Detoxification of Toxic Phorbol Esters from Malaysian Jatropha curcas Linn. Kernel by Trichoderma spp. and Endophytic Fungi

Abstract
:1. Introduction
2. Results and Discussion
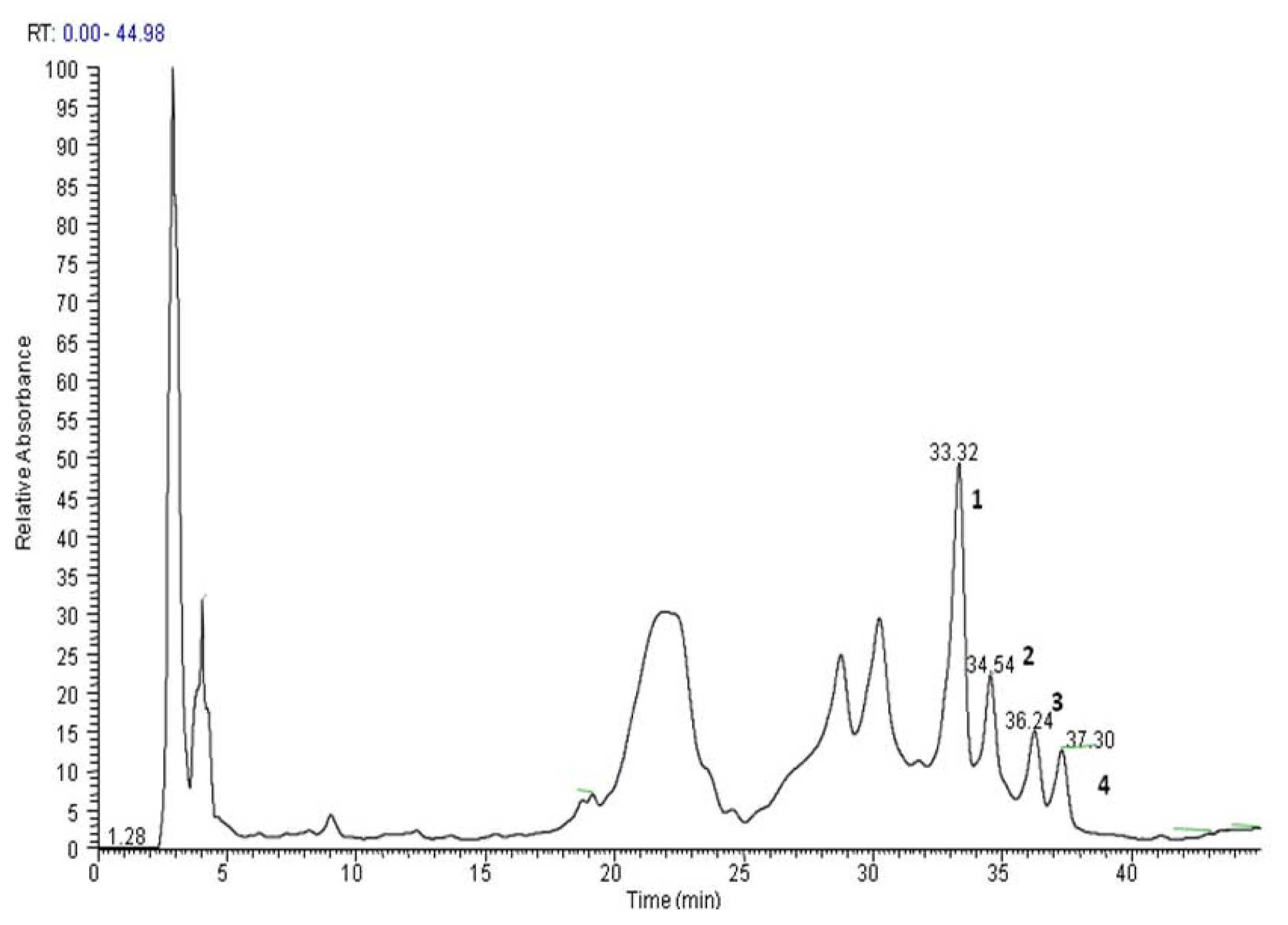
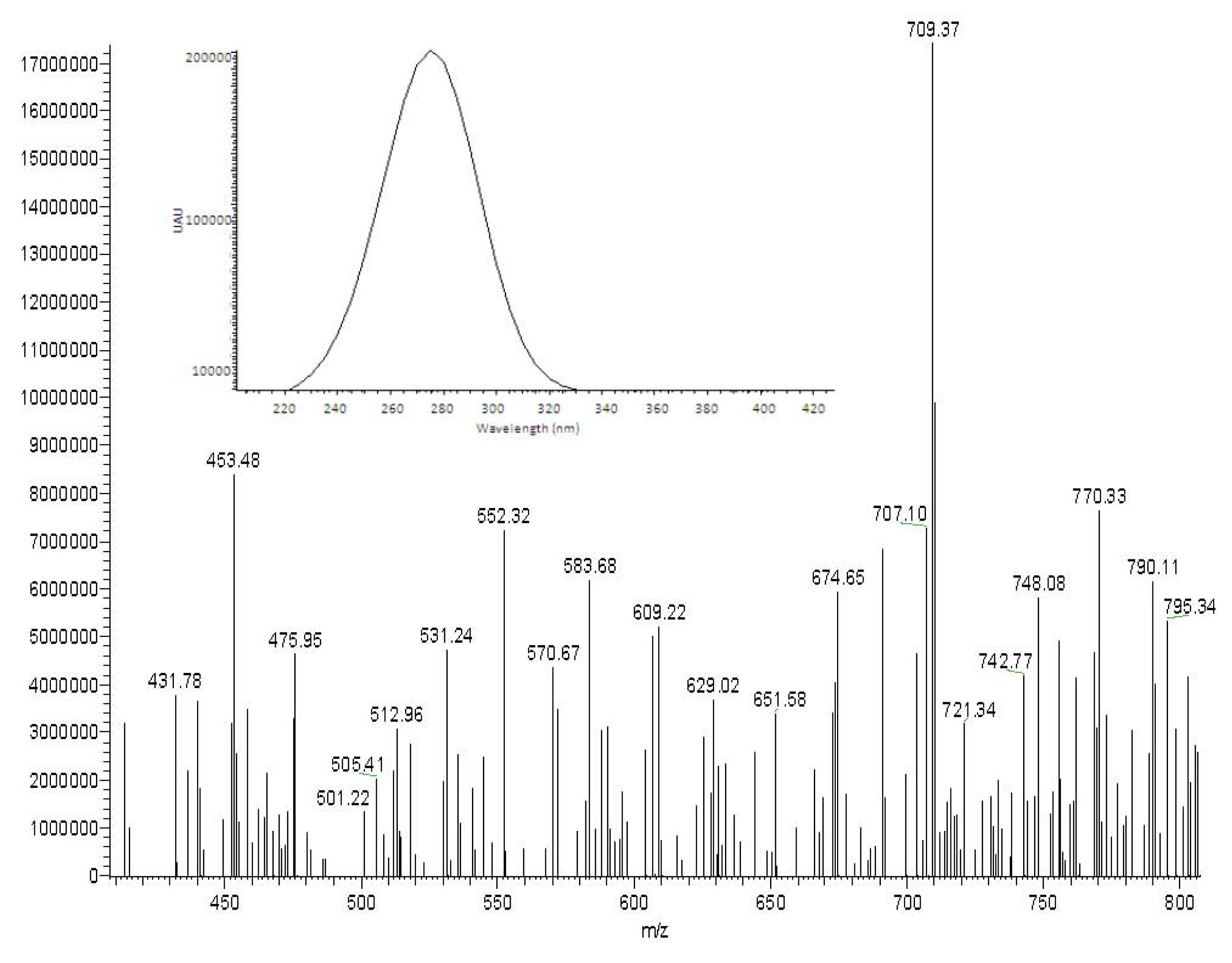
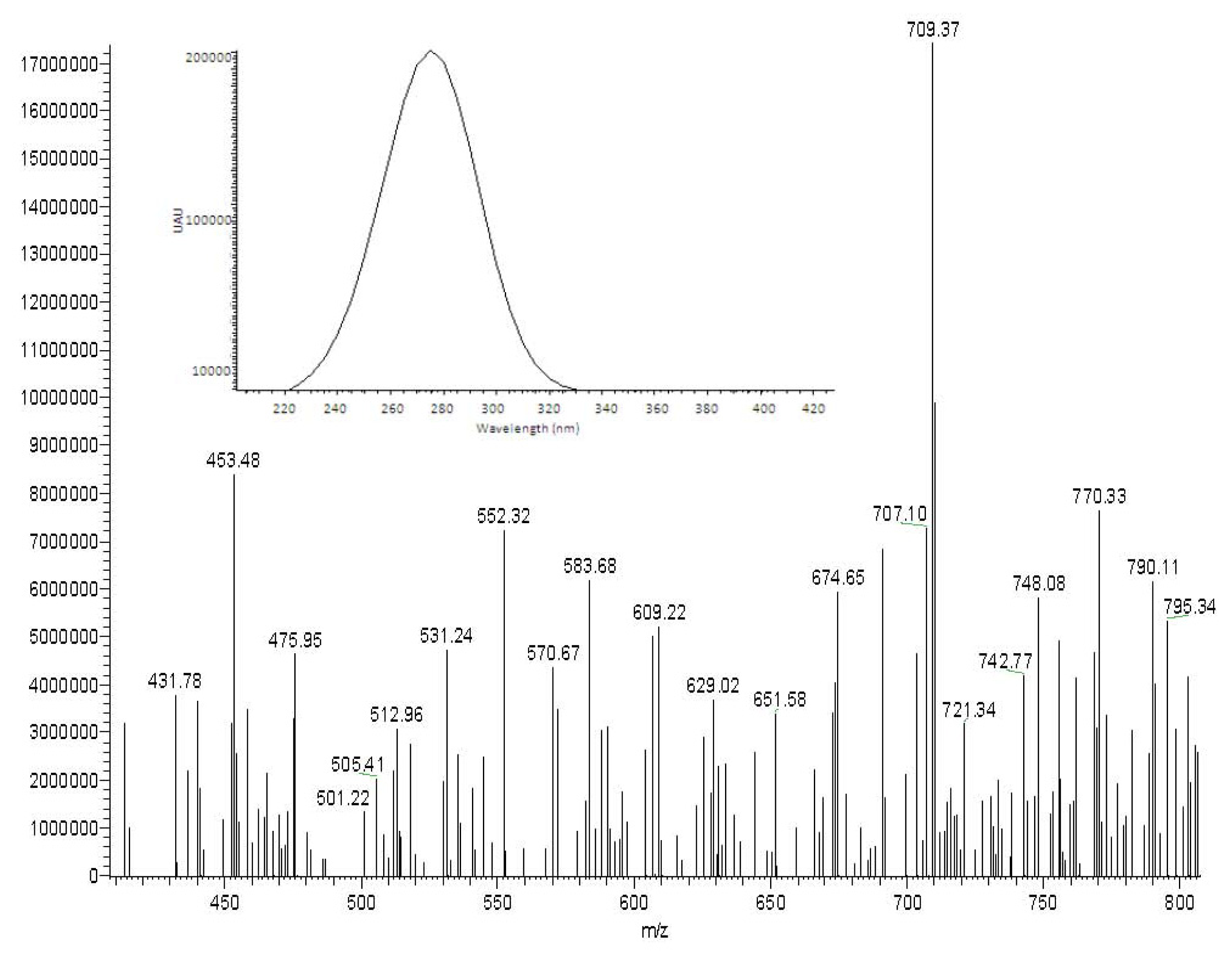
2.1. Detection of PEs-Rich Fraction
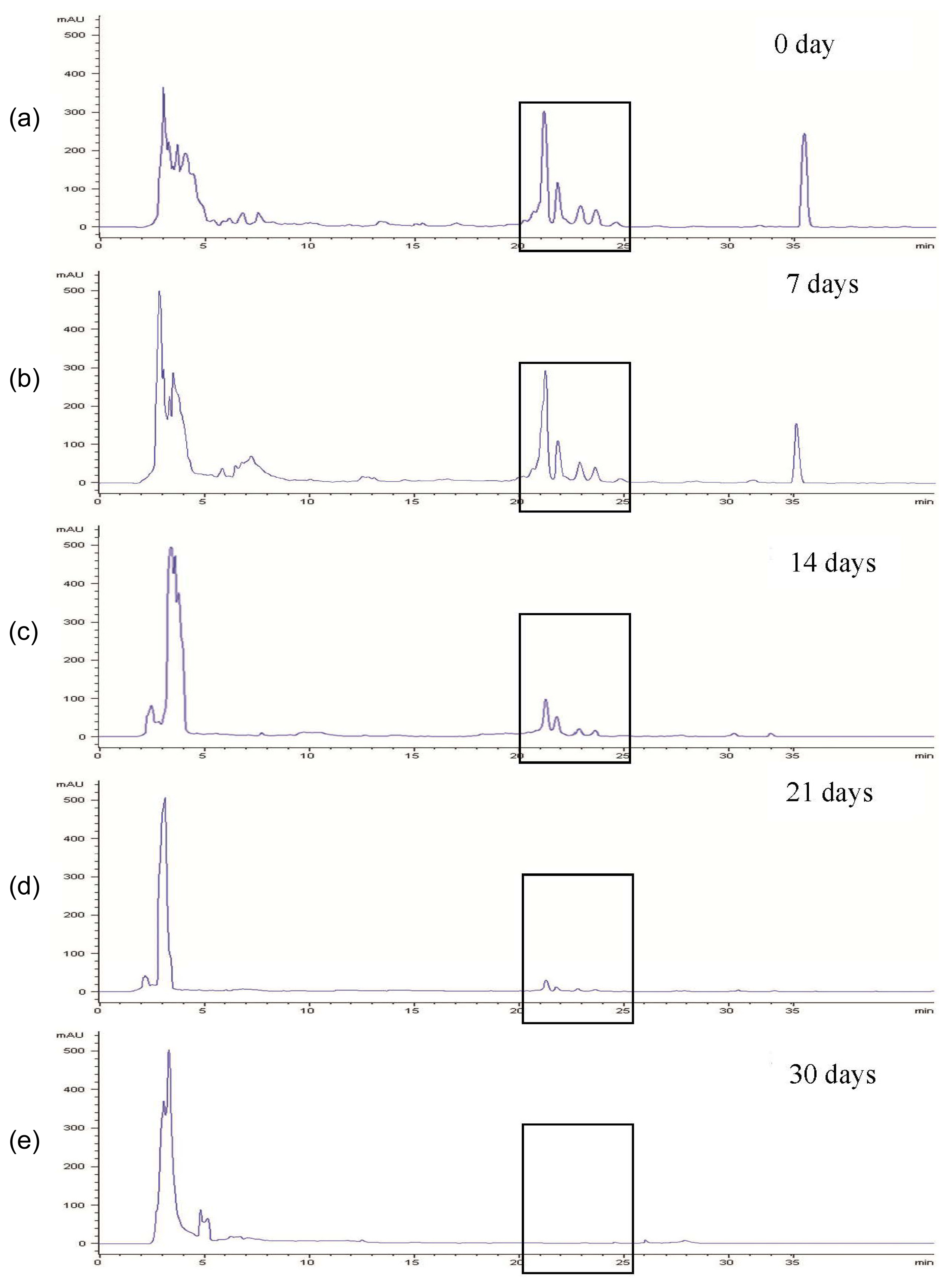
2.2. Phorbol Esters Degradation by Fungal Strains
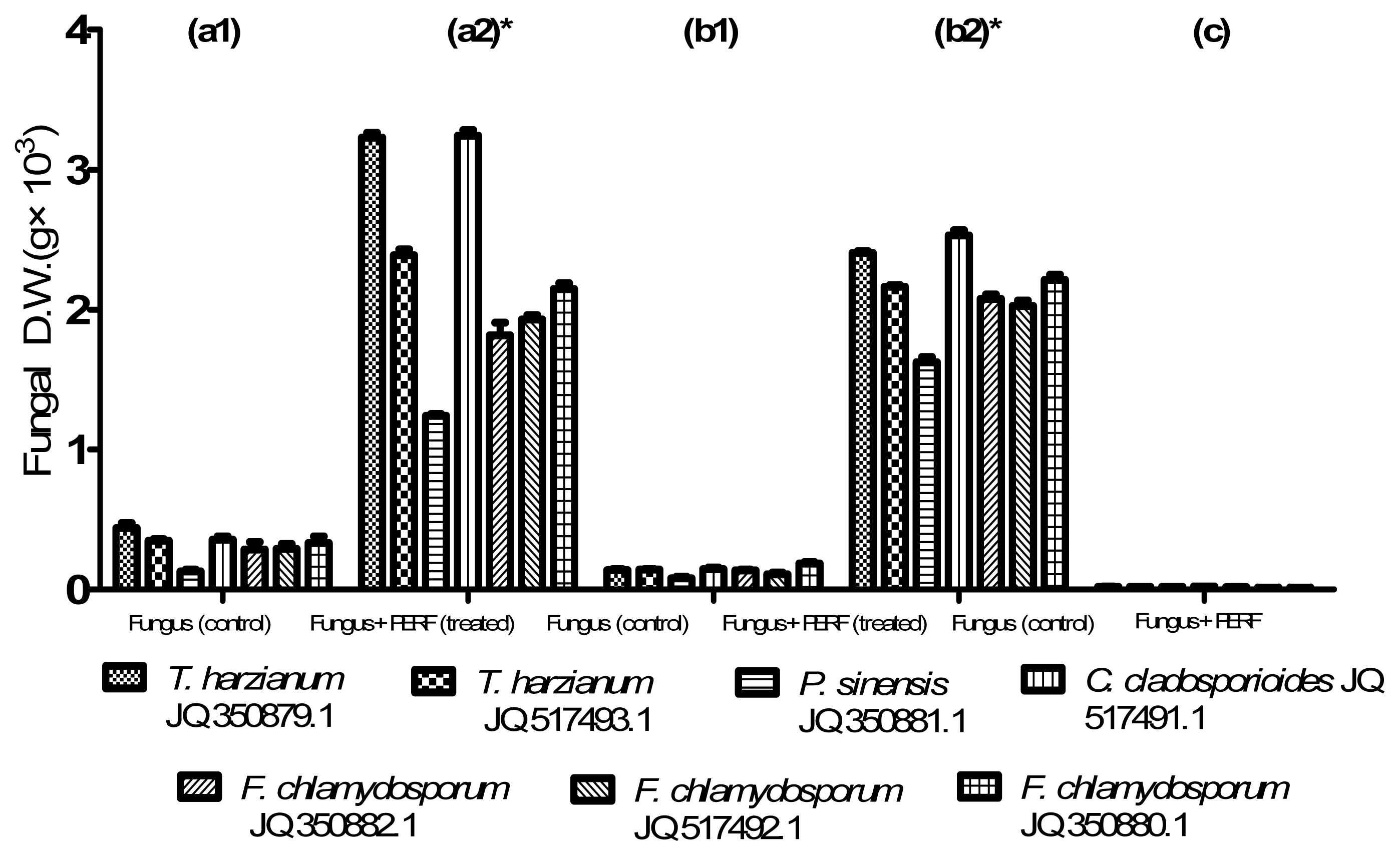
2.3. Utilization of PEs-Rich Fraction as Carbon Source for Fungal Growth
2.4. Cytotoxicity Assay
3. Experimental Section
3.1. Jatropha curcas L. seeds
3.2. Preparation of Phorbol Esters-Rich Fraction
3.3. Detection of PEs
3.4. Microorganism and Inoculum Preparation
3.5. Degradation of PEs
3.6. Utilization of PEs-Rich Fraction as Carbon Source for Fungal Growth
3.7. Cytotoxicity Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Joshi, C.; Mathur, P.; Khare, S. Degradation of phorbol esters by Pseudomonas aeruginosa PseA during solid-state fermentation of deoiled Jatropha curcas seed cake. Bioresour. Technol 2011, 102, 4815–4819. [Google Scholar]
- Oskoueian, E.; Abdullah, N.; Ahmad, S. Phorbol esters isolated from Jatropha meal induced apoptosis-mediated inhibition in proliferation of Chang and Vero cell lines. Int. J. Mol. Sci 2012, 13, 13816–13829. [Google Scholar]
- Ahmed, W.A.; Salimon, J. Phorbol ester as toxic constituents of tropical Jatropha curcas seed oil. Eur. J. Sci. Res 2009, 31, 429–436. [Google Scholar]
- Oskoueian, E.; Abdullah, N.; Saad, W.Z.; Omar, A.R.; Ahmad, S.; Kuan, W.B.; Zolkifli, N.A.; Hendra, R.; Ho, Y.W. Antioxidant, anti-inflammatory and anticancer activities of methanolic extracts from Jatropha curcas Linn. J. Med. Plants Res 2011, 5, 49–57. [Google Scholar]
- Kumar, V.; Makkar, H.P.; Amselgruber, W.; Becker, K. Physiological, haematological and histopathological responses in common carp (Cyprinus carpio L.) fingerlings fed with differently detoxified Jatropha curcas kernel meal. Food Chem. Toxicol 2010, 48, 2063–2072. [Google Scholar]
- De Barros, C.R.M.; Ferreira, L.M.M.; Nunes, F.M.; Bezerra, R.M.F.; Dias, A.A.; Guedes, C.V.; Cone, J.W.; Marques, G.S.M.; Rodrigues, M.A.M. The potential of white-rot fungi to degrade phorbol esters of Jatropha curcas L. seed cake. Eng. Life Sci 2011, 11, 107–110. [Google Scholar]
- Abdel-Hafez, A.A.M.; Nakamura, N.; Hattori, M. Biotransformation of phorbol by human intestinal bacteria. Chem. Pharm. Bull 2002, 50, 160–164. [Google Scholar]
- Jain, A.; Morlok, C.K.; Henson, J.M. Comparison of solid-state and submerged-state fermentation for the bioprocessing of switchgrass to ethanol and acetate by Clostridium phytofermentans. Appl. Microbiol. Biotechnol 2013, 97, 905–917. [Google Scholar]
- Haddadin, M.S.Y.; Haddadin, J.; Arabiyat, O.I.; Hattar, B. Biological conversion of olive pomace into compost by using Trichoderma harzianum and Phanerochaete chrysosporium. Bioresour. Technol 2009, 100, 4773–4782. [Google Scholar]
- Claus, H. Laccases: structure, reactions, distribution. Micron 2004, 35, 93–96. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microb 2004, 2, 43–56. [Google Scholar]
- Yan, X.; Sikora, R.A.; Zheng, J. Potential use of cucumber (Cucumis sativus L.) endophytic fungi as seed treatment agents against root-knot nematode Meloidogyne incognita. J. Zhejiang Uni. Sci 2011, 12, 219–225. [Google Scholar]
- Haas, W.; Sterk, H.; Mittelbach, M. Novel 12-Deoxy-16-Hydroxyphorbol diesters isolated from the seed oil of Jatropha curcas. J. Nat. Prod 2002, 65, 1434–1440. [Google Scholar]
- Hirota, M.; Suttajit, M.; Suguri, H.; Endo, Y.; Shudo, K.; Wongchai, V.; Hecker, E.; Fujiki, H. A new tumor promoter from the seed oil of Jatropha curcas L., an intramolecular diester of 12-Deoxy-16-Hydroxyphorbol. Cancer Res 1988, 48, 5800–5804. [Google Scholar]
- Makkar, H.P.; Kumar, V.; Oyeleye, O.O.; Akinleye, A.O.; Angulo-Escalante, M.A.; Becker, K. Jatropha platyphylla, a new non-toxic Jatropha species: Physical properties and chemical constituents including toxic and antinutritional factors of seeds. Food Chem 2011, 125, 63–71. [Google Scholar]
- Makkar, H.P.S.; Becker, K.; Sporer, F.; Wink, M. Studies on nutritive potential and toxic constituents of different provenances of Jatropha curcas. J. Agric. Food Chem 1997, 45, 3152–3157. [Google Scholar]
- Gopinath, S.C.B.; Anbu, P.; Hilda, A. Extracellular enzymatic activity profiles in fungi isolated from oil-rich environments. Mycoscience 2005, 46, 119–126. [Google Scholar]
- Ward, O.P. Production of recombinant proteins by filamentous fungi. Biotechnol. Adv 2012, 30, 1119–1139. [Google Scholar]
- De Lima Damásio, A.R.; da Silva, T.M.; Maller, A.; Jorge, J.A.; Terenzi, H.F.; Polizeli, M.L. Purification and partial characterization of an exo-polygalacturonase from Paecilomyces variotii liquid cultures. Appl. Biochem. Biotechnol 2010, 160, 1496–1507. [Google Scholar]
- Stamatis, H.; Christakopoulos, P.; Kekos, D.; Macris, B.; Kolisis, F. Studies on the synthesis of short-chain geranyl esters catalysed by Fusarium oxysporum esterase in organic solvents. J. Molec. Catal. B 1998, 4, 229–236. [Google Scholar]
- Belewu, M.; Akande, B. Biological upgrading of the nutritional quality of Jatropha curcas kernel cake: Effect on performance characteristics of goat. Int. Res. J. Biotech 2010, 1, 19–22. [Google Scholar]
- Kurniati, T. Detoxification through fermentation by consortium of Aspergillus niger and Neurospora sitophila towards the degree of forbol esther and nutrition value of Jatropha curcas L. for broilers feed. J. Asian Sci. Res 1998, 2, 317–324. [Google Scholar]
- Gabriel-Ajobiewe, R.A.; Akinyele, B.; Mirrila, E. Basal media formulation using Canavalia ensiformis as carbon and nitrogen source for the growth of some fungi species. J. Microbiol. Biotech. Food Sci 2012, 1, 1136–1151. [Google Scholar]
- Arfi, Y.; Chevret, D.; Henrissat, B.; Berrin, J.; Levasseur, A.; Record, E. Characterization of salt-adapted secreted lignocellulolytic enzymes from the mangrove fungus Pestalotiopsis sp. Nat. Commun 2013, 4, 1810–1838. [Google Scholar]
- Preiss, K.; Adam, I.K.; Gebauer, G. Irradiance governs exploitation of fungi: fine-tuning of carbon gain by two partially myco-heterotrophic Orchids. Proc. R. Soc. B 2010, 277, 1333–1336. [Google Scholar]
- Devappa, R.K.; Angulo-Escalante, M.A.; Makkar, H.P.S.; Becker, K. Potential of using phorbol esters as an insecticide against Spodoptera frugiperda. Ind. Crops Prod 2012, 38, 50–53. [Google Scholar]
- Oskoueian, E.; Abdullah, N.; Ahmad, S. Phorbol esters from Jatropha meal triggered apoptosis, activated PKC-δ, caspase-3 proteins and down-regulated the proto-oncogenes in Mcf-7 and Hela cancer cell lines. Molecules 2012, 17, 10816–10830. [Google Scholar]
- Calvert, T.; Aidoo, K.; Candlish, A.G.G.; Fuat, A.R.M. Comparison of in vitro cytotoxicity of Fusarium mycotoxins, deoxynivalenol, T-2 toxin and zearalenone on selected human epithelial cell lines. Mycopathologia 2005, 159, 413–419. [Google Scholar]
- Flemming, J.; Hudson, B.; Rand, T. Comparison of inflammatory and cytotoxic lung responses in mice after intratracheal exposure to spores of two different Stachybotrys chartarum strains. Toxicol. Sci 2004, 78, 267–275. [Google Scholar]
- Shi, M.; Wang, H.N.; Xie, S.T.; Luo, Y.; Sun, C.Y.; Chen, X.L.; Zhang, Y.Z. Antimicrobial peptaibols, novel suppressors of tumor cells, targeted calcium-mediated apoptosis and autophagy in human hepatocellular carcinoma cells. Mol. Cancer 2010, 9, 26–40. [Google Scholar]
- Toledo Marante, F.J.; Mioso, R.; Bermejo Barrera, J.; González González, J.E.; Santana Rodríguez, J.J.; Bravo de Laguna, I.H. Structural characterization and metabolite profiling of the facultative marine fungus Paecilomyces variotii. Ann. Microbiol 2012, 62, 1601–1607. [Google Scholar]
- Makkar, H.P.S.; Becker, K. Are Jatropha curcas phorbol esters degraded by rumen microbes? J. Sci. Food Agric 2010, 90, 1562–1565. [Google Scholar]
- Moore, J.E.; Xu, J.; Millar, B.; Elshibly, S. Edible dates (Phoenix dactylifera), a potential source of Cladosporium cladosporioides and Sporobolomyces roseus: Implications for public health. Mycopathologia 2002, 154, 25–28. [Google Scholar]
- Najjar, A.A. Determination Mycobiota and Mycotoxins of Air-borne Dust in Some Animal Houses and Computer Laboratories in Jeddah Province. Master’s Thesis, King Abdul-Aziz University, 4 March 2007. [Google Scholar]
- Roberts, W.; Davidson, P. Growth characteristics of selected fungi on polyvinyl chloride film. Appl. Environ. Microbiol 1986, 51, 673–676. [Google Scholar]
- Hammer, E.C.; Nasr, H.; Wallander, H. Effects of different organic materials and mineral nutrients on arbuscular mycorrhizal fungal growth in a mediterranean saline dryland. Soil Biol. Biochem 2011, 7, 76–81. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal strains | Percentage loss of PEs | |||
|---|---|---|---|---|
| 7 days | 14 days | 21 days | 30 days | |
| T. harzianum JQ350879.1 | 28.4 a,v ± 1.44 | 51.0 b,v ± 0.80 | 90.0 c,v ± 0.27 | 99.7 d,v ± 0.01 |
| T. harzianum JQ517493.1 | 23.4 a,v ± 0.25 | 47.6 b,w ± 0.57 | 88.8 c,v ± 0.34 | 99.4 d,v ± 0.05 |
| P. sinensis JQ350881.1 | 24.9 a,v ± 1.96 | 45.3 b,x ± 0.33 | 86.2 c,w ± 0.38 | 98.9 d,v ± 0.02 |
| C. cladosporioides JQ517491.1 | 16.4 a,w ± 0.88 | 42.9 b,y ± 0.20 | 78.0 c,x ± 0.15 | 96.9 d,w ± 0.03 |
| F. chlamydosporum JQ350882.1 | 13.7 a,w ± 1.97 | 33.6 b,z ± 0.27 | 70.1 c,y ± 0.50 | 92.2 d,x ± 0.40 |
| F. chlamydosporum JQ517492.1 | 11.5 a,w ± 1.97 | 34.8 b,z ± 1.23 | 69.8 c,y ± 0.24 | 89.1 d,y ± 0.29 |
| F. chlamydosporum JQ350880.1 | 13.1 a,w ± 1.97 | 33.6 b,z ± 0.27 | 66.9 c,z ± 0.21 | 88.9 d,y ± 0.60 |
| Fungal treated PEs | Cell viability (%) | |
|---|---|---|
| Chang | NIH3T3 | |
| Untreated PEs (control) 1 | 0.4 c,d ± 0.07 | 0.3 c,d ± 0.06 |
| T. harzianum JQ350879.1 | 96.5 a,d ± 0.57 | 93.1 a,d ± 0.64 |
| T. harzianum JQ517493.1 | 90.5 a,b,d ± 0.64 | 90.0 a,b,d ± 1.40 |
| P. sinensis JQ350881.1 | 89.1 b,d± 2.30 | 90.1 a,b,d ± 3.30 |
| C. cladosporioides JQ517491.1 | 89.8 b,d ± 2.38 | 92.5 a,b,d ± 0.56 |
| F. chlamydosporum JQ350882.1 | 88.6 b,d ± 2.29 | 89.2 a,b,d ± 0.71 |
| F. chlamydosporum JQ517492.1 | 84.3 b,d ± 4.42 | 88.0 a,b,d ± 1.53 |
| F. chlamydosporum JQ350880.1 | 86.9 b,d ± 2.41 | 87.4 b,d ± 2.01 |
| Fungal extracts | Cell viability (%) | |
|---|---|---|
| Chang | NIH3T3 | |
| Control 1 | 102.5 ± 0.52 | 103.5 ± 1.20 |
| T. harzianum JQ350879.1 | 99.4 ± 1.17 | 100.2 ± 0.53 |
| T. harzianum JQ517493.1 | 102.4 ± 1.97 | 102.8 ± 2.18 |
| P. sinensis JQ350881.1 | 100.1 ± 2.13 | 103.2 ± 1.70 |
| C. cladosporioides JQ517491.1 | 101.8 ± 1.87 | 100.7 ± 1.82 |
| F. chlamydosporum JQ350882.1 | 98.4 ± 1.23 | 100.2 ± 1.15 |
| F. chlamydosporum JQ517492.1 | 101.9 ± 1.49 | 97.5 ± 1.33 |
| F. chlamydosporum JQ350880.1 | 98.2 ± 1.08 | 101.4 ± 1.46 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Najjar, A.; Abdullah, N.; Saad, W.Z.; Ahmad, S.; Oskoueian, E.; Abas, F.; Gherbawy, Y. Detoxification of Toxic Phorbol Esters from Malaysian Jatropha curcas Linn. Kernel by Trichoderma spp. and Endophytic Fungi. Int. J. Mol. Sci. 2014, 15, 2274-2288. https://doi.org/10.3390/ijms15022274
Najjar A, Abdullah N, Saad WZ, Ahmad S, Oskoueian E, Abas F, Gherbawy Y. Detoxification of Toxic Phorbol Esters from Malaysian Jatropha curcas Linn. Kernel by Trichoderma spp. and Endophytic Fungi. International Journal of Molecular Sciences. 2014; 15(2):2274-2288. https://doi.org/10.3390/ijms15022274
Chicago/Turabian StyleNajjar, Azhar, Norhani Abdullah, Wan Zuhainis Saad, Syahida Ahmad, Ehsan Oskoueian, Faridah Abas, and Youssuf Gherbawy. 2014. "Detoxification of Toxic Phorbol Esters from Malaysian Jatropha curcas Linn. Kernel by Trichoderma spp. and Endophytic Fungi" International Journal of Molecular Sciences 15, no. 2: 2274-2288. https://doi.org/10.3390/ijms15022274
APA StyleNajjar, A., Abdullah, N., Saad, W. Z., Ahmad, S., Oskoueian, E., Abas, F., & Gherbawy, Y. (2014). Detoxification of Toxic Phorbol Esters from Malaysian Jatropha curcas Linn. Kernel by Trichoderma spp. and Endophytic Fungi. International Journal of Molecular Sciences, 15(2), 2274-2288. https://doi.org/10.3390/ijms15022274