Isolation and Characterization of the Brassinosteroid Receptor Gene (GmBRI1) from Glycine max

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation of the BRI1 Gene from Soybean
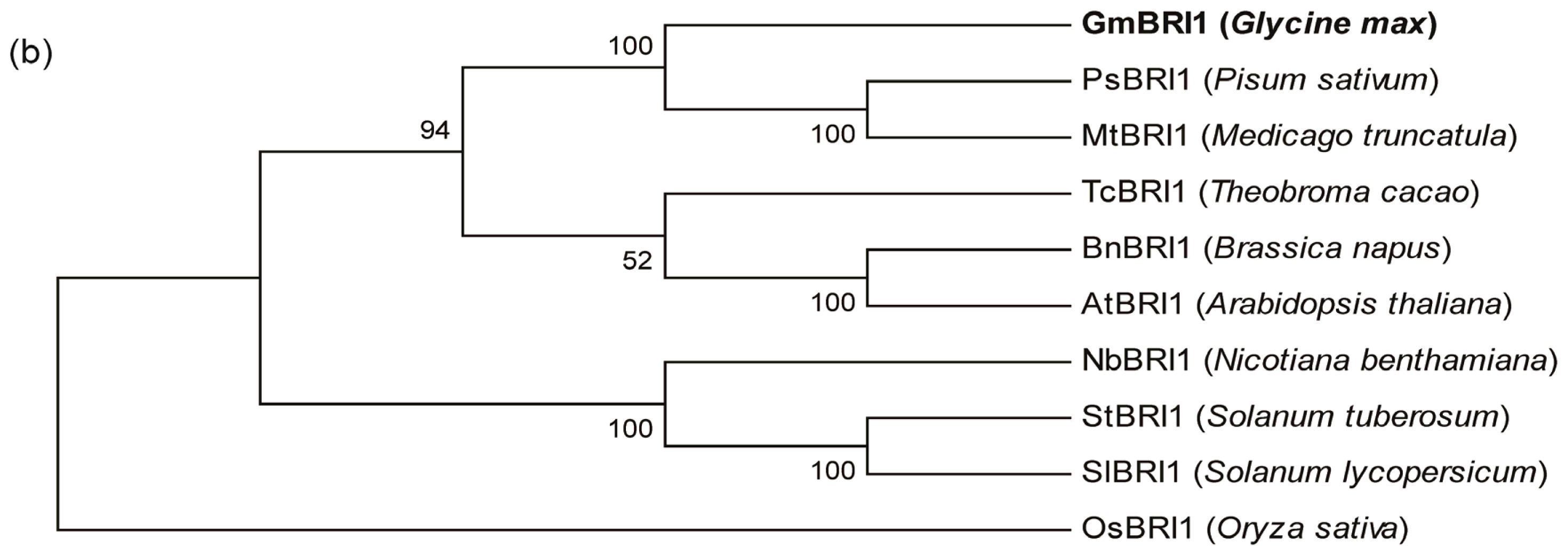
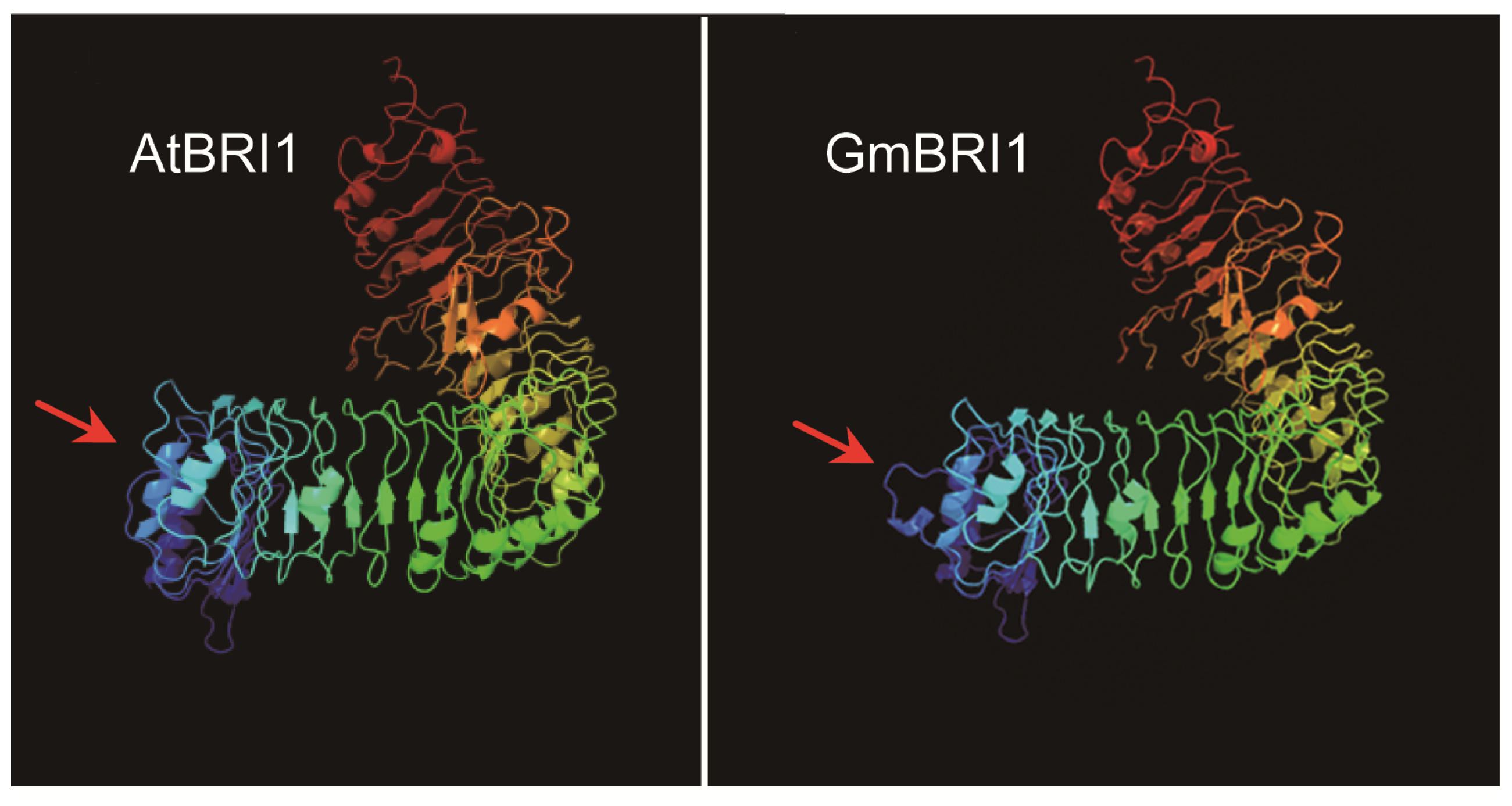
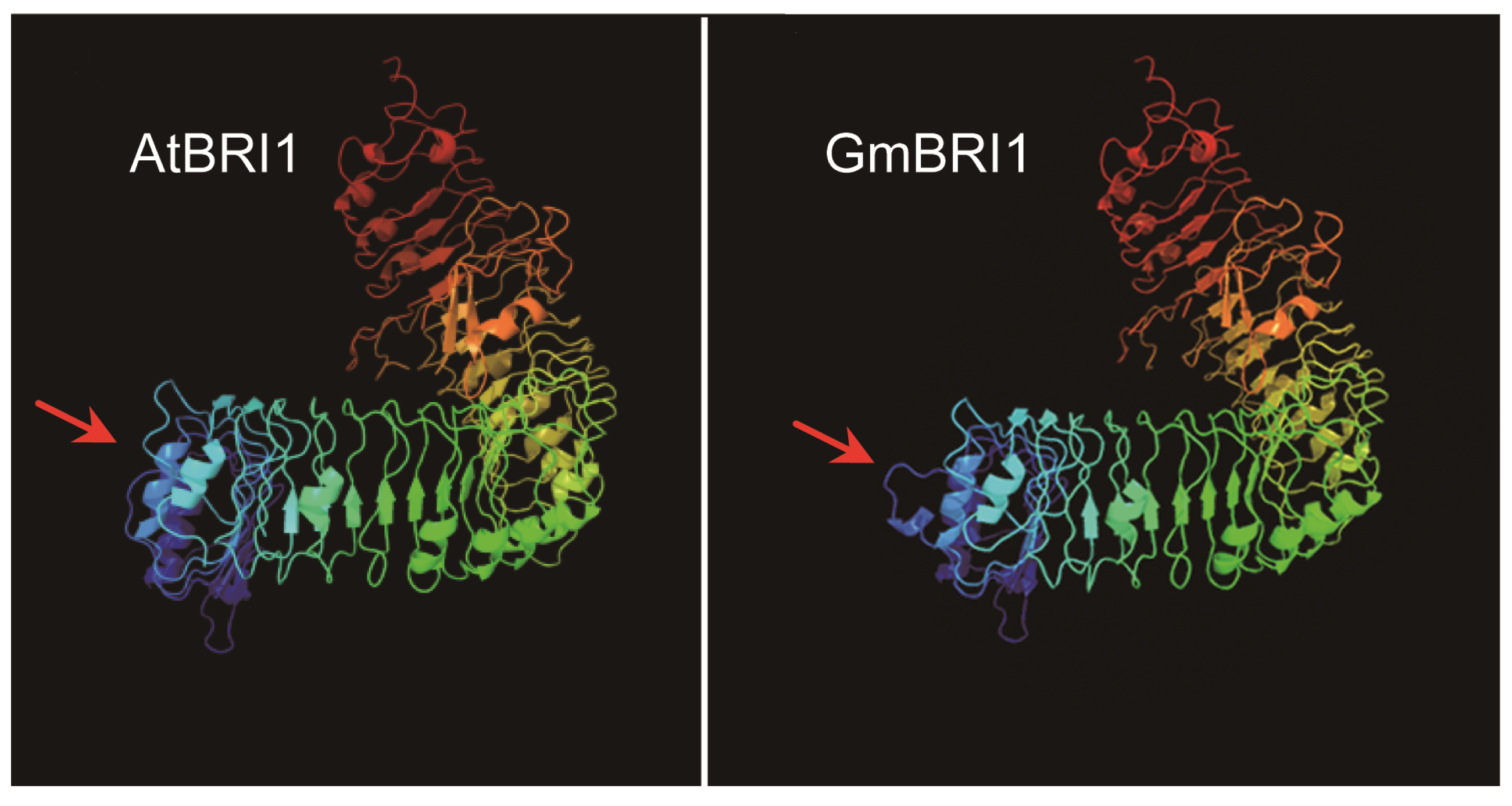
2.2. GmBRI1 Is Strongly Similar in Sequence and Structure to BRIs
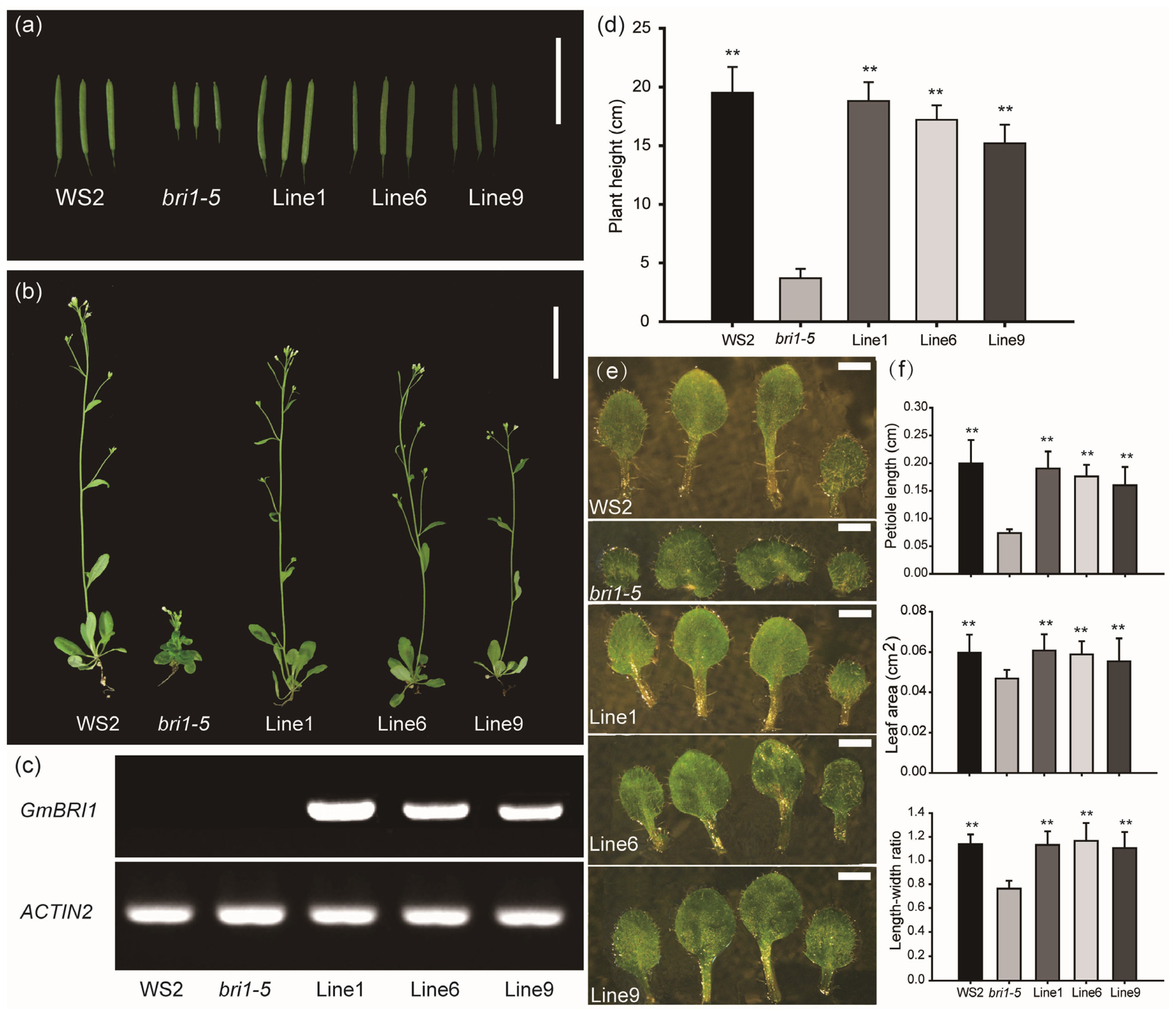
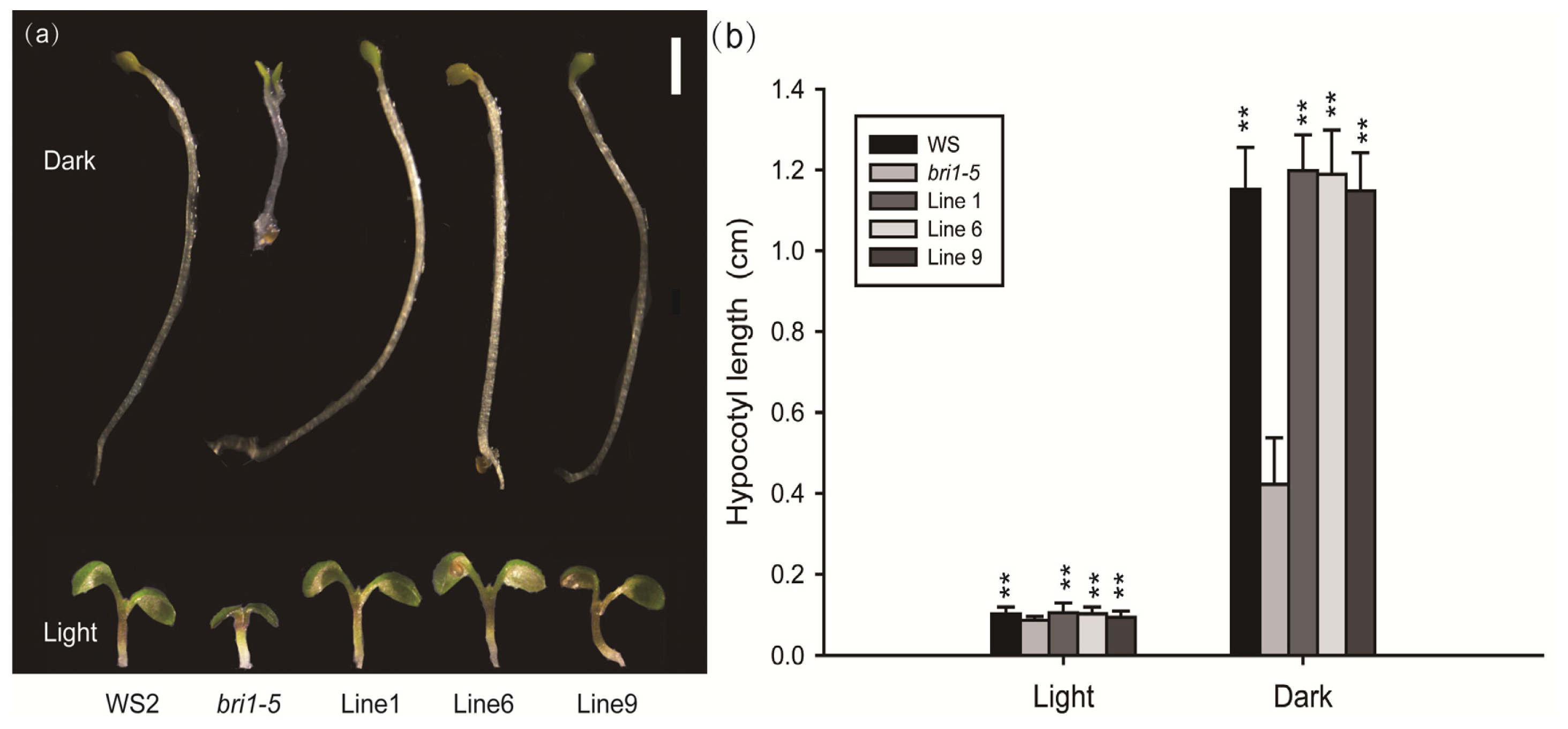
2.3. Expression of GmBRI1 Gene in the Arabidopsis Mutant Complements the Phenotype
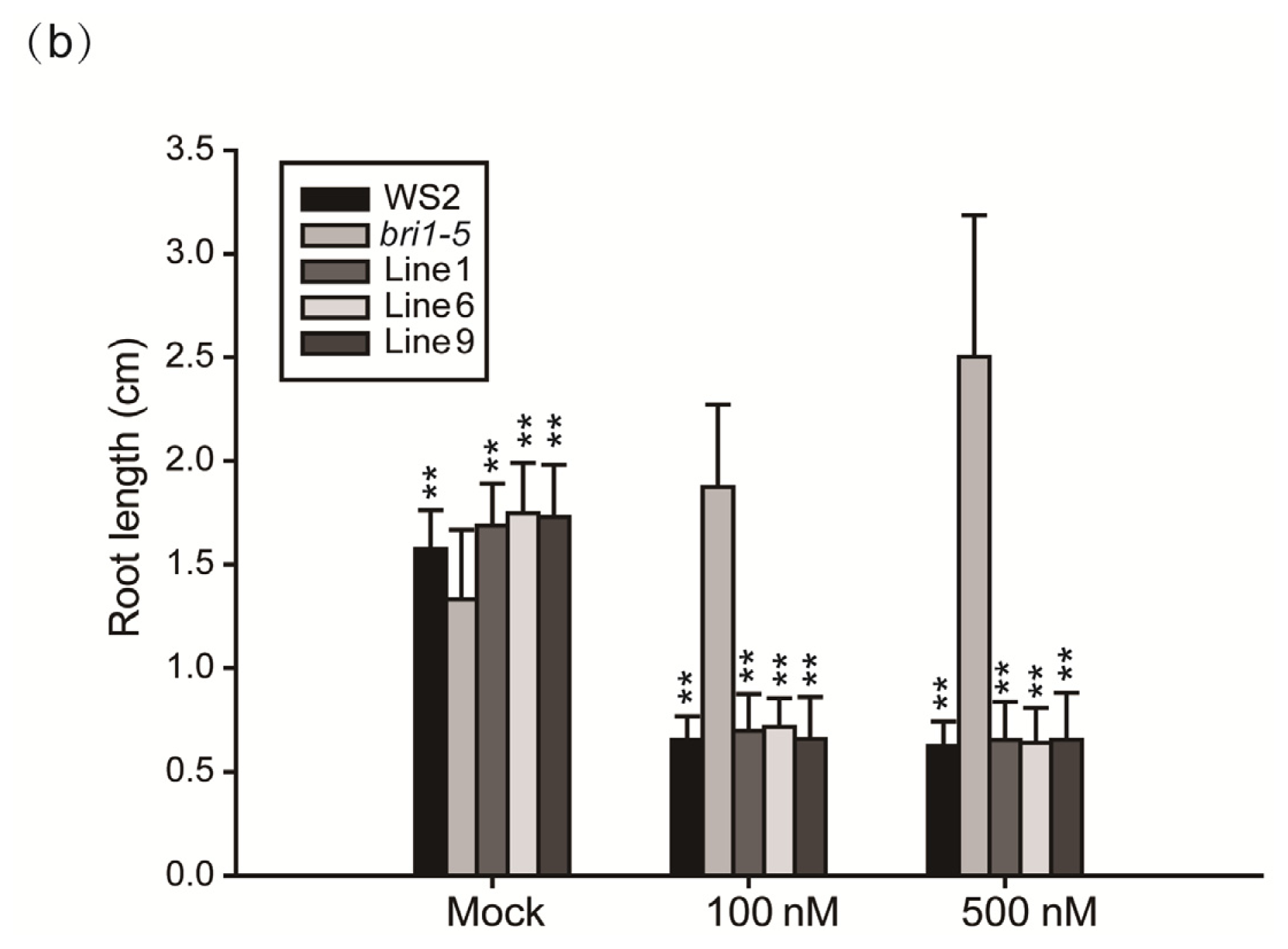
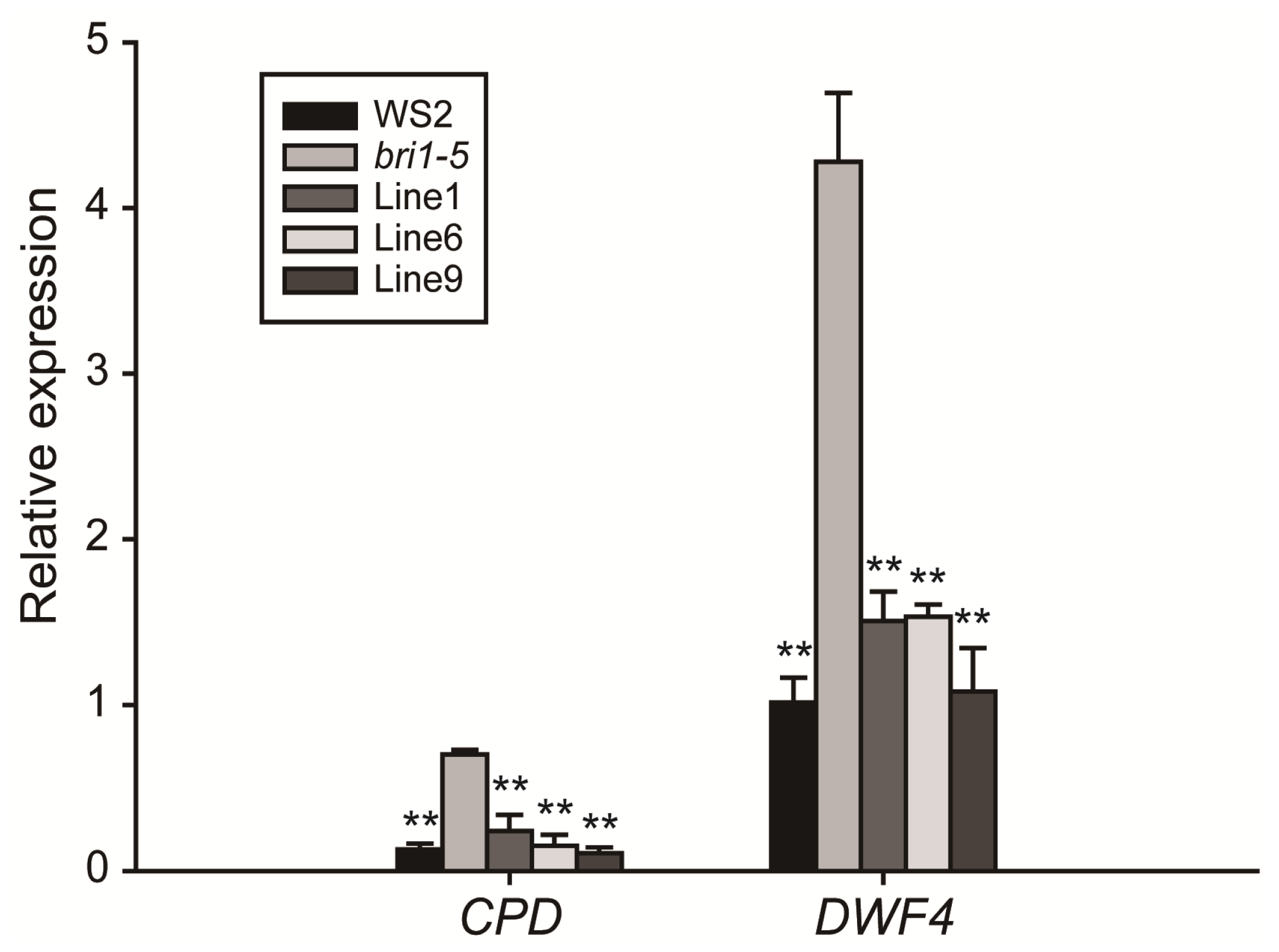
2.4. Expression of GmBRI1 Gene in the bri1-5 Restores the BR Signaling Pathway
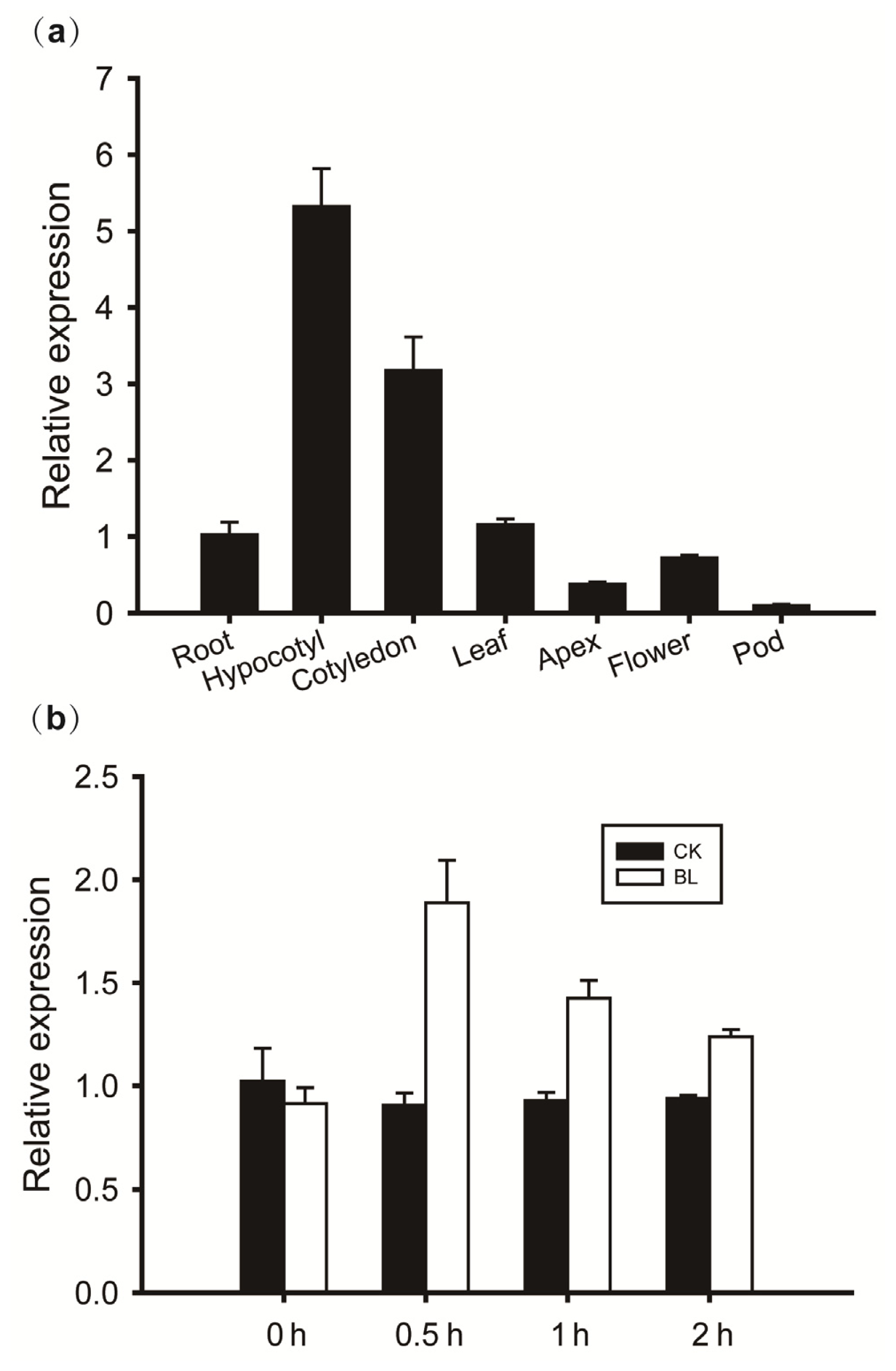
2.5. Expression of GmBRI1 in Different Soybean Organs
2.6. Transcript Levels of GmBRI1 in Response to Brassinosteroids (BRs) in Soybean Plants
3. Discussion
4. Experimental Section
4.1. Plant Material and Growth Conditions
4.2. Brassinosteroid Treatment
4.3. DNA and RNA Isolation
4.4. Sequence Analyses
4.5. Measurements and Statistical Analysis
4.6. GmBRI1 Transgene Complementation
4.7. Analysis of the GmBRI1 Gene Expression
5. Conclusions
Supplementary Information
ijms-15-03871-s001.pdfAcknowledgments
Conflicts of Interest
References
- Khush, G.S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar]
- Athwal, D.S. Semidwarf rice and wheat in global food needs. Q. Rev. Biol. 1971, 46, 1–34. [Google Scholar]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. “Green revolution” genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar]
- Hedden, P. The genes of the green revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar]
- Morinaka, Y.; Sakamoto, T.; Inukai, Y.; Agetsuma, M.; Kitano, H.; Ashikari, M.; Matsuoka, M. Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice. Plant Physiol. 2006, 141, 924–931. [Google Scholar]
- Chono, M.; Honda, I.; Zeniya, H.; Yoneyama, K.; Saisho, D.; Takeda, K.; Takatsuto, S.; Hoshino, T.; Watanabe, Y. A semidwarf phenotype of barley uzu results from a nucleotide substitution in the gene encoding a putative brassinosteroid receptor. Plant Physiol. 2003, 133, 1209–1219. [Google Scholar]
- Sasse, J.M. Physiological actions of brassinosteroids: An update. J. Plant Growth Regul. 2003, 22, 276–288. [Google Scholar]
- Krishna, P. Brassinosteroid-mediated stress responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar]
- Clouse, S.D. Molecular genetic analysis of brassinosteroid action. Physiol. Plant. 1997, 100, 702–709. [Google Scholar]
- Clouse, S.D.; Sasse, J.M. Brassinosteroids: Essential regulators of plant growth and development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 427–451. [Google Scholar]
- Yang, X.H.; Xu, Z.H.; Xue, H.W. Arabidopsis membrane steroid binding protein 1 is involved in inhibition of cell elongation. Plant Cell 2005, 17, 116–131. [Google Scholar]
- Wang, Z.Y.; Seto, H.; Fujioka, S.; Yoshida, S.; Chory, J. BRI1 is a critical component of a plasma-membrane receptor for plant steroids. Nature 2001, 410, 380–383. [Google Scholar]
- Li, J.; Chory, J. A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell 1997, 90, 929–938. [Google Scholar]
- Vert, G.; Nemhauser, J.L.; Geldner, N.; Hong, F.; Chory, J. Molecular mechanisms of steroid hormone signaling in plants. Annu. Rev. Cell Dev. Biol. 2005, 21, 177–201. [Google Scholar]
- Gruszka, D. The brassinosteroid signaling pathway-new key players and interconnections with other signaling networks crucial for plant development and stress tolerance. Int. J. Mol. Sci. 2013, 14, 8740–8774. [Google Scholar]
- Hothorn, M.; Belkhadir, Y.; Dreux, M.; Dabi, T.; Noel, J.P.; Wilson, I.A.; Chory, J. Structural basis of steroid hormone perception by the receptor kinase BRI1. Nature 2011, 474, 467–471. [Google Scholar]
- Kinoshita, T.; Cano-Delgado, A.C.; Seto, H.; Hiranuma, S.; Fujioka, S.; Yoshida, S.; Chory, J. Binding of brassinosteroids to the extracellular domain of plant receptor kinase BRI1. Nature 2005, 433, 167–171. [Google Scholar]
- She, J.; Han, Z.F.; Kim, T.W.; Wang, J.J.; Cheng, W.; Chang, J.B.; Shi, S.A.; Wang, J.W.; Yang, M.J.; Wang, Z.Y.; et al. Structural insight into brassinosteroid perception by BRI1. Nature 2011, 474, 472–476. [Google Scholar]
- Yang, C.J.; Zhang, C.; Lu, Y.N.; Jin, J.Q.; Wang, X.L. The mechanisms of brassinosteroids’ action: From signal transduction to plant development. Mol. Plant 2011, 4, 588–600. [Google Scholar]
- Nam, K.H.; Li, J.M. BRI1/BAK1 a receptor kinase pair mediating brassinosteroid signaling. Cell 2002, 110, 203–212. [Google Scholar]
- Wang, X.L.; Chory, J. Brassinosteroids regulate dissociation of BKI1 a negative regulator of BRI1 signaling from the plasma membrane. Science 2006, 313, 1118–1122. [Google Scholar]
- He, J.X.; Gendron, J.M.; Sun, Y.; Gampala, S.S.L.; Gendron, N.; Sun, C.Q.; Wang, Z.Y. BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 2005, 307, 1634–1638. [Google Scholar]
- Yin, Y.H.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar]
- Nomura, T.; Nakayama, M.; Reid, J.B.; Takeuchi, Y.; Yokota, T. Blockage of brassinosteroid biosynthesis and sensitivity causes dwarfism in garden pea. Plant Physiol. 1997, 113, 31–37. [Google Scholar]
- Nomura, T.; Kitasaka, Y.; Takatsuto, S.; Reid, J.B.; Fukami, M.; Yokota, T. Brassinosteroid/sterol synthesis and plant growth as affected by ika and ikb mutations of pea. Plant Physiol. 1999, 119, 1517–1526. [Google Scholar]
- Koka, C.V.; Cerny, R.E.; Gardner, R.G.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Clouse, S.D. A putative role for the tomato genes dumpy and curl-3 in brassinosteroid biosynthesis and response. Plant Physiol. 2000, 122, 85–98. [Google Scholar]
- Xie, F.; Dong, Z.; Wang, X.; Sun, Y. The effects of lodging on soybean yeild formation. Soybean Sci. 1993, 12, 81–85. [Google Scholar]
- Wang, L.; Sun, J.; Zhao, R.; Wang, L.Z.; Luo, G.; Li, B. Advances in soybean breeding for super high-yeilding. Soybean Sci. 2013, 32, 687–693. [Google Scholar]
- Hu, P.; Han, T. The molecular basis of stem development in plant. Chin. Bull. Bot. 2008, 25, 1–13. [Google Scholar]
- Noguchi, T.; Fujioka, S.; Choe, S.; Takatsuto, S.; Yoshida, S.; Yuan, H.; Feldmann, K.A.; Tax, F.E. Brassinosteroid-insensitive dwarf mutants of Arabidopsis accumulate brassinosteroids. Plant Physiol. 1999, 121, 743–752. [Google Scholar]
- Gonzalez-Garcia, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-Garcia, S.; Russinova, E.; Cano-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar]
- Clouse, S.D.; Langford, M.; McMorris, T.C. A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol. 1996, 111, 671–678. [Google Scholar]
- Mussig, C.; Shin, G.H.; Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271. [Google Scholar]
- Lin, L.L.; Wu, C.C.; Huang, H.C.; Chen, H.J.; Hsieh, H.L.; Juan, H.F. Identification of microrna 395a in 24-epibrassinolide-regulated root growth of Arabidopsis thaliana using microrna arrays. Int. J. Mol. Sci. 2013, 14, 14270–14286. [Google Scholar]
- Kauschmann, A.; Jessop, A.; Koncz, C.; Szekeres, M.; Willmitzer, L.; Altmann, T. Genetic evidence for an essential role of brassinosteroids in plant development. Plant J. 1996, 9, 701–713. [Google Scholar]
- Szekeres, M.; Nemeth, K.; KonczKalman, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Redei, G.P.; Nagy, F.; Schell, J.; Koncz, C. Brassinosteroids rescue the deficiency of cyp90 a cytochrome p450 controlling cell elongation and de-etiolation in arabidopsis. Cell 1996, 85, 171–182. [Google Scholar]
- Mathur, J.; Molnar, G.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Yokota, T.; Adam, G.; Voigt, B.; Nagy, F.; Maas, C.; et al. Transcription of the Arabidopsis cpd gene encoding a steroidogenic cytochrome p450 is negatively controlled by brassinosteroids. Plant J. 1998, 14, 593–602. [Google Scholar]
- Kim, H.B.; Kwon, M.; Ryu, H.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; An, C.S.; Lee, I.; Hwang, I.; Choe, S. The regulation of dwarf4 expression is likely a critical mechanism in maintaining the homeostasis of bioactive brassinosteroids in Arabidopsis. Plant Physiol. 2006, 140, 548–557. [Google Scholar]
- Grove, M.D.; Spencer, G.F.; Rohwedder, W.K.; Mandava, N.; Worley, J.F.; Warthen, J.D.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C. Brassinolide a plant growth-promoting steroid isolated from brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar]
- Zurek, D.M.; Clouse, S.D. Molecular cloning and characterization of a brassinosteroid-regulated gene from elongating soybean (Glycine max l) epicotyls. Plant Physiol. 1994, 104, 161–170. [Google Scholar]
- Zurek, D.M.; Rayle, D.L.; McMorris, T.C.; Clouse, S.D. Investigation of gene expression growth kinetics and wall extensibility during brassinosteroid-regulated stem elongation. Plant Physiol. 1994, 104, 505–513. [Google Scholar]
- Shi, J.; Gonzales, R.A.; Bhattacharyya, M.K. Identification and characterization of an S-adenosyl-l-methionine: δ 24-sterol-C-methyltransferase cDNA from soybean. J. Biol. Chem. 1996, 271, 9384–9389. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar]






© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, M.; Sun, S.; Wu, C.; Han, T.; Wang, Q. Isolation and Characterization of the Brassinosteroid Receptor Gene (GmBRI1) from Glycine max. Int. J. Mol. Sci. 2014, 15, 3871-3888. https://doi.org/10.3390/ijms15033871
Wang M, Sun S, Wu C, Han T, Wang Q. Isolation and Characterization of the Brassinosteroid Receptor Gene (GmBRI1) from Glycine max. International Journal of Molecular Sciences. 2014; 15(3):3871-3888. https://doi.org/10.3390/ijms15033871
Chicago/Turabian StyleWang, Miao, Shi Sun, Cunxiang Wu, Tianfu Han, and Qingyu Wang. 2014. "Isolation and Characterization of the Brassinosteroid Receptor Gene (GmBRI1) from Glycine max" International Journal of Molecular Sciences 15, no. 3: 3871-3888. https://doi.org/10.3390/ijms15033871