A Novel F-Box Protein CaF-Box Is Involved in Responses to Plant Hormones and Abiotic Stress in Pepper (Capsicum annuum L.)

Abstract
:1. Introduction
2. Results and Discussion
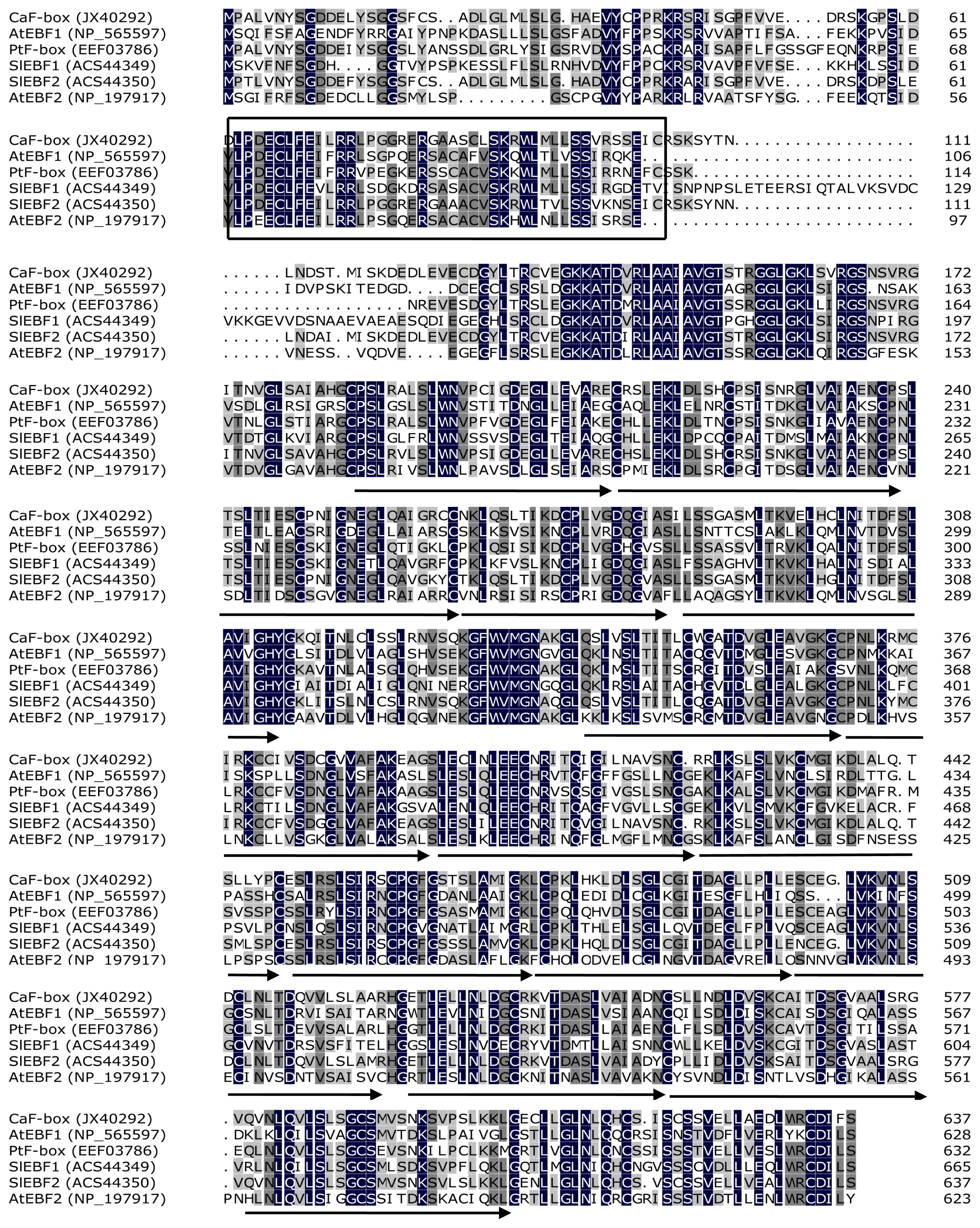
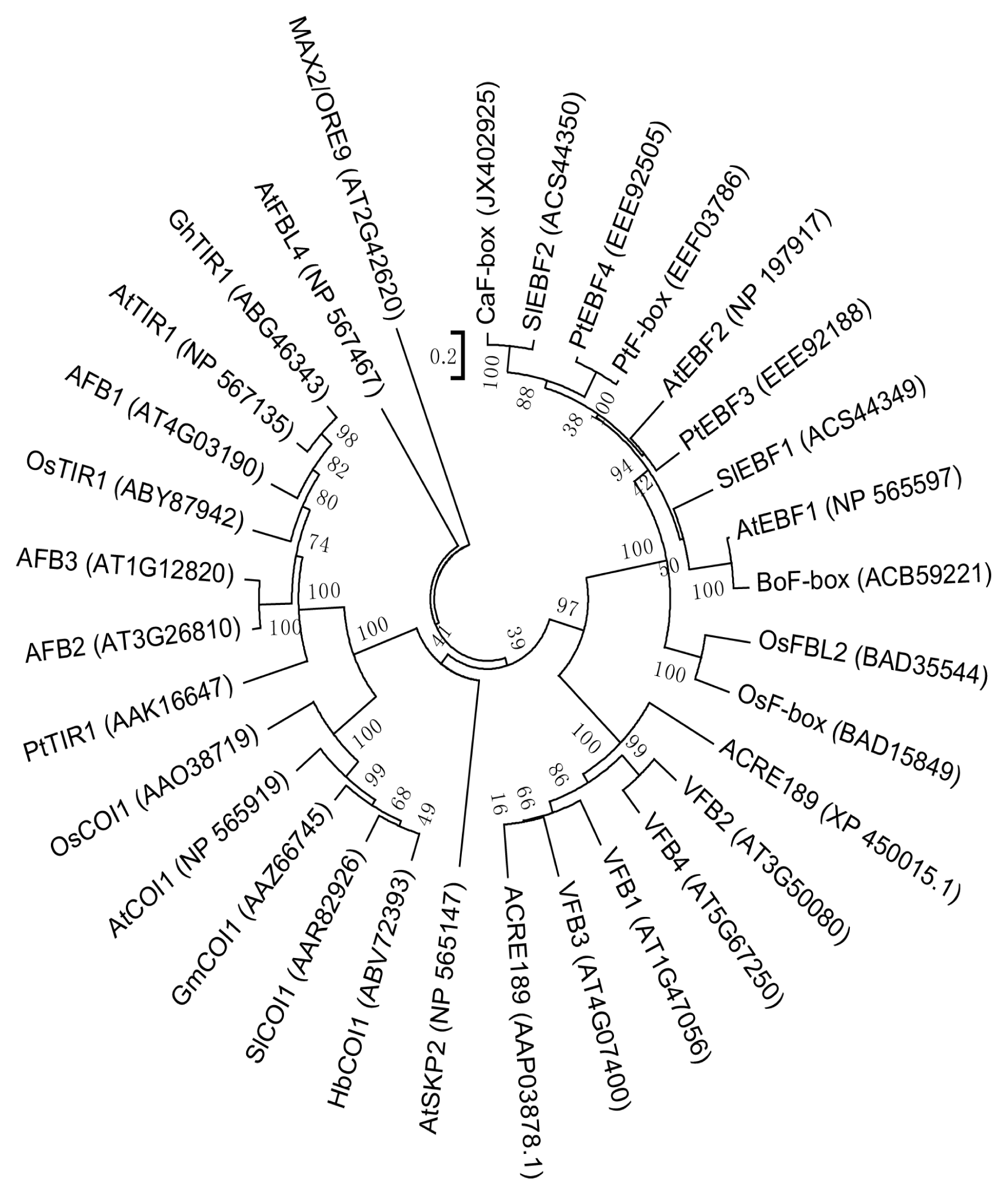
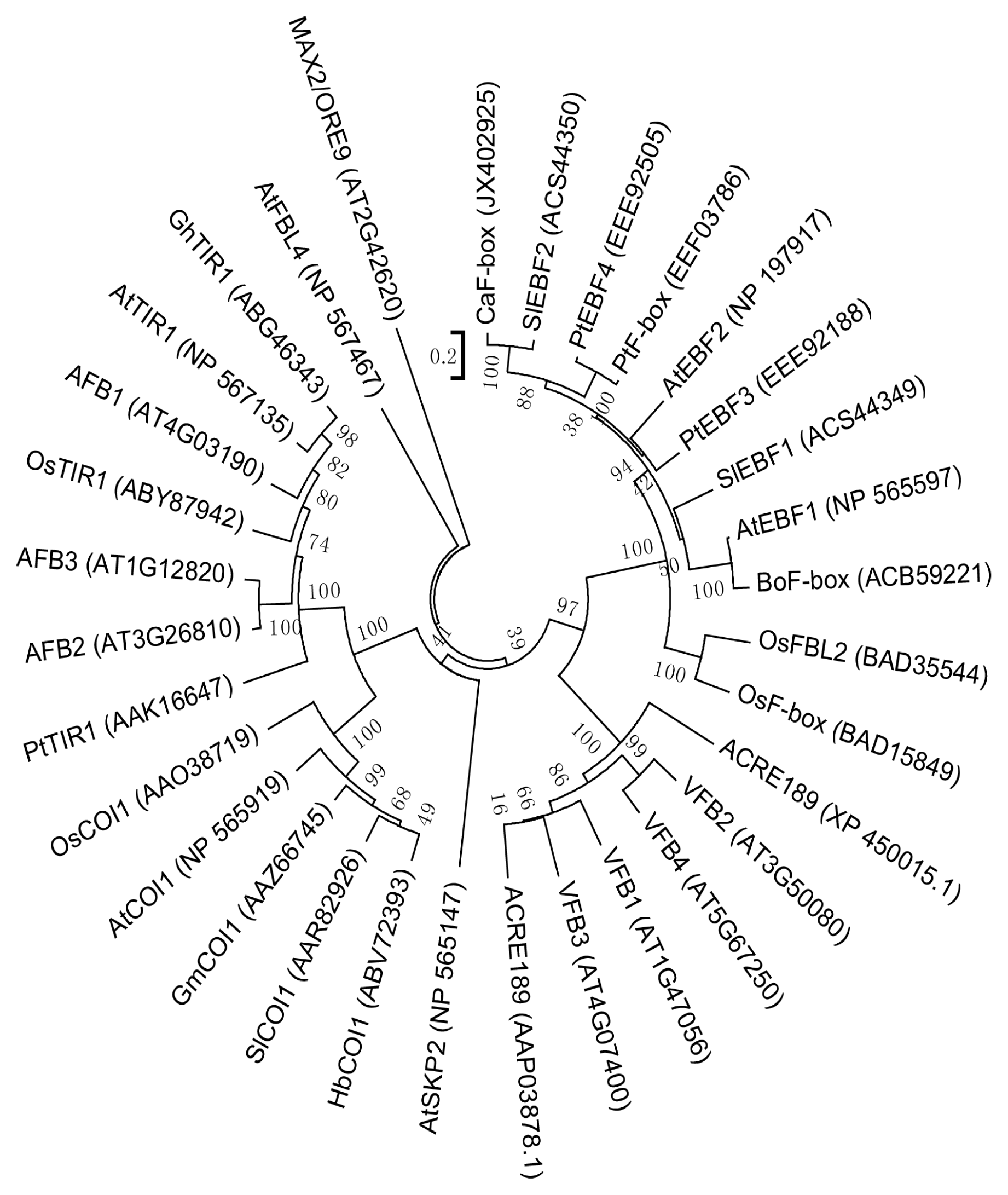
2.1. Isolation of CaF-Box and Sequence Analysis
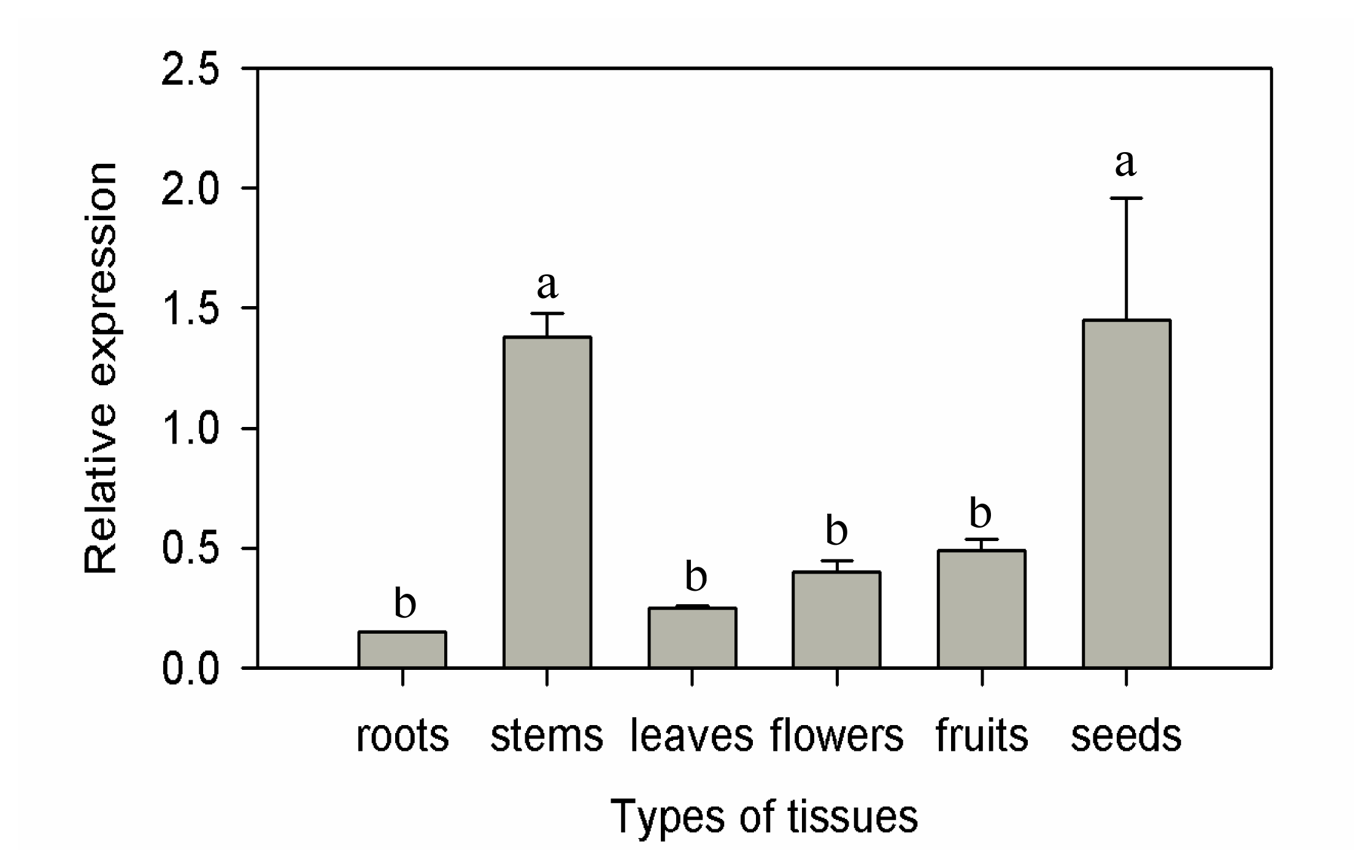
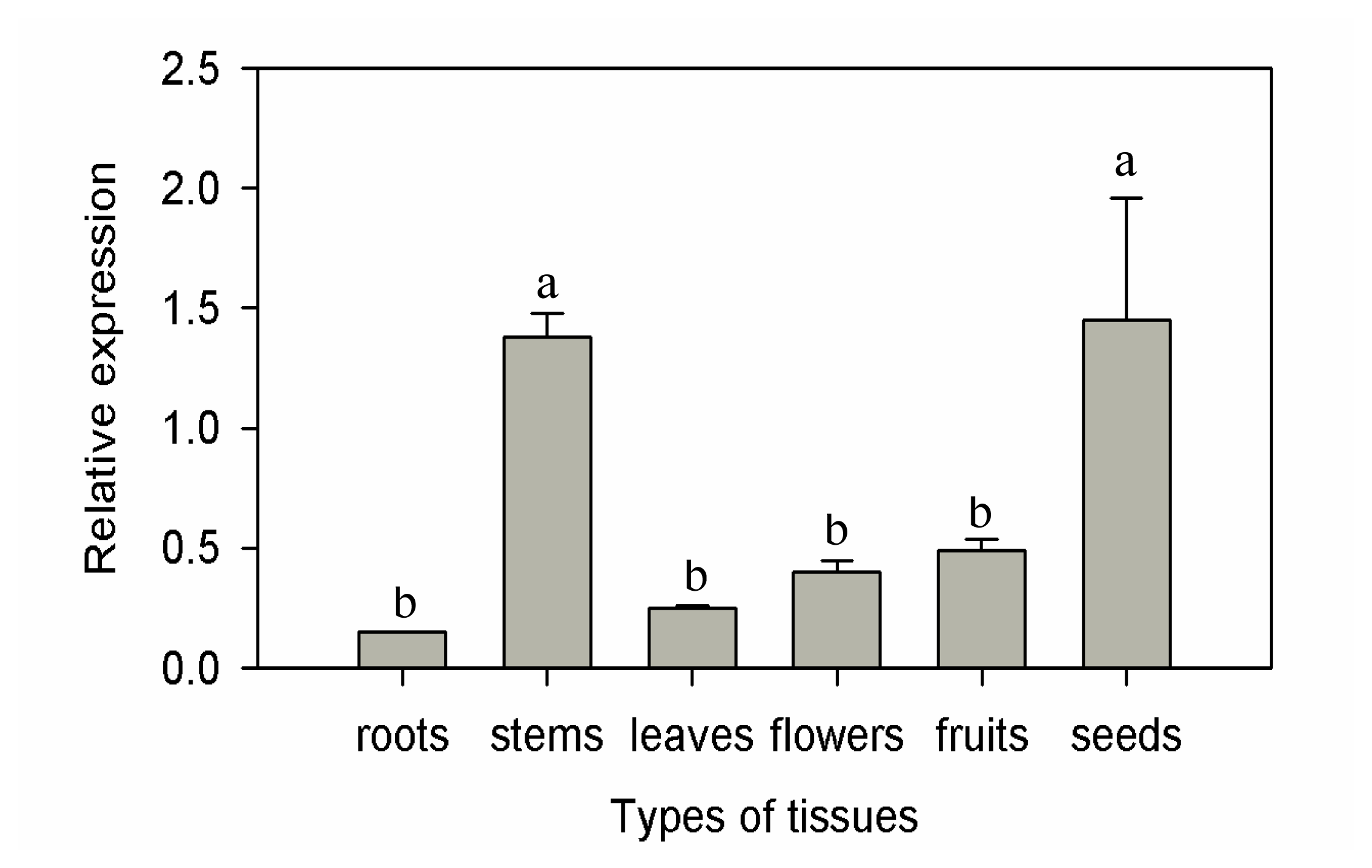
2.2. Expression Patterns of CaF-Box in Different Tissues of Pepper Plants
2.3. Expression Patterns of CaF-Box Gene in Response to Abiotic Stress and Plant Hormones Treatment
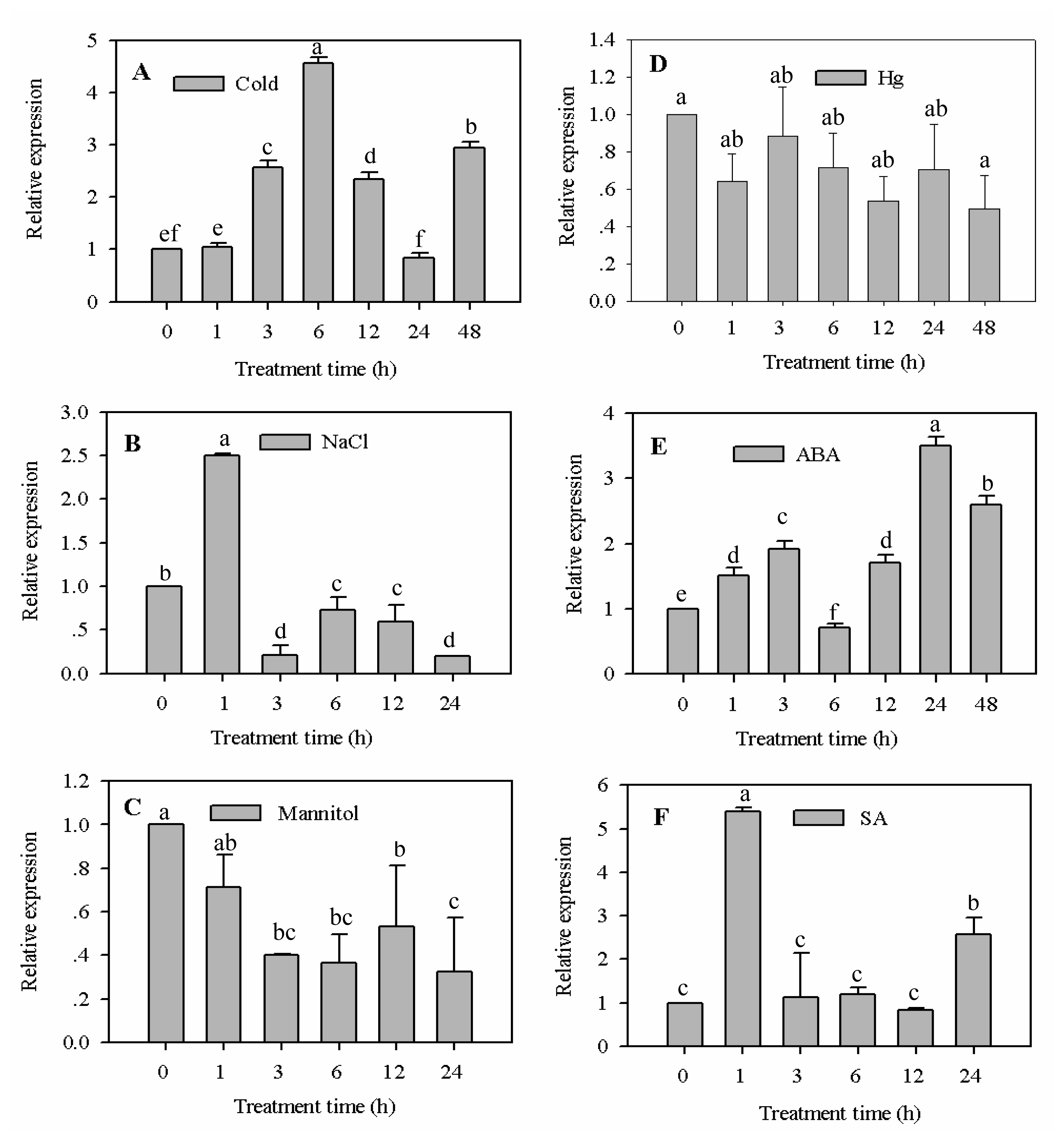
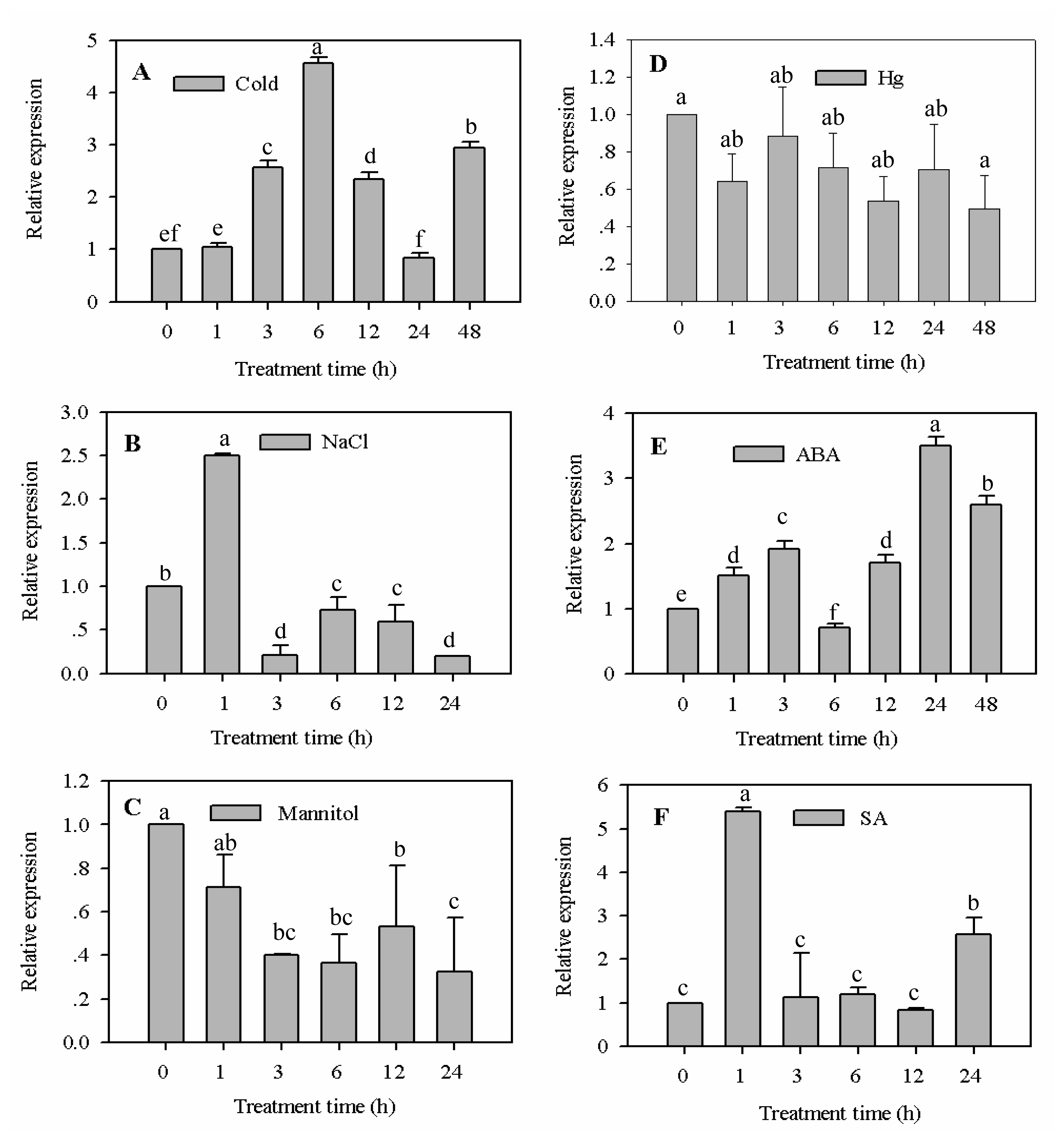
2.3.1. Expression Analysis of CaF-Box Gene in Response to Cold, Salt, Osmotic, and Heavy Metal Stress Treatments
2.3.2. Expression Analysis of CaF-Box Gene in Response to ABA and SA Treatment
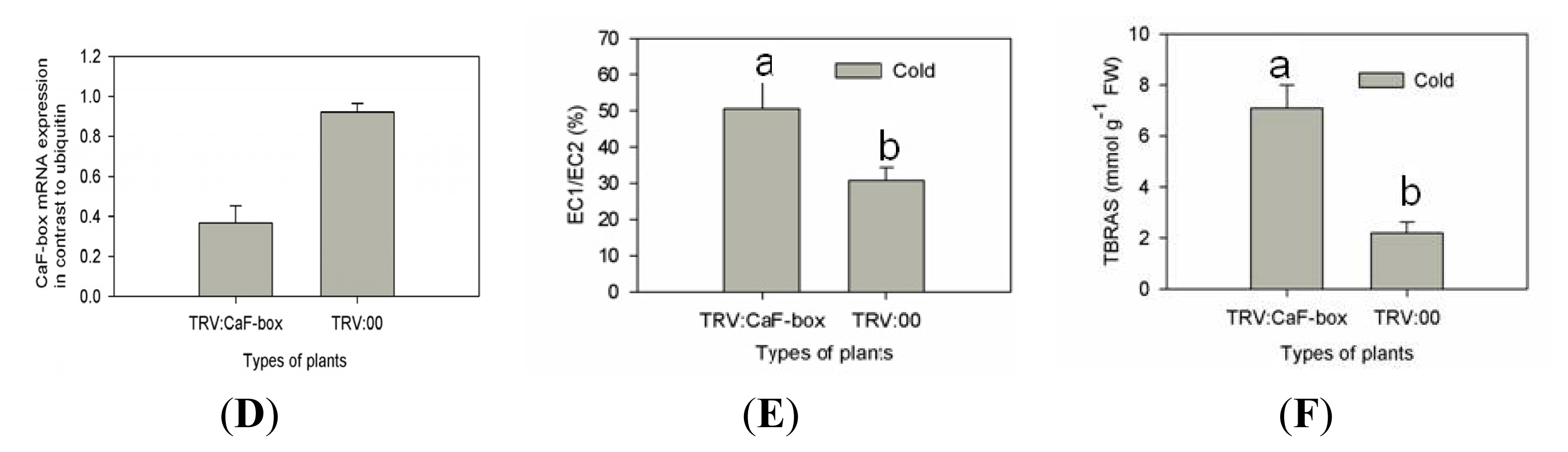
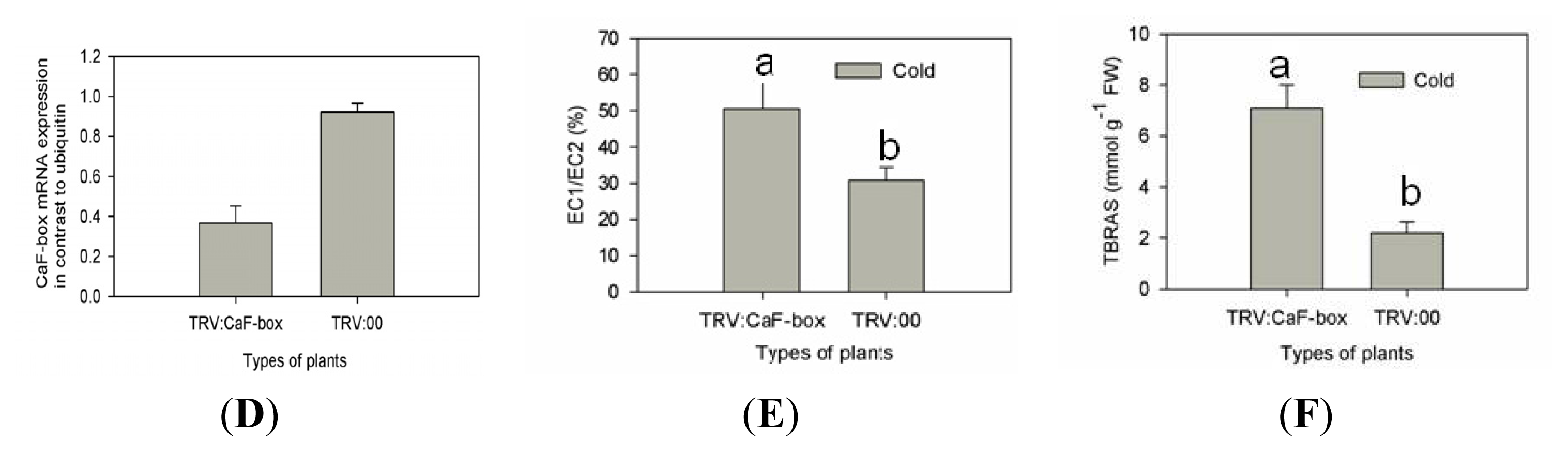
2.4. Suppression of CaF-Box Results in Reduced Tolerance to Cold Stress
3. Experimental Section
3.1. Plant Materials and Growth Conditions
3.2. Bacterial Strains, Vectors, Enzymes
3.3. Isolation of CaF-Box cDNA Clone and Sequence Analysis
3.4. CaF-Box Gene Expression Patterns Analysis
3.4.1. Tissue-Specific Expression of CaF-Box Gene
3.4.2. Stress Treatments
3.5. RNA Isolation and Real-Time RT-PCR Analysis
3.6. Virus-Induced Gene Silencing of CaF-Box in Pepper
3.7. TBARS and Electrolyte Leakage Level Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Smalle, J.; Vierstra, R.D. The ubiquity in 26S proteasome proteolytic pathway. Annu. Rev. Plant Biol 2004, 55, 555–590. [Google Scholar]
- Dreher, K.; Callis, J. Ubiquitin, hormones and biotic stress in plants. Ann. Bot 2007, 99, 787–822. [Google Scholar]
- Vierstra, R.D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol 2009, 110, 385–397. [Google Scholar]
- Hellmann, H.; Estelle, M. Plant development: Regulation by protein degradation. Science 2002, 297, 793–797. [Google Scholar]
- Cardozo, T.; Pagano, M. The SCF ubiquitin ligase: Insights into a molecular machine. Nat. Rev. Mol. Cell Biol 2004, 5, 739–751. [Google Scholar]
- Zheng, N.; Schulman, B.A.; Song, L.; Miller, J.J.; Jeffrey, P.D.; Wang, P.; Chu, C.; Koepp, D.M.; Elledge, S.J.; Pagano, M.; et al. Structure of the Cul1-Rbx1-Skp1-F box Skp2 SCF ubiquitin ligase complex. Nature 2002, 416, 703–709. [Google Scholar]
- Gagne, J.M.; Downes, B.P.; Shiu, S.H.; Durski, A.M.; Vierstra, R.D. The F-box subunit of the SCF E3 complex is encoded by a diverse superfamily of genes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2002, 99, 11519–11524. [Google Scholar]
- Jin, J.; Cardozo, T.; Lovering, R.C.; Elledge, S.J.; Pagano, M.; Harper, J.W. Systematic analysis and nomenclature of mammalian F-box proteins. Genes Dev 2004, 18, 2573–2580. [Google Scholar]
- Kipreos, E.T.; Pagano, M. The F-box protein family. Genome Biol 2000, 1, REVIEWS3002. [Google Scholar]
- Xu, G.X.; Ma, H.; Nei, M.; Kong, H.Z. Evolution of F-box genes in plants: Different modes of sequence divergence and their relationships with functional diversification. Proc. Natl. Acad. Sci. USA 2009, 106, 835–840. [Google Scholar]
- Santner, A.; Estelle, M. The ubiquitin-proteasome system regulates plant hormone signaling. Plant J 2010, 61, 1029–1040. [Google Scholar]
- Lee, J.H.; Kim, W.T. Regulation of abiotic stress signal transduction by E3 ubiquitin ligases in Arabidopsis. Mol. Cell 2011, 31, 201–208. [Google Scholar]
- Song, S.Y.; Dai, X.Y.; Zhang, W.H. A rice F box gene, OsFbx352, is involved in glucose-delayed seed germination in rice. J. Exp. Bot 2012, 63, 5550–5568. [Google Scholar]
- Levin, J.Z.; Meyerowitz, E.M. UFO: An Arabidopsis gene involved in both floral meristem and floral organ development. Plant Cell 1995, 7, 529–548. [Google Scholar]
- Hepworth, S.R.; Klenz, J.E.; Haughn, G.W. UFO in the Arabidopsis inflorescence apex is required for floral-meristem identity and bract suppression. Planta 2006, 223, 769–778. [Google Scholar]
- Kepinski, S.; Leyser, O. The Arabidopsis F-box protein TIR1 is an auxin receptor. Nature 2005, 435, 446–451. [Google Scholar]
- Xie, D.X.; Feys, B.F.; James, S.; Nieto-Rostro, M.; Turner, J.G. COI1: An Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 1998, 280, 1091–1094. [Google Scholar]
- Woo, H.R.; Chung, K.M.; Park, J.H.; Oh, S.A.; Ahn, T.; Hong, S.H.; Jang, S.K.; Nam, H.G. ORE9, an F-box protein that regulates leaf senescence in Arabidopsis. Plant Cell 2001, 13, 1779–1790. [Google Scholar]
- Kim, H.S.; Delaney, T.P. Arabidopsis SON1 is an F-box protein that regulates a novel induced defense response independent of both salicylic acid and systemic acquired resistance. Plant Cell 2002, 14, 1469–1482. [Google Scholar]
- Gou, M.Y.; Su, N.; Zheng, J.; Huai, J.L.; Wu, G.H.; Zhao, J.F.; He, J.G.; Tang, D.Z.; Yang, S.H.; Wang, G.Y. An F-box gene, CPY30, functions as a negative regulator of the defense response in Arabidopsis. Plant J 2009, 60, 757–770. [Google Scholar]
- Zhang, Y.; Xu, W.; Li, Z.; Deng, X.W.; Wu, W.; Xue, Y. F-box protein DOR functions as a novel inhibitory factor for abscisic acid-induced stomatal closure under drought stress in Arabidopsis. Plant Physiol 2008, 148, 2121–2133. [Google Scholar]
- Jia, Y.Y.; Gu, H.Y.; Wang, X.S.; Chen, Q.J.; Shi, S.B.; Zhang, J.S.; Ma, L.; Zhang, H.; Ma, H. Molecular cloning and characterization of an F-box family gene CarF-box1 from chichpea (Cicer arietinum L.). Mol. Biol. Rep 2012, 39, 2337–2345. [Google Scholar]
- Yan, Y.S.; Chen, X.Y.; Yang, K.; Sun, Z.X.; Fu, Y.P.; Zhang, Y.M.; Fang, R.X. Overexpression of an F-box protein gene reduces abiotic stress tolerance and promotes root growth in rice. Mol. Plant 2011, 4, 190–197. [Google Scholar]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Li, D.W. Suppression subtractive hybridization analysis of genes regulated by application of exogenous abscisic acid in pepper plant (Capsicum annuum L.) leaves under chilling stress. PLoS One 2013, 8, e66667. [Google Scholar]
- Chai, L.J.; Ge, X.X.; Biswas, M.K.; Deng, X.X. Molecular analysis and expression of a floral organ-relative F-box gene isolated from “Zigui shatian” pummelo (Citrus grandis Osbeck). Mol. Biol. Rep 2011, 38, 4429–2236. [Google Scholar]
- Paquis, S.; Mazeyrat-Gourbeyre, F.; Fernandez, O.; Crouzet, J.; Clément, C.; Baillieul, F.; Dorey, S. Characterization of an F-box gene up-regulated by phytohormones and upon biotic and abiotic stresses in grapevine. Mol. Biol. Rep 2011, 38, 3327–3337. [Google Scholar]
- Choi, D.S.; Hwang, B.K. The pepper extracellular peroxidase CaPO 2 is required for salt, drought and oxidative stress tolerance as well as resistance to fungal pathogens. Planta 2012, 235, 1369–1382. [Google Scholar]
- Lechner, E.; Achard, P.; Vansirc, A.; Potuschak, T.; Genschik, P. F-box proteins everywhere. Curr. Opin. Plant Biol 2006, 9, 631–638. [Google Scholar]
- Nguyen, H.T.; Leipner, J.; Stamp, P.; Guerra-Peraza, O. Low temperature stress in maize (Zea mays L.) induces genes involved in photosynthesis and signal transduction as studied by suppression subtractive hybridization. Plant Physiol. Biochem 2009, 47, 116–122. [Google Scholar]
- Peng, J.; Yu, D.S.; Wang, L.Q.; Xie, M.M.; Yuan, C.Y.; Wang, Y.; Tang, D.Y.; Zhao, X.Y.; Liu, X.M. Arabidopsis F-box gene FOA1 involved in ABA signaling. Sci. China Life Sci 2012, 55, 497–506. [Google Scholar]
- Bray, E.A. Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. J. Exp. Bot 2004, 55, 2331–2341. [Google Scholar]
- Gupta, D.K.; Vandenhove, H.; Inouhe, M. Role of phytochelatins in heavy metal stress and detoxification mechanisms in plants. Heavy Metal Stress Plants 2013, 73–94. [Google Scholar]
- An, S.H.; Sohn, K.H.; Choi, H.W.; Hwang, I.S.; Lee, S.C.; Hwang, B.K. Pepper pectin methylesterase inhibitor protein CaPMEI 1 is required for antifungal activity, basal disease resistance and abiotic stress tolerance. Planta 2008, 228, 61–78. [Google Scholar]
- Lee, S.C.; Hwang, B.K. Functional roles of the pepper antimi-crobial protein gene, CaAMP 1, in abscisic acid signaling and salt and drought tolerance in Arabidopsis. Planta 2009, 229, 383–391. [Google Scholar]
- Choi, D.S.; Hwang, B.K. Proteomics and functional analyses of pepper abscisic acid-responsive 1 (ABR 1), which is involved in cell death and defense signaling. Plant Cell 2011, 23, 823–842. [Google Scholar]
- Gao, Y.; Zhao, Y.; Li, T.T.; Liu, Y.; Ren, C.X.; Wang, M.L. Molecular cloning and expression analysis of an F-box protein gene responsive to plant hormones in Brassica napus. Mol. Biol. Rep 2010, 37, 1037–1044. [Google Scholar]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene CaLOX 1 functions in defense and cell death responses to microbial pathogens. Plant Physiol 2010, 152, 948–967. [Google Scholar]
- Wang, J.E.; Liu, K.K.; Li, D.W.; Zhang, Y.L.; Zhao, Q.; He, Y.M.; Gong, Z.H. A novel peroxidase canpod gene of pepper is involved in defense responses to phytophtora capsici infection as well as abiotic stress tolerance. Int. J. Mol. Sci 2013, 14, 3158–3177. [Google Scholar]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmedand, S.S.; He, Y.M. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper (Capsicum annuum) leaves subjected to chilling stress. Genet. Mol. Res 2012, 11, 4063–4080. [Google Scholar]
- Li, Q.; Yu, B.; Gao, Y.; Dai, A.H. Cinnamic acid pretreatment mitigates chilling stress of cucumber leaves through altering antioxidant enzyme activity. Plant Physiol 2011, 168, 927–934. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. The plant hormone abscisic acid mediates the drought-induced expression but not the seed-specific expression of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana. Mol. Gen. Genet 1993, 238, 17–25. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar]
- Kiyosue, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. ERD15, a cDNA for a dehydration-induced gene from Arabidopsis thaliana. Plant Physiol 1994, 106, 1707. [Google Scholar]
- Bjellqvist, B.; Hughes, G.J.; Pasquali, C.; Paquet, N.; Ravier, F.; Sanchez, J.C.; Frutiger, S.; Hochstrasser, D. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis 1993, 14, 1023–1031. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W-Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 1994, 22, 4673–4680. [Google Scholar]
- Wan, H.J.; Yuan, W.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Zhao, J.; Liu, S.; et al. Identification of reference genes for reverse transcription quantitative real-time PCR normalization in pepper (Capsicum annuum L.). Biochem. Biophys. Res. Commun 2011, 416, 24–30. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot 1981, 32, 93–101. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci 1998, 135, 1–9. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| CaUbi3 | QCaUbi3-F | TGTCCATCTGCTCTCTGTTG |
| QCaUbi3-R | CACCCCAAGCACAATAAGAC | |
| CaF-box | QCaF-box-F | CAAGGTTCAATGTGTGTTACC |
| QCaF-box-R | ATGATGATACAAATACAGTGCC | |
| ERD15 | QERD15-F | CCAGCGAAATGGGGAAAC |
| QERD15-R | ACAAAGGTACAGTGGTGGC | |
| RD22 | QRD22-F | GCGTTGGCAGCGGAAAA |
| QRD22-R | GCGTTAGGATCGTCGTGG | |
| KIN1 | QKIN1-F | AAATGTCAGAGACCAACAAGAA |
| QKIN1-R | CTACTTGTTCAGGCCGGTCTT |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, R.; Guo, W.; Yin, Y.; Gong, Z.-H. A Novel F-Box Protein CaF-Box Is Involved in Responses to Plant Hormones and Abiotic Stress in Pepper (Capsicum annuum L.). Int. J. Mol. Sci. 2014, 15, 2413-2430. https://doi.org/10.3390/ijms15022413
Chen R, Guo W, Yin Y, Gong Z-H. A Novel F-Box Protein CaF-Box Is Involved in Responses to Plant Hormones and Abiotic Stress in Pepper (Capsicum annuum L.). International Journal of Molecular Sciences. 2014; 15(2):2413-2430. https://doi.org/10.3390/ijms15022413
Chicago/Turabian StyleChen, Rugang, Weili Guo, Yanxu Yin, and Zhen-Hui Gong. 2014. "A Novel F-Box Protein CaF-Box Is Involved in Responses to Plant Hormones and Abiotic Stress in Pepper (Capsicum annuum L.)" International Journal of Molecular Sciences 15, no. 2: 2413-2430. https://doi.org/10.3390/ijms15022413