Elevated Th22 Cells Correlated with Th17 Cells in Peripheral Blood of Patients with Acute Myeloid Leukemia

Abstract
:1. Introduction
2. Results and Discussion
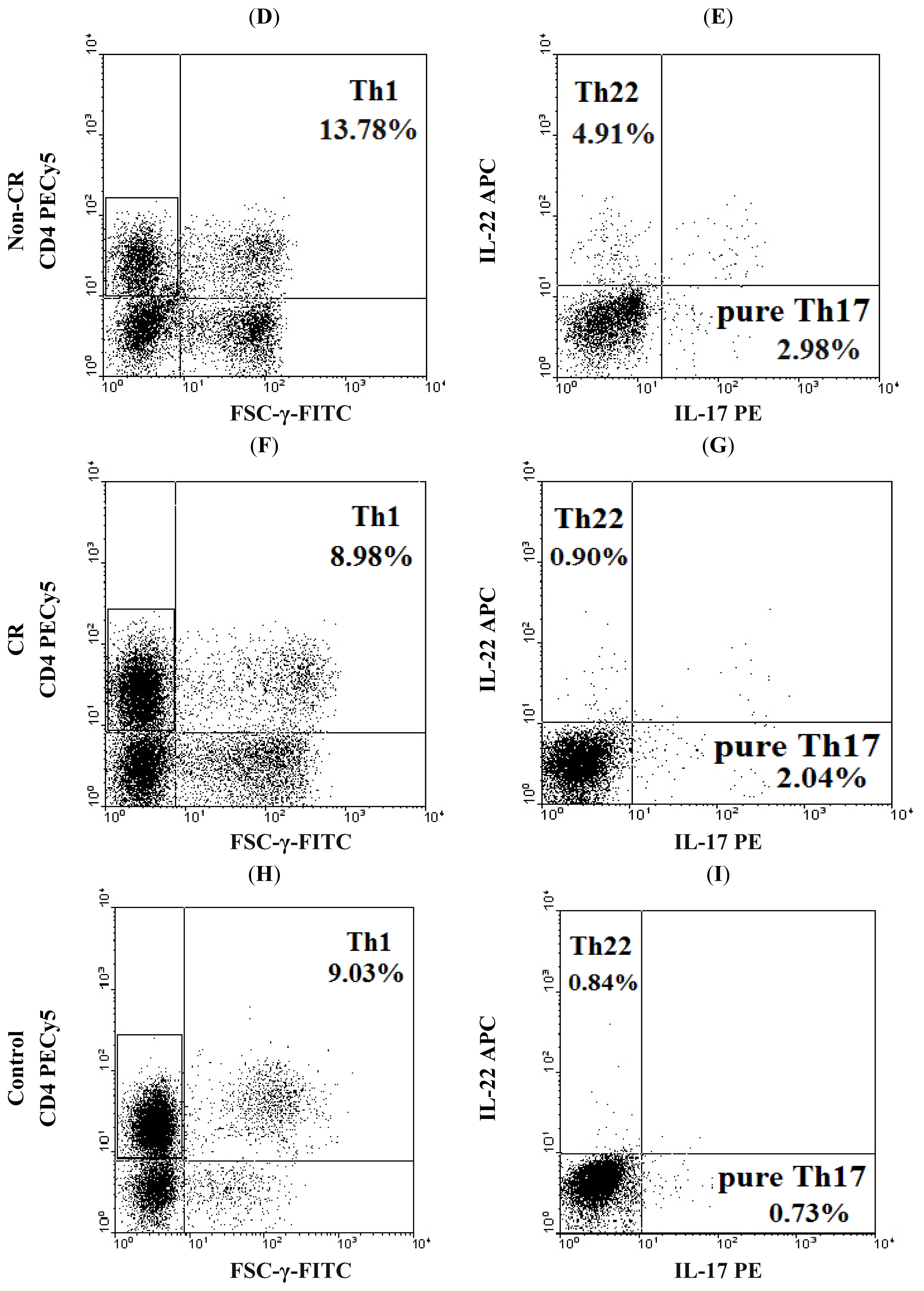
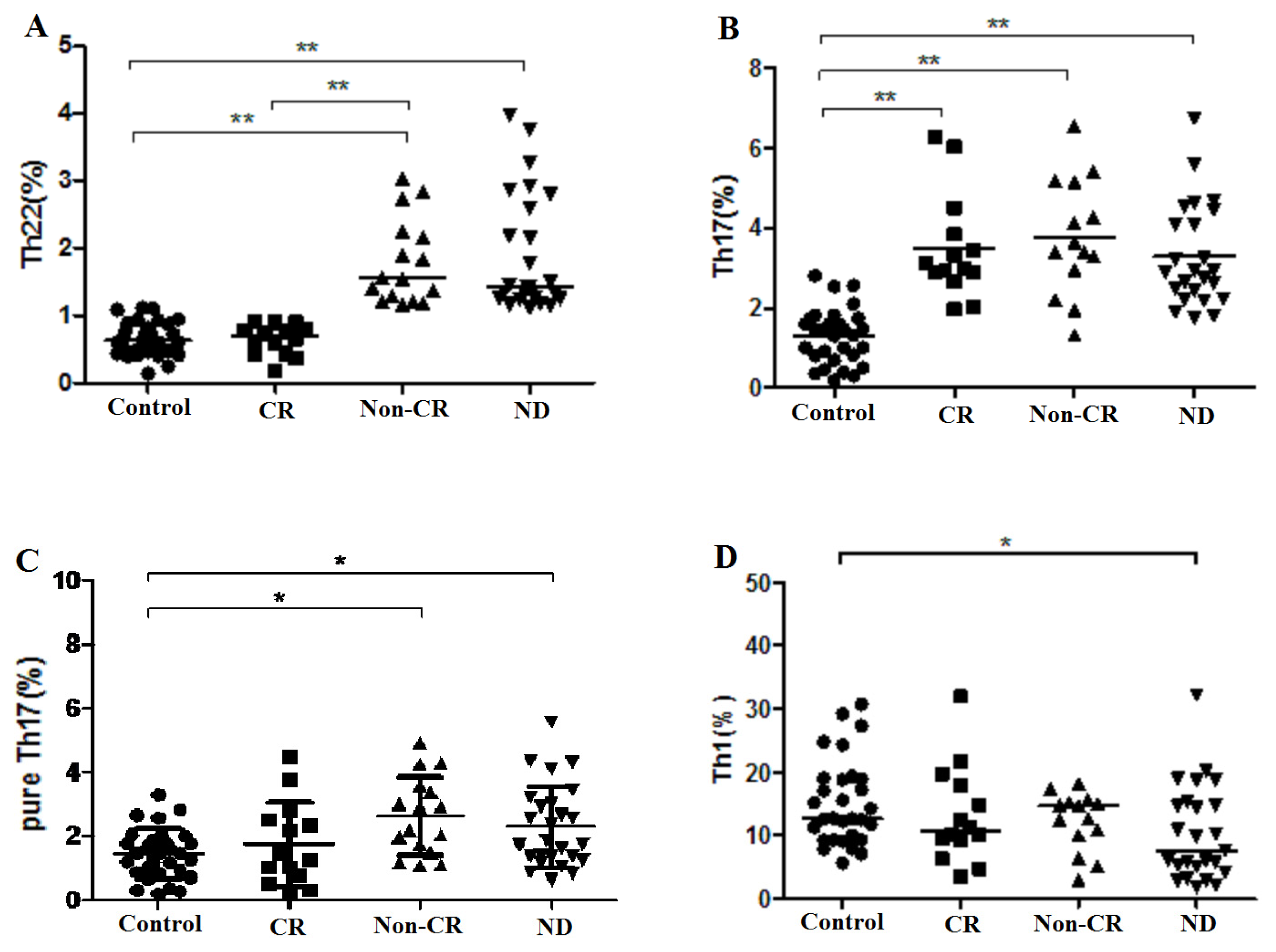
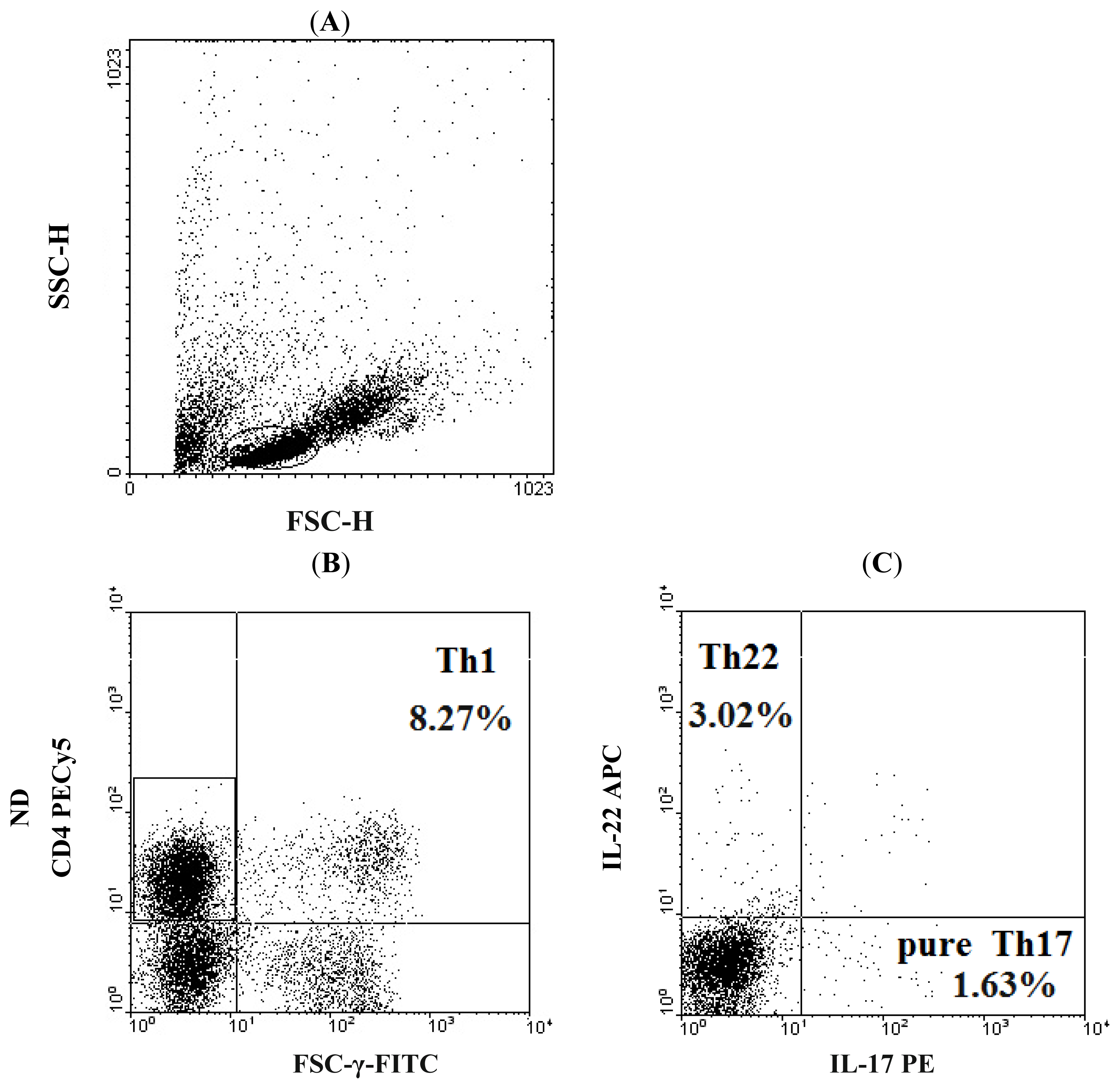
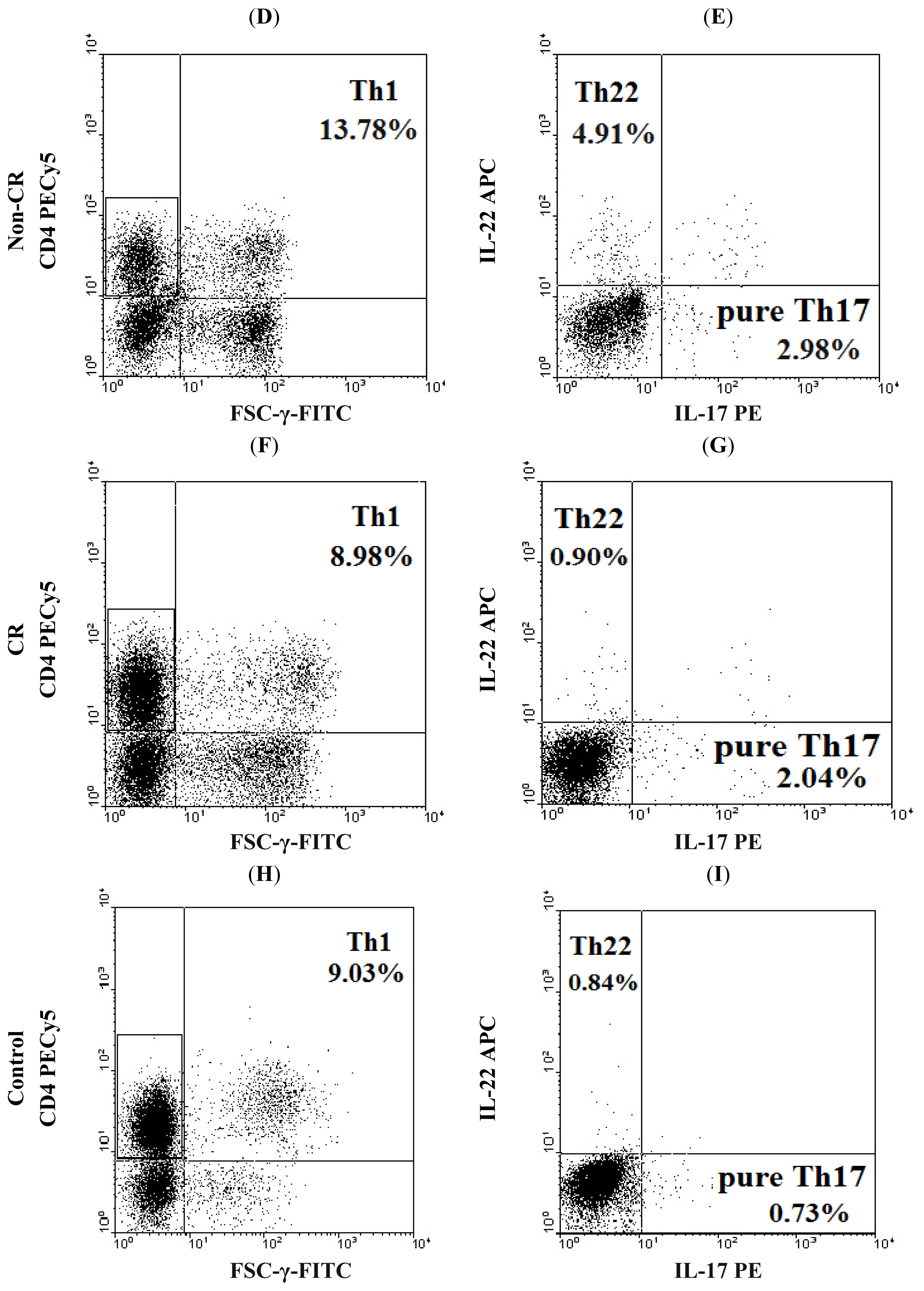
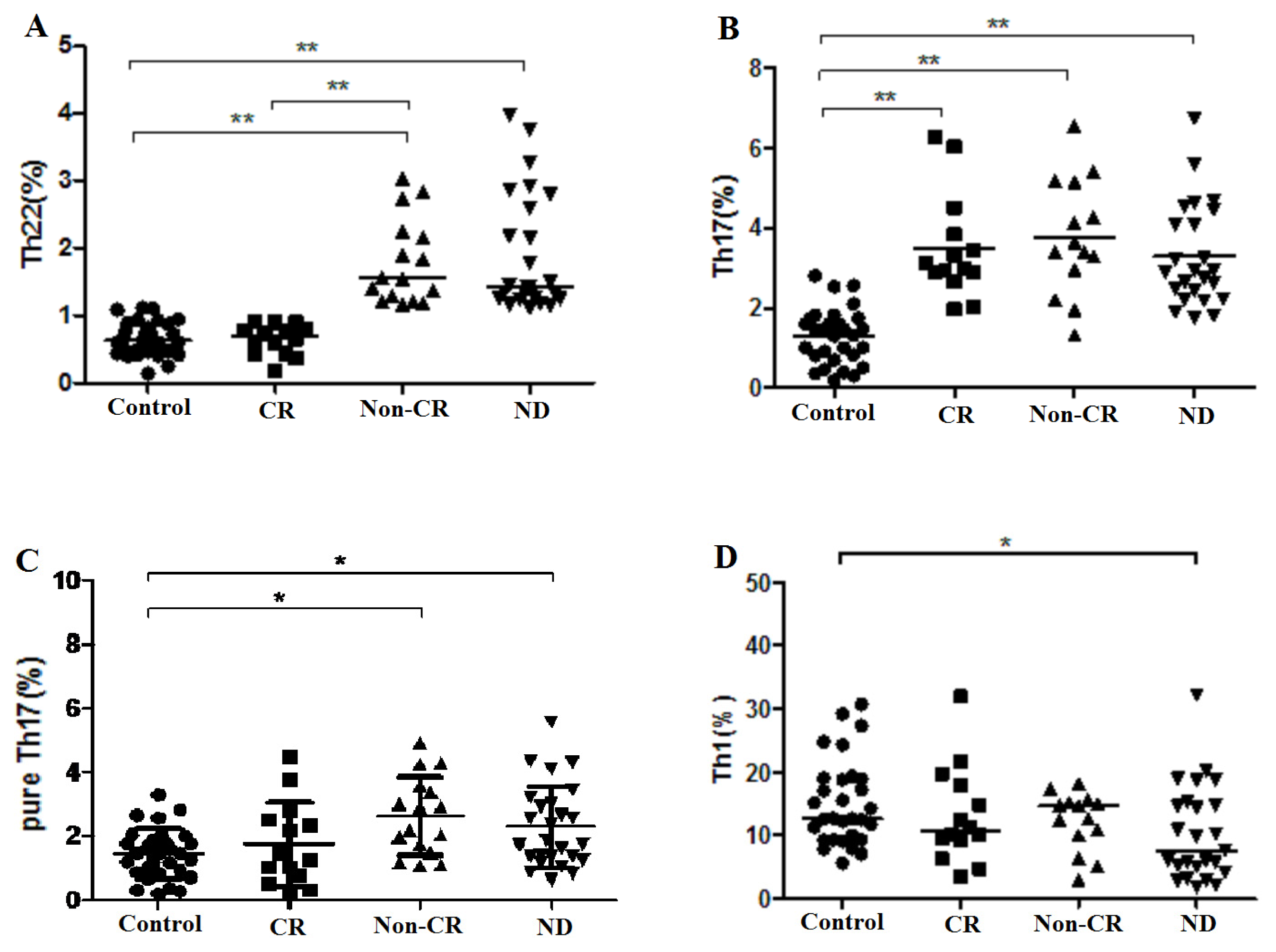
2.1. Elevated Th22 Cells and Plasma IL-22 Level in AML Patients
2.2. Elevated Th17 and Decreased Th1 Cells in AML Patients
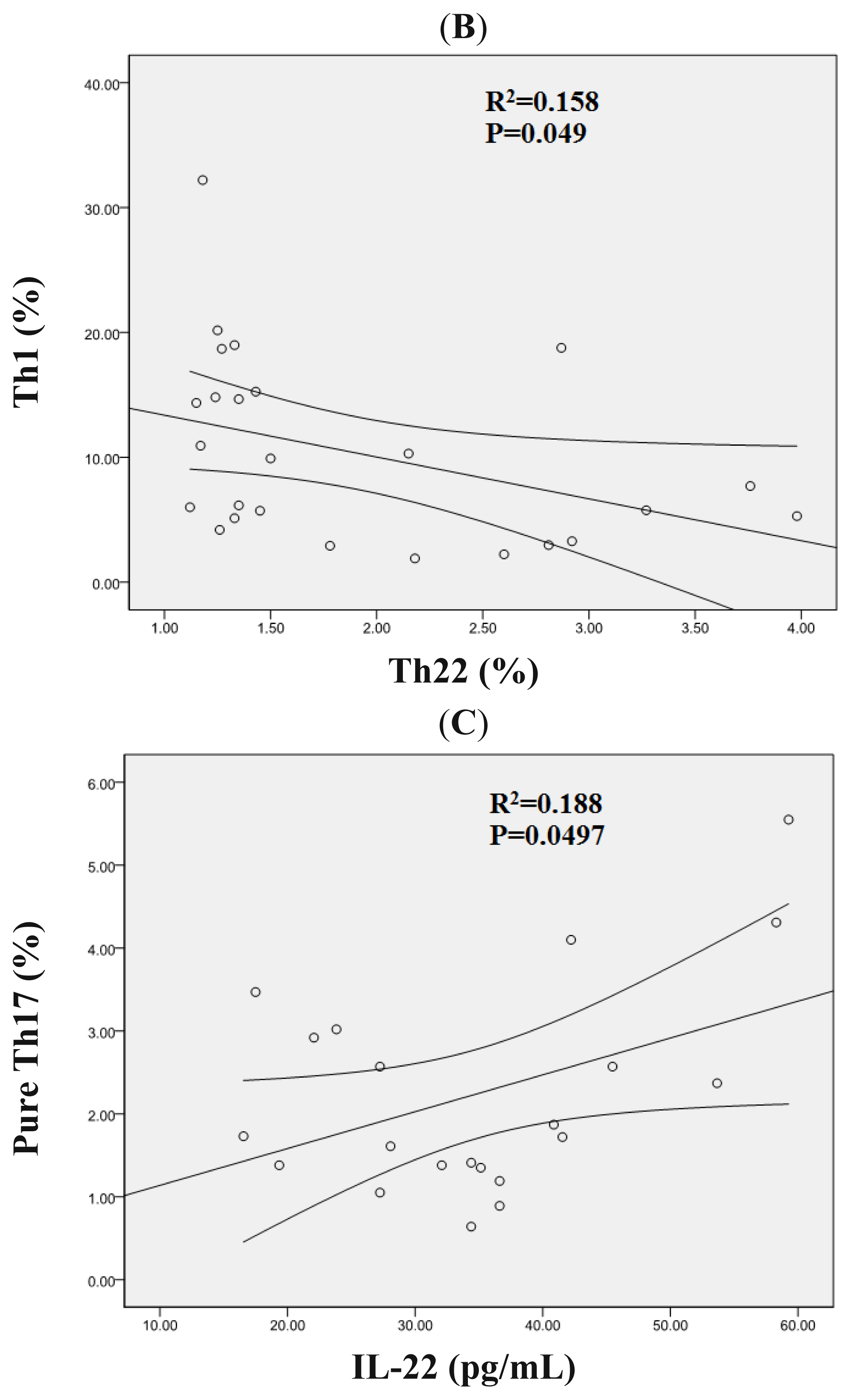
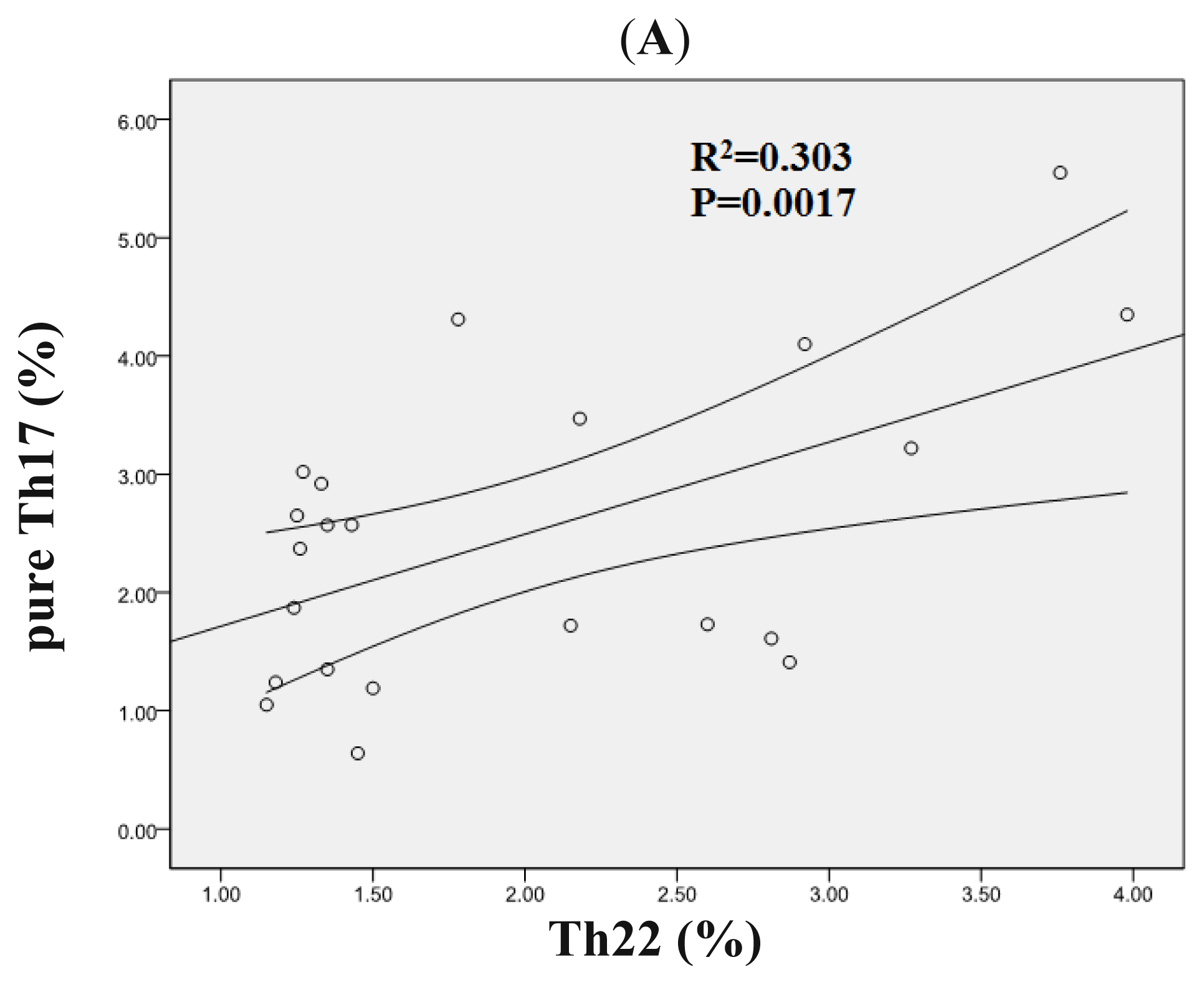
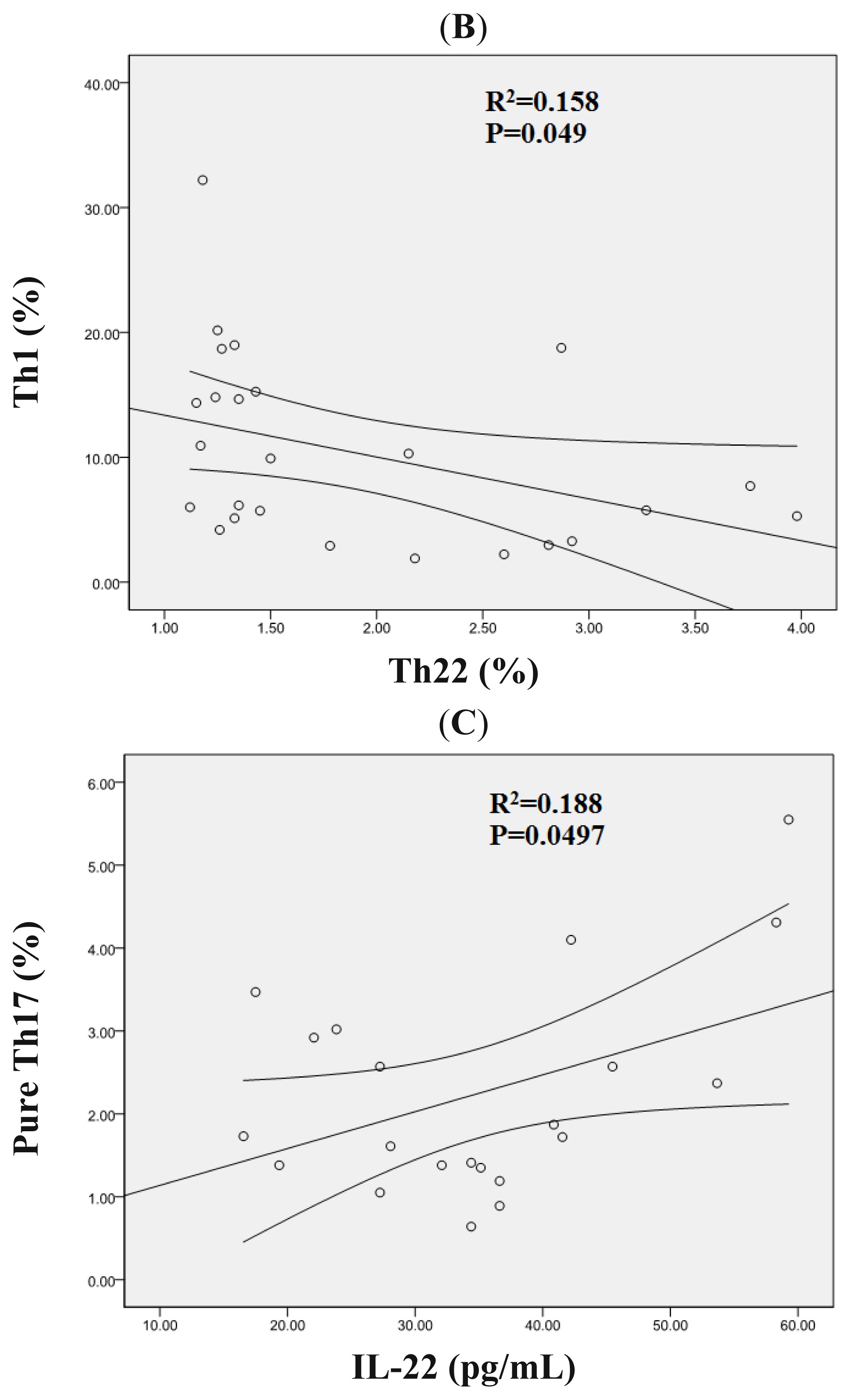
2.3. Correlation Analysis between Th22, Th17, Pure Th17, and Th1 Cells in AML Patients
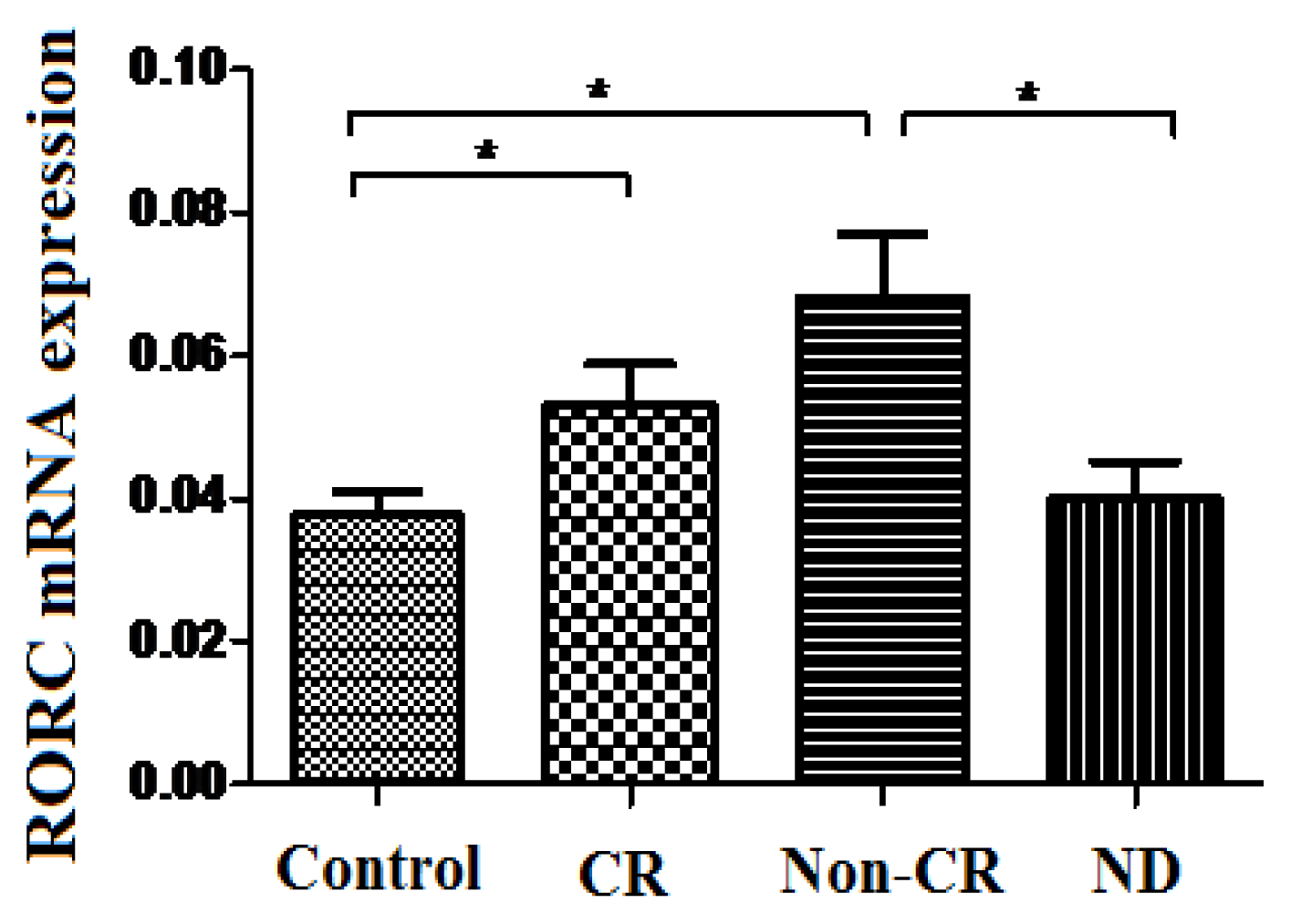
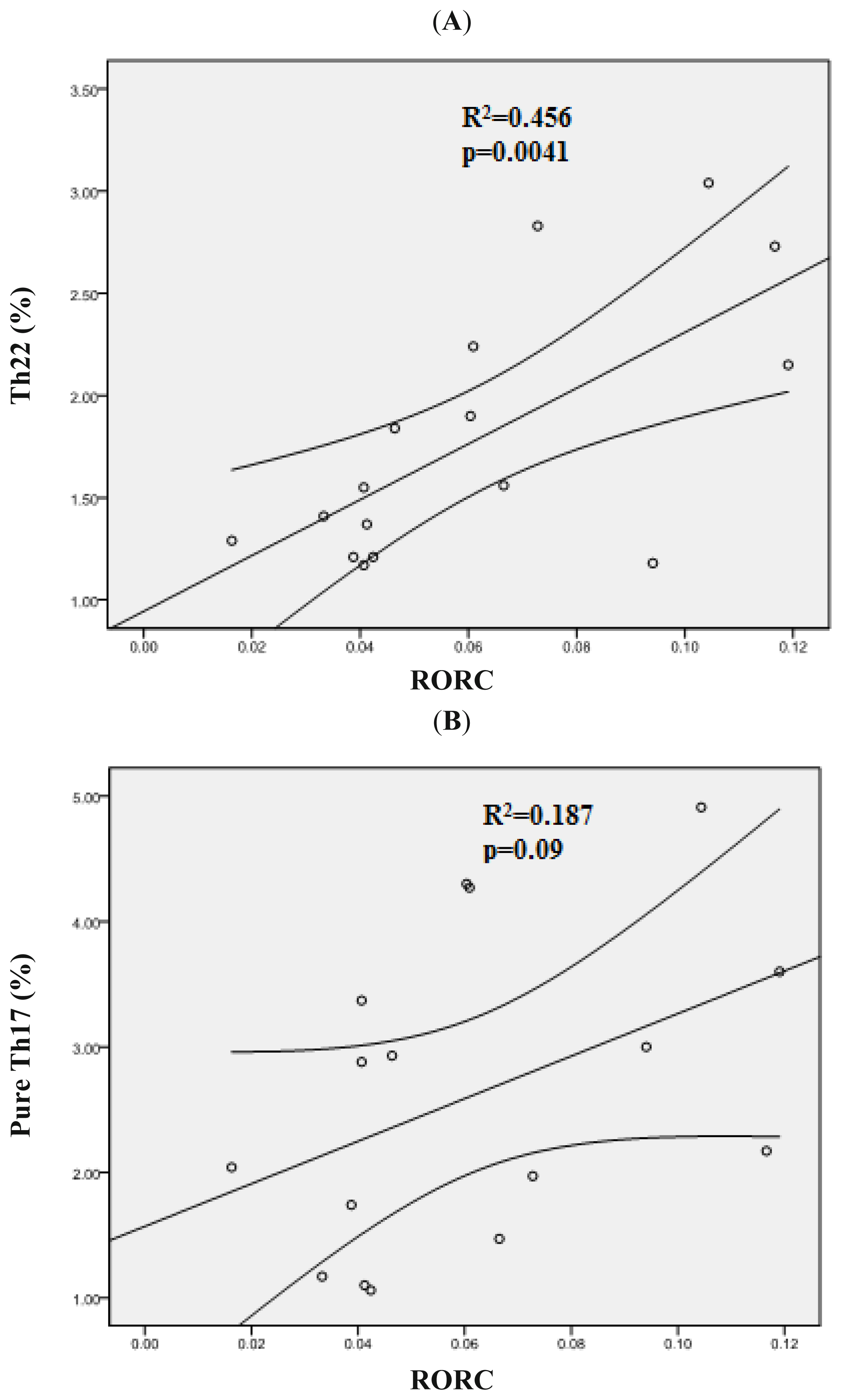
2.4. Increased Expression Profile of RORC in Patients with AML
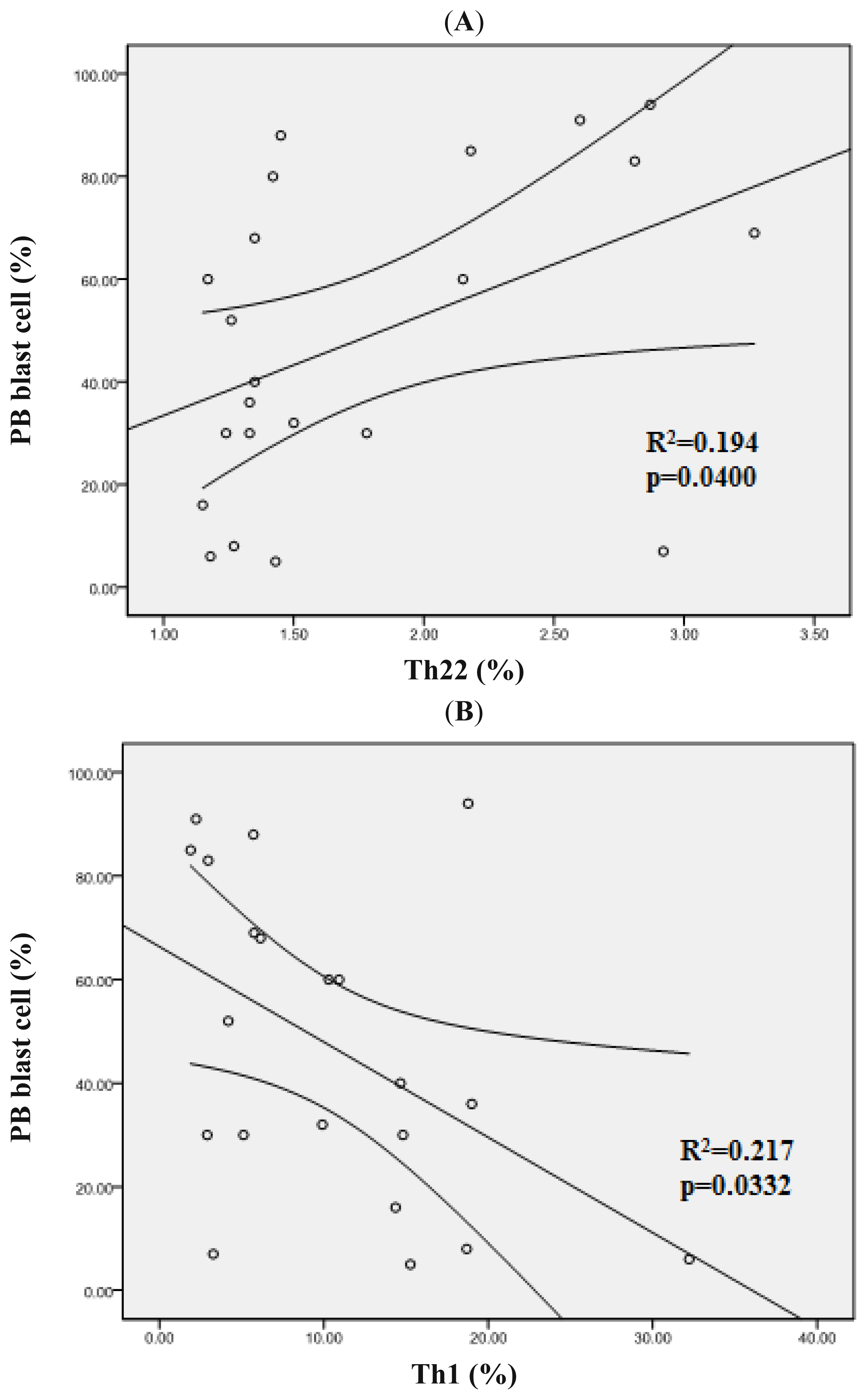
2.5. Clinical Relevance of Th22, Th17, Pure Th17 and Th1 Cells in Patients with AML
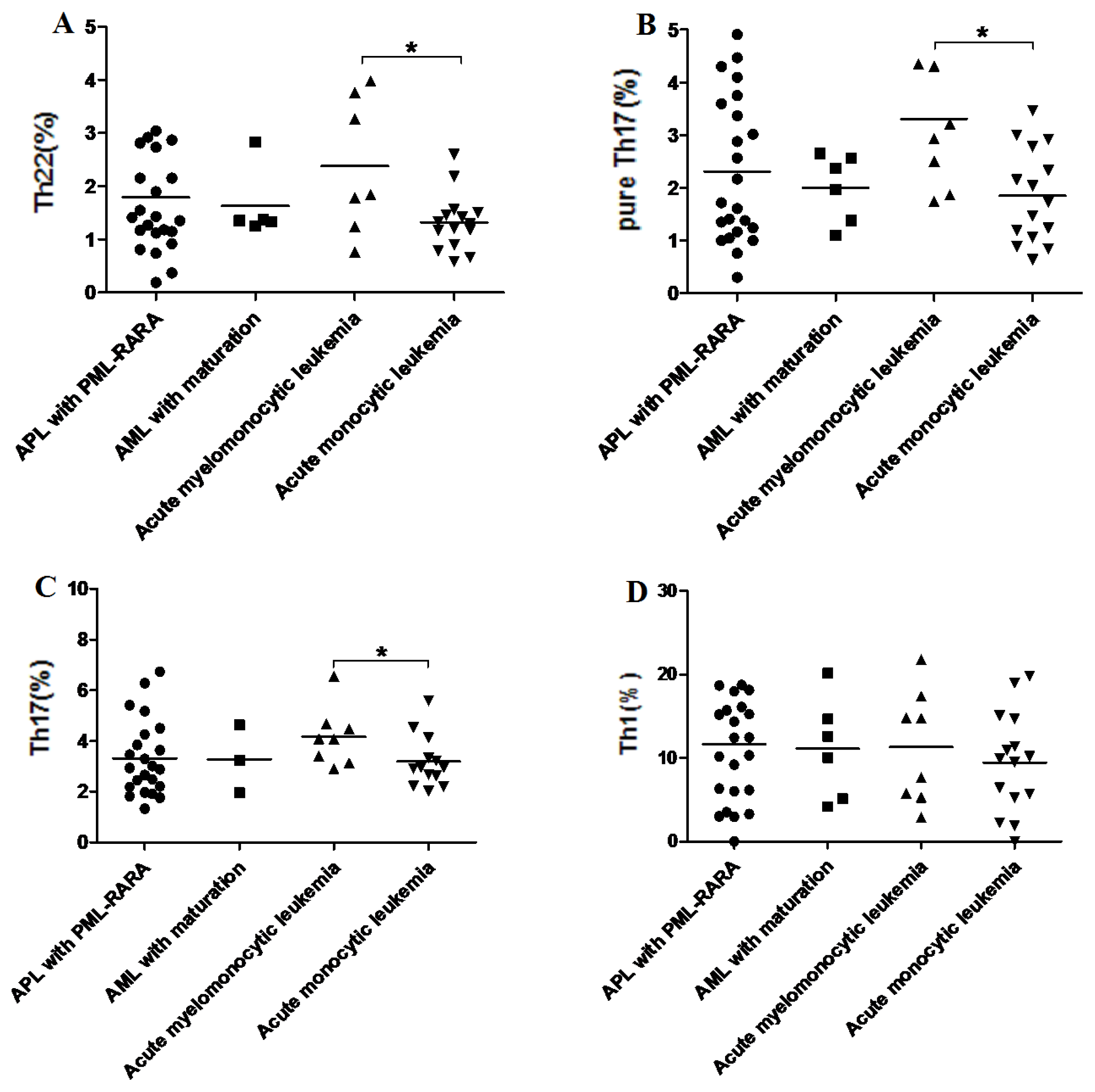
2.6. Increased Th22, Th17, Pure Th17 in Acute Myelomonocytic Leukemia
3. Experimental Section
3.1. Patients and Controls
3.2. Flow Cytometric Analysis
3.3. IL-22 and IL-17 Enzyme-Linked Immunosorbent Assay (ELISA)
3.4. Quantitative Real-Time PCR Analysis
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsS.Y. and D.M. designed and performed research, analyzed data, and wrote the paper; L.Z., B.S. and T.T. performed research; Y.H., L.S. and Y.S. collected samples; C.L. and C.J. designed research.
References
- Ersvaer, E.; Liseth, K.; Skavland, J.; Gjertsen, B.T.; Bruserud, O. Intensive chemotherapy for acute myeloid leukemia differentially affects circulating Tc1, Th1, Th17 and treg cells. BMC Immunol 2010, 11, 38. [Google Scholar]
- Yang, H.Z.; Wang, J.; Sun, Z.M. Intracellular cytokine expression characteristics of activated T lymphocytes in patients with acute myeloid leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2007, 15, 1161–1164. [Google Scholar]
- Wu, C.; Wang, S.; Wang, F.; Chen, Q.; Peng, S.; Zhang, Y.; Qian, J.; Jin, J.; Xu, H. Increased frequencies of T helper type 17 cells in the peripheral blood of patients with acute myeloid leukaemia. Clin. Exp. Immunol 2009, 158, 199–204. [Google Scholar]
- Ferrone, S.; Whiteside, T.L. Tumor microenvironment and immune escape. Surg. Oncol. Clin. N. Am 2007, 16, 755–774. [Google Scholar]
- Whiteside, T.L. Immune suppression in cancer: Effects on immune cells, mechanisms and future therapeutic intervention. Semin. Cancer Biol 2006, 16, 3–15. [Google Scholar]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immun 2005, 6, 1123–1132. [Google Scholar]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor rorgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar]
- Iwakura, Y.; Ishigame, H.; Saijo, S.; Nakae, S. Functional specialization of Interleukin-17 family members. Immunity 2011, 34, 149–162. [Google Scholar]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol 2010, 10, 479–489. [Google Scholar]
- Chae, W.J.; Gibson, T.F.; Zelterman, D.; Hao, L.; Henegariu, O.; Bothwell, A.L. Ablation of IL-17A abrogates progression of spontaneous intestinal tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 5540–5544. [Google Scholar]
- Wang, L.; Yi, T.; Zhang, W.; Pardoll, D.M.; Yu, H. IL-17 enhances tumor development in carcinogen-induced skin cancer. Cancer Res 2010, 70, 10112–10120. [Google Scholar]
- Charles, K.A.; Kulbe, H.; Soper, R.; Escorcio-Correia, M.; Lawrence, T.; Schultheis, A.; Chakravarty, P.; Thompson, R.G.; Kollias, G.; Smyth, J.F.; et al. The tumor-promoting actions of TNF-α involve TNFR1 and IL-17 in ovarian cancer in mice and humans. J. Clin. Invest 2009, 119, 3011–3023. [Google Scholar]
- Numasaki, M.; Fukushi, J.; Ono, M.; Narula, S.K.; Zavodny, P.J.; Kudo, T.; Robbins, P.D.; Tahara, H.; Lotze, M.T. Interleukin-17 promotes angiogenesis and tumor growth. Blood 2003, 101, 2620–2627. [Google Scholar]
- Numasaki, M.; Watanabe, M.; Suzuki, T.; Takahashi, H.; Nakamura, A.; McAllister, F.; Hishinuma, T.; Goto, J.; Lotze, M.T.; Kolls, J.K.; et al. IL-17 enhances the net angiogenic activity and in vivo growth of human non-small cell lung cancer in scid mice through promoting CXCR-2-dependent angiogenesis. J. Immunol 2005, 175, 6177–6189. [Google Scholar]
- Wrobel, T.; Mazur, G.; Jazwiec, B.; Kuliczkowski, K. Interleukin-17 in acute myeloid leukemia. J. Cell. Mol. Med 2003, 7, 472–474. [Google Scholar]
- Fan, Z.G.; Zhang, L.S.; Chai, Y.; Zeng, P.Y.; Wu, C.Y. The prevalence of Th17 cells in patients with acute myeloid leukemia. Zhonghua Xue Ye Xue Za Zhi 2010, 31, 617–620. [Google Scholar]
- Su, M.F.; Wang, C.F.; Zhao, Y.M.; Wu, J.X.; Zhang, Y. Expression and clinical significance of IL-17 and IL-21 in patients with acute leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2010, 18, 1143–1146. [Google Scholar]
- Eyerich, S.; Eyerich, K.; Pennino, D.; Carbone, T.; Nasorri, F.; Pallotta, S.; Cianfarani, F.; Odorisio, T.; Traidl-Hoffmann, C.; Behrendt, H.; et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J. Clin. Invest 2009, 119, 3573–3585. [Google Scholar]
- Duhen, T.; Geiger, R.; Jarrossay, D.; Lanzavecchia, A.; Sallusto, F. Production of interleukin 22 but not interleukin 17 by a subset of human skin-homing memory T cells. Nat. Immun 2009, 10, 857–863. [Google Scholar]
- Liu, Y.; Yang, B.; Zhou, M.; Li, L.; Zhou, H.; Zhang, J.; Chen, H.; Wu, C. Memory IL-22-producing CD4+ T cells specific for candida albicans are present in humans. Eur. J. Immunol 2009, 39, 1472–1479. [Google Scholar]
- Trifari, S.; Kaplan, C.D.; Tran, E.H.; Crellin, N.K.; Spits, H. Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from Th-17, Th1 and Th2 cells. Nat. Immun 2009, 10, 864–871. [Google Scholar]
- Nograles, K.E.; Zaba, L.C.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Kikuchi, T.; Ramon, M.; Bergman, R.; Krueger, J.G.; Guttman-Yassky, E. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing Th17 T cells. J. Allergy Clin. Immunol 2009, 123, 1244–1252. [Google Scholar]
- Annunziato, F.; Cosmi, L.; Santarlasci, V.; Maggi, L.; Liotta, F.; Mazzinghi, B.; Parente, E.; Fili, L.; Ferri, S.; Frosali, F.; et al. Phenotypic and functional features of human Th17 cells. J. Exp. Med 2007, 204, 1849–1861. [Google Scholar]
- Wolk, K.; Witte, E.; Wallace, E.; Docke, W.D.; Kunz, S.; Asadullah, K.; Volk, H.D.; Sterry, W.; Sabat, R. IL-22 regulates the expression of genes responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: A potential role in psoriasis. Eur. J. Immunol 2006, 36, 1309–1323. [Google Scholar]
- Zhang, N.; Pan, H.F.; Ye, D.Q. Th22 in inflammatory and autoimmune disease: Prospects for therapeutic intervention. Mol. Cell. Biochem 2011, 353, 41–46. [Google Scholar]
- Dumoutier, L.; Louahed, J.; Renauld, J.C. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. J. Immunol 2000, 164, 1814–1819. [Google Scholar]
- Dumoutier, L.; Van Roost, E.; Colau, D.; Renauld, J.C. Human interleukin-10-related T cell-derived inducible factor: Molecular cloning and functional characterization as an hepatocyte-stimulating factor. Proc. Natl. Acad. Sci. USA 2000, 97, 10144–10149. [Google Scholar]
- Cheng, F.; Guo, Z.; Xu, H.; Yan, D.; Li, Q. Decreased plasma il22 levels, but not increased IL17 and IL23 levels, correlate with disease activity in patients with systemic lupus erythematosus. Ann. Rheum. Dis 2009, 68, 604–606. [Google Scholar]
- Nickoloff, B.J. Cracking the cytokine code in psoriasis. Nat. Med 2007, 13, 242–244. [Google Scholar]
- Brand, S.; Beigel, F.; Olszak, T.; Zitzmann, K.; Eichhorst, S.T.; Otte, J.M.; Diepolder, H.; Marquardt, A.; Jagla, W.; Popp, A.; et al. IL-22 is increased in active Crohn’s disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am. J. Physiol 2006, 290, G827–G838. [Google Scholar]
- Shao, L.L.; Zhang, L.; Hou, Y.; Yu, S.; Liu, X.G.; Huang, X.Y.; Sun, Y.X.; Tian, T.; He, N.; Ma, D.X.; et al. Th22 cells as well as Th17 cells expand differentially in patients with early-stage and late-stage myelodysplastic syndrome. PLoS One 2012, 7, e51339. [Google Scholar]
- Tian, T.; Sun, Y.; Li, M.; He, N.; Yuan, C.; Yu, S.; Wang, M.; Ji, C.; Ma, D. Increased Th22 cells as well as Th17 cells in patients with adult T-cell acute lymphoblastic leukemia. Clin. Chim. Acta 2013, 426, 108–113. [Google Scholar]
- Shi, L.; Chen, S.; Yang, L.; Li, Y. The role of PD-1 and PD-l1 in T-cell immune suppression in patients with hematological malignancies. J. Hematol. Oncol 2013, 6, 74. [Google Scholar]
- Zha, X.; Yan, X.; Shen, Q.; Zhang, Y.; Wu, X.; Chen, S.; Li, B.; Yang, L.; Geng, S.; Weng, J.; et al. Alternative expression of TCRζ related genes in patients with chronic myeloid leukemia. J. Hematol. Oncol 2012, 5, 74. [Google Scholar]
- Liu, L.M.; Zhang, X.X.; Zhang, Y.M. The change of peripheral blood Th22 cells in patients with acute myeloid leukemia. Zhonghua Xue Ye Xue Za Zhi 2012, 33, 133–135. [Google Scholar]
- Wolk, K.; Witte, E.; Witte, K.; Warszawska, K.; Sabat, R. Biology of interleukin-22. Semin. Immunopathol 2010, 32, 17–31. [Google Scholar]
- Ikeuchi, H.; Kuroiwa, T.; Hiramatsu, N.; Kaneko, Y.; Hiromura, K.; Ueki, K.; Nojima, Y. Expression of interleukin-22 in rheumatoid arthritis: Potential role as a proinflammatory cytokine. Arthritis Rheum 2005, 52, 1037–1046. [Google Scholar]
- Pan, H.F.; Zhao, X.F.; Yuan, H.; Zhang, W.H.; Li, X.P.; Wang, G.H.; Wu, G.C.; Tang, X.W.; Li, W.X.; Li, L.H.; et al. Decreased serum IL-22 levels in patients with systemic lupus erythematosus. Clin. Chim. Acta 2009, 401, 179–180. [Google Scholar]
- Kryczek, I.; Wei, S.; Zou, L.; Altuwaijri, S.; Szeliga, W.; Kolls, J.; Chang, A.; Zou, W. Cutting edge: Th17 and regulatory T cell dynamics and the regulation by IL-2 in the tumor microenvironment. J. Immunol 2007, 178, 6730–6733. [Google Scholar]
- Zhang, B.; Rong, G.; Wei, H.; Zhang, M.; Bi, J.; Ma, L.; Xue, X.; Wei, G.; Liu, X.; Fang, G. The prevalence of Th17 cells in patients with gastric cancer. Biochem. Biophys. Res. Commun 2008, 374, 533–537. [Google Scholar]
- Wang, K.; Wei, G.; Liu, D. CD19: A biomarker for B cell development, lymphoma diagnosis and therapy. Exp. Hematol. Oncol 2012, 1, 36. [Google Scholar]
- Huang, X.; Du, X.; Li, Y. The role of BCL11B in hematological malignancy. Exp. Hematol. Oncol 2012, 1, 22. [Google Scholar]
- Vardiman, J.W.; Thiele, J.; Arber, D.A.; Brunning, R.D.; Borowitz, M.J.; Porwit, A.; Harris, N.L.; Le Beau, M.M.; Hellstrom-Lindberg, E.; Tefferi, A.; et al. The 2008 revision of the world health organization (WHO) classification of myeloid neoplasms and acute leukemia: Rationale and important changes. Blood 2009, 114, 937–951. [Google Scholar]
- Cheson, B.D.; Bennett, J.M.; Kopecky, K.J.; Buchner, T.; Willman, C.L.; Estey, E.H.; Schiffer, C.A.; Doehner, H.; Tallman, M.S.; Lister, T.A.; et al. Revised recommendations of the international working group for diagnosis, standardization of response criteria, treatment outcomes, and reporting standards for therapeutic trials in acute myeloid leukemia. J. Clin. Oncol 2003, 21, 4642–4649. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND AML patients (n = 26) | Non-CR AML patients (n = 16) | CR AML patients (n = 14) | |
|---|---|---|---|
| Age (year) | 39 (19–77) | 43 (15–74) | 37 (13–57) |
| Gender (male/female) | 11/15 | 11/5 | 5/9 |
| PB blast cell (%) | 48.64 ± 30.61 | 34 ± 34.53 | 0 |
| BM blast cell (%) | 76.25 ± 17.78 | 47.33 ± 28.52 | 0.5 (0–2) |
| WHO classification | |||
| APL with t (15; 17) (q22; q12); PML-RARA | 11 | 5 | 7 |
| AML with t (8; 21) (q22; q22) | 2 | 0 | 0 |
| AML, not otherwise specified | |||
| AML without maturation | 0 | 1 | 0 |
| AML with maturation | 4 | 2 | 0 |
| Acute myelomonocytic leukemia | 5 | 2 | 1 |
| Acute monocytic leukemia | 7 | 4 | 4 |
| Acute erythroid leukemia | 0 | 1 | 0 |
| Treatment regimens | Number of samples |
|---|---|
| ATRA | 4 |
| ATRA and DNR | 2 |
| HA | 1 |
| IA | 1 |
| Multiple cycles of chemotherapy | 22 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yu, S.; Liu, C.; Zhang, L.; Shan, B.; Tian, T.; Hu, Y.; Shao, L.; Sun, Y.; Ji, C.; Ma, D. Elevated Th22 Cells Correlated with Th17 Cells in Peripheral Blood of Patients with Acute Myeloid Leukemia. Int. J. Mol. Sci. 2014, 15, 1927-1945. https://doi.org/10.3390/ijms15021927
Yu S, Liu C, Zhang L, Shan B, Tian T, Hu Y, Shao L, Sun Y, Ji C, Ma D. Elevated Th22 Cells Correlated with Th17 Cells in Peripheral Blood of Patients with Acute Myeloid Leukemia. International Journal of Molecular Sciences. 2014; 15(2):1927-1945. https://doi.org/10.3390/ijms15021927
Chicago/Turabian StyleYu, Shuang, Chuanfang Liu, Lei Zhang, Baozhong Shan, Tian Tian, Yu Hu, Linlin Shao, Yuanxin Sun, Chunyan Ji, and Daoxin Ma. 2014. "Elevated Th22 Cells Correlated with Th17 Cells in Peripheral Blood of Patients with Acute Myeloid Leukemia" International Journal of Molecular Sciences 15, no. 2: 1927-1945. https://doi.org/10.3390/ijms15021927