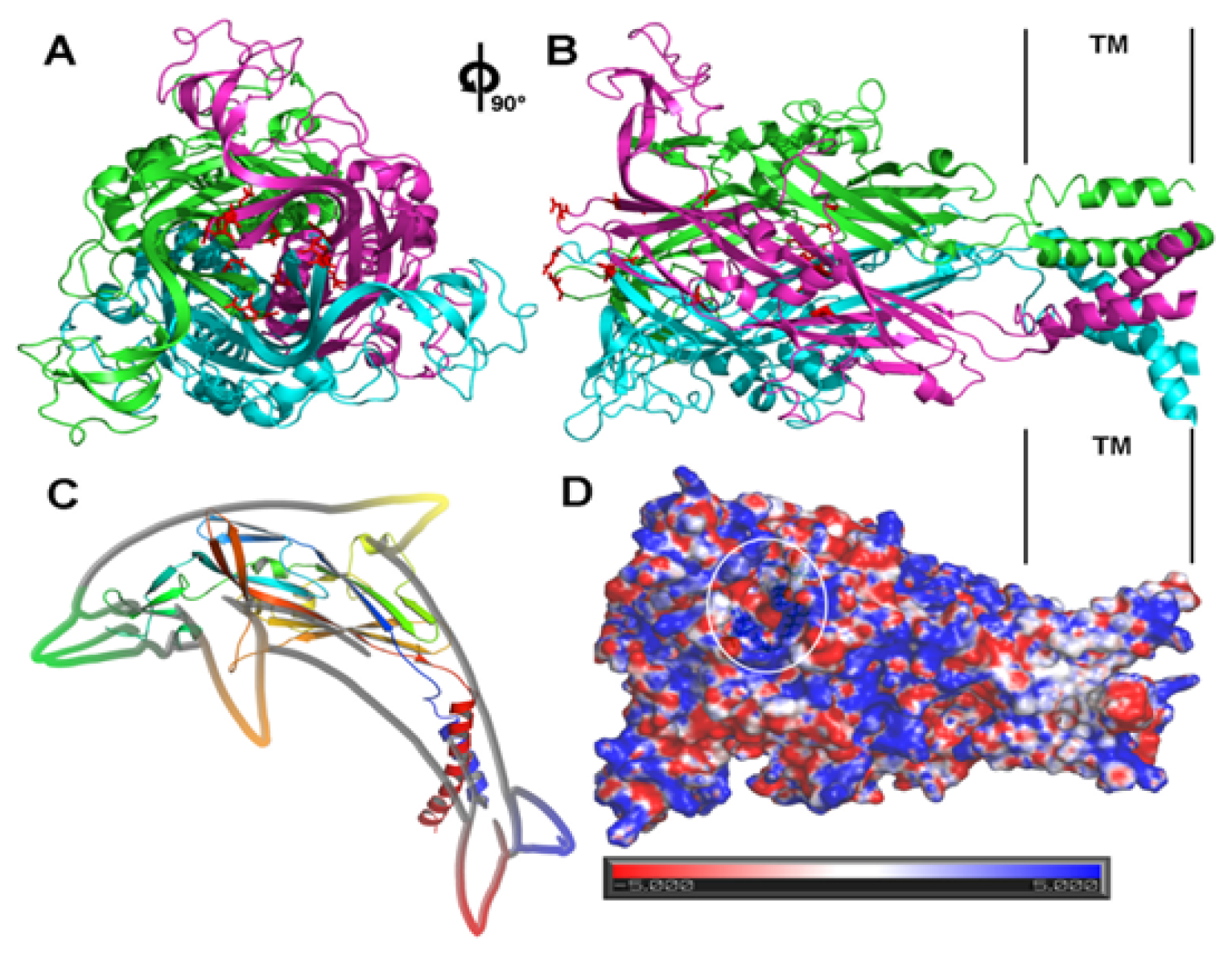
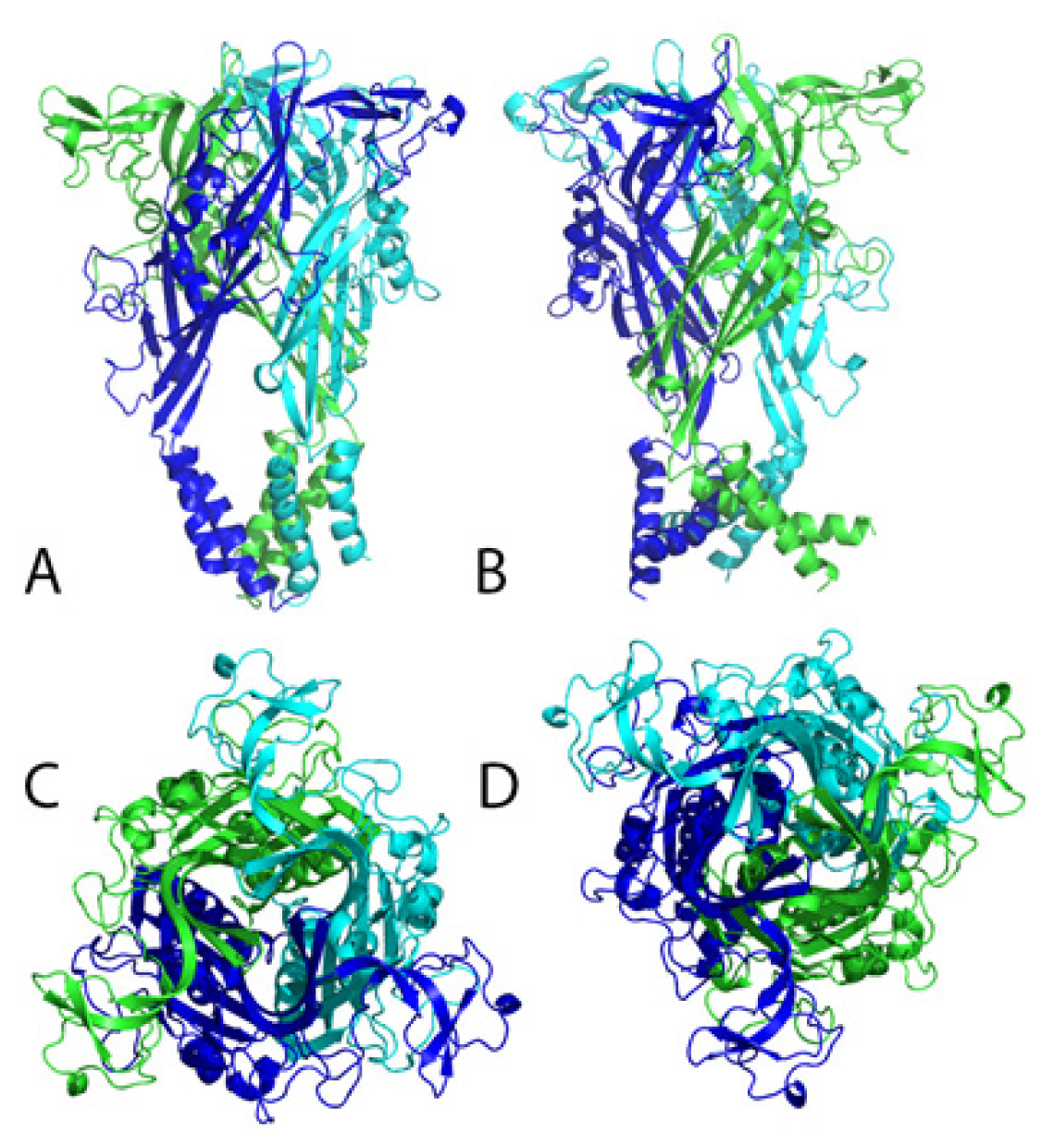
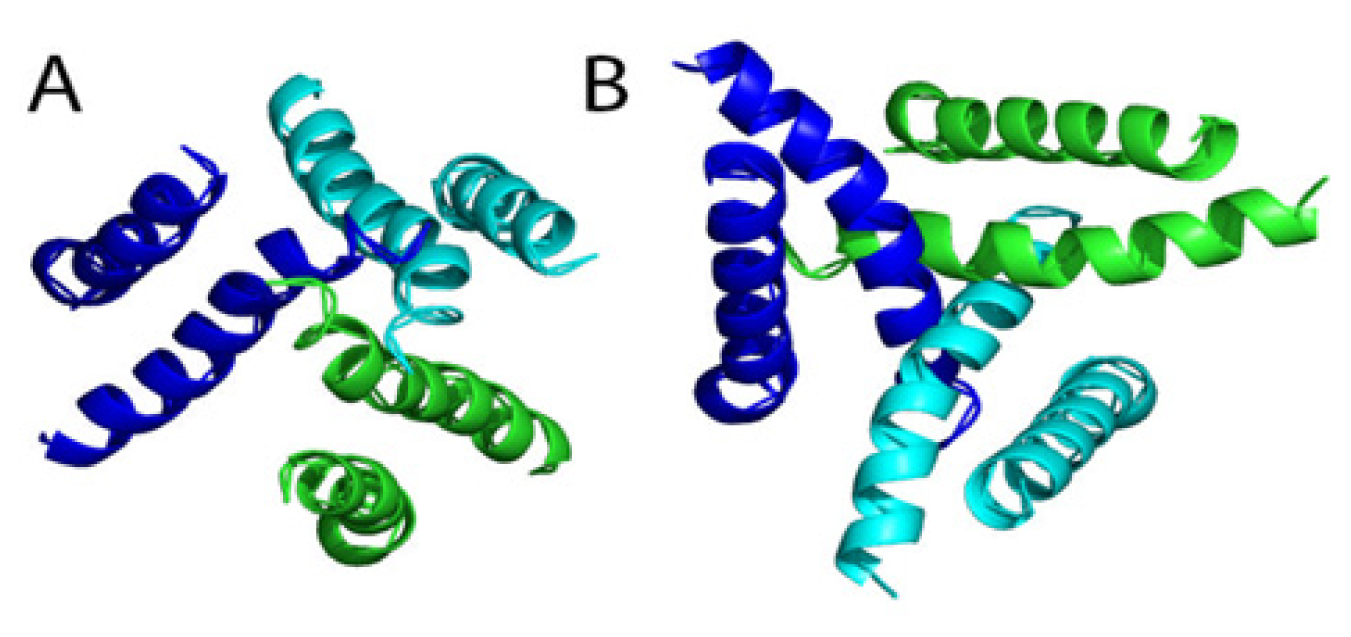
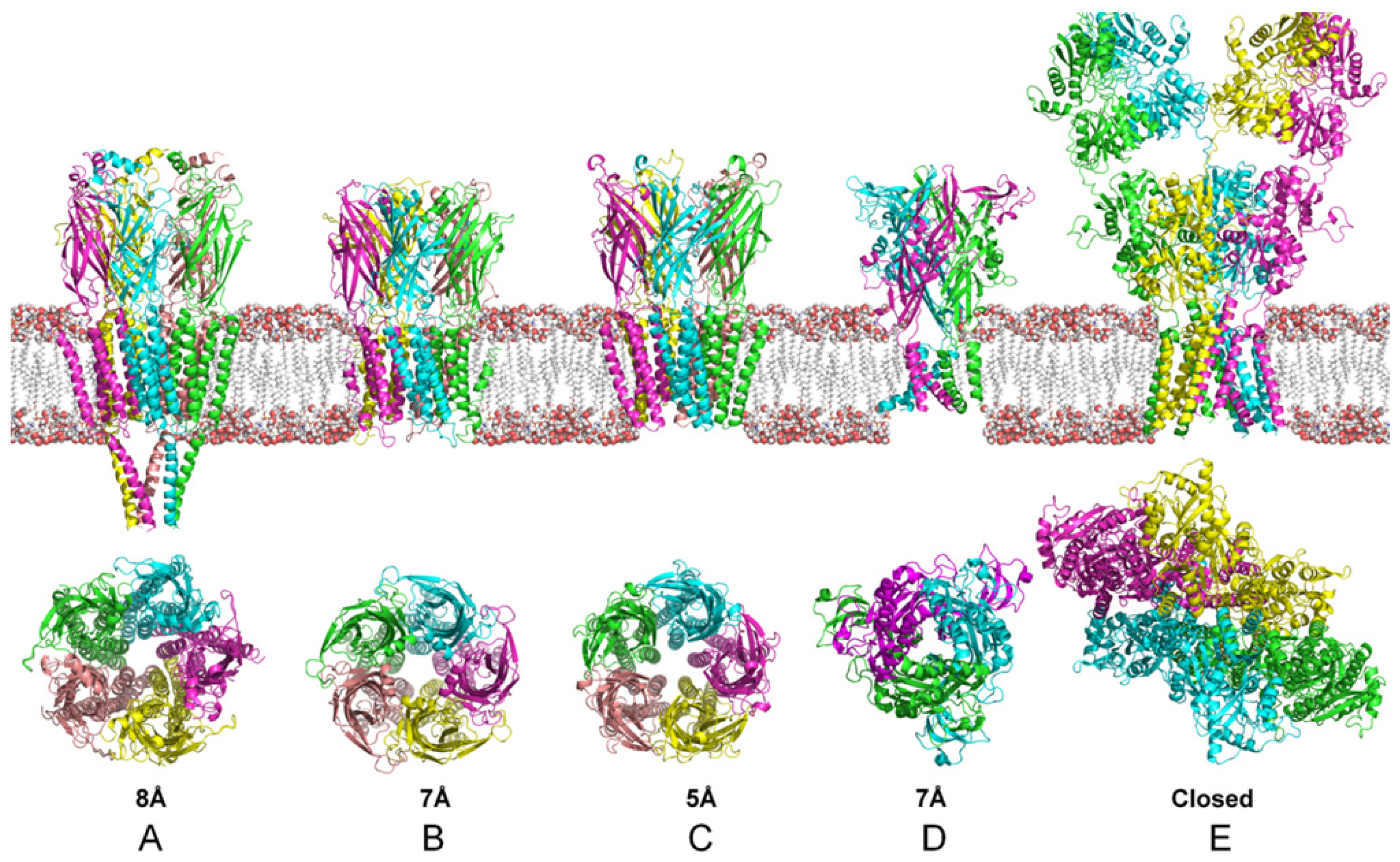
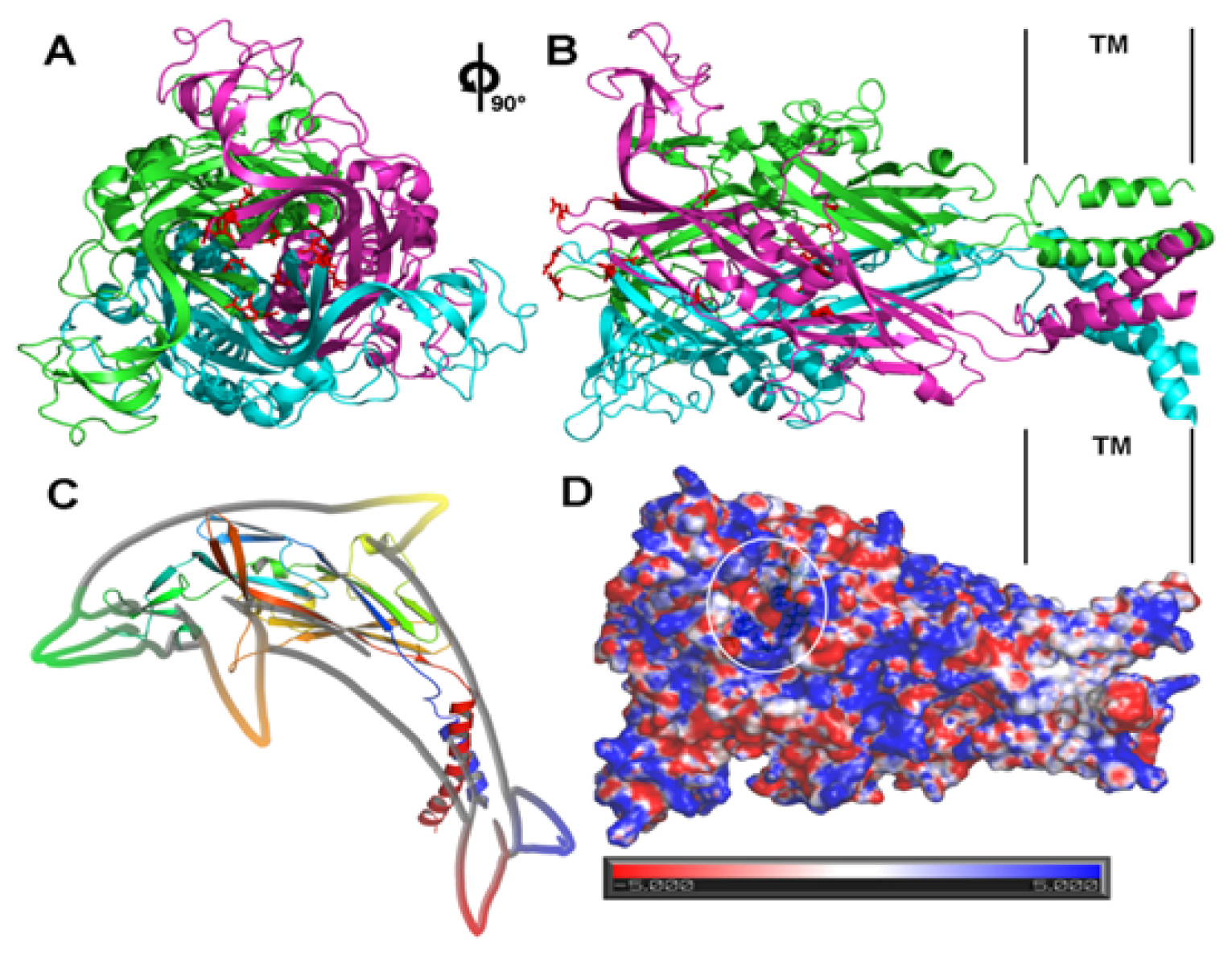
4. Structural and Modeling Aspects of P2XRs
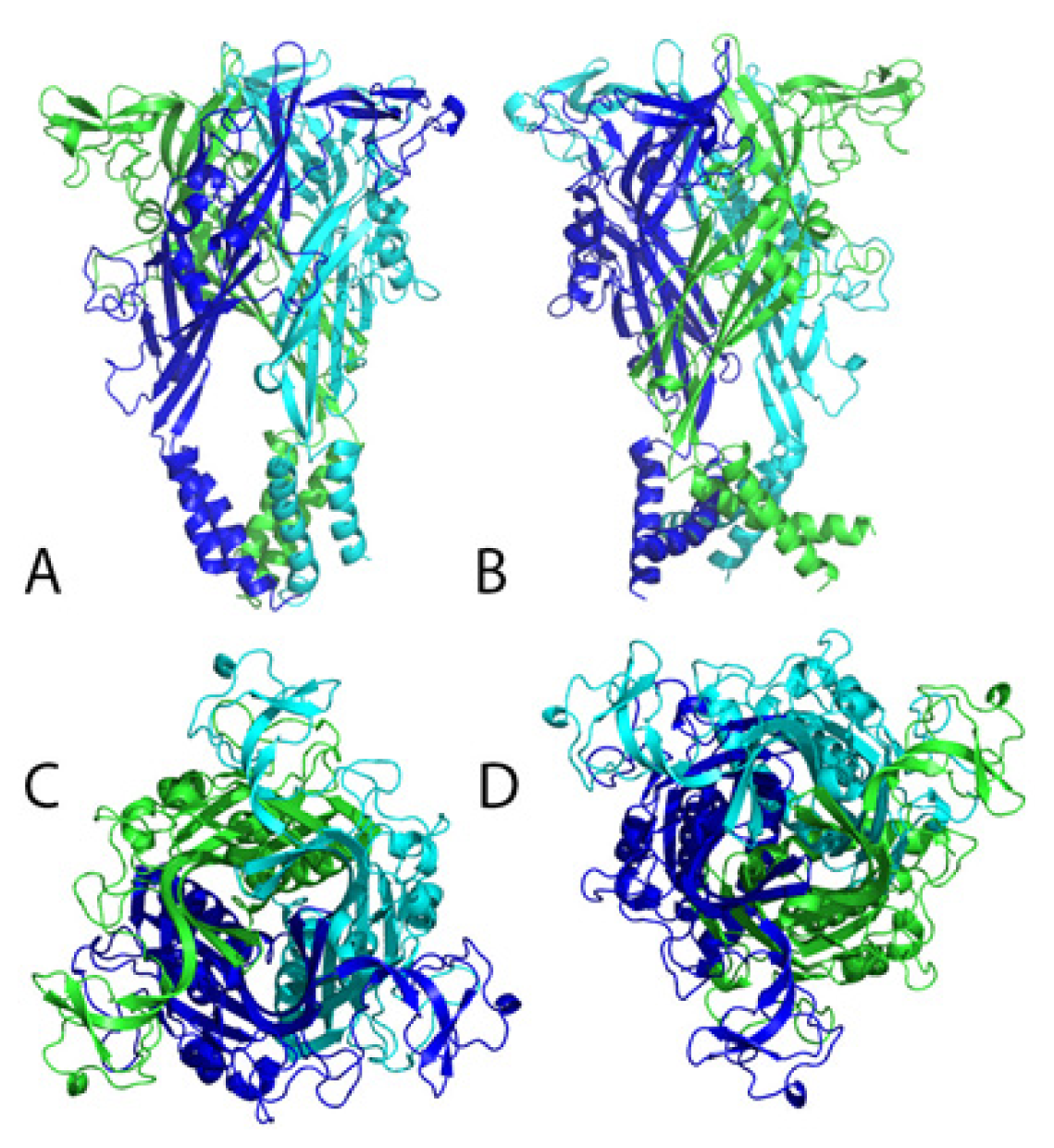
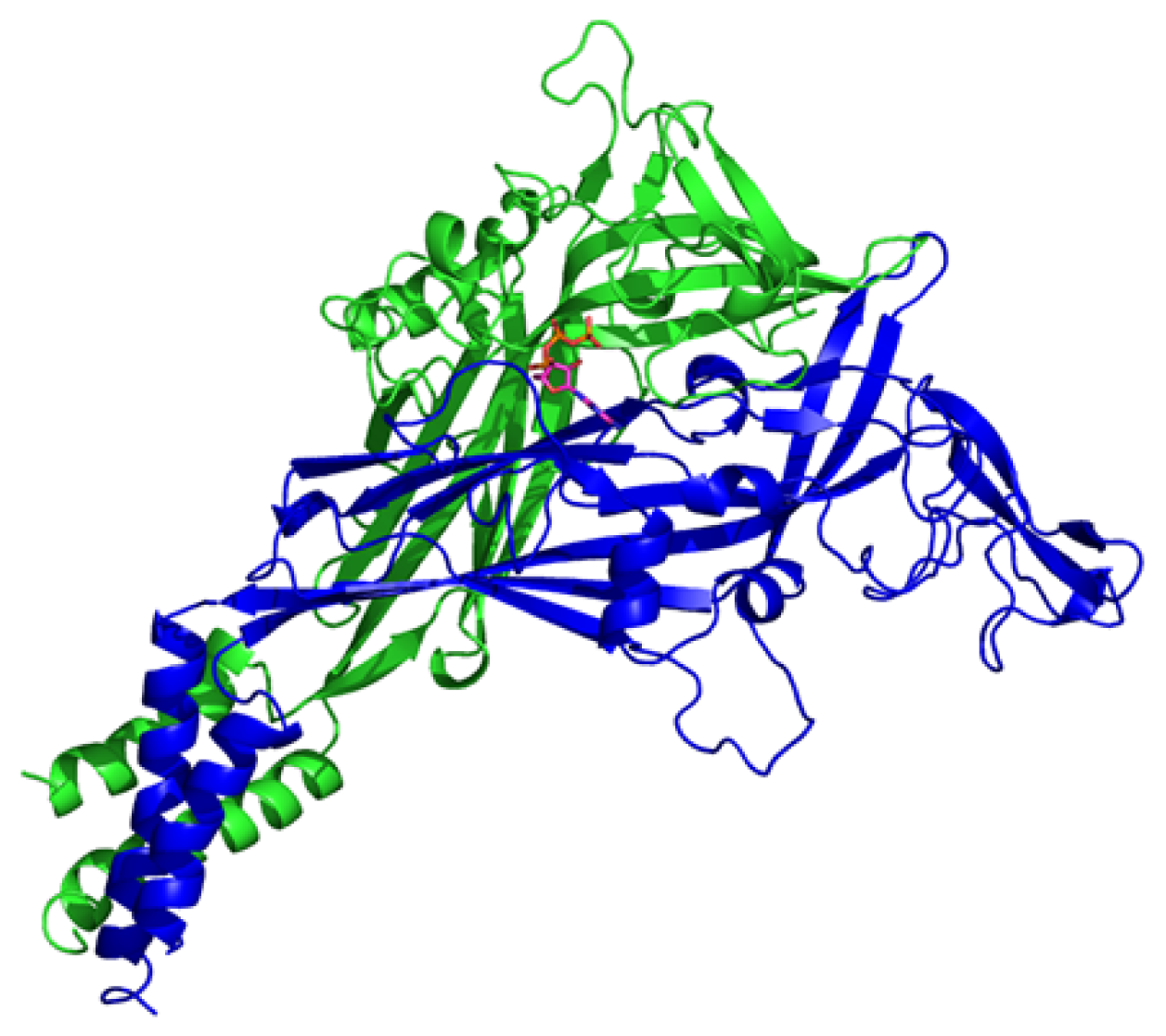
4.1. P2X1 Receptor
4.2. P2X2 Receptor
4.3. P2X3 Receptor
4.4. P2X2/3 Receptors
4.5. P2X4 Receptor
4.6. P2X5 Receptor
4.7. P2X6 Receptor
4.8. P2X7 Receptor
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Holton, F.A.; Holton, P. The capillary dilator substances in dry powders of spinal roots; a possible role of adenosine triphosphate in chemical transmission from nerve endings. J. Physiol. 1954, 126, 124–140. [Google Scholar]
- Burnstock, G. Introductory overview of purinergic signalling. Front. Biosci. 2011, 3, 896–900. [Google Scholar]
- Burnstock, G. Purine and pyrimidine receptors. Cell. Mol. Life Sci. 2007, 64, 1471–1483. [Google Scholar]
- Behrends, J.C. Evolution of the ion channel concept: The historical perspective. Chem. Rev. 2012, 112, 6218–6226. [Google Scholar]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar]
- Changeux, J.P.; Gautron, J.; Israel, M.; Podleski, T. Separation of excitable membranes from the electric organ of electrophorus electricus. C. R. Acad. Sci. D 1969, 269, 1788–1791. [Google Scholar]
- Dupont, Y.; Cohen, J.B.; Changeux, J.P. X-ray diffraction study of membrane fragments rich in acetylcholine receptor protein prepared from the electric organ of torpedo marmorata. FEBS Lett. 1974, 40, 130–133. [Google Scholar]
- MacKinnon, R. Potassium channels. FEBS Lett. 2003, 555, 62–65. [Google Scholar]
- Gouaux, E.; Mackinnon, R. Principles of selective ion transport in channels and pumps. Science 2005, 310, 1461–1465. [Google Scholar]
- Jensen, M.O.; Jogini, V.; Borhani, D.W.; Leffler, A.E.; Dror, R.O.; Shaw, D.E. Mechanism of voltage gating in potassium channels. Science 2012, 336, 229–233. [Google Scholar]
- Law, R.J.; Tieleman, D.P.; Sansom, M.S. Pores formed by the nicotinic receptor m2delta peptide: A molecular dynamics simulation study. Biophys. J. 2003, 84, 14–27. [Google Scholar]
- Law, R.J.; Forrest, L.R.; Ranatunga, K.M.; la Rocca, P.; Tieleman, D.P.; Sansom, M.S. Structure and dynamics of the pore-lining helix of the nicotinic receptor: Md simulations in water lipid bilayers and transbilayer bundles. Proteins 2000, 39, 47–55. [Google Scholar]
- Kozuska, J.L.; Paulsen, I.M. The Cys-loop pentameric ligand-gated ion channel receptors: 50 years on. Can. J. Physiol. Pharmacol. 2012, 90, 771–782. [Google Scholar]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure regulation and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar]
- Gonzales, E.B.; Kawate, T.; Gouaux, E. Pore architecture and ion sites in acid-sensing ion channels and P2X receptors. Nature 2009, 460, 599–604. [Google Scholar]
- Keramidas, A.; Moorhouse, A.J.; Schofield, P.R.; Barry, P.H. Ligand-gated ion channels: Mechanisms underlying ion selectivity. Prog. Biophys. Mol. Biol. 2004, 86, 161–204. [Google Scholar]
- Bertrand, D. Neurocircuitry of the nicotinic cholinergic system. Dialog. Clin. Neurosci. 2010, 12, 463–470. [Google Scholar]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar]
- Dingledine, R.; Borges, K.; Bowie, D.; Traynelis, S.F. The glutamate receptor ion channels. Pharmacol. Rev. 1999, 51, 7–61. [Google Scholar]
- Erreger, K.; Chen, P.E.; Wyllie, D.J.; Traynelis, S.F. Glutamate receptor gating. Crit. Rev. Neurobiol. 2004, 16, 187–224. [Google Scholar]
- Mayer, M.L.; Westbrook, G.L.; Guthrie, P.B. Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature 1984, 309, 261–263. [Google Scholar]
- Johnson, J.W.; Ascher, P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature 1987, 325, 529–531. [Google Scholar]
- North, R.A.; Jarvis, M.F. P2X receptors as drug targets. Mol. Pharmacol. 2013, 83, 759–769. [Google Scholar]
- Lemoine, D.; Jiang, R.; Taly, A.; Chataigneau, T.; Specht, A.; Grutter, T. Ligand-gated ion channels: New insights into neurological disorders and ligand recognition. Chem. Rev. 2012, 112, 6285–6318. [Google Scholar]
- Egan, T.M.; Haines, W.R.; Voigt, M.M. A domain contributing to the ion channel of ATP-gated P2X2 receptors identified by the substituted cysteine accessibility method. J. Neurosci. 1998, 18, 2350–2359. [Google Scholar]
- Kracun, S.; Chaptal, V.; Abramson, J.; Khakh, B.S. Gated access to the pore of a P2X receptor: Structural implications for closed-open transitions. J. Boil. Chem. 2010, 285, 10110–10121. [Google Scholar]
- Rassendren, F.; Buell, G.; Newbolt, A.; North, R.A.; Surprenant, A. Identification of amino acid residues contributing to the pore of a P2X receptor. EMBO J. 1997, 16, 3446–3454. [Google Scholar]
- Li, M.; Chang, T.H.; Silberberg, S.D.; Swartz, K.J. Gating the pore of P2X receptor channels. Nat. Neurosci. 2008, 11, 883–887. [Google Scholar]
- Khakh, B.S.; North, R.A. Neuromodulation by extracellular ATP and P2X receptors in the CNS. Neuron 2012, 76, 51–69. [Google Scholar]
- North, R.A.; Surprenant, A. Pharmacology of cloned P2X receptors. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 563–580. [Google Scholar]
- Sun, C.; Heid, M.E.; Keyel, P.A.; Salter, R.D. The second transmembrane domain of P2X7 contributes to dilated pore formation. PLoS One 2013, 8, e61886. [Google Scholar]
- Browne, L.E.; Compan, V.; Bragg, L.; North, R.A. P2X7 receptor channels allow direct permeation of nanometer-sized dyes. J. Neurosci. 2013, 33, 3557–3566. [Google Scholar]
- Roberts, J.A.; Evans, R.J. ATP binding at human P2X1 receptors Contribution of aromatic and basic amino acids revealed using mutagenesis and partial agonists. J. Biol. Chem. 2004, 279, 9043–9055. [Google Scholar]
- Hattori, M.; Gouaux, E. Molecular mechanism of ATP binding and ion channel activation in P2X receptors. Nature 2012, 485, 207–212. [Google Scholar]
- Kawate, T.; Michel, J.C.; Birdsong, W.T.; Gouaux, E. Crystal structure of the ATP-gated P2X(4) ion channel in the closed state. Nature 2009, 460, 592–598. [Google Scholar]
- Werner, P.; Seward, E.P.; Buell, G.N.; North, R.A. Domains of P2X receptors involved in desensitization. Proc. Natl. Acad. Sci. USA 1996, 93, 15485–15490. [Google Scholar]
- Nicke, A.; Kerschensteiner, D.; Soto, F. Biochemical and functional evidence for heteromeric assembly of P2X1 and P2X4 subunits. J. Neurochem. 2005, 92, 925–933. [Google Scholar]
- Allsopp, R.C.; El Ajouz, S.; Schmid, R.; Evans, R.J. Cysteine scanning mutagenesis (residues glu52-gly96) of the human P2X1 receptor for ATP: Mapping agonist binding and channel gating. J. Biol. Chem. 2011, 286, 29207–29217. [Google Scholar]
- Roberts, J.A.; Allsopp, R.C.; El Ajouz, S.; Vial, C.; Schmid, R.; Young, M.T.; Evans, R.J. Agonist binding evokes extensive conformational changes in the extracellular domain of the ATP-gated human P2X1 receptor ion channel. Proc. Natl. Acad. Sci. USA 2012, 109, 4663–4667. [Google Scholar]
- Roberts, J.A.; Vial, C.; Digby, H.R.; Agboh, K.C.; Wen, H.; Atterbury-Thomas, A.; Evans, R.J. Molecular properties of P2X receptors. Pflugers Arch. 2006, 452, 486–500. [Google Scholar]
- Virginio, C.; North, R.A.; Surprenant, A. Calcium permeability and block at homomeric and heteromeric P2X2 and P2X3 receptors and P2X receptors in rat nodose neurones. J. Physiol. 1998, 510, 27–35. [Google Scholar]
- Jiang, L.H.; Rassendren, F.; Surprenant, A.; North, R.A. Identification of amino acid residues contributing to the ATP-binding site of a purinergic P2X receptor. J. Biol. Chem. 2000, 275, 34190–34196. [Google Scholar]
- Jiang, R.; Lemoine, D.; Martz, A.; Taly, A.; Gonin, S.; Prado de Carvalho, L.; Specht, A.; Grutter, T. Agonist trapped in ATP-binding sites of the P2X2 receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 9066–9071. [Google Scholar]
- Jiang, R.; Taly, A.; Grutter, T. Moving through the gate in ATP-activated P2X receptors. Trends Biochem. Sci. 2013, 38, 20–29. [Google Scholar]
- Torres, G.E.; Egan, T.M.; Voigt, M.M. Hetero-oligomeric assembly of P2X receptor subunits Specificities exist with regard to possible partners. J. Biol. Chem. 1999, 274, 6653–6659. [Google Scholar]
- Rothwell, S.W.; Stansfeld, P.J.; Bragg, L.; Verkhratsky, A.; North, R.A. Direct gating of ATP-activated ion channels (P2X2 receptors) by lipophilic attachment at the outer end of the second transmembrane domain. J. Biol. Chem. 2013. [Google Scholar] [CrossRef]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar]
- Cockayne, D.A.; Dunn, P.M.; Zhong, Y.; Rong, W.; Hamilton, S.G.; Knight, G.E.; Ruan, H.Z.; Ma, B.; Yip, P.; Nunn, P.; et al. P2X2 knockout mice and P2X2/P2X3 double knockout mice reveal a role for the P2X2 receptor subunit in mediating multiple sensory effects of ATP. J. Physiol. 2005, 567, 621–639. [Google Scholar]
- Bodnar, M.; Wang, H.; Riedel, T.; Hintze, S.; Kato, E.; Fallah, G.; Groger-Arndt, H.; Giniatullin, R.; Grohmann, M.; Hausmann, R.; et al. Amino acid residues constituting the agonist binding site of the human P2X3 receptor. J. Biol. Chem. 2011, 286, 2739–2749. [Google Scholar]
- Petrenko, N.; Khafizov, K.; Tvrdonova, V.; Skorinkin, A.; Giniatullin, R. Role of the ectodomain serine 275 in shaping the binding pocket of the ATP-gated P2X3 receptor. Biochemistry 2011, 50, 8427–8436. [Google Scholar]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar]
- The PyMOL Molecular Graphics System. Available online: http://www.pymol.org/ (accessed on 13 December 2013).
- Duckwitz, W.; Hausmann, R.; Aschrafi, A.; Schmalzing, G. P2X5 subunit assembly requires scaffolding by the second transmembrane domain and a conserved aspartate. J. Biol. Chem. 2006, 281, 39561–39572. [Google Scholar]
- Lalo, U.; Pankratov, Y.; Wichert, S.P.; Rossner, M.J.; North, R.A.; Kirchhoff, F.; Verkhratsky, A. P2X1 and P2X5 subunits form the functional P2X receptor in mouse cortical astrocytes. J. Neurosci. 2008, 28, 5473–5480. [Google Scholar]
- Compan, V.; Ulmann, L.; Stelmashenko, O.; Chemin, J.; Chaumont, S.; Rassendren, F. P2X2 and P2X5 subunits define a new heteromeric receptor with P2X7-like properties. J. Neurosci. 2012, 32, 4284–4296. [Google Scholar]
- King, B.F.; Townsend-Nicholson, A.; Wildman, S.S.; Thomas, T.; Spyer, K.M.; Burnstock, G. Coexpression of rat P2X2 and P2X6 subunits in Xenopus oocytes. J. Neurosci. 2000, 20, 4871–4877. [Google Scholar]
- Le, K.T.; Babinski, K.; Seguela, P. Central P2X4 and P2X6 channel subunits coassemble into a novel heteromeric atp receptor. J. Neurosci. 1998, 18, 7152–7159. [Google Scholar]
- Ormond, S.J.; Barrera, N.P.; Qureshi, O.S.; Henderson, R.M.; Edwardson, J.M.; Murrell-Lagnado, R.D. An uncharged region within the N terminus of the P2X6 receptor inhibits its assembly and exit from the endoplasmic reticulum. Mol. Pharmacol. 2006, 69, 1692–1700. [Google Scholar]
- Costa-Junior, H.M.; Sarmento Vieira, F.; Coutinho-Silva, R. C terminus of the P2X7 receptor: Treasure hunting. Purinergic Signal. 2011, 7, 7–19. [Google Scholar]
- Denlinger, L.C.; Fisette, P.L.; Sommer, J.A.; Watters, J.J.; Prabhu, U.; Dubyak, G.R.; Proctor, R.A.; Bertics, P.J. Cutting edge: The nucleotide receptor P2X7 contains multiple proteinand lipid-interaction motifs including a potential binding site for bacterial lipopolysaccharide. J. Immunol. 2001, 167, 1871–1876. [Google Scholar]
- Adriouch, S.; Bannas, P.; Schwarz, N.; Fliegert, R.; Guse, A.H.; Seman, M.; Haag, F.; Koch-Nolte, F. Adp-ribosylation at R125 gates the P2X7 ion channel by presenting a covalent ligand to its nucleotide binding site. FASEB J. 2008, 22, 861–869. [Google Scholar]
- Smart, M.L.; Gu, B.; Panchal, R.G.; Wiley, J.; Cromer, B.; Williams, D.A.; Petrou, S. P2X7 receptor cell surface expression and cytolytic pore formation are regulated by a distal C-terminal region. J. Biol. Chem. 2003, 278, 8853–8860. [Google Scholar]
- Guile, S.D.; Alcaraz, L.; Birkinshaw, T.N.; Bowers, K.C.; Ebden, M.R.; Furber, M.; Stocks, M.J. Antagonists of the P2X(7) receptor From lead identification to drug development. J. Med. Chem. 2009, 52, 3123–3141. [Google Scholar]
- Gum, R.J.; Wakefield, B.; Jarvis, M.F. P2X receptor antagonists for pain management: Examination of binding and physicochemical properties. Purinergic Signal. 2012, 8, 41–56. [Google Scholar]
- Jiang, L.H.; Baldwin, J.M.; Roger, S.; Baldwin, S.A. Insights into the molecular mechanisms underlying mammalian P2X7 receptor functions and contributions in diseases revealed by structural modeling and single nucleotide polymorphisms. Front. Pharmacol. 2013, 4, 55. [Google Scholar]
- Roger, S.; Mei, Z.Z.; Baldwin, J.M.; Dong, L.; Bradley, H.; Baldwin, S.A.; Surprenant, A.; Jiang, L.H. Single nucleotide polymorphisms that were identified in affective mood disorders affect atp-activated P2X7 receptor functions. J. Psychiatr. Res. 2010, 44, 347–355. [Google Scholar]
- Liu, X.; Surprenant, A.; Mao, H.J.; Roger, S.; Xia, R.; Bradley, H.; Jiang, L.H. Identification of key residues coordinating functional inhibition of P2X7 receptors by zinc and copper. Mol. Pharmacol. 2008, 73, 252–259. [Google Scholar]
- Adriouch, S.; Scheuplein, F.; Bahring, R.; Seman, M.; Boyer, O.; Koch-Nolte, F.; Haag, F. Characterisation of the R276a gain-of-function mutation in the ectodomain of murine P2X7. Purinergic Signal. 2009, 5, 151–161. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Pentameric receptors | Permeant ions |
|---|---|---|
| Glycine | GyR (vertebrates) | anions |
| GABA | GABAAR (vertebrates) | anions |
| ACh | nAChR (vertebrates) | cations |
| 5-HT | 5-HT3R (vertebrates) | cations |
| Zn2+ | ZACR (vertebrates) | cations |
| Glutamate | GluR (invertebrates) | cations |
| 5-HT | MOD-1 (invertebrates) | anions |
| GABA | EXP-1 (invertebrates) | cations |
| Tetrameric-Ionotropic glutamate receptors | ||
| Glutamate | iGluRs | |
| Glutamate | AMPARs | |
| Glycine and Glutamate | Kainate | cations |
| NMDARs | ||
| Trimeric-Purinergic receptors | ||
| ATP | P2XRs | cations |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Alves, L.A.; Da Silva, J.H.M.; Ferreira, D.N.M.; Fidalgo-Neto, A.A.; Teixeira, P.C.N.; De Souza, C.A.M.; Caffarena, E.R.; De Freitas, M.S. Structural and Molecular Modeling Features of P2X Receptors. Int. J. Mol. Sci. 2014, 15, 4531-4549. https://doi.org/10.3390/ijms15034531
Alves LA, Da Silva JHM, Ferreira DNM, Fidalgo-Neto AA, Teixeira PCN, De Souza CAM, Caffarena ER, De Freitas MS. Structural and Molecular Modeling Features of P2X Receptors. International Journal of Molecular Sciences. 2014; 15(3):4531-4549. https://doi.org/10.3390/ijms15034531
Chicago/Turabian StyleAlves, Luiz Anastacio, João Herminio Martins Da Silva, Dinarte Neto Moreira Ferreira, Antonio Augusto Fidalgo-Neto, Pedro Celso Nogueira Teixeira, Cristina Alves Magalhães De Souza, Ernesto Raúl Caffarena, and Mônica Santos De Freitas. 2014. "Structural and Molecular Modeling Features of P2X Receptors" International Journal of Molecular Sciences 15, no. 3: 4531-4549. https://doi.org/10.3390/ijms15034531