Long Non-Coding RNA HOTAIR Promotes Cell Migration and Invasion via Down-Regulation of RNA Binding Motif Protein 38 in Hepatocellular Carcinoma Cells

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
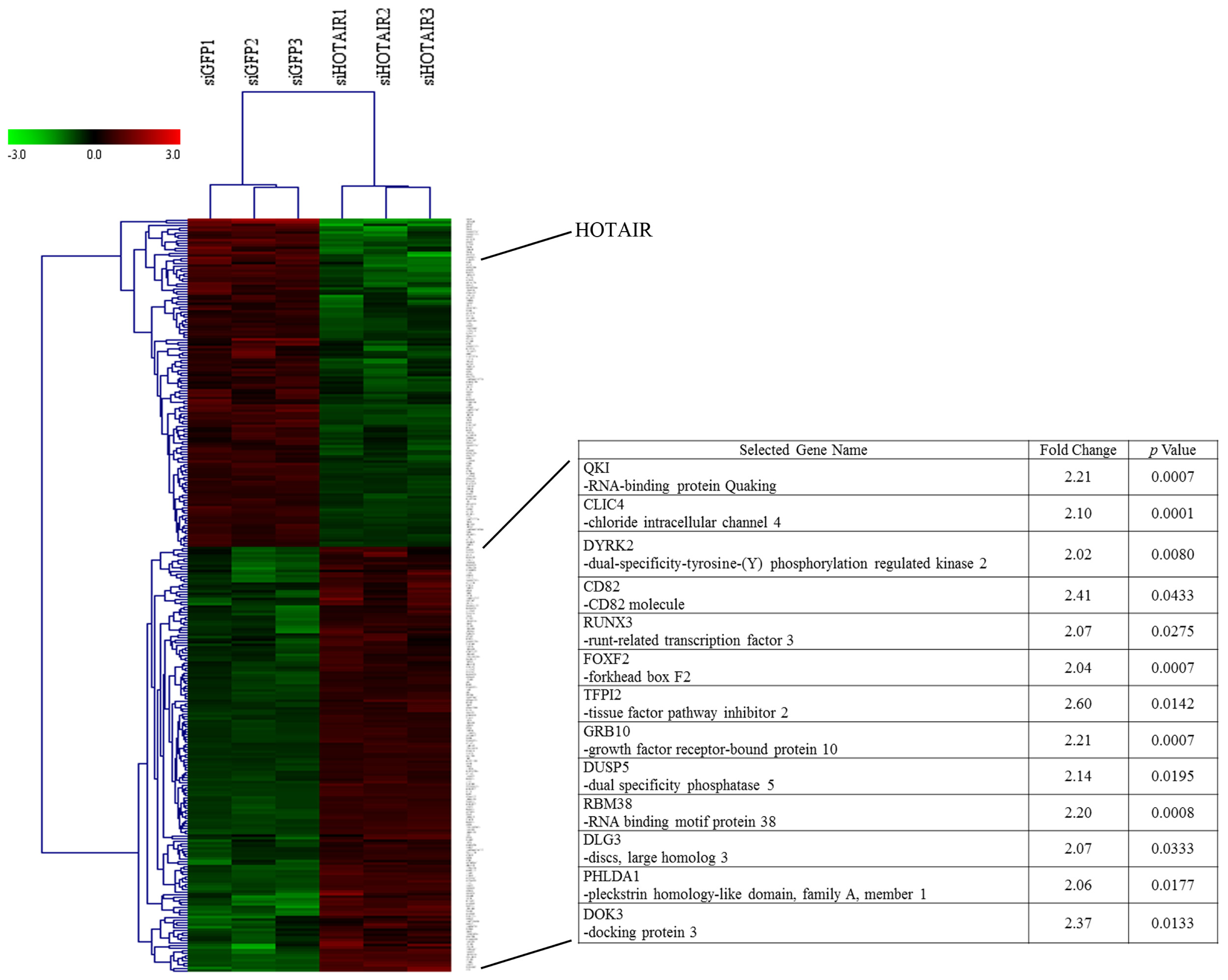
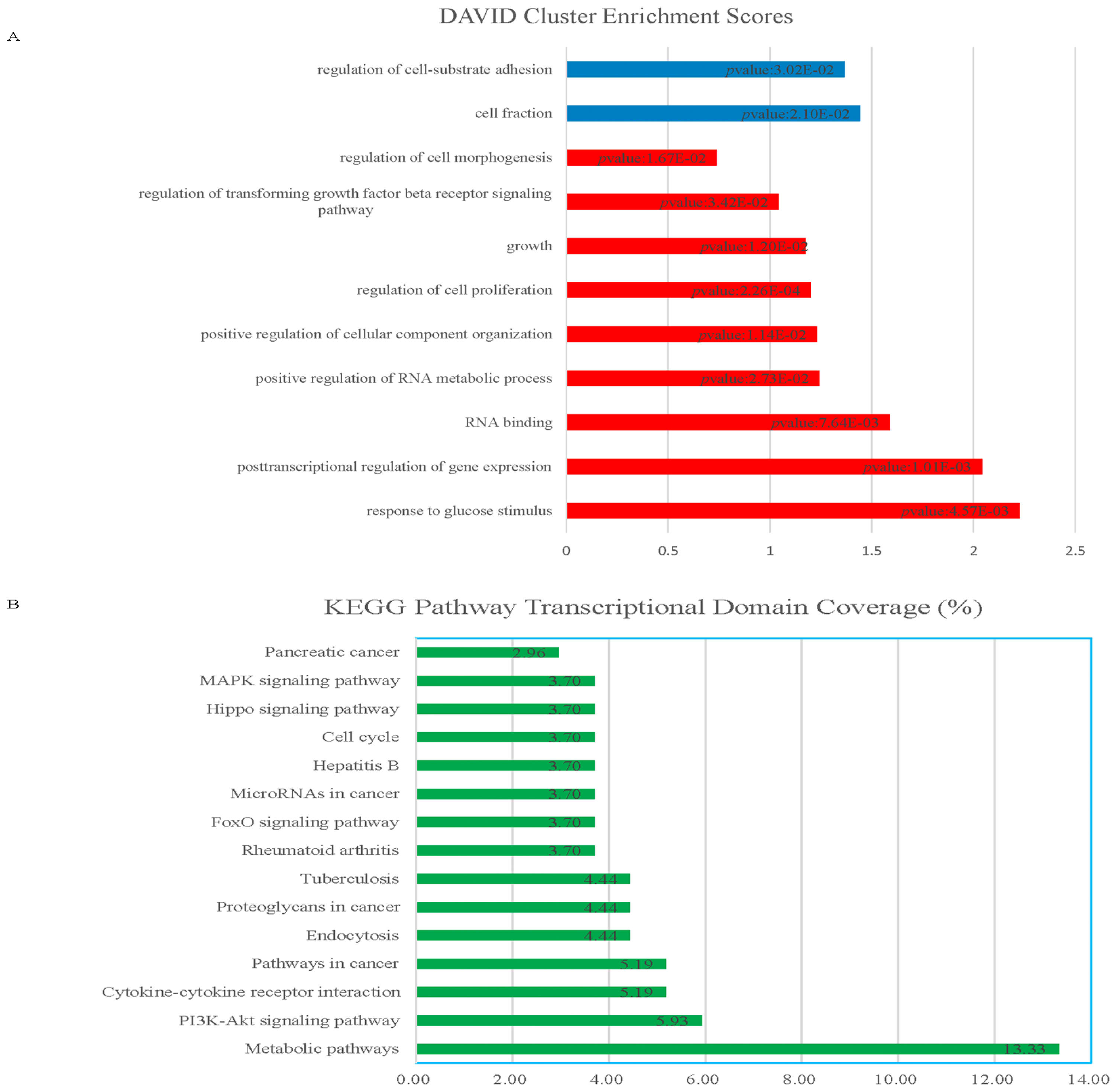
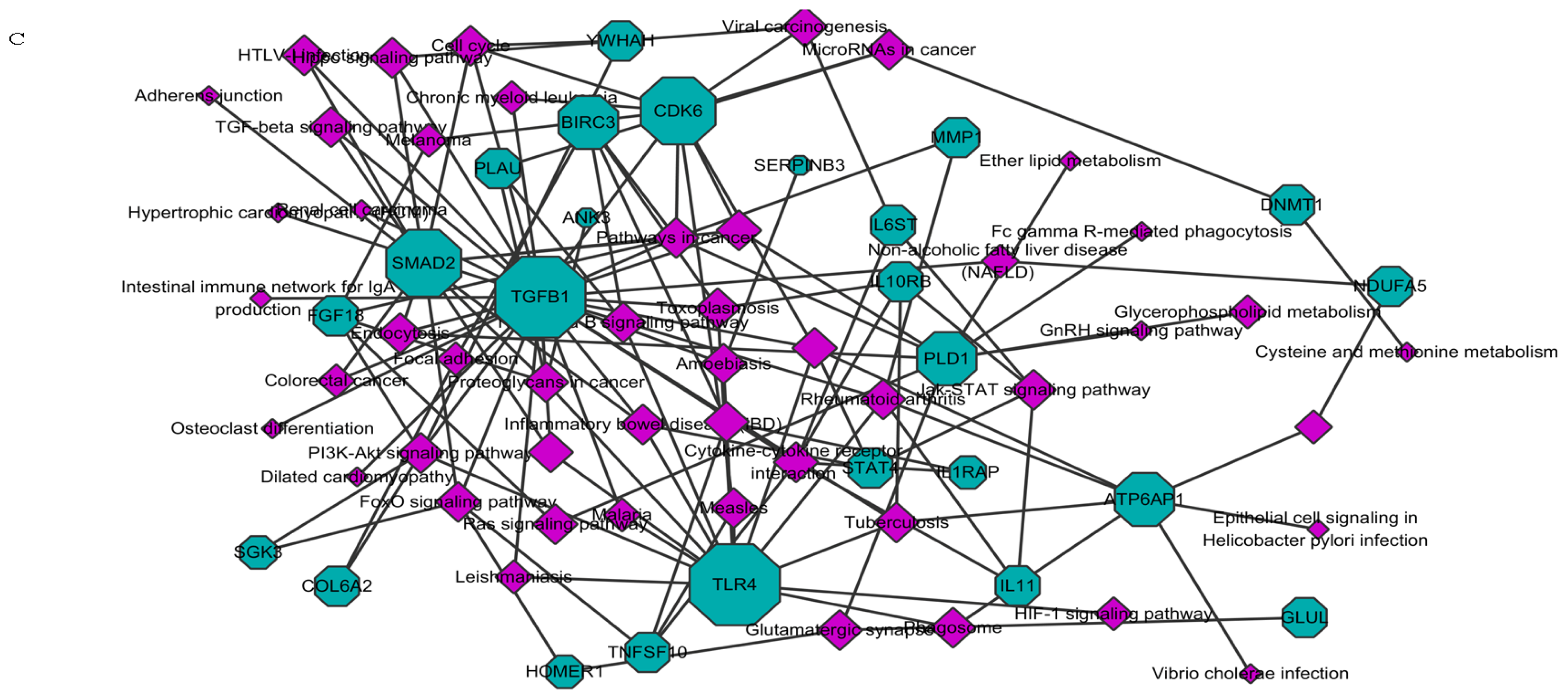
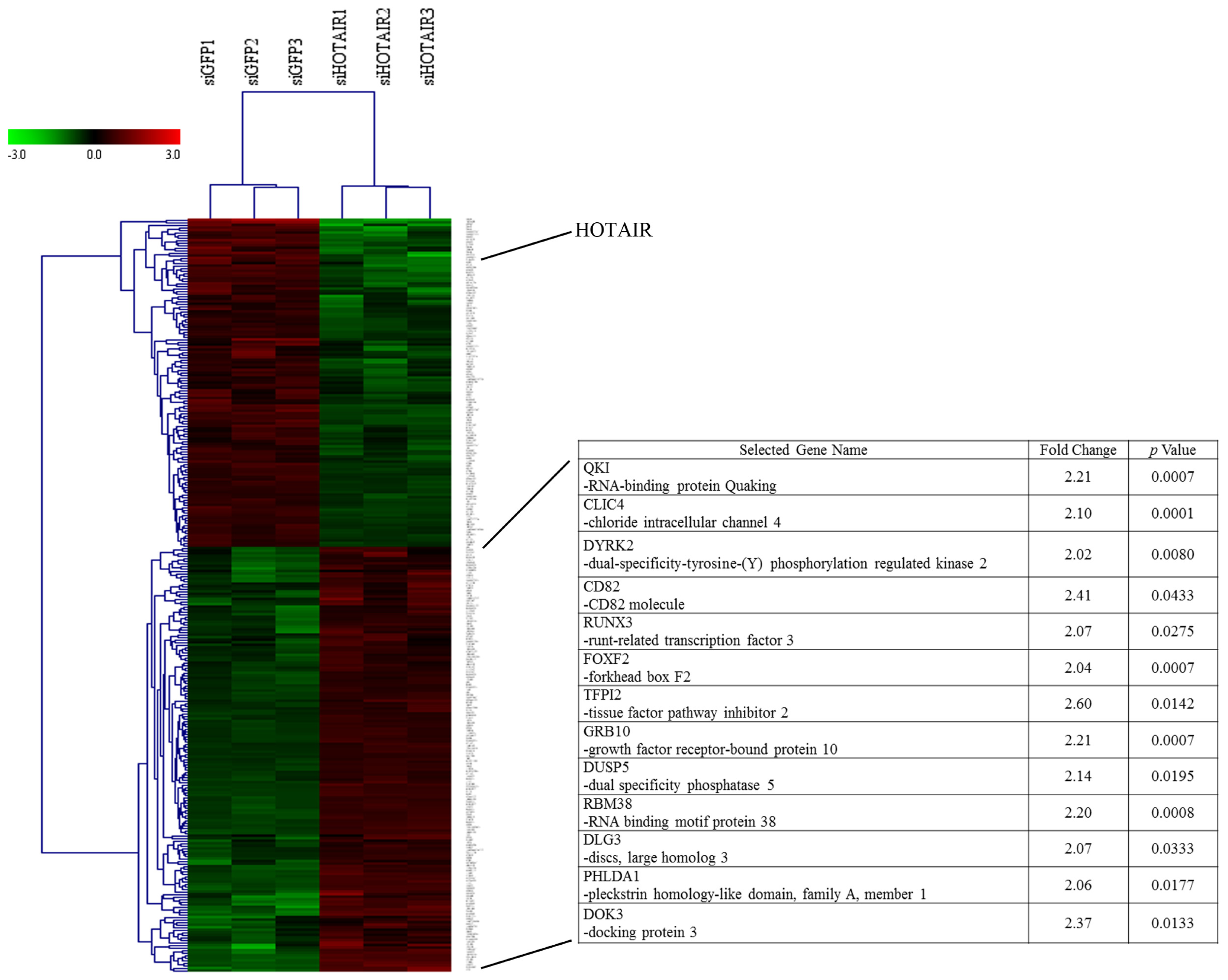
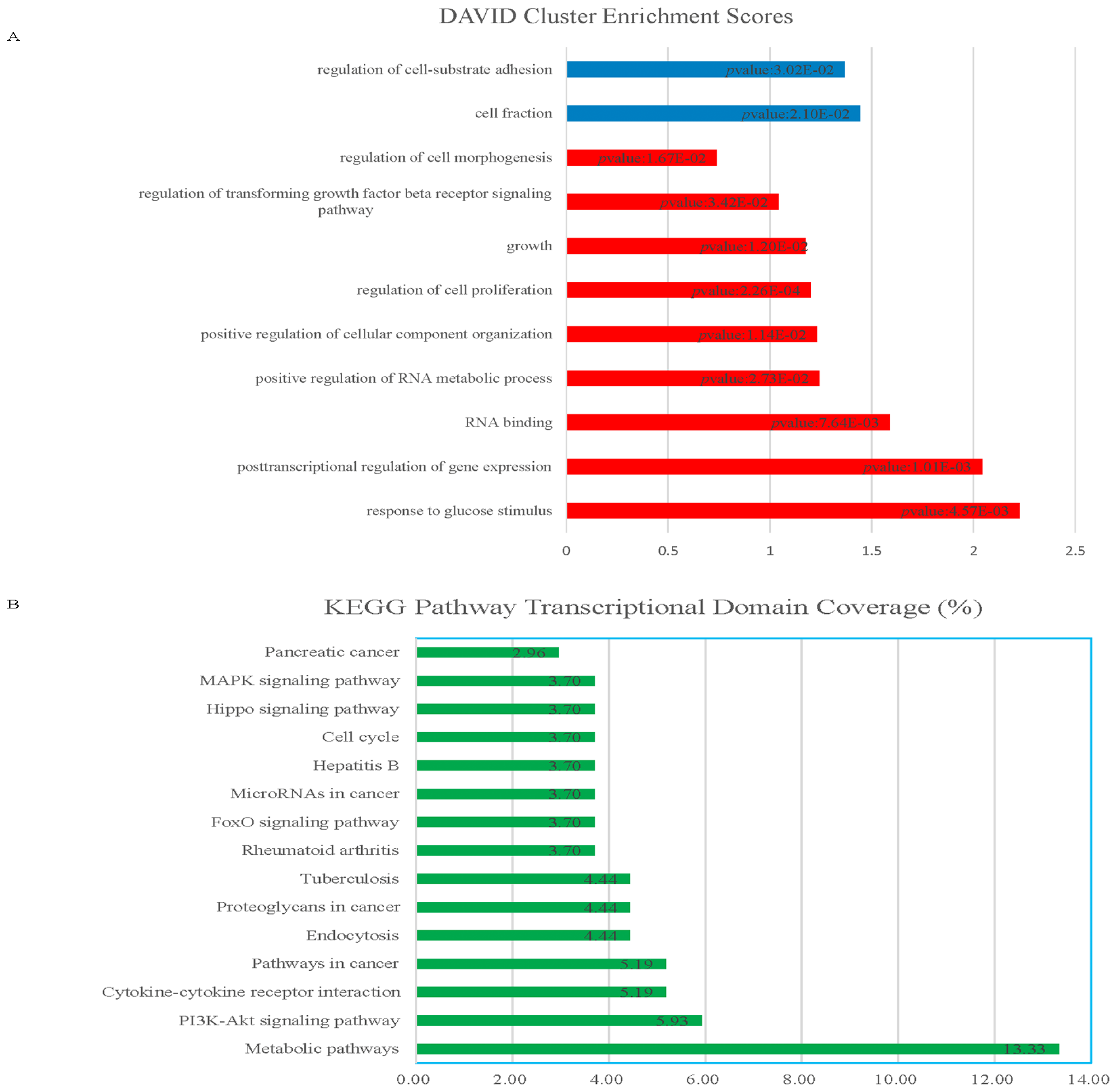
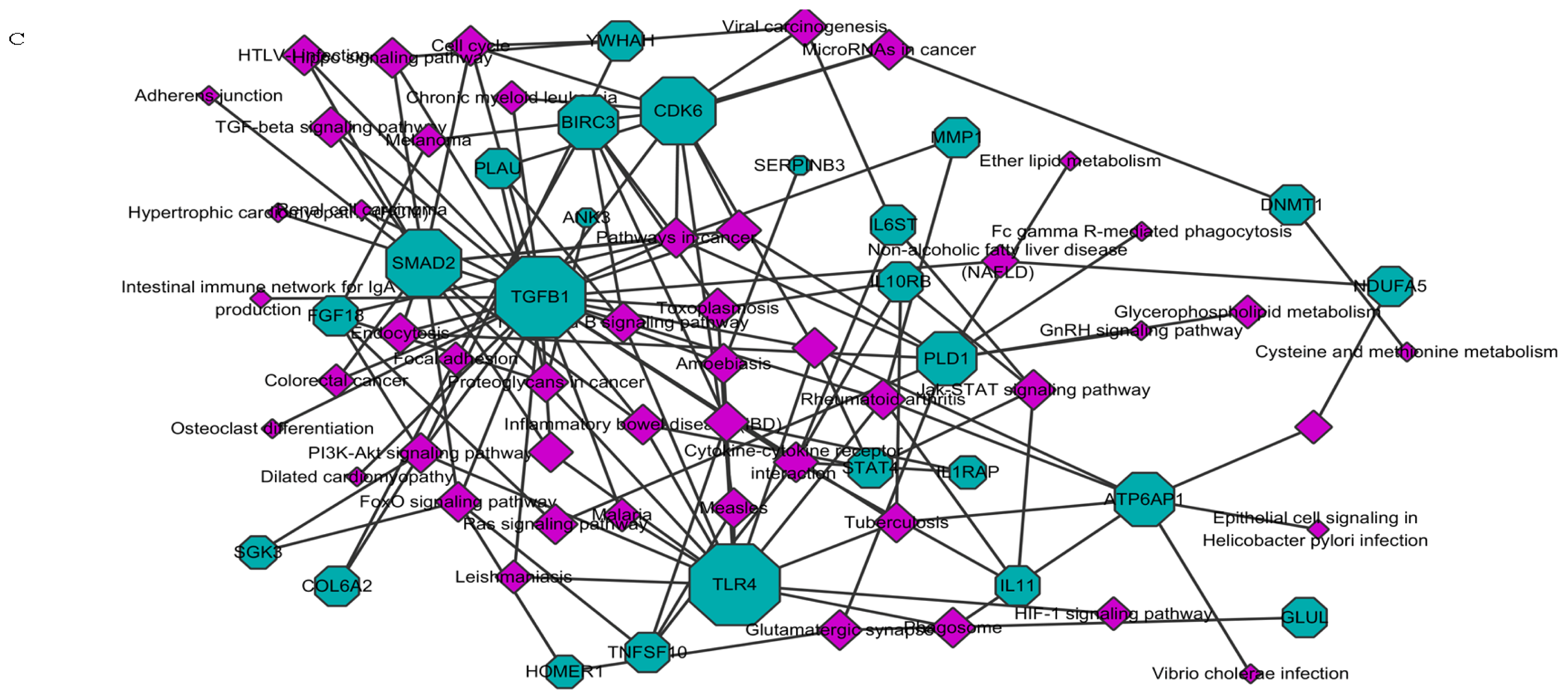
2.1.1. Knockdown of HOTAIR Altered Global Gene Expression Patterns in HCC Cells
2.1.2. Upregulation of QKI, CD82, and RBM38 Were Validated after Knockdown of HOTAIR
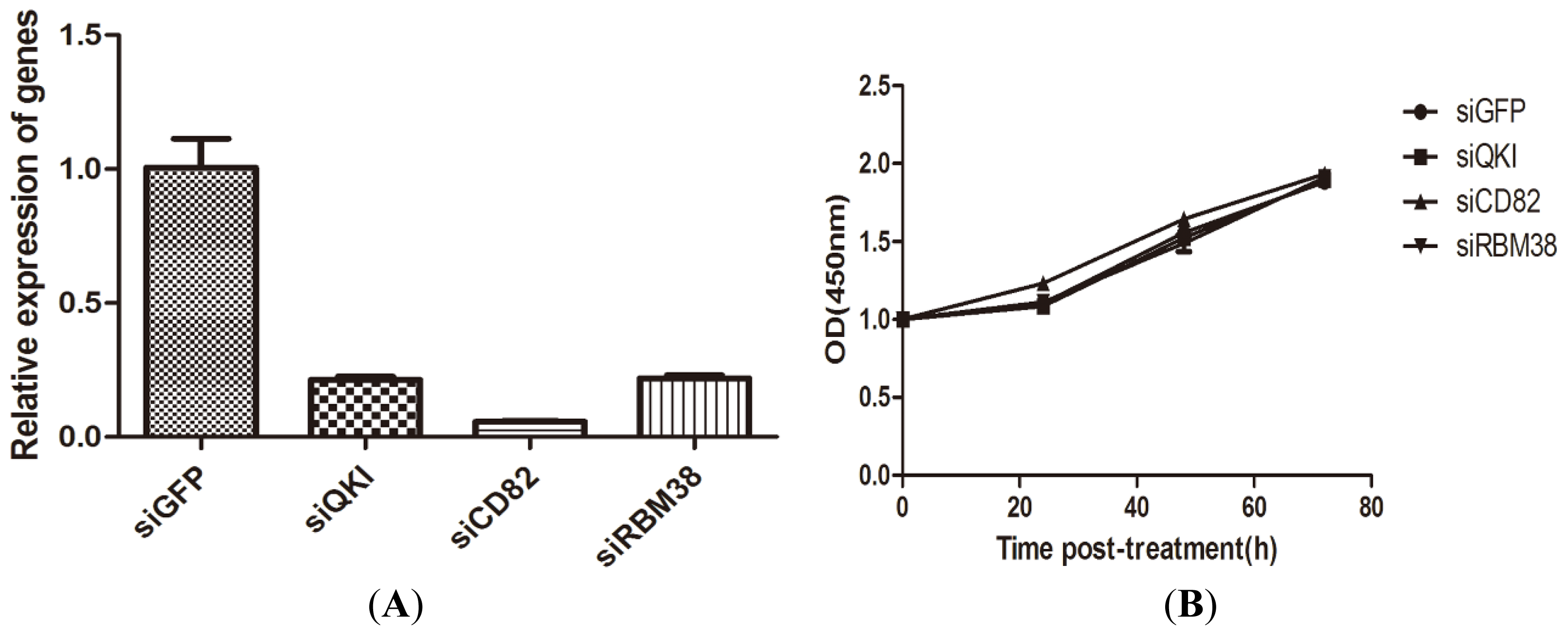
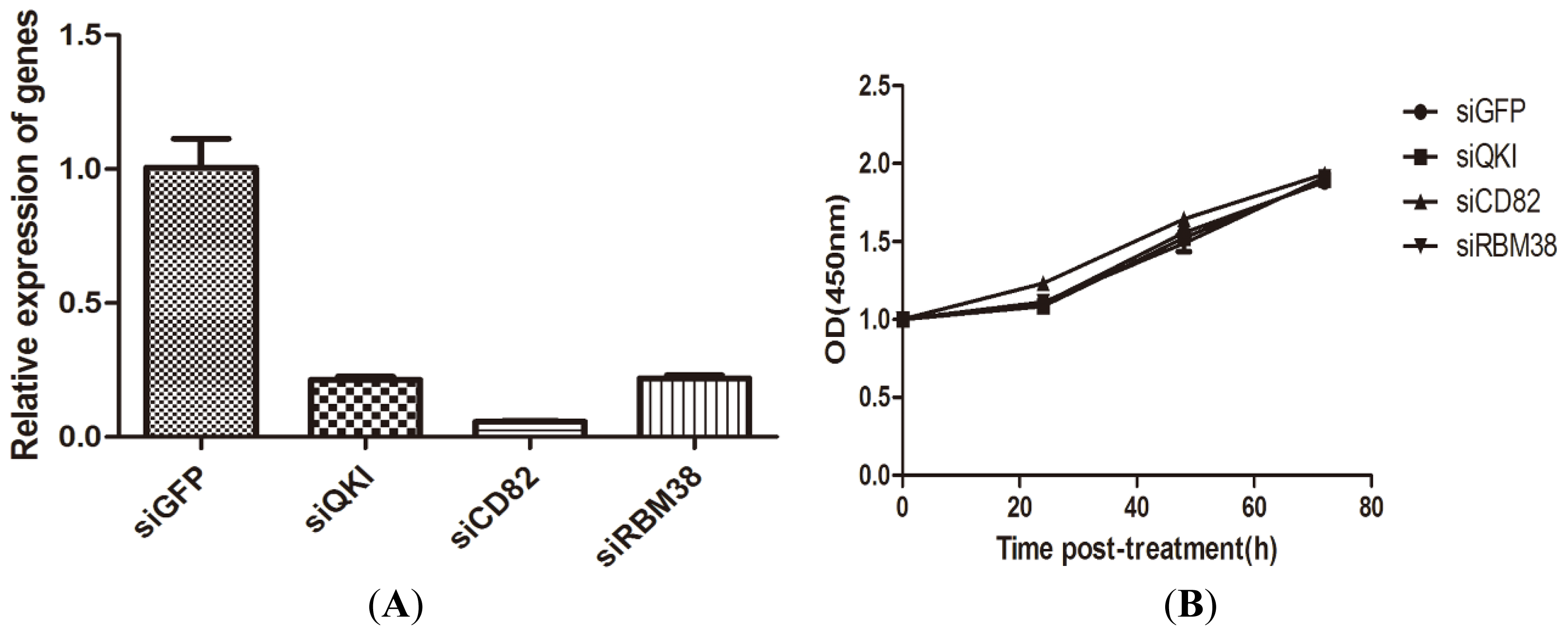
2.1.3. QKI, CD82, and RBM38 Knockdown by the Corresponding siRNAs Did not Alter Cell Proliferation
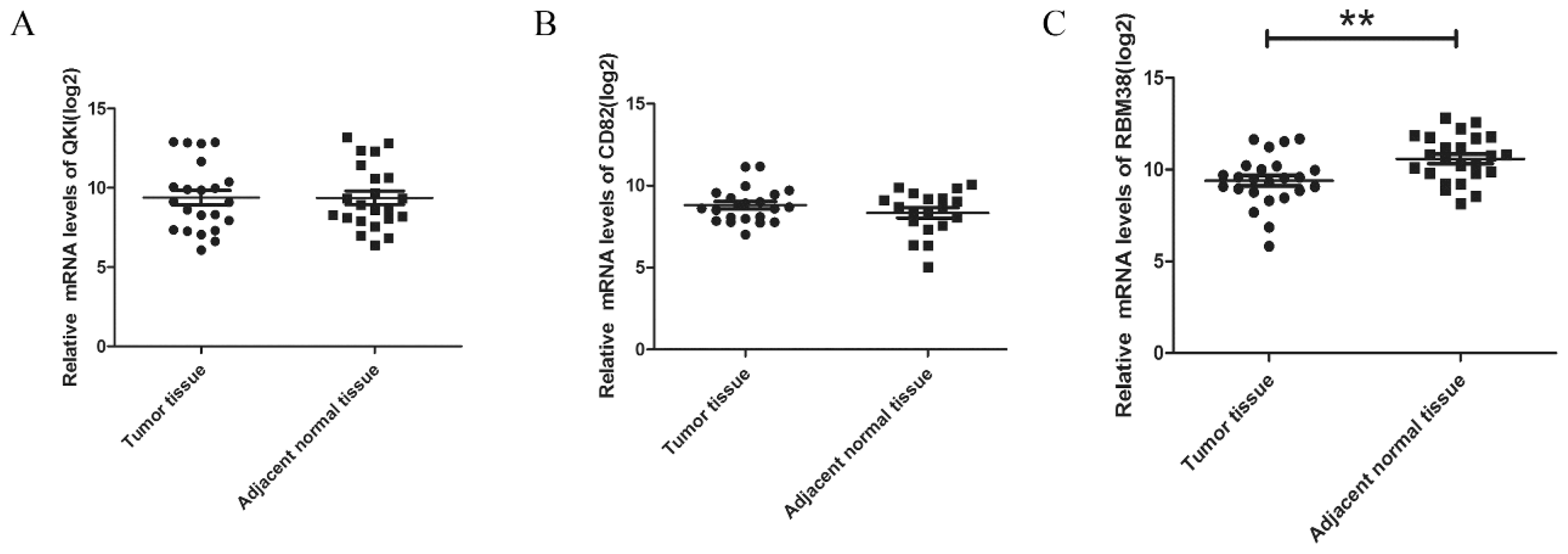
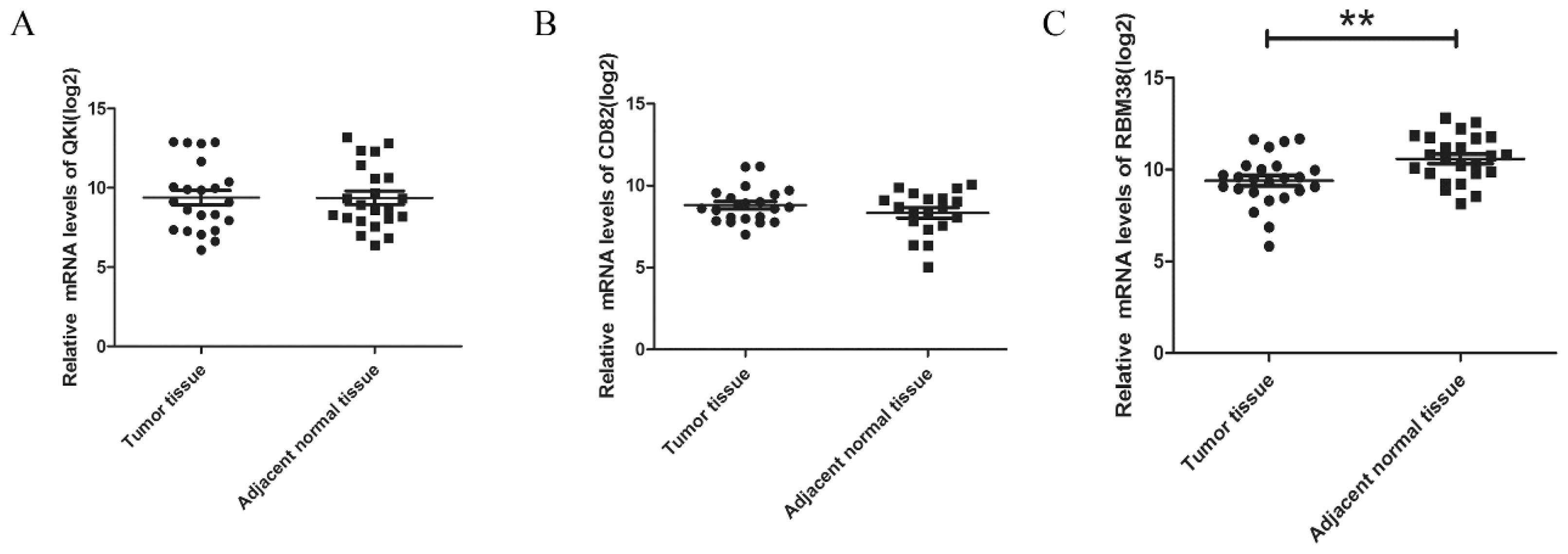
2.1.4. The Expression Levels of RBM38 Were Decreased in HCC Samples
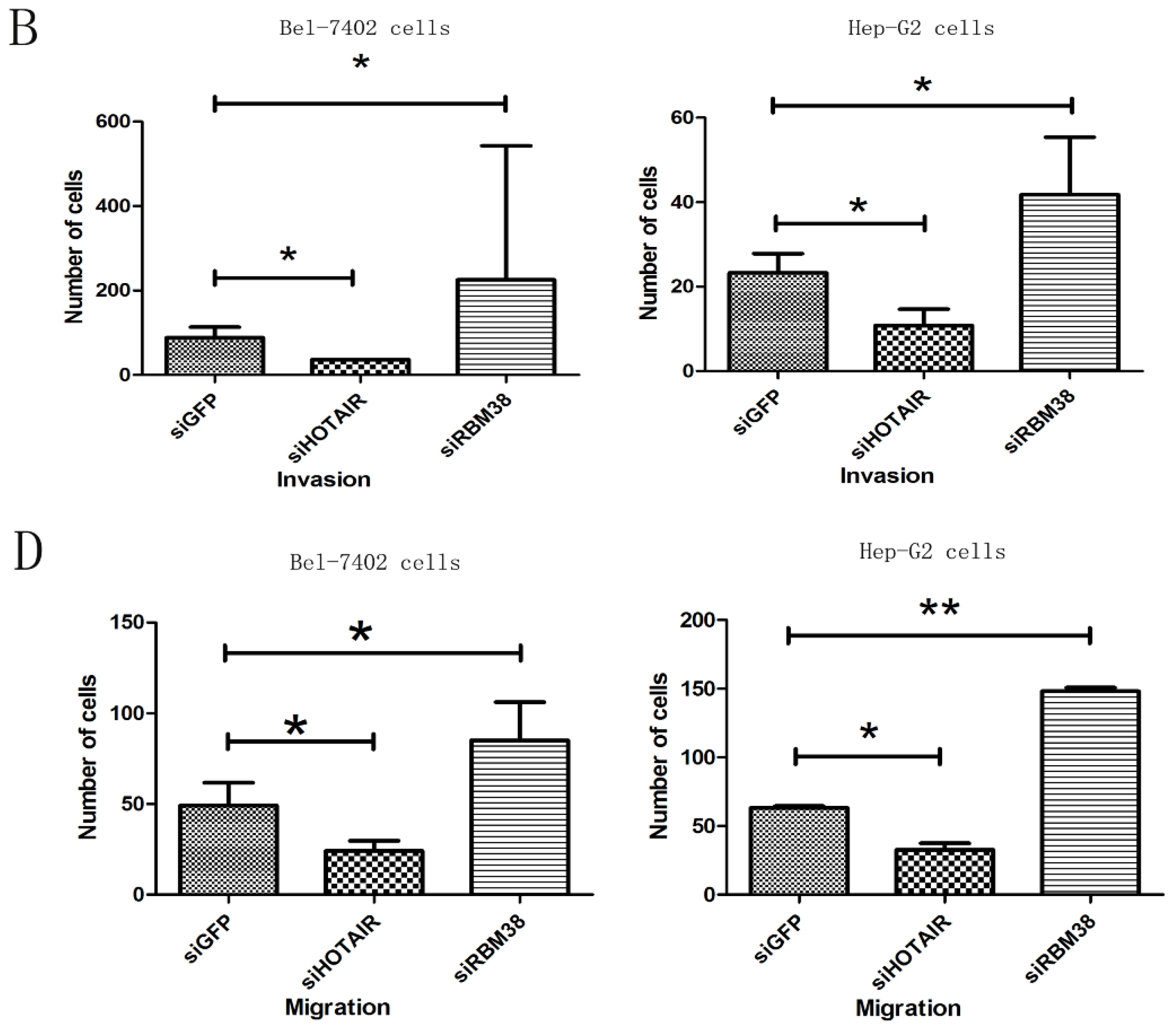
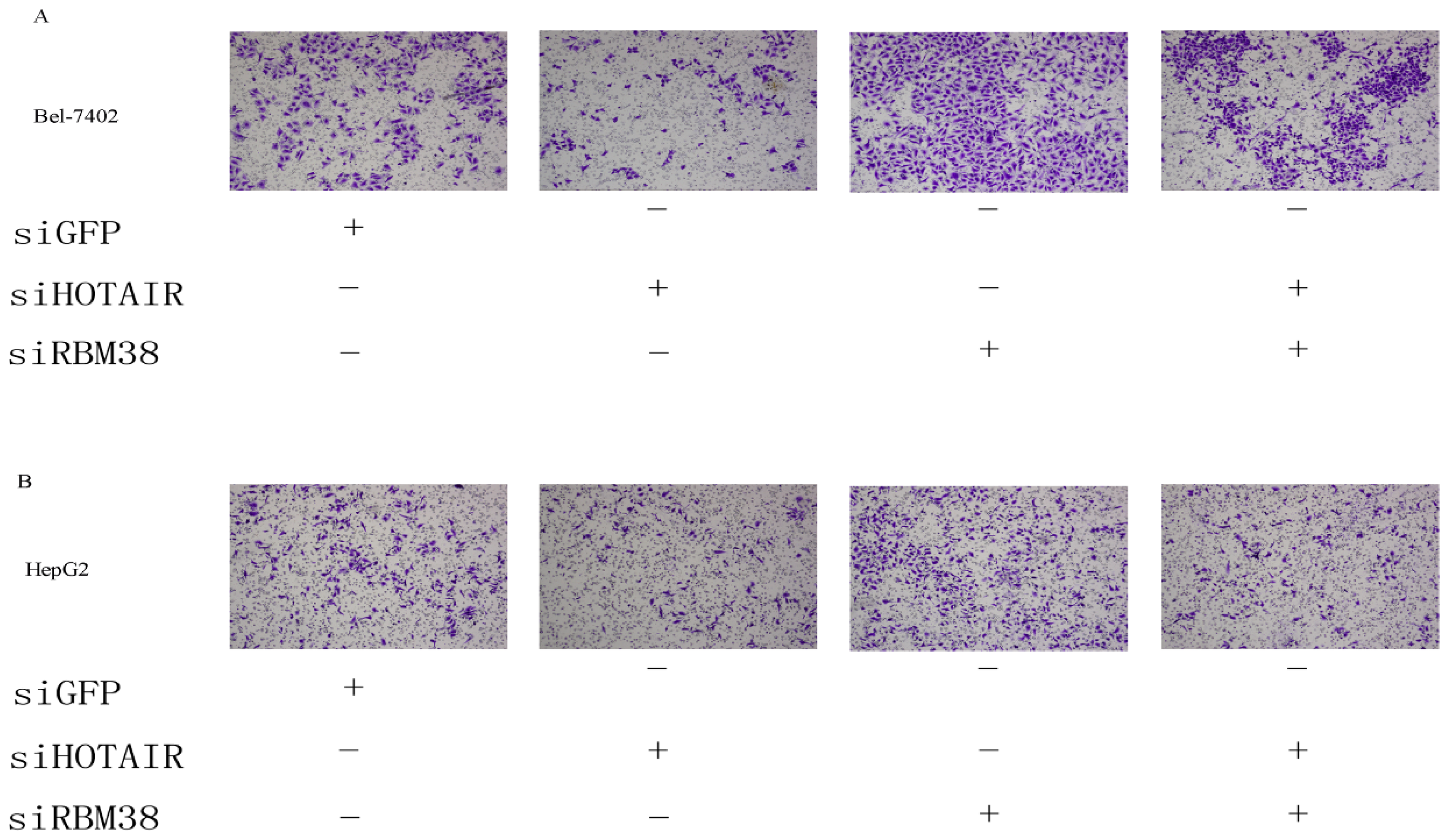
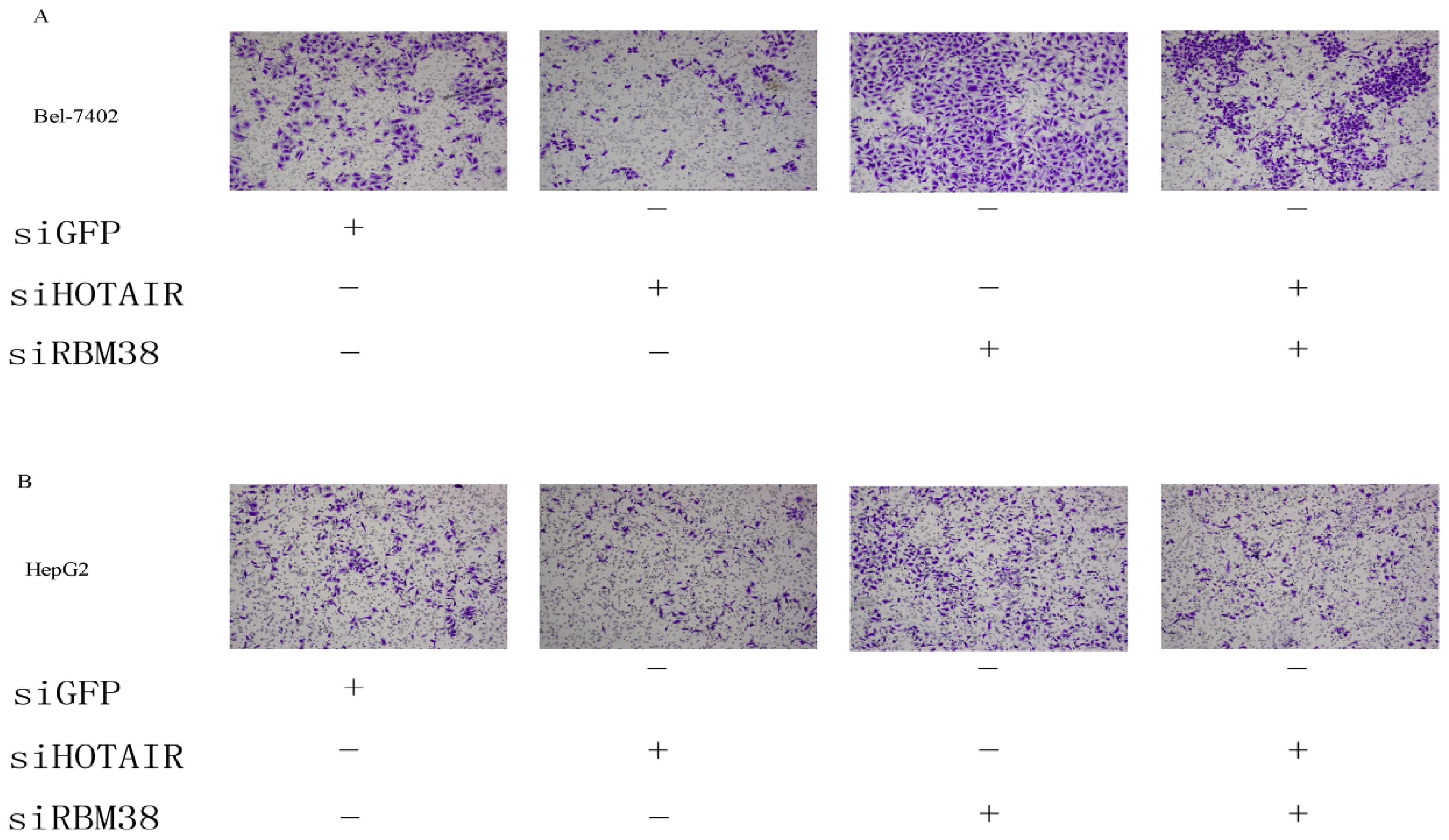
2.1.5. RBM38 Knockdown Promoted Migration and Invasion of HCC Cells
2.1.6. HOTAIR Is Functionally Related to RBM38
2.2. Discussion
3. Experimental Section
3.1. Patient Samples
3.2. Materials
3.3. Microarray Experiments and Data Analysis
3.4. RNA Isolation and Real-Time Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
- GAPDH-F: ATGGGGAAGGTGAAGGTCG
- GAPDH-R: GGGGTCATTGATGGCAACAATA
- CD82-F: CTGCAGGATGCCTGGGACTA
- CD82-R: CTCAGCGTTGTCTGTCCAGTTGTA
- QKI-F: GATGCAGCTGATGAACGACAAG
- QKI-R: CAGCATCAGGCAATTCTGCAC
- RBM38-F: GATACTGCCGCCACCAAGA
- RBM38-R: CGAGCCGGCAGACACTTTATTATAC
3.5. Small Interfering RNA Transfection
- siQKI: CCUUGAGUAUCCUAUUGAACCUAGU
- siRBM38: GACACCACGUUCACCAAGA
- siCD82: UAUUUGGUGACUUUGAUACAGGCUG
- siGFP: CUACAACAGCCACAACGUCTT
- siHOTAIR: GAACGGGAGUACAGAGAGAUU
3.6. Cell Proliferation Assay
3.7. Cell Migration and Invasion Assay
3.8. Protein Extraction and Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-15-04060-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsZheng S.S. and Wu J. conceived and designed the experiments; Ding C.F., Cheng S.B., Yang Z., Lv Z., Xiao H. performed the experiments; Ding C.F., Cheng S.B., Du C.L, Peng C.H., Xie H.Y., Zhou L. analyzed the data; Ding C.F., Cheng S.B. wrote the manuscript.
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar]
- El-Serag, H.B.; Rudolph, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar]
- Tang, Z.Y.; Ye, S.L.; Liu, Y.K.; Qin, L.X.; Sun, H.C.; Ye, Q.H.; Wang, L.; Zhou, J.; Qiu, S.J.; Li, Y.; et al. A decade’s studies on metastasis of hepatocellular carcinoma. J. Cancer Res. Clin. Oncol. 2004, 130, 187–196. [Google Scholar]
- Yang, Y.; Nagano, H.; Ota, H.; Morimoto, O.; Nakamura, M.; Wada, H.; Noda, T.; Damdinsuren, B.; Marubashi, S.; Miyamoto, A.; et al. Patterns and clinicopathologic features of extrahepatic recurrence of hepatocellular carcinoma after curative resection. Surgery 2007, 141, 196–202. [Google Scholar]
- Boyault, S.; Rickman, D.S.; de Reynies, A.; Balabaud, C.; Rebouissou, S.; Jeannot, E.; Herault, A.; Saric, J.; Belghiti, J.; Franco, D.; et al. Transcriptome classification of HCC is related to gene alterations and to new therapeutic targets. Hepatology 2007, 45, 42–52. [Google Scholar] [Green Version]
- Pogribny, I.P.; Rusyn, I. Role of epigenetic aberrations in the development and progression of human hepatocellular carcinoma. Cancer Lett. 2014, 342, 223–230. [Google Scholar]
- Huang, J.L.; Zheng, L.; Hu, Y.W.; Wang, Q. Characteristics of long noncoding RNA and its relation to hepatocellular carcinoma. Carcinogenesis 2013. [Google Scholar] [CrossRef]
- Deng, G.; Sui, G. Noncoding RNA in oncogenesis: A new era of identifying key players. Int. J. Mol. Sci. 2013, 14, 18319–18349. [Google Scholar]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics monitoring and therapeutics: A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar]
- Lee, J.T.; Bartolomei, M.S. X-inactivation imprinting and long noncoding RNAs in health and disease. Cell 2013, 152, 1308–1323. [Google Scholar]
- Gibb, E.A.; Brown, C.J.; Lam, W.L. The functional role of long non-coding RNA in human carcinomas. Mol. Cancer 2011, 10, 38:1–38:17. [Google Scholar]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar]
- Lu, L.; Zhu, G.; Zhang, C.; Deng, Q.; Katsaros, D.; Mayne, S.T.; Risch, H.A.; Mu, L.; Canuto, E.M.; Gregori, G.; et al. Association of large noncoding RNA HOTAIR expression and its downstream intergenic CpG island methylation with survival in breast cancer. Breast Cancer Res. Treat. 2012, 136, 875–883. [Google Scholar]
- Sorensen, K.P.; Thomassen, M.; Tan, Q.; Bak, M.; Cold, S.; Burton, M.; Larsen, M.J.; Kruse, T.A. Long non-coding RNA HOTAIR is an independent prognostic marker of metastasis in estrogen receptor-positive primary breast cancer. Breast Cancer Res. Treat. 2013, 142, 529–536. [Google Scholar]
- Yang, Z.; Zhou, L.; Wu, L.M.; Lai, M.C.; Xie, H.Y.; Zhang, F.; Zheng, S.S. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol. 2011, 18, 1243–1250. [Google Scholar]
- Kogo, R.; Shimamura, T.; Mimori, K.; Kawahara, K.; Imoto, S.; Sudo, T.; Tanaka, F.; Shibata, K.; Suzuki, A.; Komune, S.; et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011, 71, 6320–6326. [Google Scholar]
- Kim, K.; Jutooru, I.; Chadalapaka, G.; Johnson, G.; Frank, J.; Burghardt, R.; Kim, S.; Safe, S. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene 2013, 32, 1616–1625. [Google Scholar]
- Liu, X.H.; Liu, Z.L.; Sun, M.; Liu, J.; Wang, Z.X.; De, W. The long non-coding RNA HOTAIR indicates a poor prognosis and promotes metastasis in non-small cell lung cancer. BMC Cancer 2013, 13, 464:1–464:10. [Google Scholar]
- Ge, X.S.; Ma, H.J.; Zheng, X.H.; Ruan, H.L.; Liao, X.Y.; Xue, W.Q.; Chen, Y.B.; Zhang, Y.; Jia, W.H. HOTAIR a prognostic factor in esophageal squamous cell carcinoma inhibits WIF-1 expression and activates Wnt pathway. Cancer Sci. 2013, 104, 1675–1682. [Google Scholar]
- Li, X.; Wu, Z.; Mei, Q.; Guo, M.; Fu, X.; Han, W. Long non-coding RNA HOTAIR a driver of malignancy predicts negative prognosis and exhibits oncogenic activity in oesophageal squamous cell carcinoma. Br. J. Cancer 2013, 109, 2266–2278. [Google Scholar]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer: A long non-coding RNA point of view. RNA Biol. 2012, 9, 703–719. [Google Scholar]
- Alves, C.P.; Fonseca, A.S.; Muys, B.R.; de Barros, E.L.B.R.; Burger, M.C.; de Souza, J.E.; Valente, V.; Zago, M.A.; Silva, W.A., Jr. The lincRNA Hotair is required for epithelial-to-mesenchymal transition and stemness maintenance of cancer cells lines. Stem Cells 2013, 31, 2827–2832. [Google Scholar]
- Leveille, N.; Elkon, R.; Davalos, V.; Manoharan, V.; Hollingworth, D.; Oude Vrielink, J.; le Sage, C.; Melo, C.A.; Horlings, H.M.; Wesseling, J.; et al. Selective inhibition of microRNA accessibility by RBM38 is required for p53 activity. Nat. Commun. 2011, 2, 513:1–513:11. [Google Scholar]
- Shu, L.; Yan, W.; Chen, X. RNPC1 an RNA-binding protein and a target of the p53 family is required for maintaining the stability of the basal and stress-induced p21 transcript. Genes Dev. 2006, 20, 2961–2972. [Google Scholar]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar]
- Gumireddy, K.; Li, A.; Yan, J.; Setoyama, T.; Johannes, G.J.; Orom, U.A.; Tchou, J.; Liu, Q.; Zhang, L.; Speicher, D.W.; et al. Identification of a long non-coding RNA-associated RNP complex regulating metastasis at the translational step. EMBO J. 2013, 32, 2672–2684. [Google Scholar]
- Yoon, J.H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. LincRNA-p21 suppresses target mRNA translation. Mol. Cell 2012, 47, 648–655. [Google Scholar]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | N = 53 | RBM38 expression | χ2 | p value | |
|---|---|---|---|---|---|
| high | low | ||||
| Age (years) | |||||
| ≤50 | 21 | 6 (11.3%) | 15 (28.3%) | 0.083 | 0.773 |
| >50 | 32 | 8 (15.1%) | 24 (45.3%) | ||
| Gender | |||||
| Male | 44 | 13 (24.5%) | 31 (58.5%) | 1.306 | 0.253 |
| Female | 9 | 1 (1.9%) | 8 (15.1%) | ||
| Size of tumor (cm) | |||||
| ≤5 | 32 | 7 (13.2%) | 25 (47.2%) | 0.856 | 0.355 |
| >5 | 21 | 7 (13.2%) | 14 (26.4%) | ||
| Number of tumor | |||||
| Single | 29 | 8 (15.1%) | 21 (39.6%) | 0.045 | 0.832 |
| Multiple | 24 | 6 (11.3%) | 18 (33.9%) | ||
| Vascular invasion | |||||
| Negative | 41 | 11 (20.8%) | 30 (56.6%) | 0.016 | 0.899 |
| Positive | 12 | 3 (5.7%) | 9 (17.0%) | ||
| Pathological Grading | |||||
| Well/Moderately | 25 | 5 (9.4%) | 20 (37.7%) | 1.002 | 0.317 |
| Poorly | 28 | 9 (17.0%) | 19 (35.8%) | ||
| AFP (ng/mL) | |||||
| ≤400 | 20 | 9 (17.0%) | 11 (20.8%) | 5.708 | 0.017 |
| >400 | 33 | 5 (9.4%) | 28 (52.8%) | ||
| TBIL (μmol/L) | |||||
| ≤20 | 33 | 10 (18.9%) | 23 (43.4%) | 0.680 | 0.410 |
| >20 | 20 | 4 (7.5%) | 16 (30.2%) | ||
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ding, C.; Cheng, S.; Yang, Z.; Lv, Z.; Xiao, H.; Du, C.; Peng, C.; Xie, H.; Zhou, L.; Wu, J.; et al. Long Non-Coding RNA HOTAIR Promotes Cell Migration and Invasion via Down-Regulation of RNA Binding Motif Protein 38 in Hepatocellular Carcinoma Cells. Int. J. Mol. Sci. 2014, 15, 4060-4076. https://doi.org/10.3390/ijms15034060
Ding C, Cheng S, Yang Z, Lv Z, Xiao H, Du C, Peng C, Xie H, Zhou L, Wu J, et al. Long Non-Coding RNA HOTAIR Promotes Cell Migration and Invasion via Down-Regulation of RNA Binding Motif Protein 38 in Hepatocellular Carcinoma Cells. International Journal of Molecular Sciences. 2014; 15(3):4060-4076. https://doi.org/10.3390/ijms15034060
Chicago/Turabian StyleDing, Chaofeng, Shaobing Cheng, Zhe Yang, Zhen Lv, Heng Xiao, Chengli Du, Chuanhui Peng, Haiyang Xie, Lin Zhou, Jian Wu, and et al. 2014. "Long Non-Coding RNA HOTAIR Promotes Cell Migration and Invasion via Down-Regulation of RNA Binding Motif Protein 38 in Hepatocellular Carcinoma Cells" International Journal of Molecular Sciences 15, no. 3: 4060-4076. https://doi.org/10.3390/ijms15034060