Lipidomic Analysis of Serum from High Fat Diet Induced Obese Mice



Abstract
:1. Introduction
2. Results
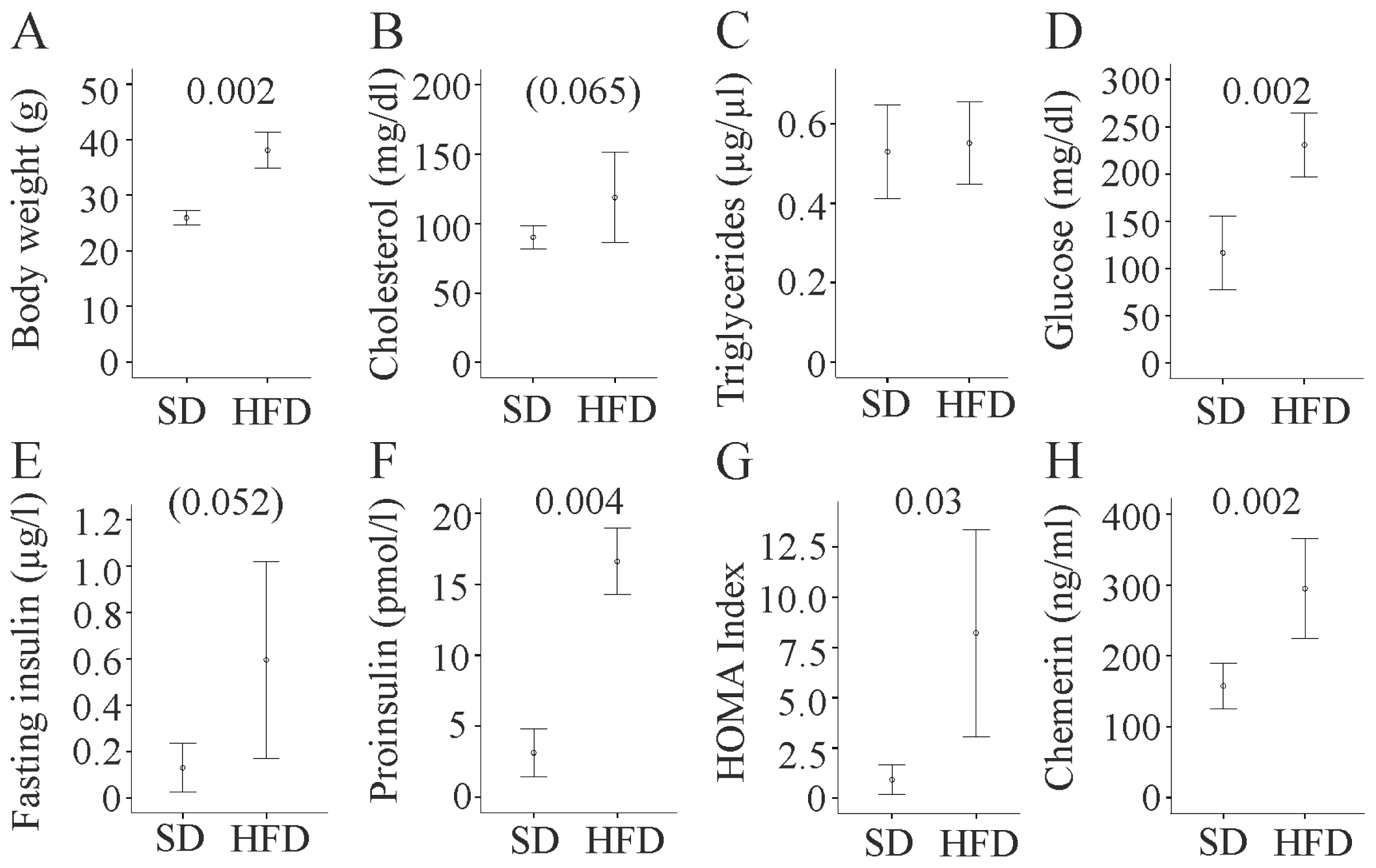
2.1. Metabolic Profile of Fat Fed Mice
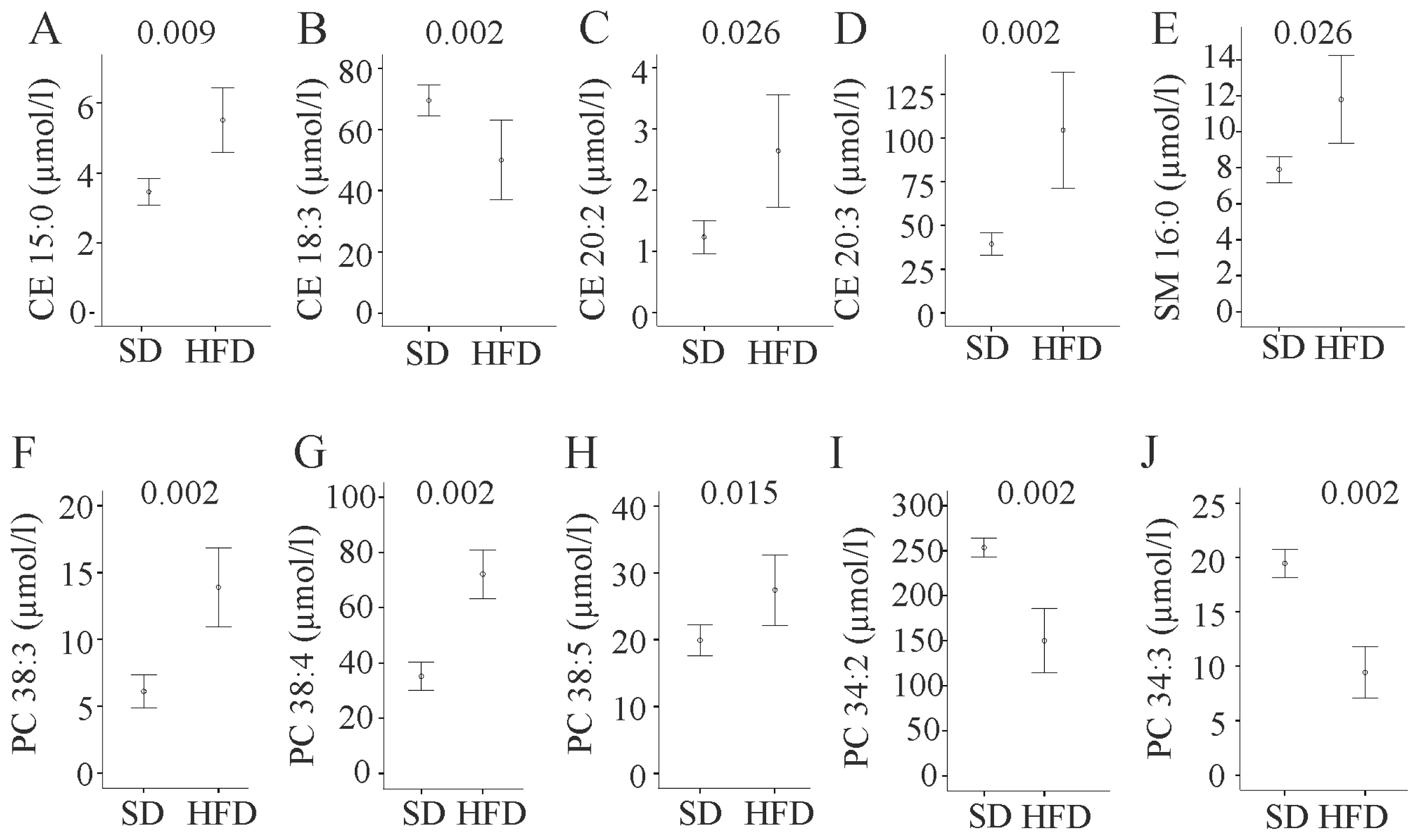
2.2. Cholesterol Species
2.3. Sphingomyelin and Ceramides
2.4. Phosphatidylcholine
2.5. Lysophosphatidylcholine
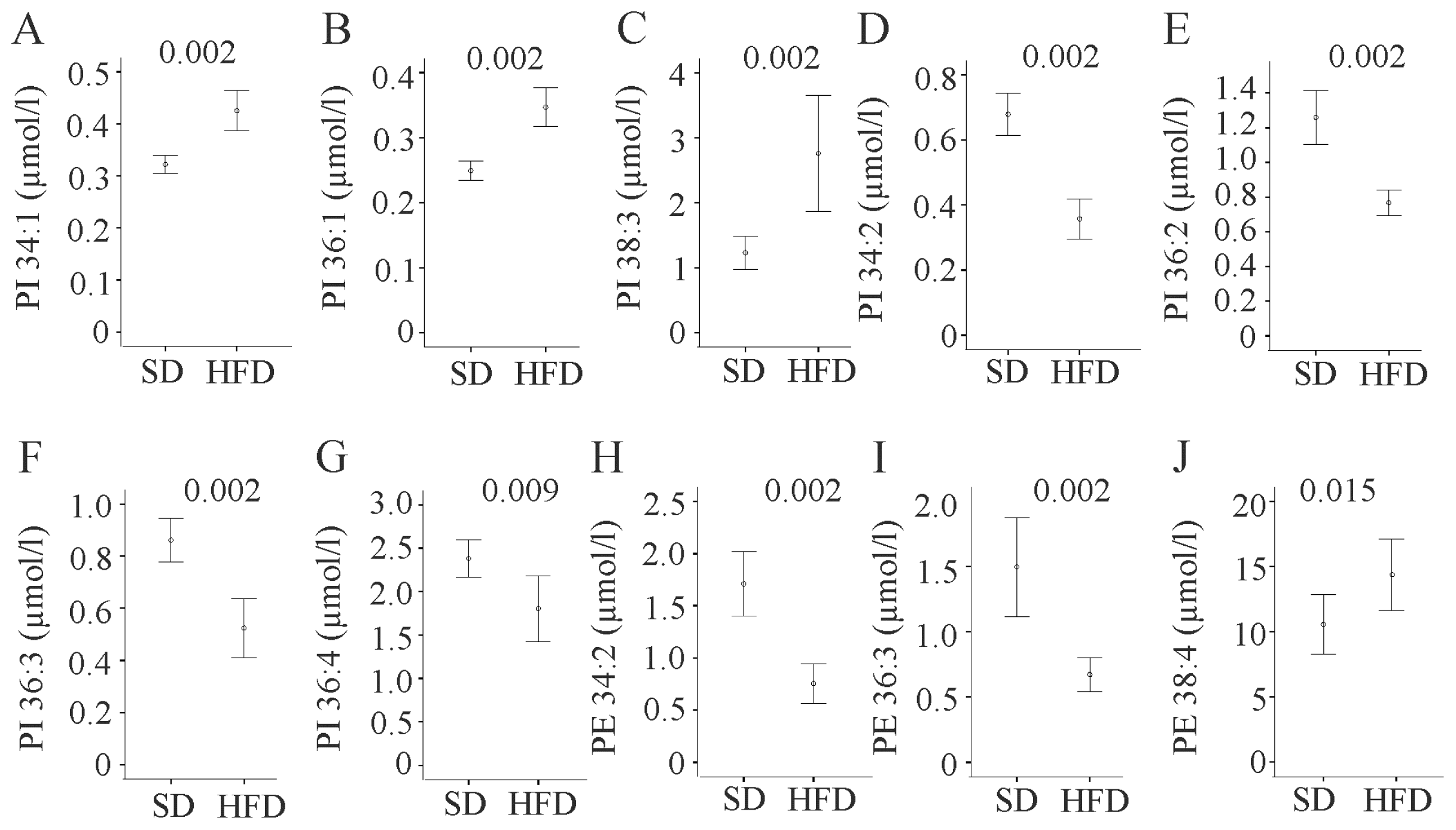
2.6. Phosphatidylinositol
2.7. Phosphatidylethanolamine
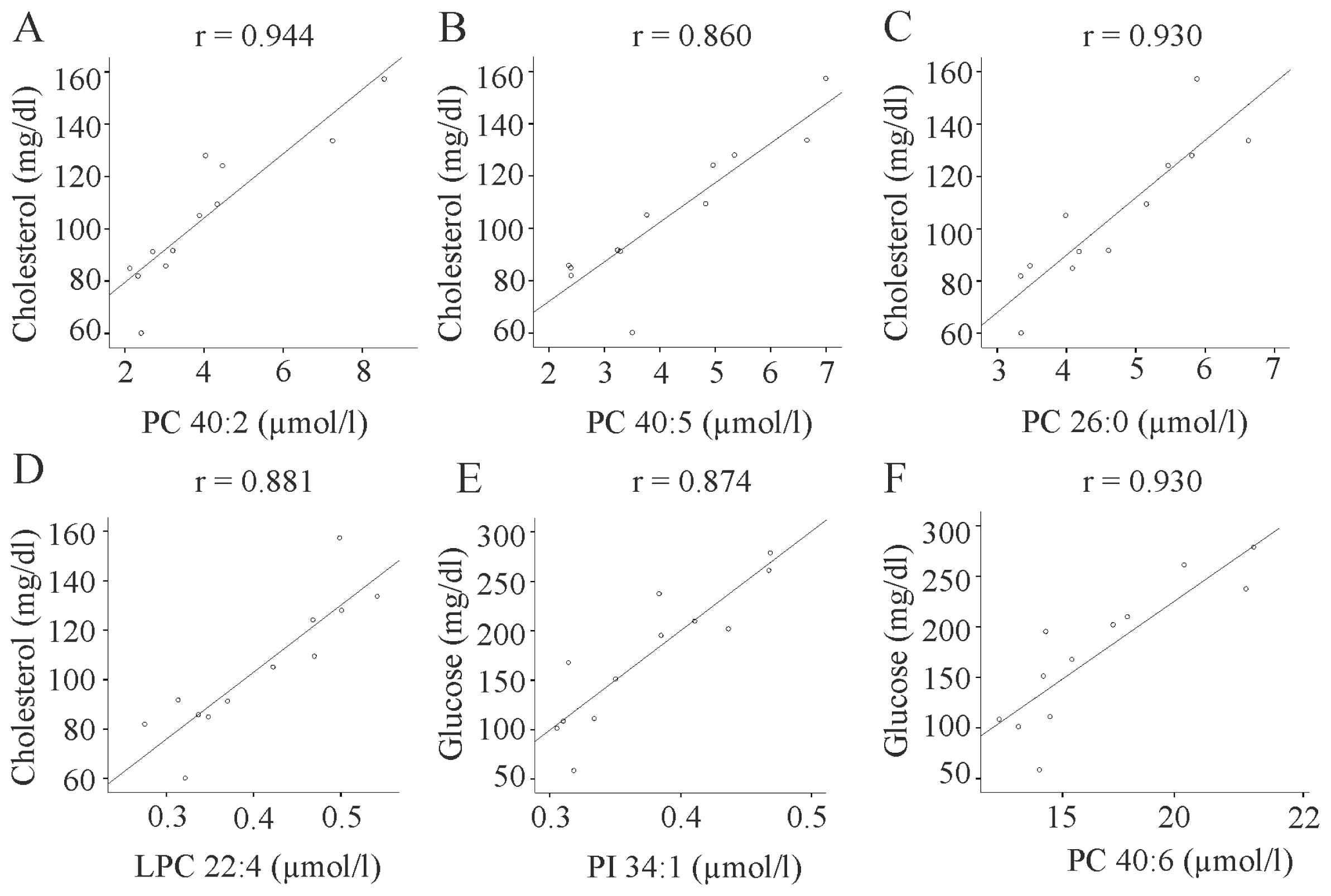
2.8. Body Weight Independent Correlations
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Animal Model
4.3. Quantification of Lipids
4.4. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-15-02991-s001.pdfAcknowledgments
Conflicts of Interest
References
- Boden, G.; Shulman, G.I. Free fatty acids in obesity and type 2 diabetes: Defining their role in the development of insulin resistance and beta-cell dysfunction. Eur. J. Clin. Investig 2002, 32, 14–23. [Google Scholar]
- Ogden, C.L.; Carroll, M.D.; Curtin, L.R.; McDowell, M.A.; Tabak, C.J.; Flegal, K.M. Prevalence of overweight and obesity in the United States, 1999–2004. JAMA 2006, 295, 1549–1555. [Google Scholar]
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol 2011, 17, 2801–2811. [Google Scholar]
- Taskinen, M.R. Type 2 diabetes as a lipid disorder. Curr. Mol. Med 2005, 5, 297–308. [Google Scholar]
- Yusuf, S.; Hawken, S.; Ounpuu, S.; Dans, T.; Avezum, A.; Lanas, F.; McQueen, M.; Budaj, A.; Pais, P.; Varigos, J.; et al. Effect of potentially modifiable risk factors associated with myocardial infarction in 52 countries (the INTERHEART study): Case-control study. Lancet 2004, 364, 937–952. [Google Scholar]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid Res 2010, 51, 3299–3305. [Google Scholar]
- Pietilainen, K.H.; Sysi-Aho, M.; Rissanen, A.; Seppanen-Laakso, T.; Yki-Jarvinen, H.; Kaprio, J.; Oresic, M. Acquired obesity is associated with changes in the serum lipidomic profile independent of genetic effects--a monozygotic twin study. PLoS One 2007, 2, e218. [Google Scholar]
- Kim, J.Y.; Park, J.Y.; Kim, O.Y.; Ham, B.M.; Kim, H.J.; Kwon, D.Y.; Jang, Y.; Lee, J.H. Metabolic profiling of plasma in overweight/obese and lean men using ultra performance liquid chromatography and Q-TOF mass spectrometry (UPLC-Q-TOF MS). J. Proteome Res 2010, 9, 4368–4375. [Google Scholar]
- Floegel, A.; Stefan, N.; Yu, Z.; Muhlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Haring, H.U.; Hrabe de Angelis, M.; Peters, A.; et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes 2013, 62, 639–648. [Google Scholar]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma lysophosphatidylcholine levels are reduced in obesity and type 2 diabetes. PLoS One 2012, 7, e41456. [Google Scholar]
- Kim, H.J.; Kim, J.H.; Noh, S.; Hur, H.J.; Sung, M.J.; Hwang, J.T.; Park, J.H.; Yang, H.J.; Kim, M.S.; Kwon, D.Y.; et al. Metabolomic analysis of livers and serum from high-fat diet induced obese mice. J. Proteome Res 2011, 10, 722–731. [Google Scholar]
- An, Y.; Xu, W.; Li, H.; Lei, H.; Zhang, L.; Hao, F.; Duan, Y.; Yan, X.; Zhao, Y.; Wu, J.; et al. High-fat diet induces dynamic metabolic alterations in multiple biological matrices of rats. J. Proteome Res 2013, 12, 3755–3768. [Google Scholar]
- Boini, K.M.; Zhang, C.; Xia, M.; Poklis, J.L.; Li, P.L. Role of sphingolipid mediator ceramide in obesity and renal injury in mice fed a high-fat diet. J. Pharmacol. Exp. Ther 2010, 334, 839–846. [Google Scholar]
- Shah, C.; Yang, G.; Lee, I.; Bielawski, J.; Hannun, Y.A.; Samad, F. Protection from high fat diet-induced increase in ceramide in mice lacking plasminogen activator inhibitor 1. J. Biol. Chem 2008, 283, 13538–13548. [Google Scholar]
- Fox, T.E.; Bewley, M.C.; Unrath, K.A.; Pedersen, M.M.; Anderson, R.E.; Jung, D.Y.; Jefferson, L.S.; Kim, J.K.; Bronson, S.K.; Flanagan, J.M.; et al. Circulating sphingolipid biomarkers in models of type 1 diabetes. J. Lipid Res 2011, 52, 509–517. [Google Scholar]
- Boon, J.; Hoy, A.J.; Stark, R.; Brown, R.D.; Meex, R.C.; Henstridge, D.C.; Schenk, S.; Meikle, P.J.; Horowitz, J.F.; Kingwell, B.A.; et al. Ceramides contained in LDL are elevated in type 2 diabetes and promote inflammation and skeletal muscle insulin resistance. Diabetes 2013, 62, 401–410. [Google Scholar]
- Haus, J.M.; Kashyap, S.R.; Kasumov, T.; Zhang, R.; Kelly, K.R.; Defronzo, R.A.; Kirwan, J.P. Plasma ceramides are elevated in obese subjects with type 2 diabetes and correlate with the severity of insulin resistance. Diabetes 2009, 58, 337–343. [Google Scholar]
- Holland, W.L.; Brozinick, J.T.; Wang, L.P.; Hawkins, E.D.; Sargent, K.M.; Liu, Y.; Narra, K.; Hoehn, K.L.; Knotts, T.A.; Siesky, A.; et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab 2007, 5, 167–179. [Google Scholar]
- Liu, S.; Brown, J.D.; Stanya, K.J.; Homan, E.; Leidl, M.; Inouye, K.; Bhargava, P.; Gangl, M.R.; Dai, L.; Hatano, B.; et al. A diurnal serum lipid integrates hepatic lipogenesis and peripheral fatty acid use. Nature 2013, 502, 550–554. [Google Scholar]
- Bauer, S.; Wanninger, J.; Schmidhofer, S.; Weigert, J.; Neumeier, M.; Dorn, C.; Hellerbrand, C.; Zimara, N.; Schaffler, A.; Aslanidis, C.; et al. Sterol regulatory element-binding protein 2 (SREBP2) activation after excess triglyceride storage induces Chemerin in hypertrophic adipocytes. Endocrinology 2011, 152, 26–35. [Google Scholar]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin is a novel adipokine associated with obesity and metabolic syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar]
- Lee, R.G.; Kelley, K.L.; Sawyer, J.K.; Farese, R.V., Jr.; Parks, J.S.; Rudel, L.L. Plasma cholesteryl esters provided by lecithin:cholesterol acyltransferase and acyl-coenzyme a: Cholesterol acyltransferase 2 have opposite atherosclerotic potential. Circ. Res 2004, 95, 998–1004. [Google Scholar]
- Chavez, J.A.; Summers, S.A. A ceramide-centric view of insulin resistance. Cell Metab 2012, 15, 585–594. [Google Scholar]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of lysophosphatidylcholine (LPC) in atherosclerosis. Curr. Med. Chem 2007, 14, 3209–3220. [Google Scholar]
- Roberts, C.K.; Liang, K.; Barnard, R.J.; Kim, C.H.; Vaziri, N.D. HMG-CoA reductase, cholesterol 7alpha-hydroxylase, LDL receptor, SR-B1, and ACAT in diet-induced syndrome X. Kidney Int 2004, 66, 1503–1511. [Google Scholar]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered adipose and plasma sphingolipid metabolism in obesity: A potential mechanism for cardiovascular and metabolic risk. Diabetes 2006, 55, 2579–2587. [Google Scholar]
- Ichi, I.; Nakahara, K.; Kiso, K.; Kojo, S. Effect of dietary cholesterol and high fat on ceramide concentration in rat tissues. Nutrition 2007, 23, 570–574. [Google Scholar]
- Masi, L.N.; Rodrigues, A.C.; Curi, R. Fatty acids regulation of inflammatory and metabolic genes. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 418–424. [Google Scholar]
- Lankinen, M.; Schwab, U.; Kolehmainen, M.; Paananen, J.; Poutanen, K.; Mykkanen, H.; Seppanen-Laakso, T.; Gylling, H.; Uusitupa, M.; Oresic, M. Whole grain products, fish and bilberries alter glucose and lipid metabolism in a randomized, controlled trial: The Sysdimet study. PLoS One 2011, 6, e22646. [Google Scholar]
- Szymanska, E.; van Dorsten, F.A.; Troost, J.; Paliukhovich, I.; van Velzen, E.J.; Hendriks, M.M.; Trautwein, E.A.; van Duynhoven, J.P.; Vreeken, R.J.; Smilde, A.K. A lipidomic analysis approach to evaluate the response to cholesterol-lowering food intake. Metabolomics 2012, 8, 894–906. [Google Scholar]
- Floegel, A.; von Ruesten, A.; Drogan, D.; Schulze, M.B.; Prehn, C.; Adamski, J.; Pischon, T.; Boeing, H. Variation of serum metabolites related to habitual diet: A targeted metabolomic approach in EPIC-Potsdam. Eur. J. Clin. Nutr 2013, 67, 1100–1108. [Google Scholar]
- Li, Z.; Agellon, L.B.; Vance, D.E. A role for high density lipoproteins in hepatic phosphatidylcholine homeostasis. Biochim. Biophys. Acta 2007, 1771, 893–900. [Google Scholar]
- Ober, C.; Loisel, D.A.; Gilad, Y. Sex-specific genetic architecture of human disease. Nat. Rev. Genet 2008, 9, 911–922. [Google Scholar]
- Liebisch, G.; Lieser, B.; Rathenberg, J.; Drobnik, W.; Schmitz, G. High-throughput quantification of phosphatidylcholine and sphingomyelin by electrospray ionization tandem mass spectrometry coupled with isotope correction algorithm. Biochim. Biophys. Acta 2004, 1686, 108–117. [Google Scholar]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res 2008, 49, 1137–1146. [Google Scholar]
- Liebisch, G.; Drobnik, W.; Reil, M.; Trumbach, B.; Arnecke, R.; Olgemoller, B.; Roscher, A.; Schmitz, G. Quantitative measurement of different ceramide species from crude cellular extracts by electrospray ionization tandem mass spectrometry (ESI-MS/MS). J. Lipid Res 1999, 40, 1539–1546. [Google Scholar]
- Liebisch, G.; Binder, M.; Schifferer, R.; Langmann, T.; Schulz, B.; Schmitz, G. High throughput quantification of cholesterol and cholesteryl ester by electrospray ionization tandem mass spectrometry (ESI-MS/MS). Biochim. Biophys. Acta 2006, 1761, 121–128. [Google Scholar]
- Liebisch, G.; Vizcaino, J.A.; Kofeler, H.; Trotzmuller, M.; Griffiths, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res 2013, 54, 1523–1530. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid | SD | Std. dev. | HFD | Std. dev. | p-value | Regulation | Regulation Kim et al. | Regulation Barber et al. |
|---|---|---|---|---|---|---|---|---|
| LPC 15:0 | 0.48 | 0.05 | 0.55 | 0.10 | 0.093 | - | ↓ | ↓ |
| LPC 16:1 | 9.36 | 0.67 | 5.51 | 0.94 | 0.002 | ↓ | ↓ | ↓ |
| LPC 16:0 | 69.84 | 4.11 | 60.02 | 13.01 | 0.180 | - | ↓ | ↓ |
| LPC 18:3 | 1.80 | 0.11 | 0.90 | 0.16 | 0.002 | ↓ | ↑ | n.d. |
| LPC 18:2 | 39.02 | 2.24 | 18.04 | 3.14 | 0.002 | ↓ | ↓ | - |
| LPC 18:1 | 82.73 | 5.60 | 84.19 | 16.24 | 0.310 | - | ↓ | ↓ |
| LPC 18:0 | 26.76 | 2.60 | 39.35 | 8.13 | 0.065 | - | ↑ | ↑ |
| LPC 20:5 | 0.48 | 0.09 | 0.57 | 0.08 | 0.180 | - | ↓ | ↓ |
| LPC 20:4 | 21.15 | 2.60 | 26.00 | 2.83 | 0.026 | ↑ | ↓ | ↑ |
| LPC 20:3 | 2.21 | 0.44 | 4.35 | 0.69 | 0.002 | ↑ | n.d. | - |
| LPC 20:0 | 2.10 | 0.25 | 1.24 | 0.26 | 0.002 | ↓ | n.d. | - |
| LPC 22:6 | 6.65 | 0.58 | 6.93 | 1.22 | 0.485 | - | n.d. | - |
| LPC 22:5 | 0.81 | 0.10 | 1.11 | 0.31 | 0.065 | - | n.d. | n.d. |
| LPC 22:4 | 0.34 | 0.05 | 0.47 | 0.07 | 0.026 | ↑ | n.d. | n.d. |
| LPC 22:0 | 3.81 | 0.35 | 5.23 | 1.50 | 0.065 | - | n.d. | n.d. |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Eisinger, K.; Liebisch, G.; Schmitz, G.; Aslanidis, C.; Krautbauer, S.; Buechler, C. Lipidomic Analysis of Serum from High Fat Diet Induced Obese Mice. Int. J. Mol. Sci. 2014, 15, 2991-3002. https://doi.org/10.3390/ijms15022991
Eisinger K, Liebisch G, Schmitz G, Aslanidis C, Krautbauer S, Buechler C. Lipidomic Analysis of Serum from High Fat Diet Induced Obese Mice. International Journal of Molecular Sciences. 2014; 15(2):2991-3002. https://doi.org/10.3390/ijms15022991
Chicago/Turabian StyleEisinger, Kristina, Gerhard Liebisch, Gerd Schmitz, Charalampos Aslanidis, Sabrina Krautbauer, and Christa Buechler. 2014. "Lipidomic Analysis of Serum from High Fat Diet Induced Obese Mice" International Journal of Molecular Sciences 15, no. 2: 2991-3002. https://doi.org/10.3390/ijms15022991