A Proteomic Approach of Bradyrhizobium/Aeschynomene Root and Stem Symbioses Reveals the Importance of the fixA Locus for Symbiosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
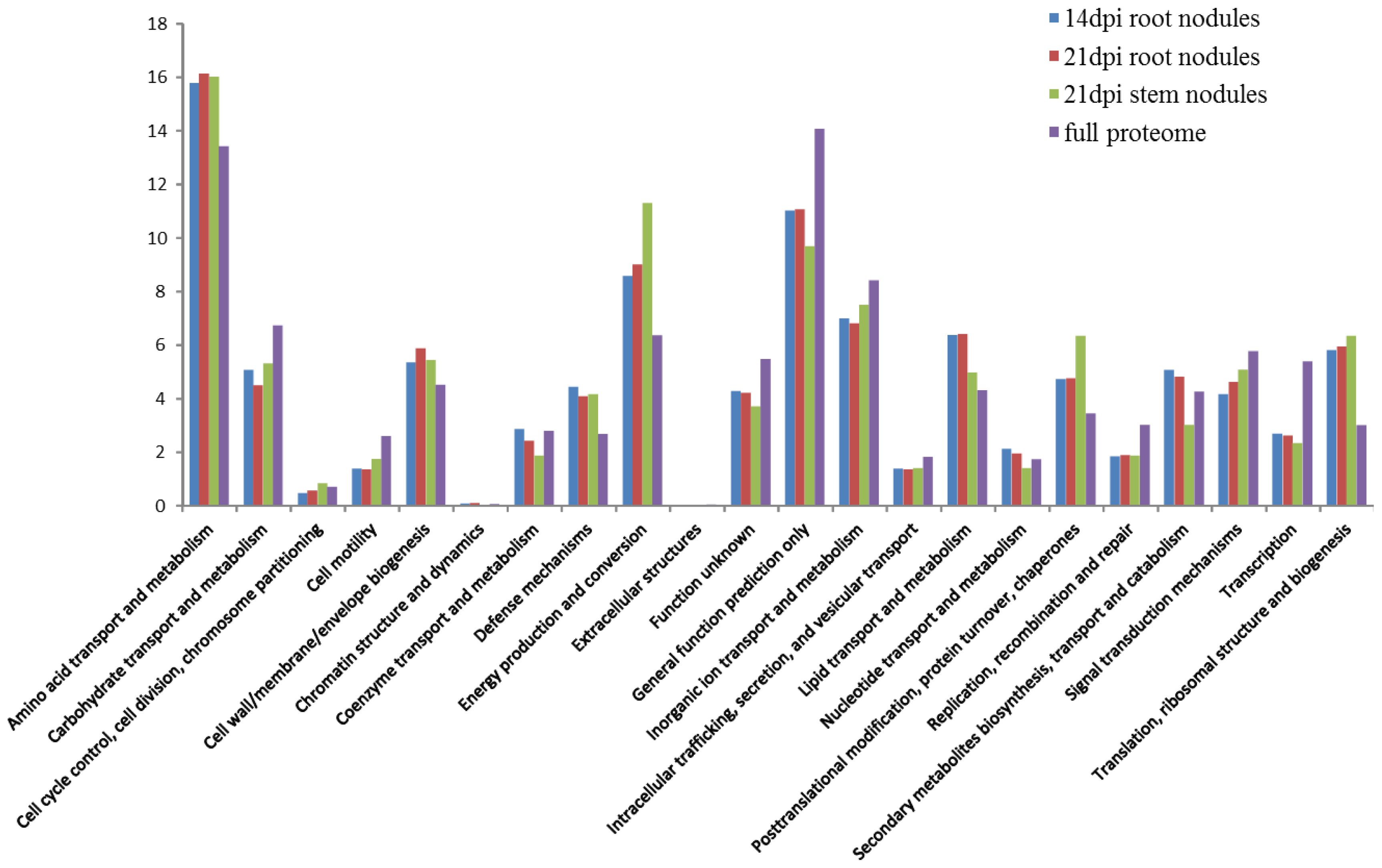
2. Results and Discussion
2.1. Bradyrhizobium sp. ORS278 Samples Overview
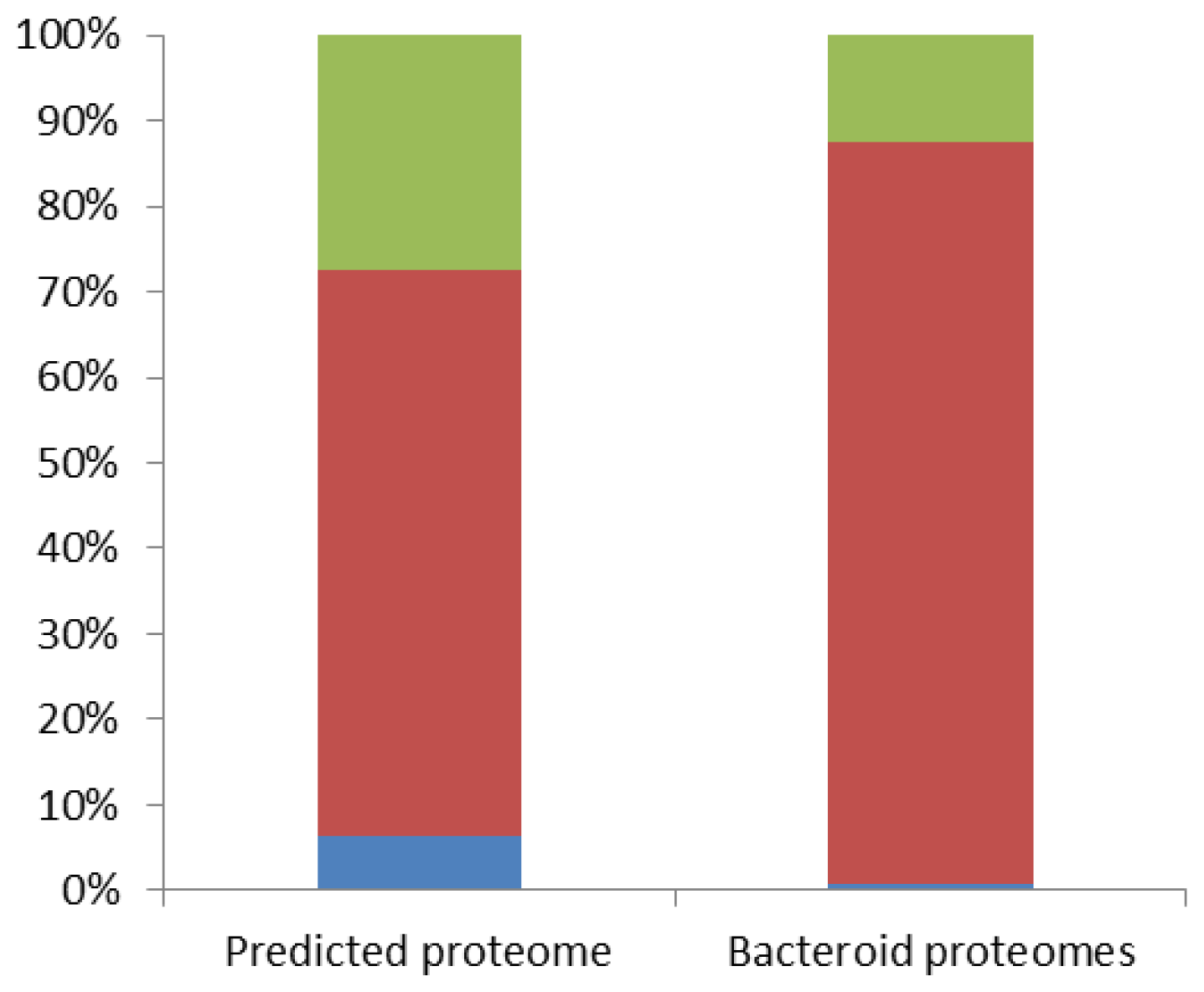
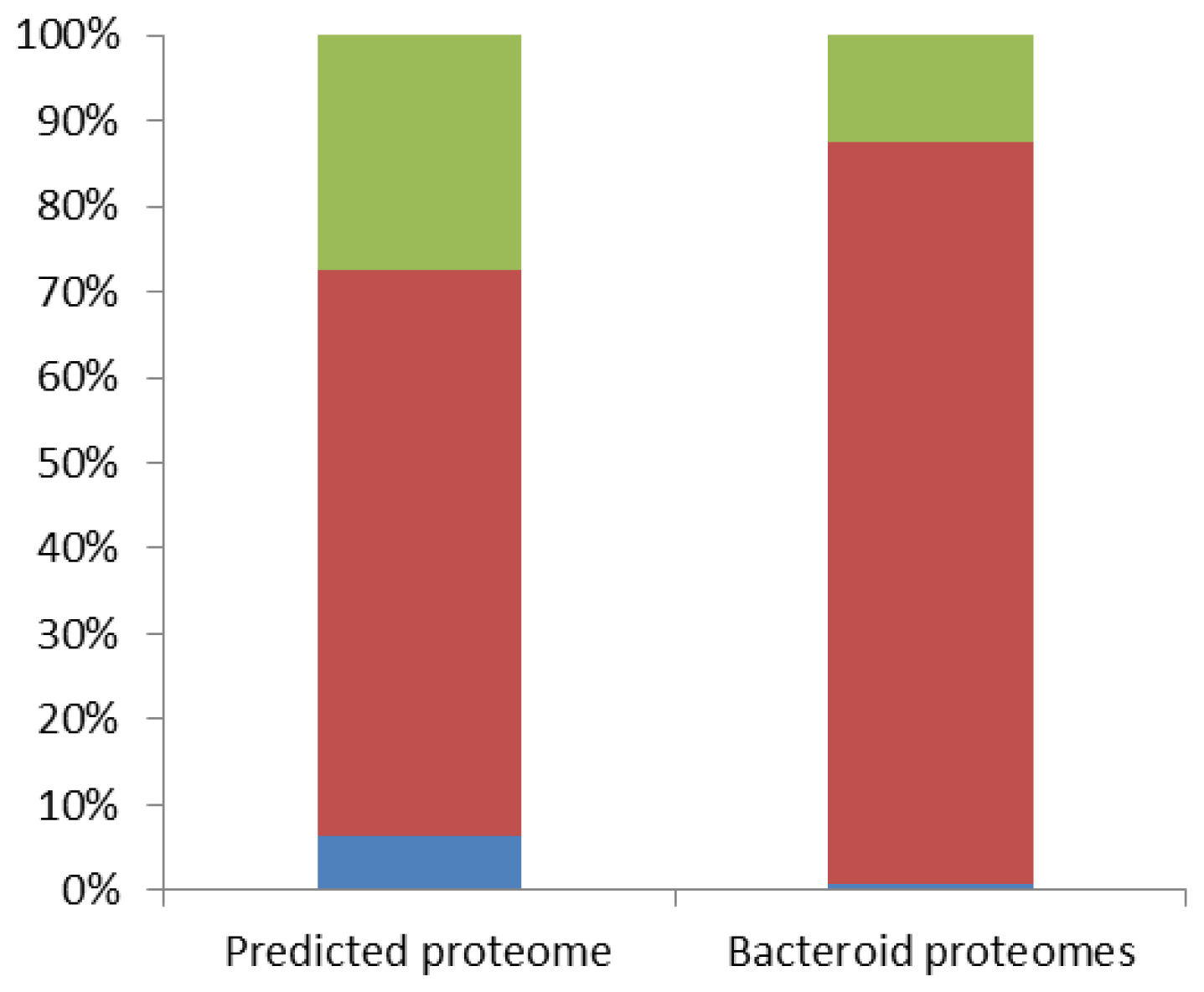
2.2. Bacteroid Proteomes Are Essentially Encoded by the Bradyrhizobium Core Genome
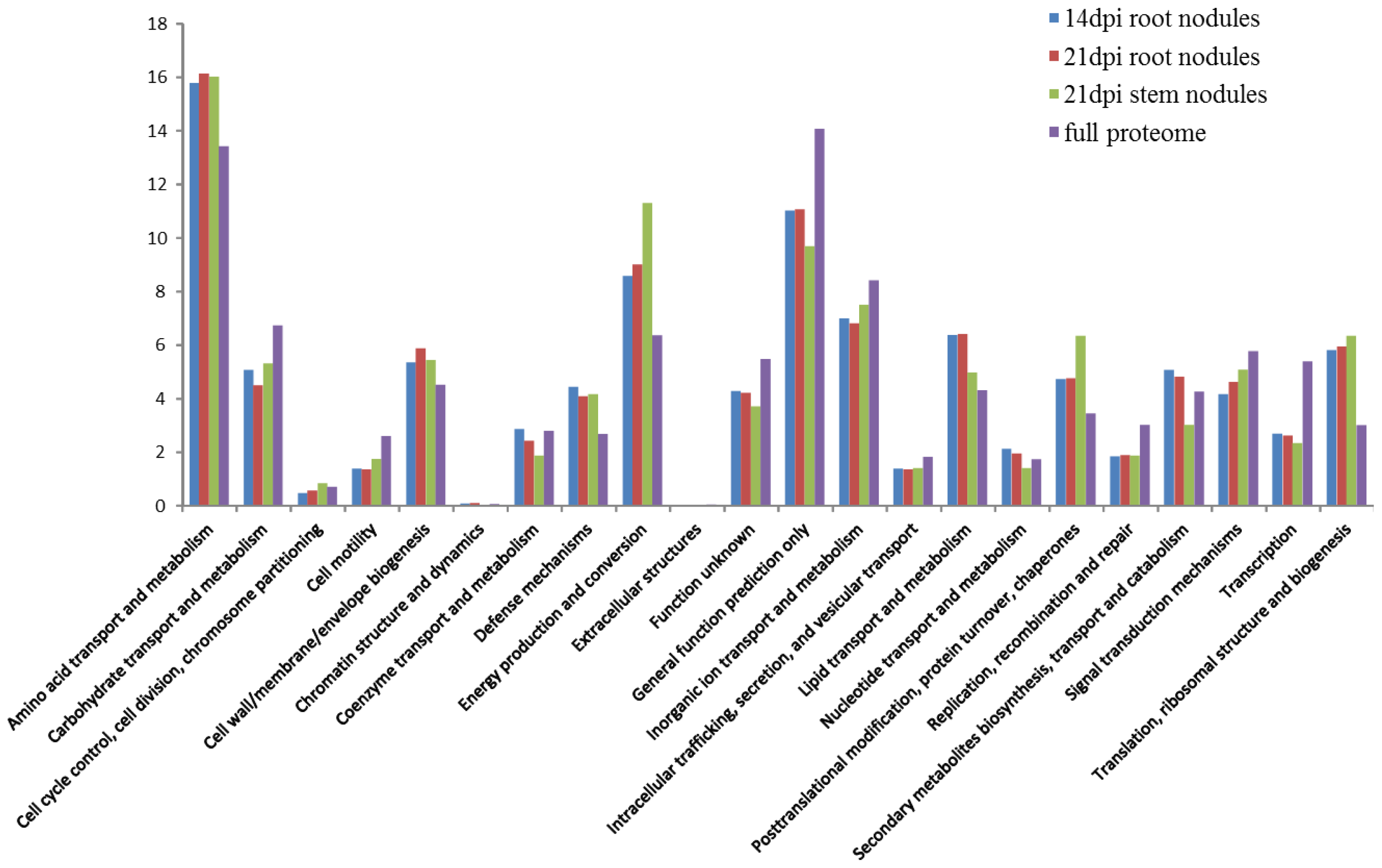
2.3. Functional Distribution of the Symbiotic Proteins
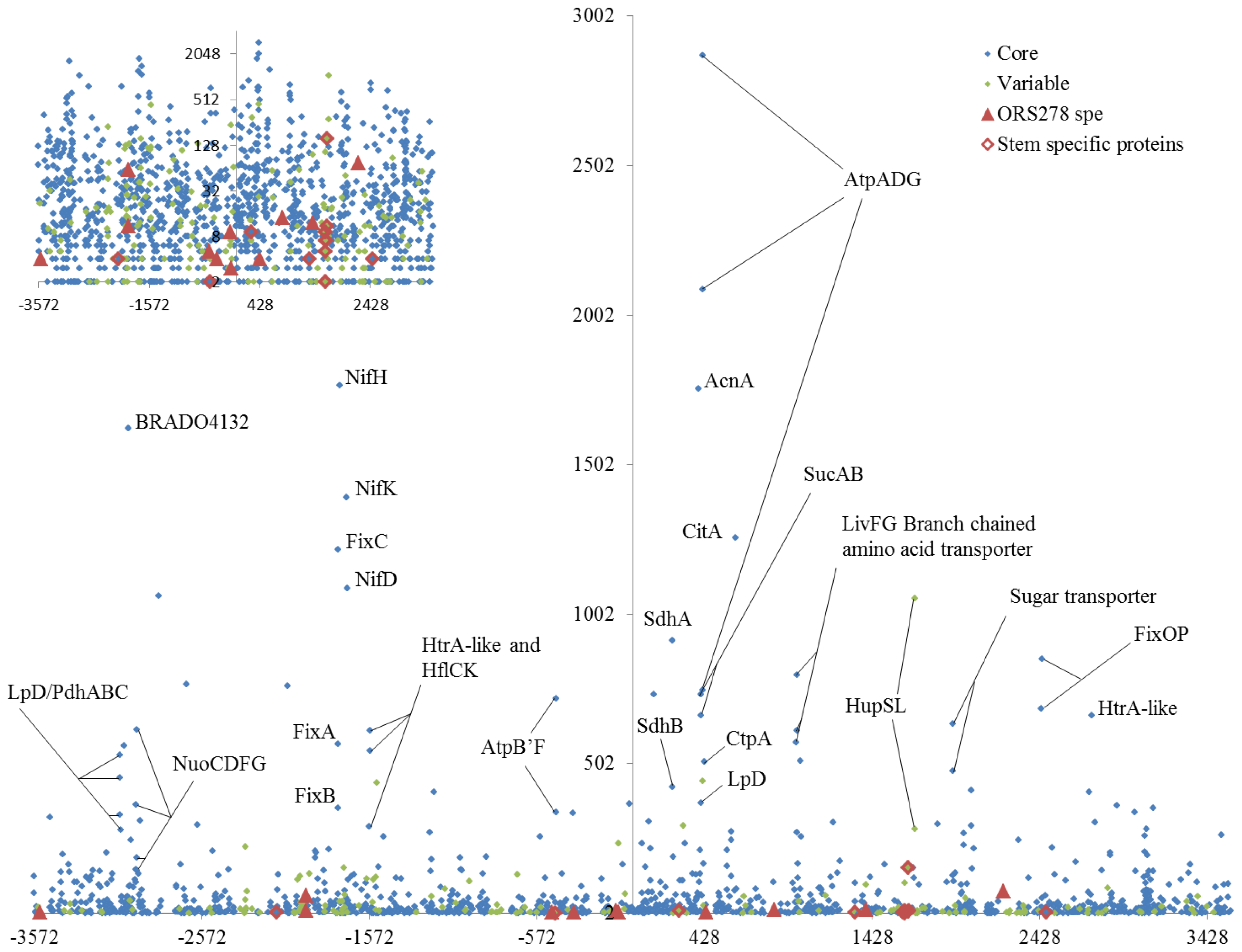
2.4. Bacteroid Most Abundant Proteins Are Involved in Central Metabolism
2.5. Comparison of Root and Stem Nodule Bacteroids Highlights the Importance of Photosynthesis for Stem Nodulation
2.6. Comparison of Proteomics and Genetics Data for Selection of Candidate Proteins for Further Functional Analysis
2.7. FixA Is Essential for the Symbiotic Process
3. Experimental Section
4. Conclusions
Acknowledgments
Conflicts of Interest
- Authors ContributionsN.D., B.G., E.G. and J.A.V. designed the experiments; N.D., B.G., C.C. and J.F. performed the experiments. B.G., E.G., S.M., B.A. analyzed the data. B.G., E.G. wrote the manuscript.
References
- Udvardi, M.; Poole, P.S. Transport and metabolism in legume-rhizobia symbioses. Ann. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar]
- Terpolilli, J.J.; Hood, G.A.; Poole, P.S. What determines the efficiency of N2-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol. 2012, 60, 325–389. [Google Scholar]
- Prell, J.; Poole, P. Metabolic changes of rhizobia in legume nodules. Trends Microbiol. 2006, 14, 161–168. [Google Scholar]
- White, J.; Prell, J.; James, E.K.; Poole, P. Nutrient Sharing between Symbionts. Plant Physiol. 2007, 144, 604–614. [Google Scholar]
- Dreyfus, B.L.; Dommergues, Y.R. Nitrogen-fixing nodules induced by Rhizobium on the stem of the tropical legume Sesbania rostrata. FEMS Microbiol. Lett. 1981, 10, 313–317. [Google Scholar]
- Alazard, D. Stem and Root Nodulation in Aeschynomene spp. Appl. Environ. Microbiol. 1985, 50, 732–734. [Google Scholar]
- Suzuki, S.; Aono, T.; Lee, K.B.; Suzuki, T.; Liu, C.T.; Miwa, H.; Wakao, S.; Iki, T.; Oyaizu, H. Rhizobial factors required for stem nodule maturation and maintenance in Sesbania rostrata–Azorhizobium caulinodans ORS571 symbiosis. Appl. Environ. Microbiol. 2007, 73, 6650–6659. [Google Scholar]
- Giraud, E.; Hannibal, L.; Fardoux, J.; Vermeglio, A.; Dreyfus, B. Effect of Bradyrhizobium photosynthesis on stem nodulation of Aeschynomene sensitiva. Proc. Natl. Acad. Sci. USA 2000, 97, 14795–14800. [Google Scholar]
- Alazard, D. Nitrogen fixation in pure culture by rhizobia isolated from stem nodules of tropical Aeschynomene species. FEMS Microbiol. Lett. 1990, 68, 177–182. [Google Scholar]
- Giraud, E.; Moulin, L.; Vallenet, D.; Barbe, V.; Cytryn, E.; Avarre, J.C.; Jaubert, M.; Simon, D.; Cartieaux, F.; Prin, Y.; et al. Legumes symbioses: Absence of Nod genes in photosynthetic bradyrhizobia. Science 2007, 316, 1307–1312. [Google Scholar]
- Knief, C.; Delmotte, N.; Vorholt, J.A. Bacterial adaptation to life in association with plants— A proteomic perspective from culture to in situ conditions. Proteomics 2011, 11, 3086–3105. [Google Scholar]
- Sarma, A.D.; Emerich, D.W. Global protein expression pattern of Bradyrhizobium japonicum bacteroids: A prelude to functional proteomics. Proteomics 2005, 5, 4170–4184. [Google Scholar]
- Delmotte, N.; Ahrens, C.H.; Knief, C.; Qeli, E.; Koch, M.; Fischer, H.M.; Vorholt, J.A.; Hennecke, H.; Pessi, G. An integrated proteomics and transcriptomics reference data set provides new insights into the Bradyrhizobium japonicum bacteroid metabolism in soybean root nodules. Proteomics 2010, 10, 1391–1400. [Google Scholar]
- Koch, M.; Delmotte, N.; Rehrauer, H.; Vorholt, J.A.; Pessi, G.; Hennecke, H. Rhizobial adaptation to hosts a new facet in the legume root-nodule symbiosis. Mol. Plant Microbe Interact. 2010, 2, 784–790. [Google Scholar]
- Kumar, D.; Yadav, A.K.; Kadimi, P.K.; Nagaraj, S.H.; Grimmond, S.M.; Dash, D. Proteogenomic analysis of Bradyrhizobium japonicum USDA110 using GenoSuite an automated multi-algorithmic pipeline. Mol. Cell. Proteomics 2014, 12, 3388–3397. [Google Scholar]
- Tatsukami, Y.; Nambu, M.; Morisaka, H.; Kuroda, K.; Ueda, M. Disclosure of the differences of Mesorhizobium loti under the free-living and symbiotic conditions by comparative proteome analysis without bacteroid isolation. BMC Microbiol. 2013, 13, 180. [Google Scholar]
- Larrainzar, E.; Wienkoop, S.; Weckwerth, W.; Ladrera, R.; Arrese-Igor, C.; Gonzalez, E.M. Medicago truncatula root nodule proteome analysis reveals differential plant and bacteroid responses to drought stress. Plant Physiol. 2007, 144, 1495–1507. [Google Scholar]
- Djordjevic, M.A. Sinorhizobium meliloti metabolism in the root nodule: A proteomic perspective. Proteomics 2004, 4, 1859–1872. [Google Scholar]
- Bonaldi, K.; Gourion, B.; Fardoux, J.; Hannibal, L.; Cartieaux, F.; Boursot, M.; Vallenet, D.; Chaintreuil, C.; Prin, Y.; Nouwen, N.; et al. Large-scale transposon mutagenesis of photosynthetic Bradyrhizobium sp strain ORS278 reveals new genetic loci putatively important for nod-independent symbiosis with Aeschynomene indica. Mol. Plant Microbe Interact. 2010, 23, 760–770. [Google Scholar]
- Vallenet, D.; Belda, E.; Calteau, A.; Cruveiller, S.; Engelen, S.; Lajus, A.; le Fevre, F.; Longin, C.; Mornico, D.; Roche, D.; et al. MicroScope—An integrated microbial resource for the curation and comparative analysis of genomic and metabolic data. Nucleic Acids Res. 2013, 41, D636–D647. [Google Scholar]
- Schubert, K.R.; Evans, H.J. Hydrogen evolution: A major factor affecting the efficiency of nitrogen fixation in nodulated symbionts. Proc. Natl. Acad. Sci. USA 1976, 73, 1207–1211. [Google Scholar]
- Ruiz-Argueso, T.; Emerich, D.W.; Evans, H.J. Hydrogenase system in legume nodules: A mechanism of providing nitrogenase with energy and protection from oxygen damage. Biochem. Biophys. Res. Commun. 1979, 86, 259–264. [Google Scholar]
- Pham, T.V.; Jimenez, C.R. An accurate paired sample test for count data. Bioinformatics 2012, 28, i596–i602. [Google Scholar]
- Giraud, E.; Fardoux, J.; Fourrier, N.; Hannibal, L.; Genty, B.; Bouyer, P.; Dreyfus, B.; Vermeglio, A. Bacteriophytochrome controls photosystem synthesis in anoxygenic bacteria. Nature 2002, 417, 202–205. [Google Scholar]
- Giraud, E.; Vermeglio, A. Bacteriophytochromes in anoxygenic photosynthetic bacteria. Photosynth. Res. 2008, 97, 141–153. [Google Scholar]
- Gubler, M.; Hennecke, H. fixA B and C genes are essential for symbiotic and free-living microaerobic nitrogen fixation. FEBS Lett. 1986, 200, 186–192. [Google Scholar]
- Kaminski, P.A.; Norel, F.; Desnoues, N.; Kush, A.; Salzano, G.; Elmerich, C. Characterization of the fixABC region of Azorhizobium caulinodans ORS571 and identification of a new nitrogen fixation gene. Mol. Gen. Genet. 1988, 214, 496–502. [Google Scholar]
- Oke, V.; Long, S.R. Bacterial genes induced within the nodule during the Rhizobium-legume symbiosis. Mol. Microbiol. 1999, 32, 837–849. [Google Scholar]
- Ehrhardt, D.W.; Atkinson, E.M.; Long, S.R. Depolarization of alfalfa root hair membrane potential by Rhizobium meliloti Nod factors. Science 1992, 256, 998–1000. [Google Scholar]
- Gourion, B.; Delmotte, N.; Bonaldi, K.; Nouwen, N.; Vorholt, J.A.; Giraud, E. Bacterial RuBisCO is required for efficient Bradyrhizobium/Aeschynomene symbiosis. PLoS One 2011, 6, e21900. [Google Scholar]
- Delmotte, N.; Knief, C.; Chaffron, S.; Innerebner, G.; Roschitzki, B.; Schlapbach, R.; von Mering, C.; Vorholt, J.A. Community proteogenomics reveals insights into the physiology of phyllosphere bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 16428–16433. [Google Scholar] [Green Version]
- Giraud, E.; Lavergne, J.; Vermeglio, A. Characterization of bacteriophytochromes from photosynthetic bacteria: Histidine kinase signaling triggered by light and redox sensing. Methods Enzymol. 2010, 471, 135–159. [Google Scholar]


© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Delmotte, N.; Mondy, S.; Alunni, B.; Fardoux, J.; Chaintreuil, C.; Vorholt, J.A.; Giraud, E.; Gourion, B. A Proteomic Approach of Bradyrhizobium/Aeschynomene Root and Stem Symbioses Reveals the Importance of the fixA Locus for Symbiosis. Int. J. Mol. Sci. 2014, 15, 3660-3670. https://doi.org/10.3390/ijms15033660
Delmotte N, Mondy S, Alunni B, Fardoux J, Chaintreuil C, Vorholt JA, Giraud E, Gourion B. A Proteomic Approach of Bradyrhizobium/Aeschynomene Root and Stem Symbioses Reveals the Importance of the fixA Locus for Symbiosis. International Journal of Molecular Sciences. 2014; 15(3):3660-3670. https://doi.org/10.3390/ijms15033660
Chicago/Turabian StyleDelmotte, Nathanael, Samuel Mondy, Benoit Alunni, Joel Fardoux, Clémence Chaintreuil, Julia A. Vorholt, Eric Giraud, and Benjamin Gourion. 2014. "A Proteomic Approach of Bradyrhizobium/Aeschynomene Root and Stem Symbioses Reveals the Importance of the fixA Locus for Symbiosis" International Journal of Molecular Sciences 15, no. 3: 3660-3670. https://doi.org/10.3390/ijms15033660