Down-Regulation by Resveratrol of Basic Fibroblast Growth Factor-Stimulated Osteoprotegerin Synthesis through Suppression of Akt in Osteoblasts

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
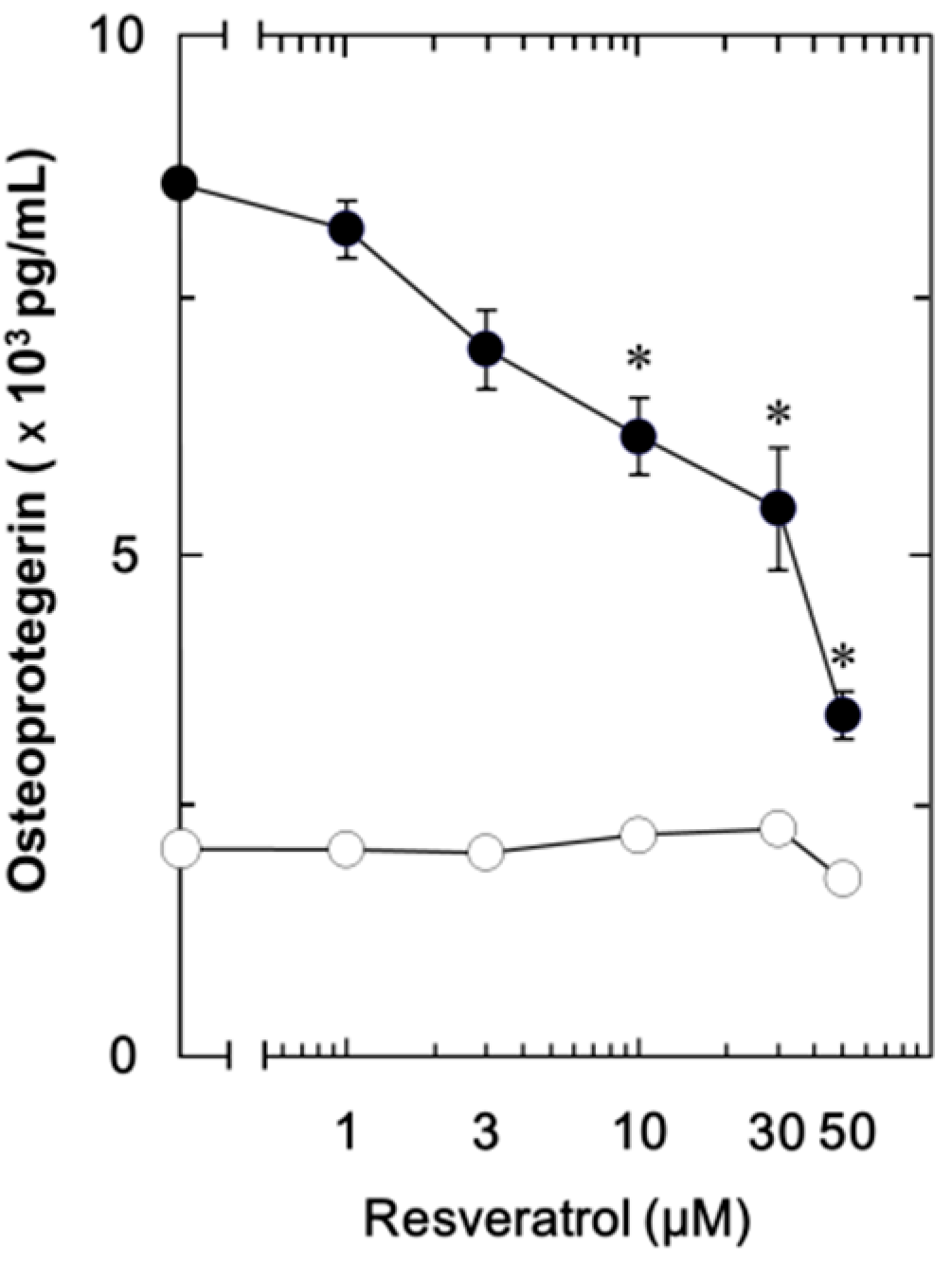
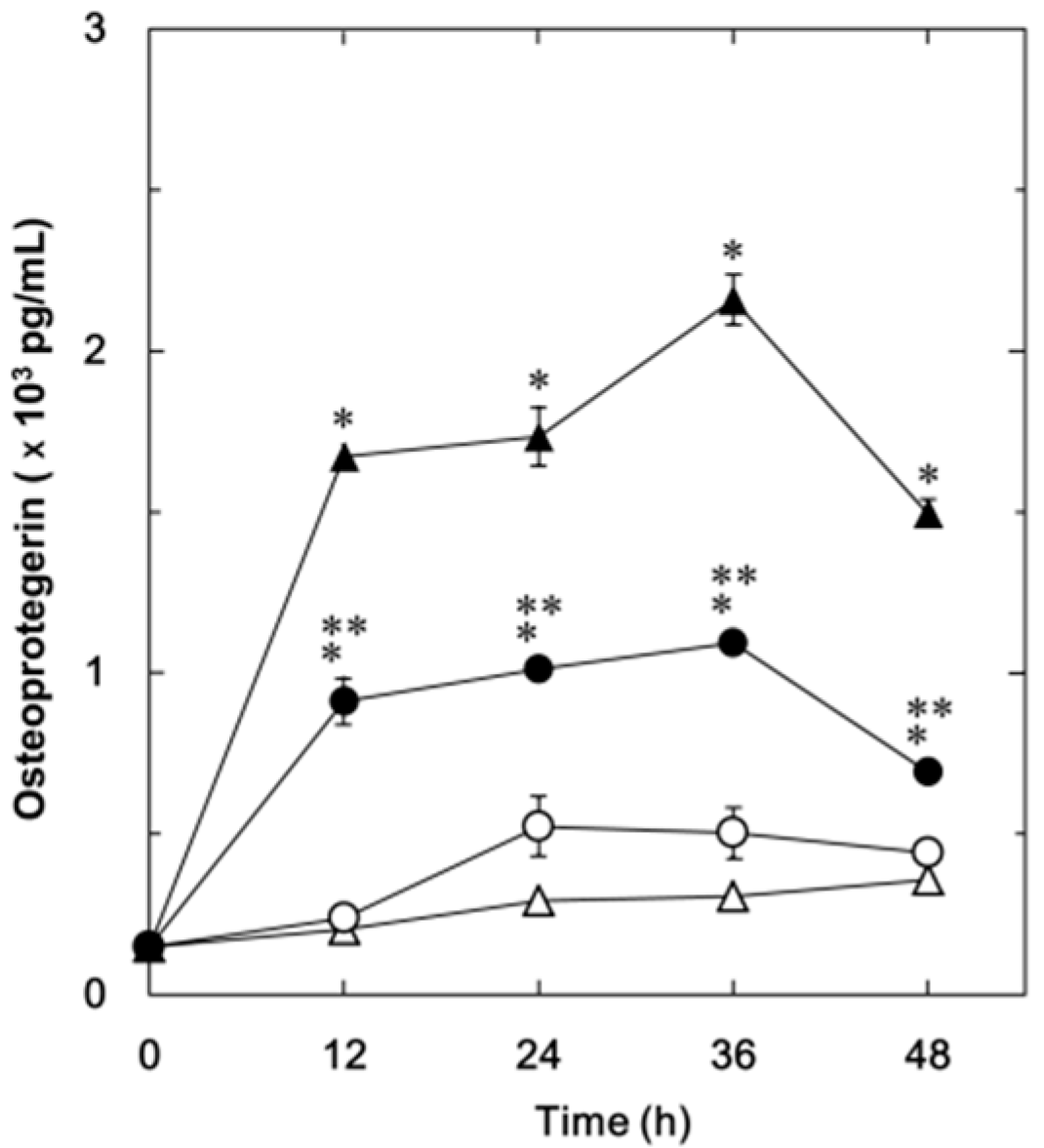
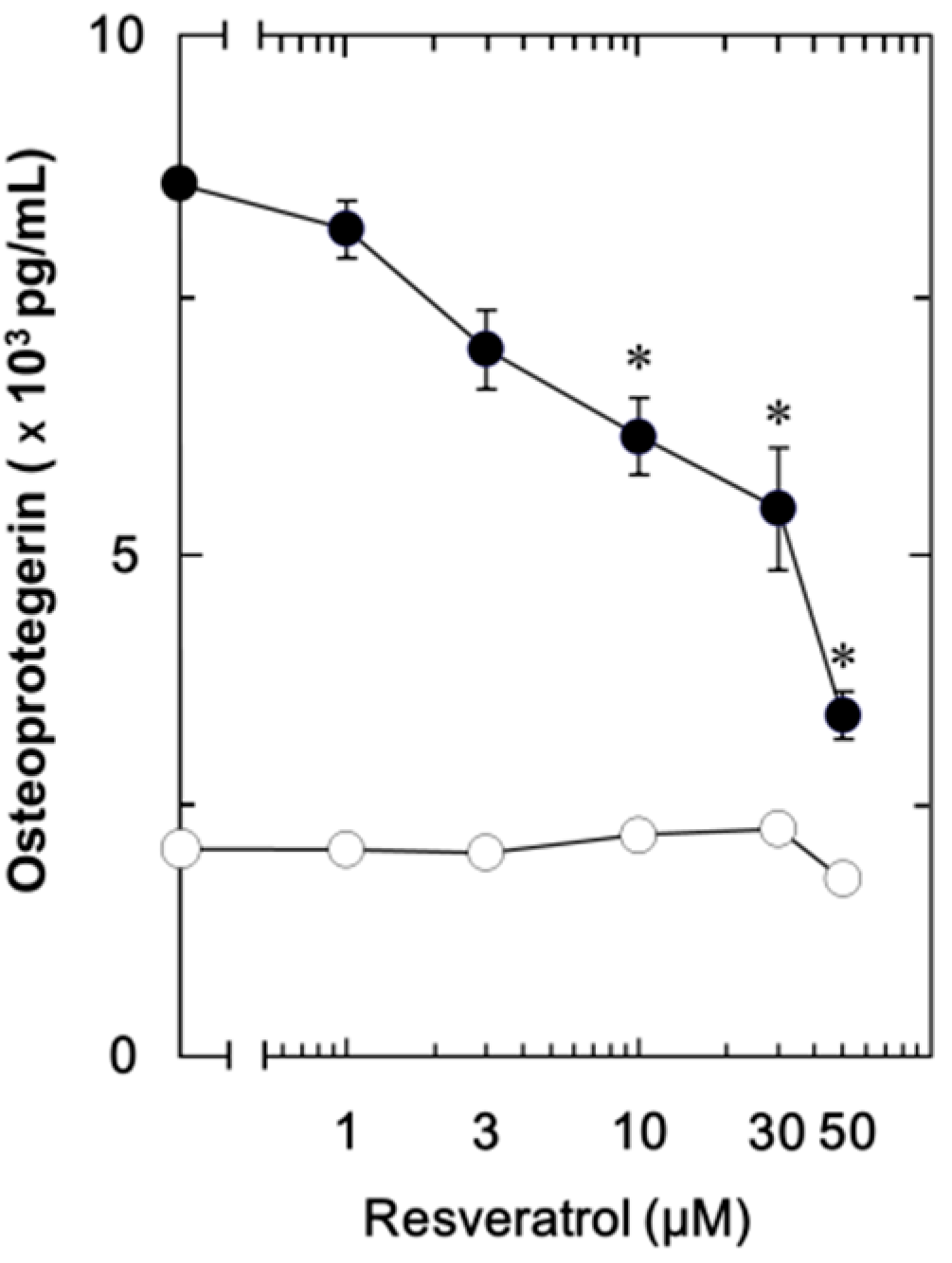
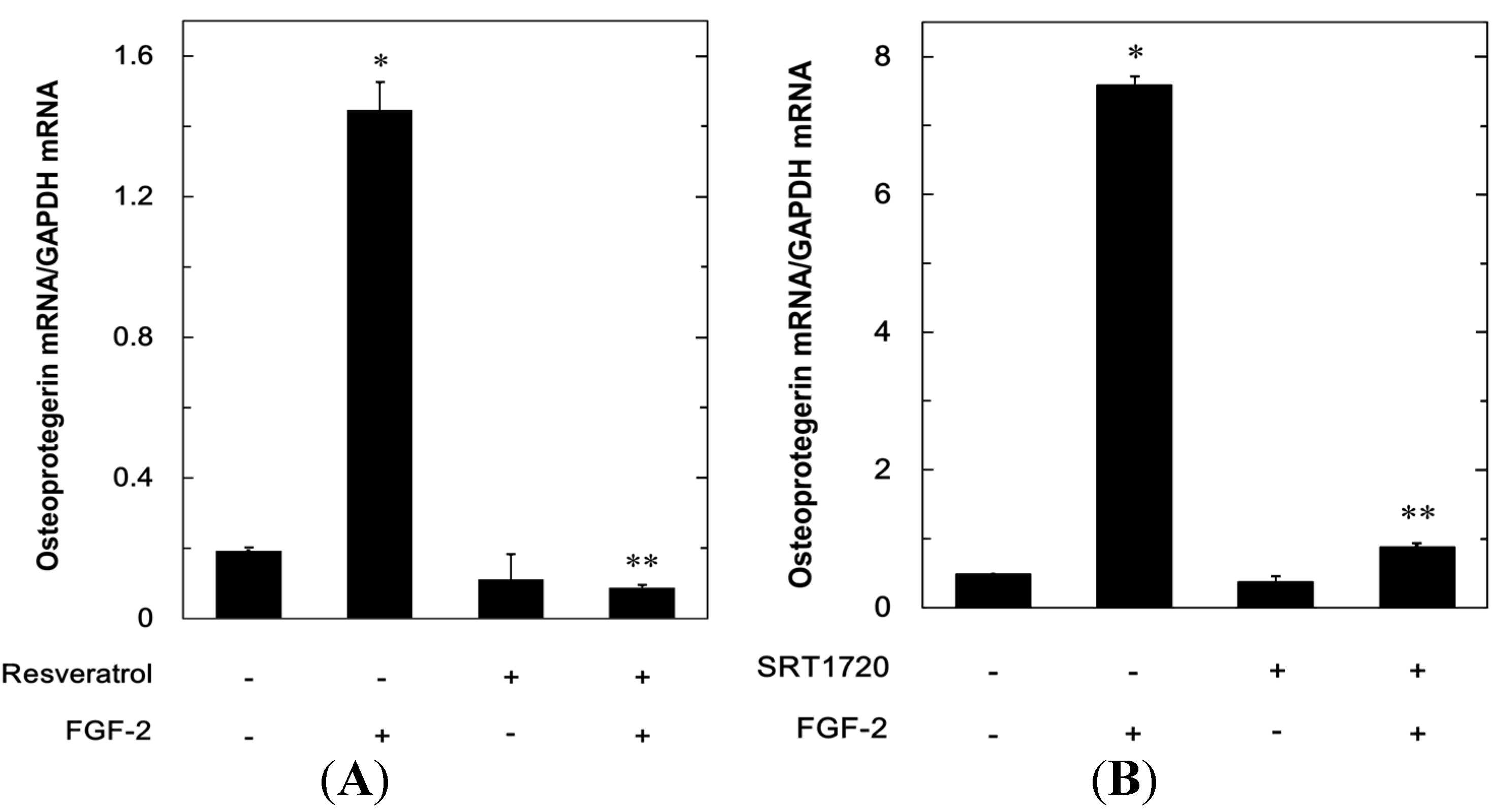
2.1.1. Effect of Resveratrol on the Fibroblast Growth Factor (FGF-2)-Stimulated Osteoprotegerin Release in MC3T3-E1 Cells
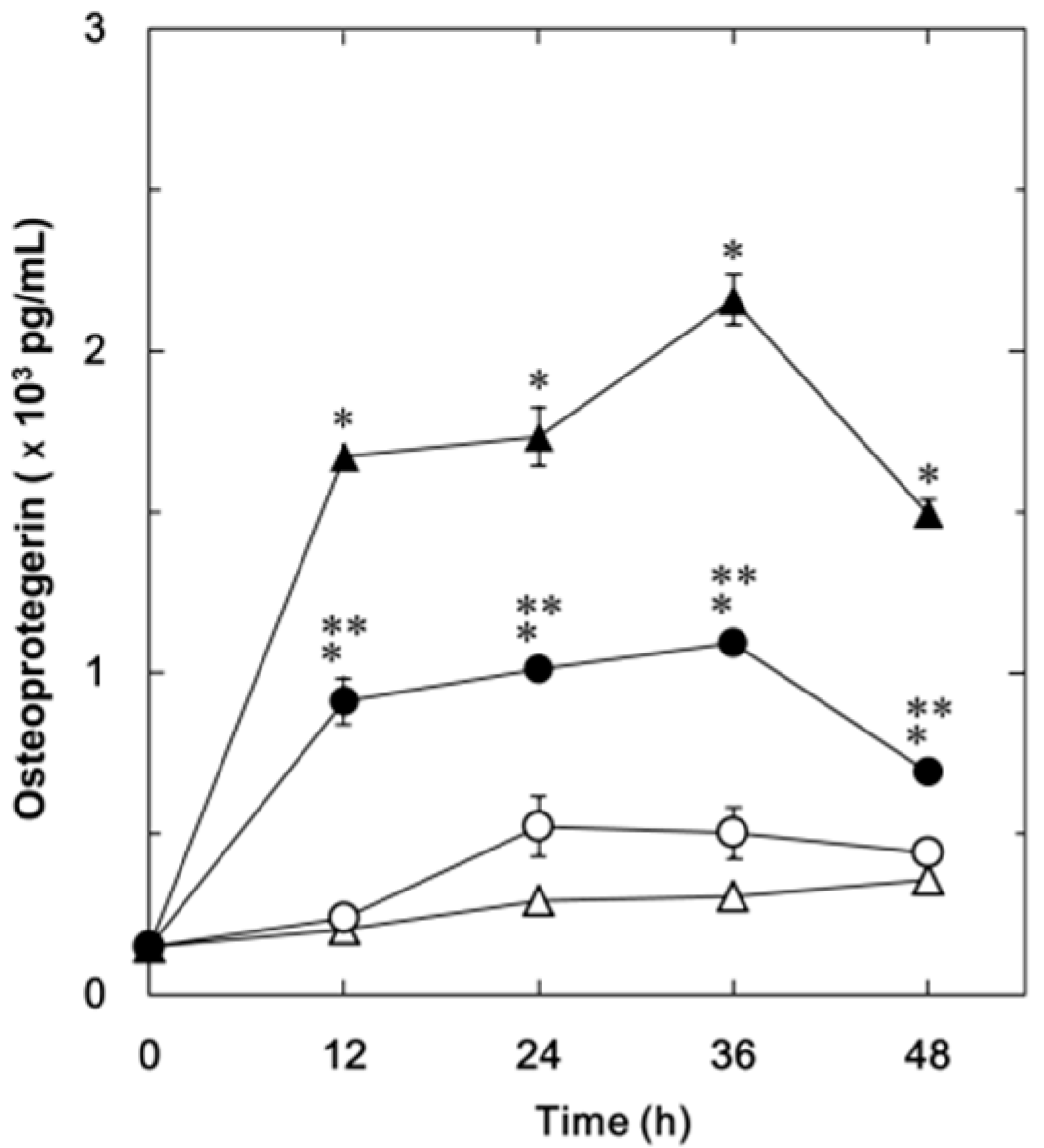
2.1.2. Effect of SRT1720 on the FGF-2-Stimulated Osteoprotegerin Release in MC3T3-E1 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SRT1720 | FGF-2 | Osteoprotegerin (pg/mL) |
|---|---|---|
| − | − | 575 ± 9 |
| − | + | 3,590 ± 299 * |
| + | − | 514 ± 38 |
| + | + | 2,665 ± 72 ** |
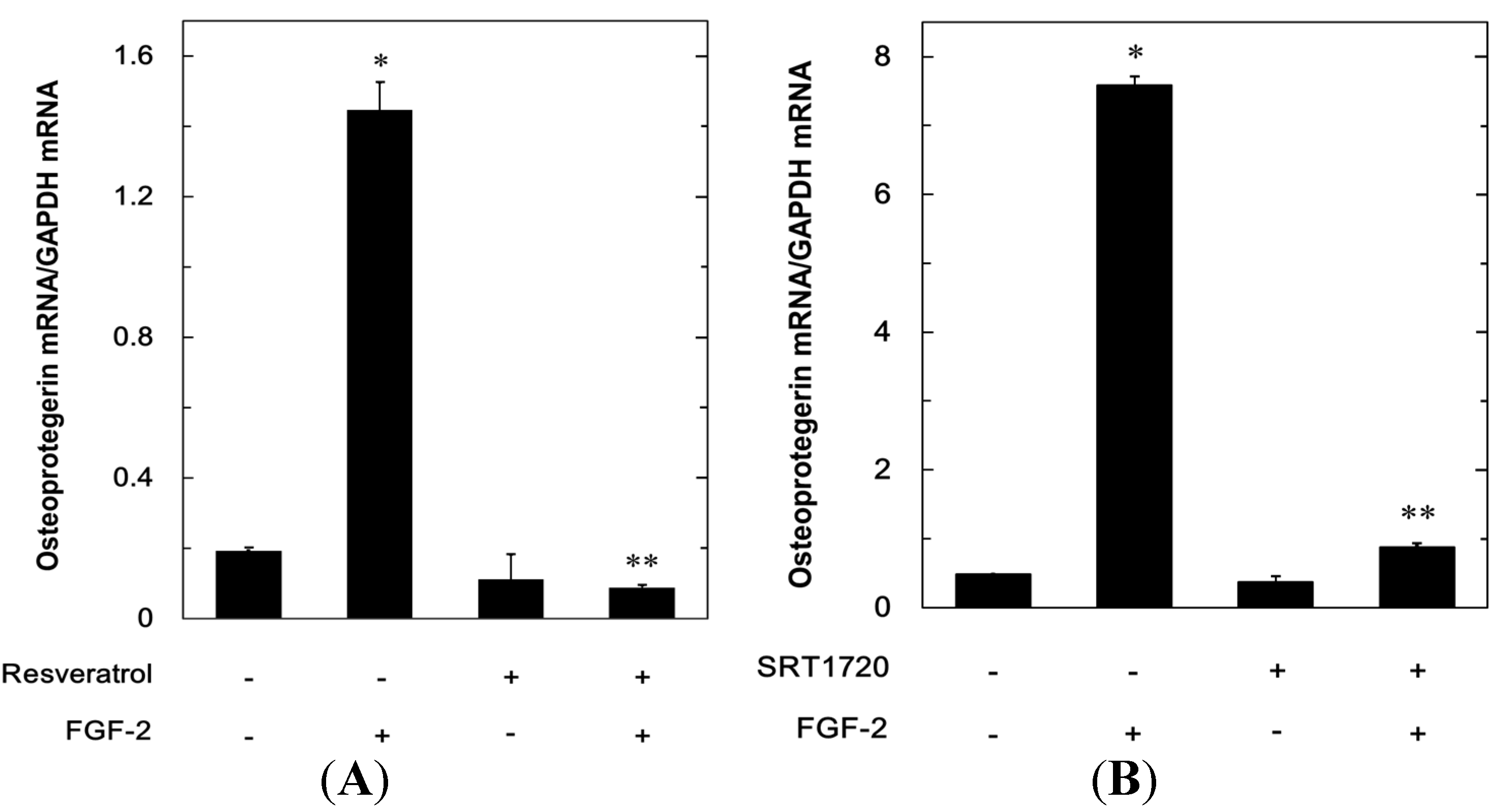
2.1.3. Effects of Resveratrol or SRT1720 on the FGF-2-Induced Expression of mRNA Osteoprotegerin in MC3T3-E1 Cells
2.1.4. Effects of PD98059, SB203580, SP600125 or Akt Inhibitor on the FGF-2-Stimulated Osteoprotegerin Release in MC3T3-E1 Cells
| Inhibitors | FGF-2 | Osteoprotegerin (pg/mL) |
|---|---|---|
| − | − | 1,052 ± 52 |
| − | + | 2,143 ± 80 * |
| PD98059 | − | 1,088 ± 48 |
| PD98059 | + | 2,791 ± 104 |
| SB203580 | − | 609 ± 29 |
| SB203580 | + | 1,005 ± 13 ** |
| SP600125 | − | 879 ± 28 |
| SP600125 | + | 986 ± 64 ** |
| Akt inhibitor | − | 693 ± 28 |
| Akt inhibitor | + | 986 ± 64 ** |
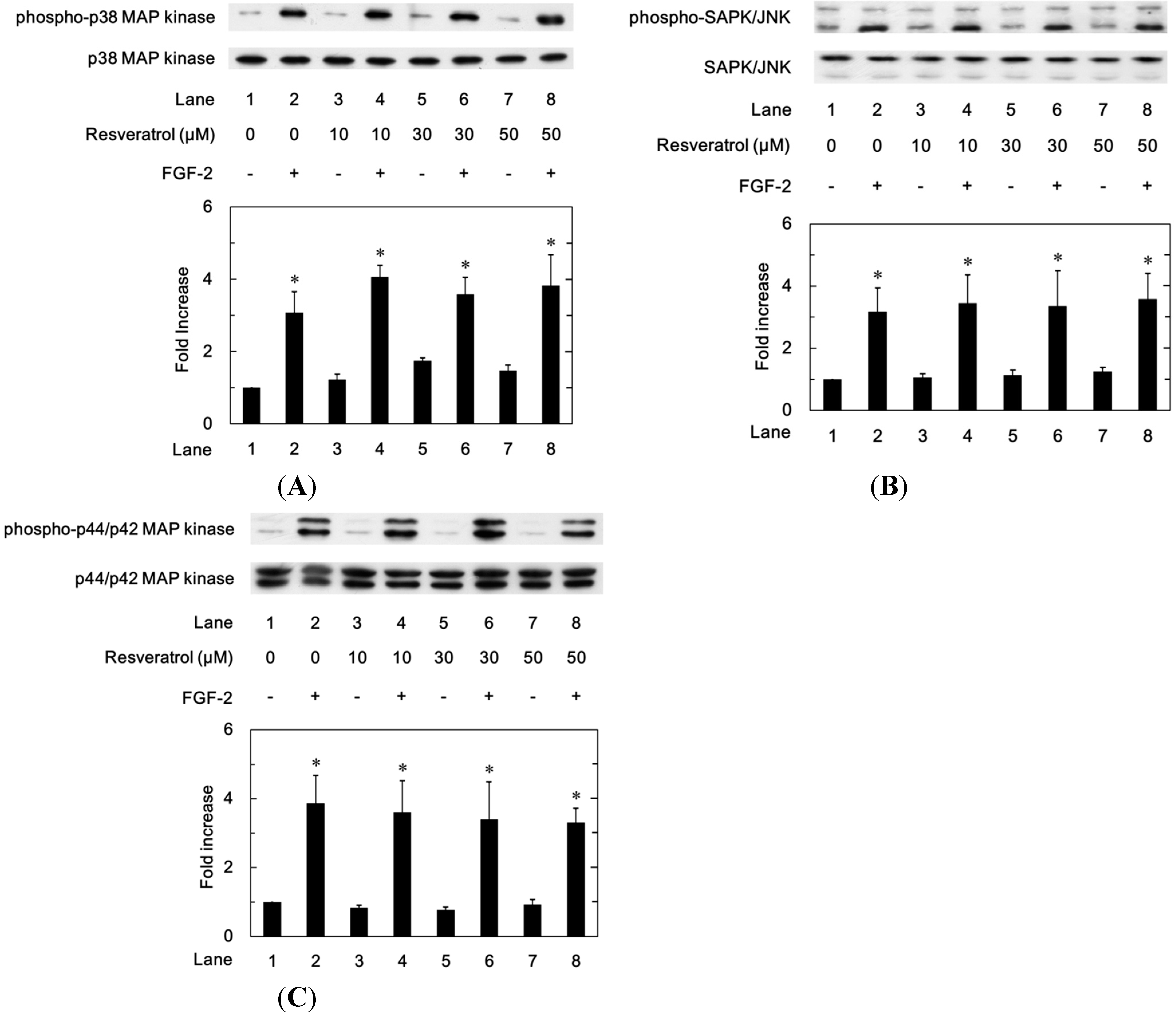
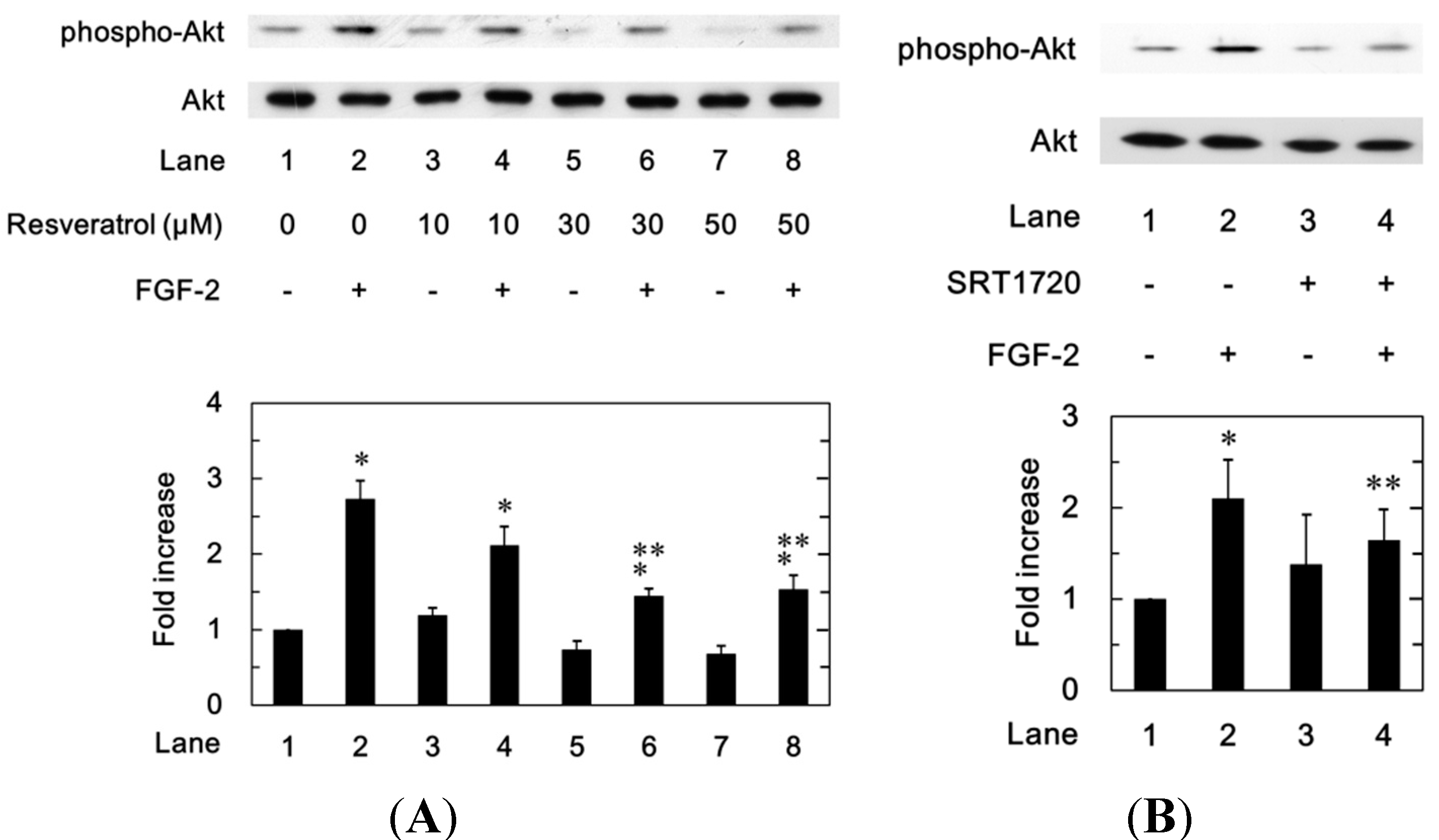
2.1.5. Effects of Resveratrol on the FGF-2-Induced Phosphorylation of p44/p42 MAP Kinase, p38 MAP Kinase or SAPK/JNK in MC3T3-E1 Cells

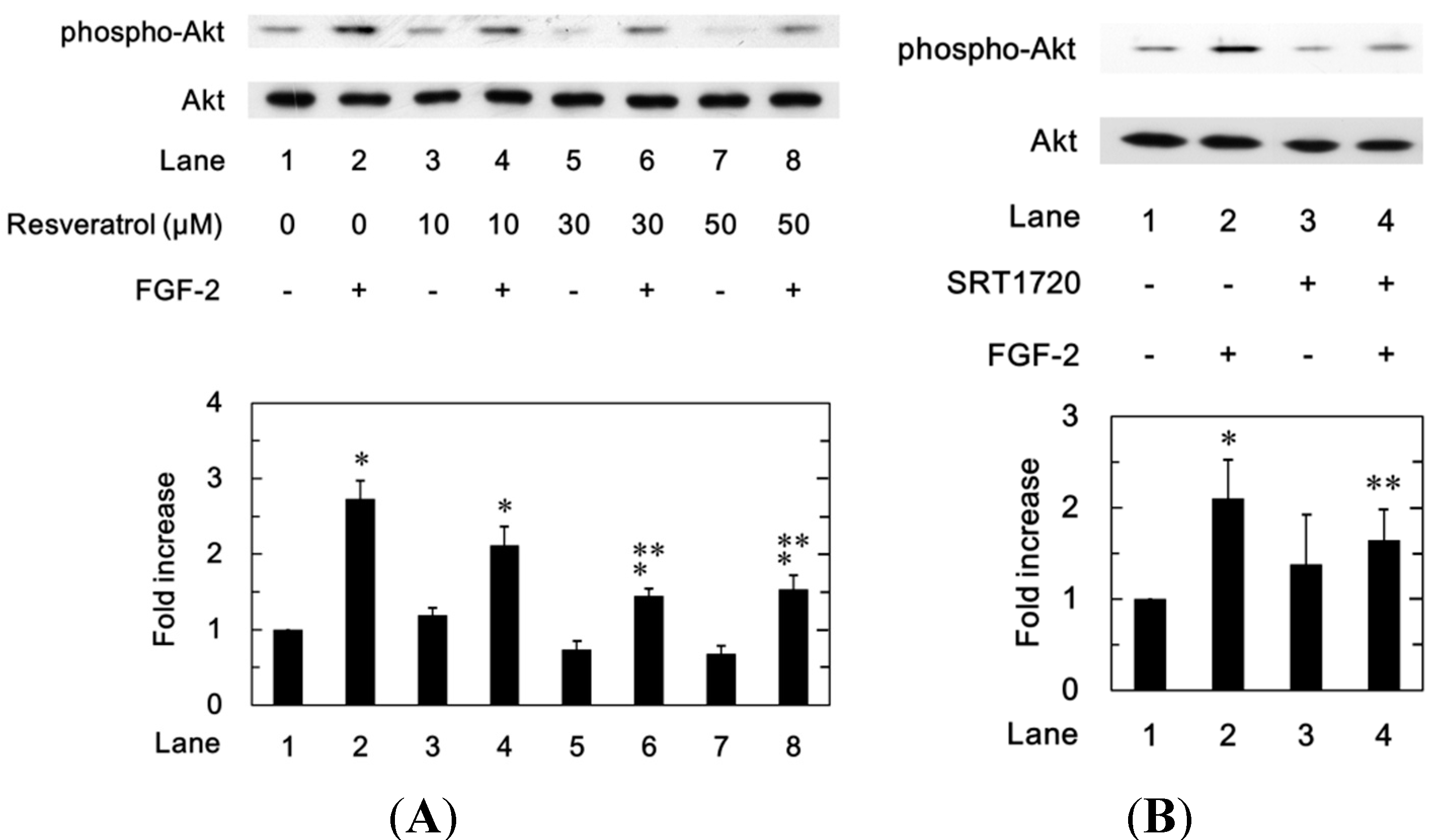
2.1.6. Effects of Resveratrol or SRT1720 on the FGF-2-Induced Phosphorylation of Akt in MC3T3-E1 Cells
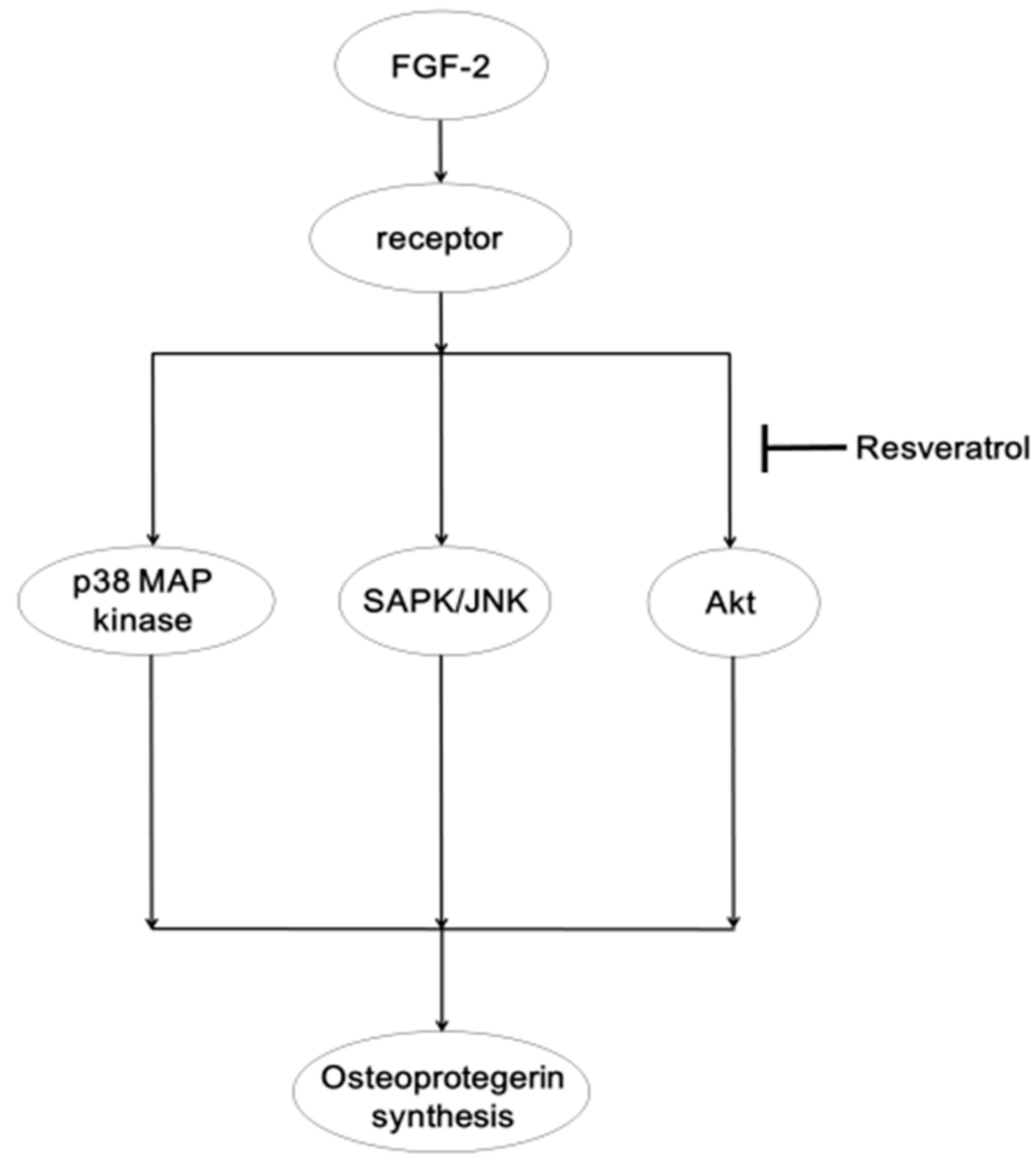
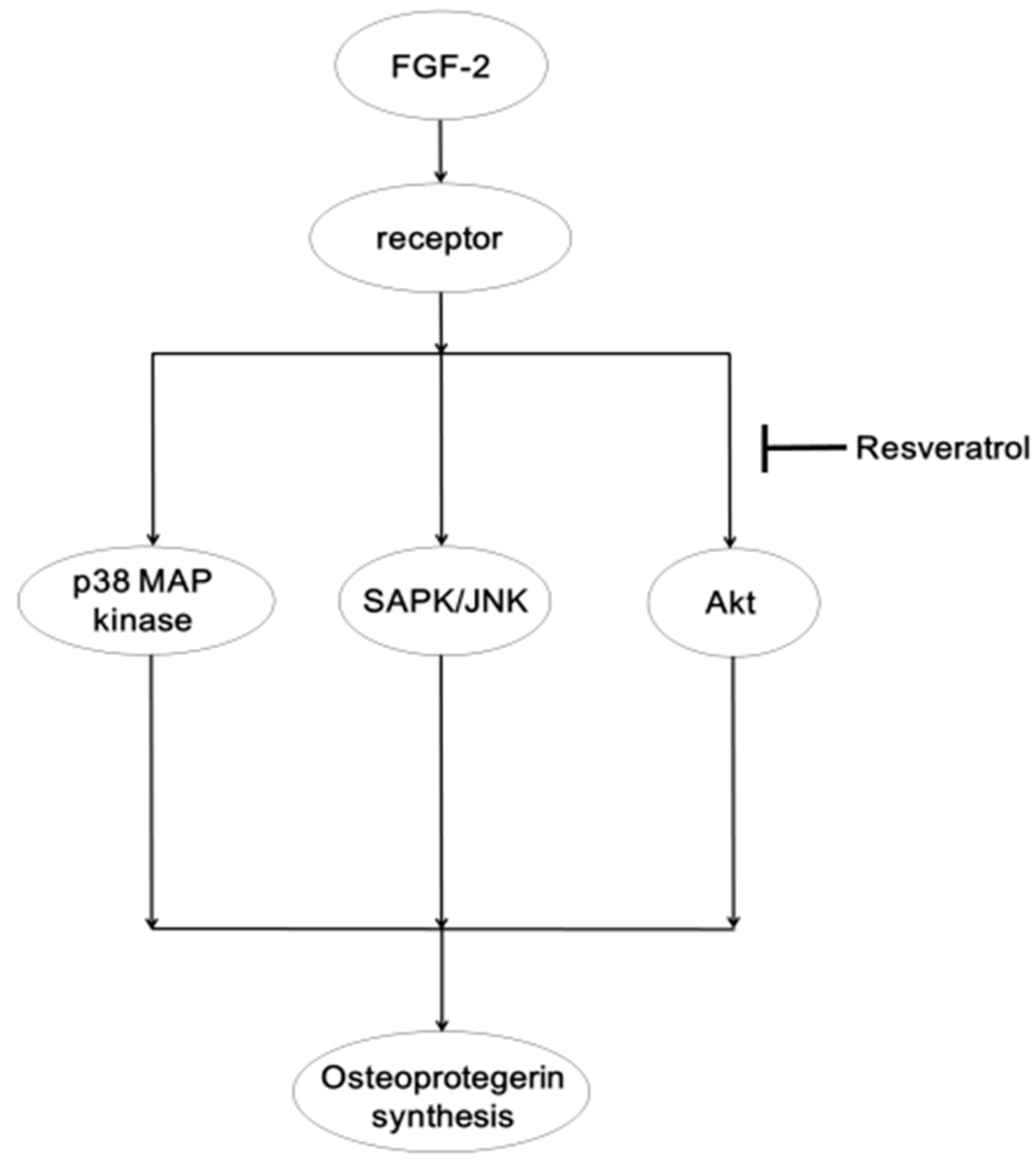
2.2. Discussion
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Assay for Osteoprotegerin
3.4. Real-Time RT-PCR
3.5. Western Blot Analysis
3.6. Densitometric Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Karsenty, G.; Wagner, E.F. Reaching a genetic and molecular understanding of skeletal development. Dev. Cell. 2002, 2, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Parfit, A.M. Targeted and nontargeted bone remodeling: Relationship to basic multicellular unit origination and progression. Bone 2002, 30, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Lüthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, A.; Amizuka, N.; Irie, K.; Murakami, A.; Fujise, N.; Kanno, T.; Sato, Y.; Nakagawa, N.; Yasuda, H.; Mochizuki, S.; et al. Severe osteoporosis in mice lacking osteoclastogenesis inhibitory factor/osteoprotegerin. Biochem. Biophys. Res. Commun. 1998, 247, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Kwan, T.S.; Padrines, M.; Théoleyre, S.; Heymann, D.; Fortun, Y. IL-6, RANKL, TNF-α/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Baylink, D.J.; Finkleman, R.D.; Mohan, S. Growth factor to stimulate bone formation. J. Bone Miner. Res. 1993, 8, 565–572. [Google Scholar] [CrossRef]
- Hurley, M.M.; Abreu, C.; Harrison, J.R.; Lichtler, A.C.; Raisz, L.G.; Kream, B.E. Basic fibroblast growth factor inhibits type I collagen gene expression in osteoblastic MC3T3-E1 cells. J. Biol. Chem. 1993, 268, 5588–5593. [Google Scholar] [PubMed]
- Aguirre, J.I.; Leal, M.E.; Rivera, M.F.; Vanegas, S.M.; Jorgensen, M.; Wronski, T.J. Effects of basic fibroblast growth factor and a prostaglandin E2 receptor subtype 4 agonist on osteoblastogenesis and adipogenesis in aged ovariectomized rats. J. Bone Miner. Res. 2007, 22, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Bolander, M.E. Regulation of fracture repair by growth factors. Proc. Soc. Exp. Biol. Med. 1992, 200, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J. Fibroblast growth factor signaling controlling osteoblast differentiation. Gene 2003, 316, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, H.; Kozawa, O.; Uematsu, T. Basic fibroblast growth factor stimulates vascular endothelial growth factor release in osteoblasts: Divergent regulation by p44/p42 mitogen-activated protein kinase and p38 mitogenactivated protein kinase. J. Bone Miner. Res. 2003, 15, 2371–2379. [Google Scholar] [CrossRef]
- Tokuda, H.; Hirade, K.; Wang, X.; Oiso, Y.; Kozawa, O. Involvement of SAPK/JNK in basic fibroblast growth factor-induced VEGF release in osteoblasts. J. Endocrinol. 2003, 177, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, O.; Tokuda, H.; Matsuno, H.; Uematsu, T. Involvement of p38 mitogen-activated protein kinase in basic fibroblast growth factor-induced interleukin-6 synthesis in osteoblasts. J. Cell. Biochem. 1999, 74, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.O.; Rittenhouse, S.E.; Tsichlis, P.N. AKT/PKB and other D3 phosphoinositide-regulated kinases: Kinase activation by phosphoinositidedependent phosphorylation. Annu. Rev. Biochem. 1999, 68, 965–1014. [Google Scholar] [CrossRef] [PubMed]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, L.R.; Hruska, K.A. The cell survival signal Akt is differently activated by PDGF-BB, EGF, and FGF-2 in osteoblastic cells. J. Cell. Biochem. 2001, 81, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Debiais, F.; Lefevre, G.; Lemonnier, J.; Le Mee, S.; Lasmoles, F. Fibroblast growth factor-2 induces osteoblast survival through a phosphatidylinositol 3-kinase-dependent-β-catenin-independent signaling pathway. Exp. Cell Res. 2004, 297, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Takai, S.; Tokuda, H.; Hanai, Y.; Kozawa, O. Activation of phosphatidylinositol 3-kinase/Akt limits FGF-2-induced VEGF release in osteoblasts. Mol. Cell. Endcrinol. 2007, 267, 46–54. [Google Scholar] [CrossRef]
- Jankun, J.; Selman, S.H.; Wiercz, R.S.; Skrzypczak-Jankun, E. Why drinking green tea could prevent cancer. Nature 1997, 387, 561. [Google Scholar] [CrossRef] [PubMed]
- Harbourne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef] [PubMed]
- Blander, G.; Guarente, L. The Sir2 family of protein deacetylases. Annu. Rev. Biochem. 2004, 73, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.H.; Montminy, M. In vino veritas: A tale of two Sirt1s? Cell 2006, 127, 1091–1093. [Google Scholar] [CrossRef]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef] [PubMed]
- Kubo, J.T.; Stefanick, M.L.; Robbins, J.; Wactawski-Wende, J.; Cullen, M.R.; Freiberg, M.; Desai, M. Preference for wine is associated with lower hip fracture incidence in post-menopausal women. BMC Womens Health 2013, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Allard, V.V.; Prabhu, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Milne, J.C.; Lambert, P.D.; Schenk, S.; Carney, D.P.; Smith, J.J.; Gagne, D.J.; Jin, L.; Boss, O.; Perni, R.B.; Vu, C.B.; et al. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature 2007, 450, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Alessi, D.R.; Cuenda, A.; Cohen, P.; Dudley, D.T.; Saltiel, A.R. PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J. Biol. Chem. 1995, 270, 27489–27494. [Google Scholar] [PubMed]
- Cuenda, A.; Rouse, J.; Doza, Y.N.; Meier, R.; Cohen, P.; Gallagher, T.F.; Young, P.R.; Lee, J.C. SB 203580 is a specific inhibitor of a MAP kinase homologue which is stimulated by cellular stresses and interleukin-1. FEBS Lett. 1995, 364, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.L.; Sasaki, D.T.; Murray, B.W.; O’Leary, E.C.; Sakata, S.T.; Xu, W.; Leisten, J.C.; Motiwala, A.; Pierce, S.; Satoh, Y.; et al. SP600125, an anthrapyrazolone inhibitor of Jun N-terminal kinase. Gene 2001, 98, 13681–13686. [Google Scholar]
- Hu, Y.; Qiao, L.; Wang, S.; Rong, S.B.; Meuillet, E.J.; Berggren, M.; Gallegos, A.; Powis, G.; Kozikowski, A.P. 3-(Hydroxymethyl)-bearing phosphatidylinositol ether lipid analogues and carbonate surrogates block PI3-K, Akt, and cancer cell growth. J. Med. Chem. 2000, 43, 3045–3051. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, N.; Yasuda, H.; Yano, K.; Mochizuki, Si.; Kobayashi, N.; Fujimoto, H.; Shima, N.; Morinaga, T.; Chikazu, D.; Kawaguchi, H.; et al. Basic fibroblast growth factor induces osteoclast formation by reciprocally regulating the production of osteoclast differentiation factor and osteoclastogenesis inhibitory factor in mouse osteoblastic cells. Biochem. Biophys. Res. Commun. 1999, 265, 158–163. [Google Scholar] [PubMed]
- Boily, G.; He, X.H.; Pearce, B.; Jardine, K.; McBurney, M.W. SirT1-null mice develop tumors at normal rates but are poorly protected by resveratrol. Oncogene 2009, 28, 2882–2893. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L.; Picard, F. Calorie restriction-the SIR2 connection. Cell 2005, 120, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [PubMed]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [PubMed]
- Kuroyanagi, G.; Mizutani, J.; Kondo, A.; Yamamoto, N.; Matsushima-Nishiwaki, R.; Otsuka, T.; Kozawa, O.; Tokuda, H. Suppression by resveratrol of prostaglandin D2-stimulated osteoprotegerin synthesis in osteoblasts. Prostaglandins Leukot. Essent. Fatty Acids 2014, 91, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kuroyanagi, G.; Tokuda, H.; Matsushima-Nishiwaki, R.; Kondo, A.; Mizutani, J.; Kozawa, O.; Otsuka, T. Resveratrol suppresses prostaglandin F2α-induced osteoprotegerin synthesis in osteoblasts: Inhibition of the MAP kinase signaling. Arch. Biochem. Biophys. 2014, 542, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S.; Sugimoto, Y.; Ushikubi, F. Prostanoid receptors: Structures, properties, and functions. Physiol. Rev. 1999, 79, 1193–1226. [Google Scholar] [PubMed]
- Eswarakumar, V.P.; Lax, I.; Schlessinger, J. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev. 2005, 16, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Pillai, V.B.; Sundaresan, N.R.; Gupta, M.P. Regulation of Akt signaling by sirtuins: Its implication in cardiac hypertrophy and aging. Circ. Res. 2014, 114, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Sudo, H.; Kodama, H.; Amagai, Y.; Yamamoto, S.; Kasai, S. In vitro differentiation and calcification in a new clonal osteogenic cell line derived from newborn mouse calvaria. J. Cell Biol. 1983, 96, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, O.; Tokuda, H.; Miwa, M.; Kotoyori, J.; Oiso, Y. Cross-talk regulation between cyclic AMP production and phosphoinositide hydrolysis induced by prostaglandin E2 in osteoblast-like cells. Exp. Cell Res. 1992, 198, 130–134. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Ito, H.; Hasegawa, K.; Inaguma, Y.; Kozawa, O.; Asano, T. Modulation of the stress-induced synthesis of hsp27 and αB-crystallin by cyclic AMP in C6 rat glioma cells. J. Neurochem. 1996, 66, 946–950. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuroyanagi, G.; Otsuka, T.; Yamamoto, N.; Matsushima-Nishiwaki, R.; Nakakami, A.; Mizutani, J.; Kozawa, O.; Tokuda, H. Down-Regulation by Resveratrol of Basic Fibroblast Growth Factor-Stimulated Osteoprotegerin Synthesis through Suppression of Akt in Osteoblasts. Int. J. Mol. Sci. 2014, 15, 17886-17900. https://doi.org/10.3390/ijms151017886
Kuroyanagi G, Otsuka T, Yamamoto N, Matsushima-Nishiwaki R, Nakakami A, Mizutani J, Kozawa O, Tokuda H. Down-Regulation by Resveratrol of Basic Fibroblast Growth Factor-Stimulated Osteoprotegerin Synthesis through Suppression of Akt in Osteoblasts. International Journal of Molecular Sciences. 2014; 15(10):17886-17900. https://doi.org/10.3390/ijms151017886
Chicago/Turabian StyleKuroyanagi, Gen, Takanobu Otsuka, Naohiro Yamamoto, Rie Matsushima-Nishiwaki, Akira Nakakami, Jun Mizutani, Osamu Kozawa, and Haruhiko Tokuda. 2014. "Down-Regulation by Resveratrol of Basic Fibroblast Growth Factor-Stimulated Osteoprotegerin Synthesis through Suppression of Akt in Osteoblasts" International Journal of Molecular Sciences 15, no. 10: 17886-17900. https://doi.org/10.3390/ijms151017886
APA StyleKuroyanagi, G., Otsuka, T., Yamamoto, N., Matsushima-Nishiwaki, R., Nakakami, A., Mizutani, J., Kozawa, O., & Tokuda, H. (2014). Down-Regulation by Resveratrol of Basic Fibroblast Growth Factor-Stimulated Osteoprotegerin Synthesis through Suppression of Akt in Osteoblasts. International Journal of Molecular Sciences, 15(10), 17886-17900. https://doi.org/10.3390/ijms151017886