Association of Polymorphisms of Exon 2 of the Growth Hormone Gene with Production Performance in Huoyan Goose

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results of Genomic DNA Extraction
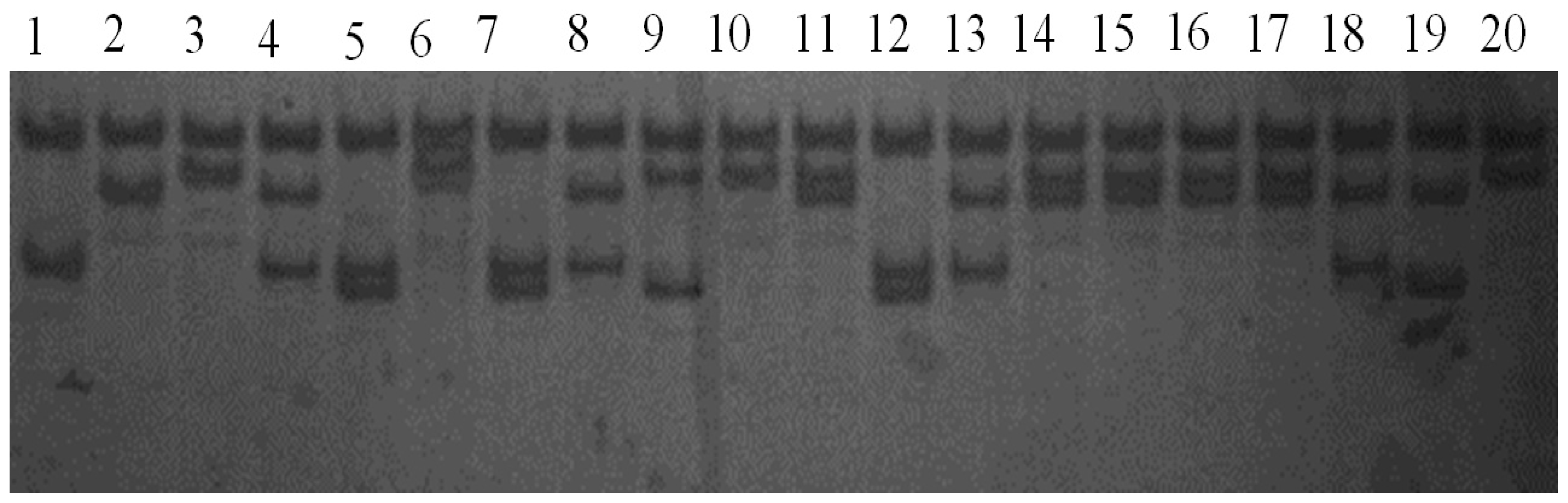
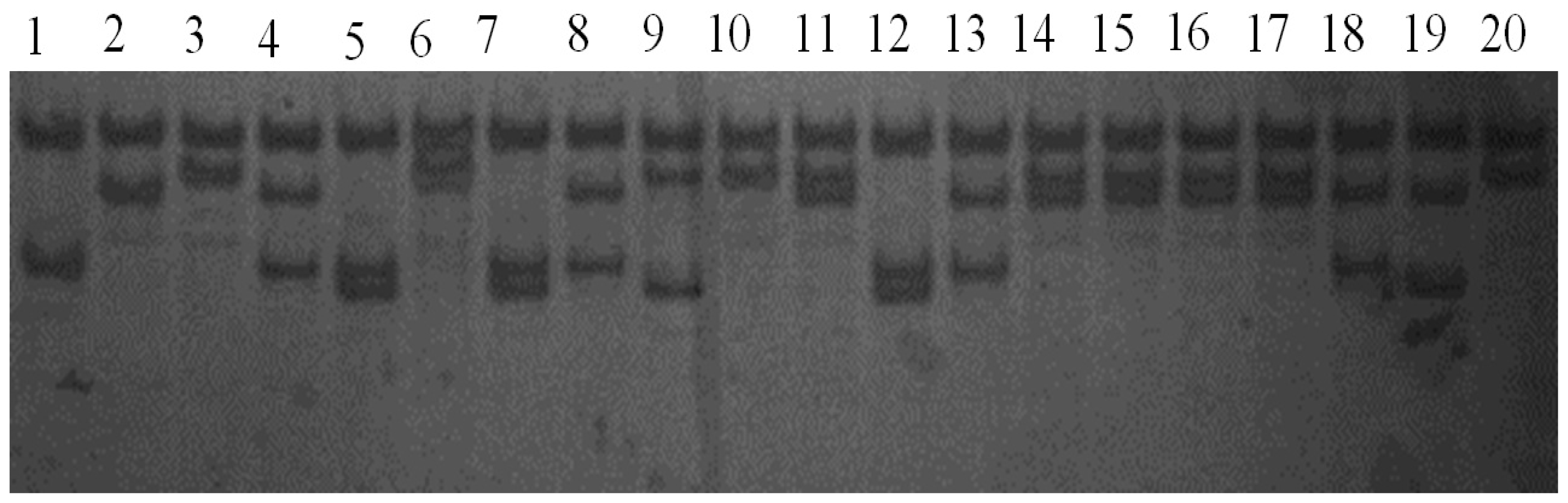
2.2. Results of PCR-SSCP Detection
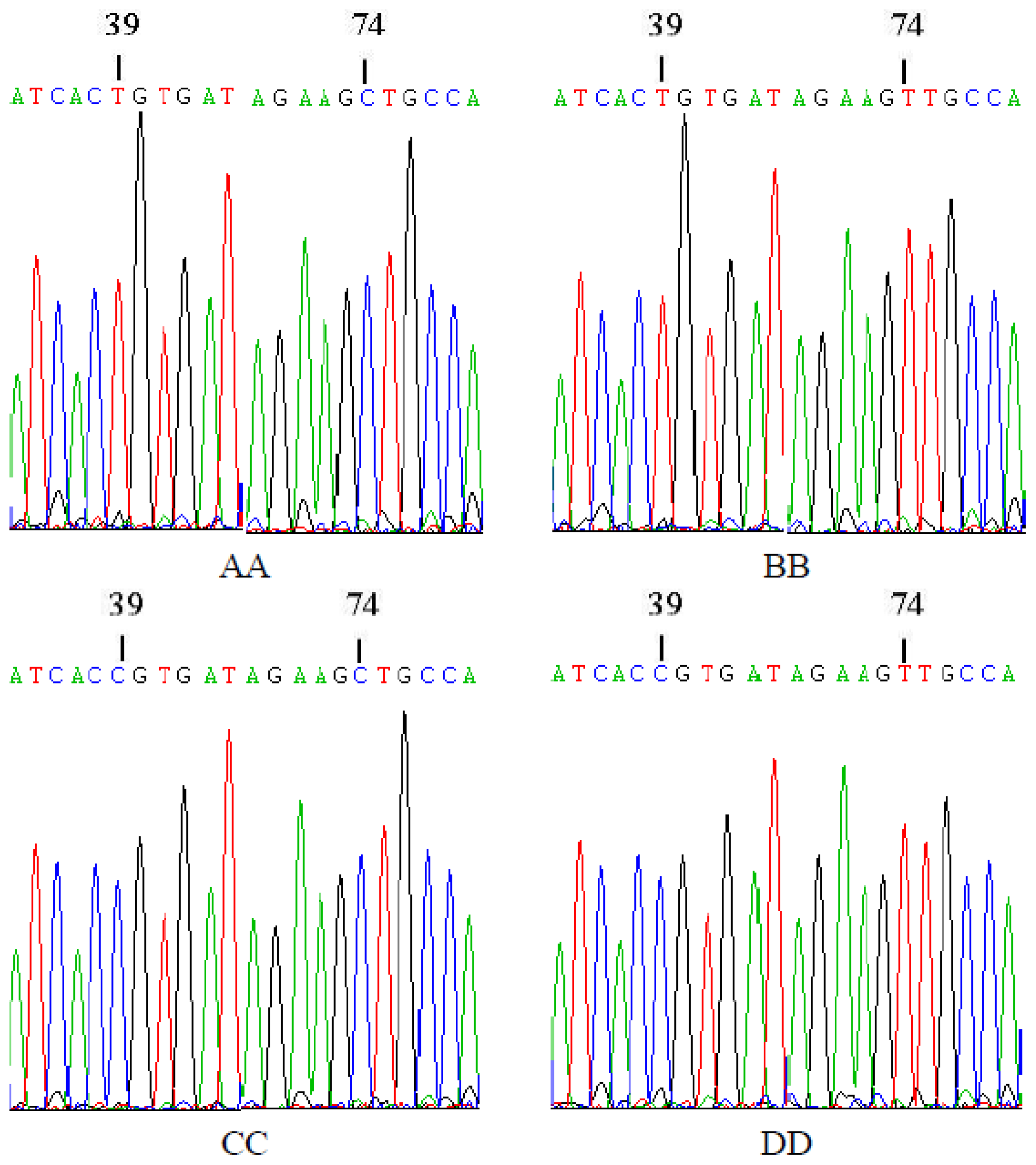
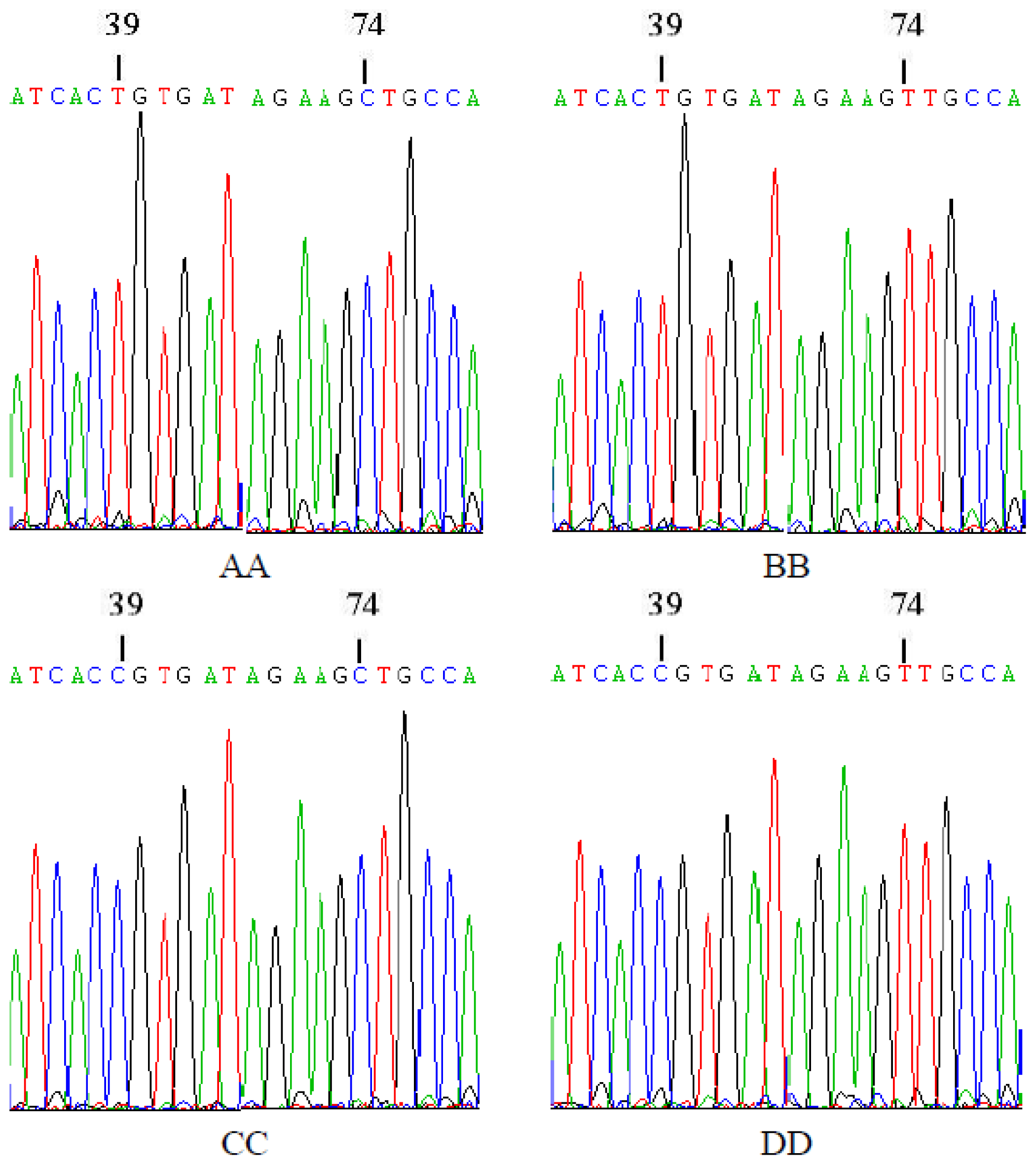
2.3. Sequencing of Homozygous Genotypes
2.4. Genotypes and Variations in the Allelic Frequencies of Exon 2 of the GH Gene
2.5. Body Weight and Body Traits of Geese with Different Genotypes
2.6. Carcass Traits of Genotypes
2.7. Meat Quality of Genotypes
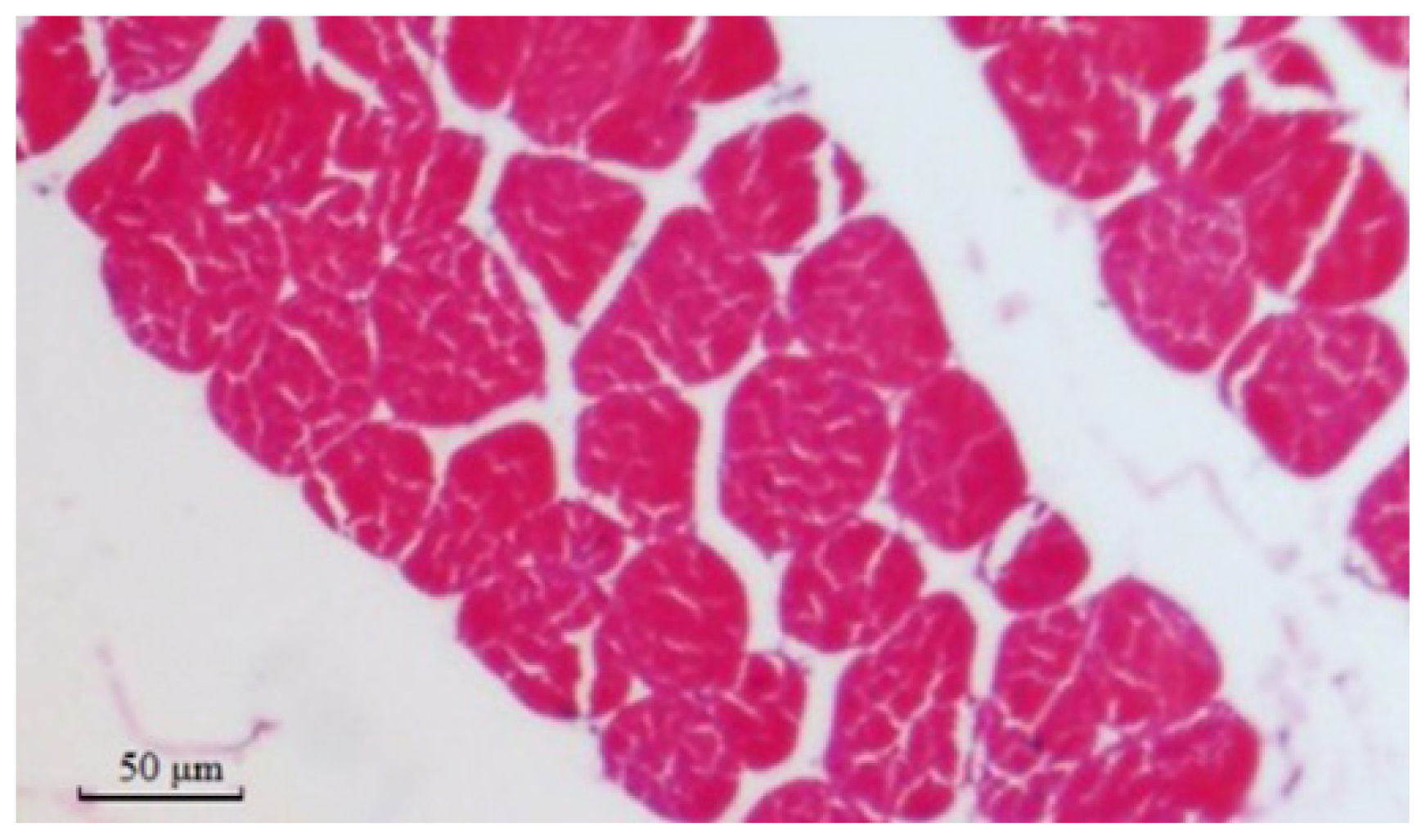
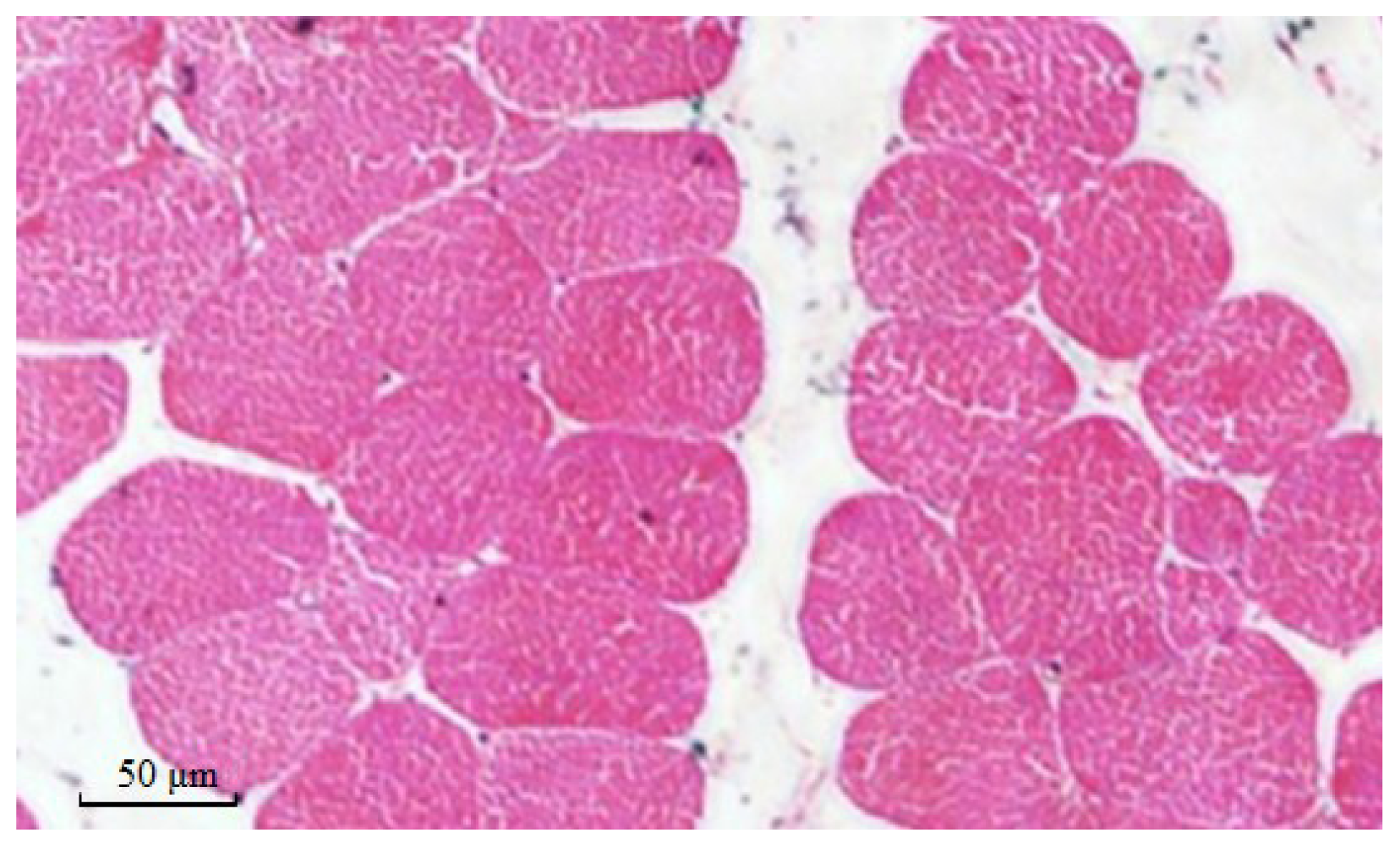
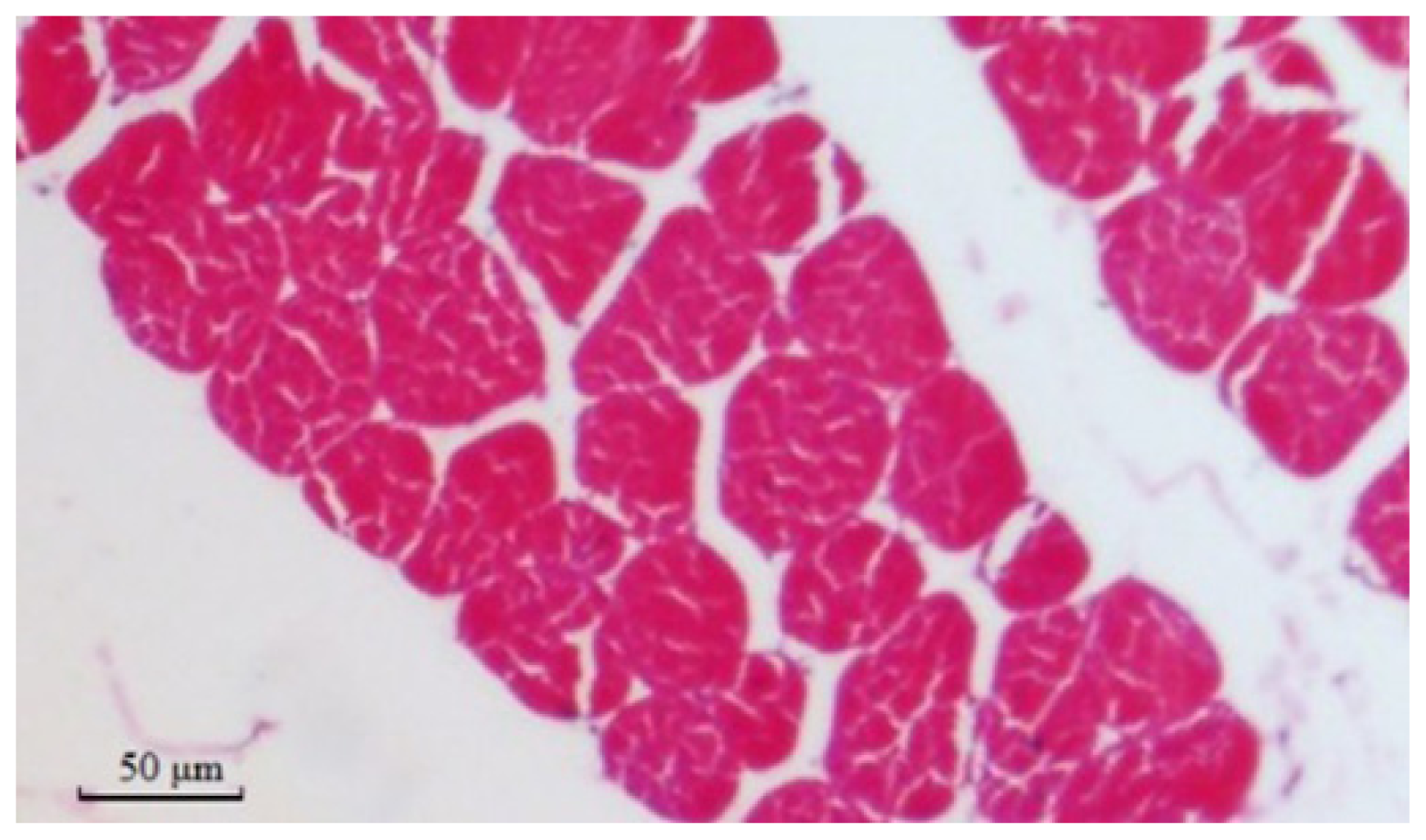
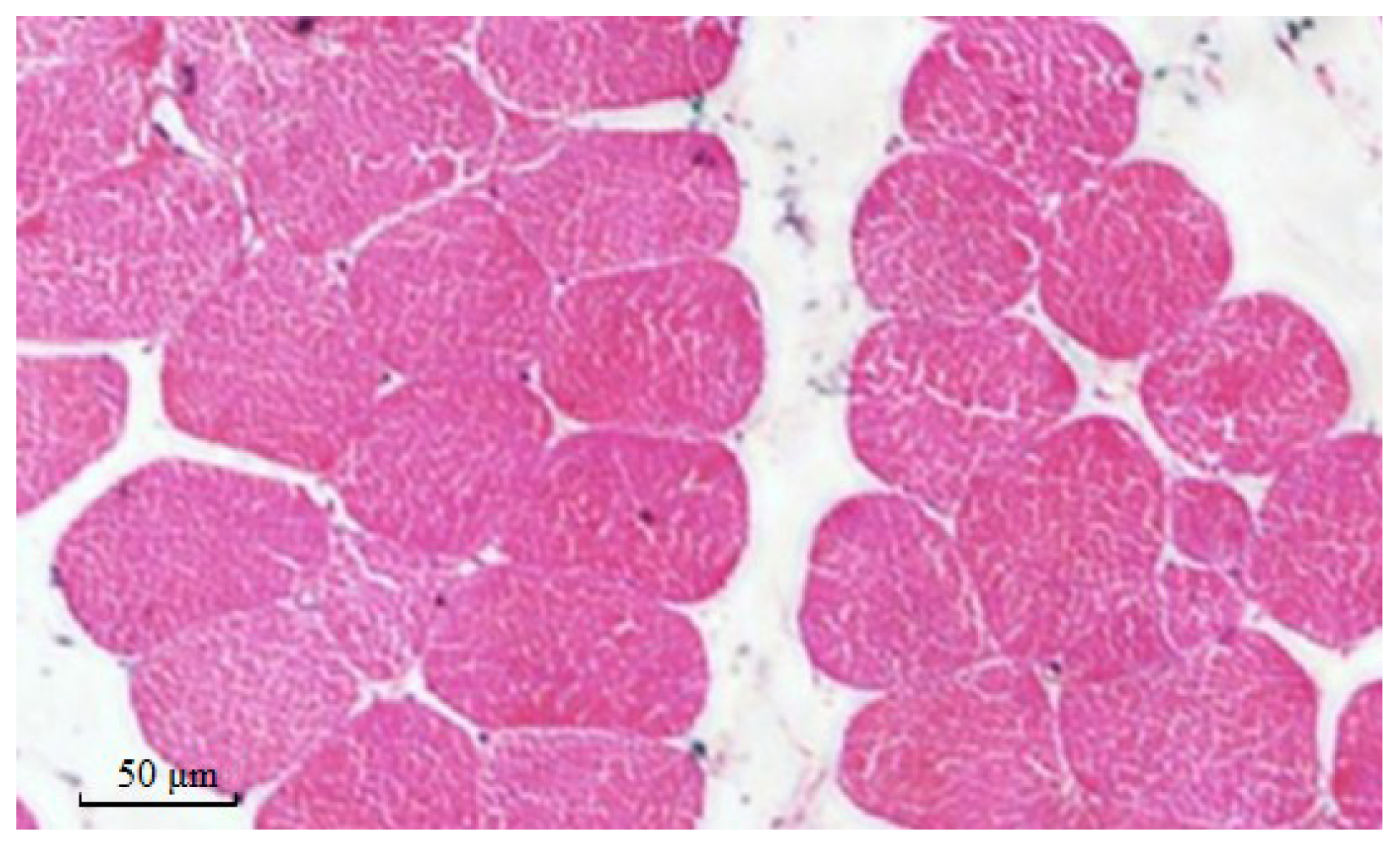
2.8. Muscle Fiber Traits in Different Genotypes
3. Materials and Methods
3.1. Experimental Animals and Blood Collection
3.2. Genomic DNA Extraction and Isolation
3.3. PCR Primer Design and Reagents
3.4. PCR-SSCP Analysis
3.5. Sequencing of Homozygous Genotypes
3.6. Statistical Analyses of Production Performance Measurements
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Min, Y.N.; Hou, S.S.; Gao, Y.P.; Huang, W.; Liu, F.Z. Effect of dietary crude protein and energy on gosling growth performance and carcass trait. Poult. Sci 2007, 86, 661–664. [Google Scholar]
- Chen, G.H.; Wang, K.H.; Wang, J.Y. Goose breeds. In Poultry Genetic Resources in China; Shanghai Science and Technology Press: Shanghai, China, 2004; p. 120. [Google Scholar]
- Wang, J.W.; Qiu, X.P.; Zeng, F.T.; Shi, X.W.; Zhang, Y.P. Genetic differentiation of domestic goose breeds in China. Acta Genet. Sin 2005, 32, 1053–1059. [Google Scholar]
- Kato, Y.; Murakami, Y.; Sohmiya, M.; Nishiki, M. Regulation of human growth hormone secretion and its disorders. Jpn. J. Med 2002, 41, 7–13. [Google Scholar]
- Qian, M.; Liu, S.F.; Zhuang, Z.; Lin, M.L.; Sun, Z.Z.; Liu, C.L.; Ma, H.; Su, Y.Q.; Tang, Q.S. Genomic structure, polymorphism and expression analysis of the growth hormone(GH) gene in female and male Half-smooth tongue sole (Cynoglossus semilaevis). Gene 2012, 493, 92–104. [Google Scholar]
- Ohlsson, C.; Bengtsson, B.A.; Isaksson, O.G.P.; Andreassen, M.C. Slootweg, growth hormone and bone. Endocr. Rev 1998, 19, 55–79. [Google Scholar]
- Millar, D.S.; Horan, M.; Chuzhanova, A.N.; Cooper, D.N. Characterisation of a functional intronic polymorphism in the human growth hormone (GH1) gene. Hum. Genomics 2010, 4, 289–301. [Google Scholar]
- Gordona, D.F.; Quickb, D.P.; Erwinc, C.R.; Donelsonc, J.E.; Maurer, R.A. Nucleotide sequence of the bovine growth hormone chromosomal gene. Mol. Cell. Endocrinol 1983, 33, 81–95. [Google Scholar]
- Mou, L.J.; Liu, N.; Zadworny, D.; Chalifour, L.; Kuhnlein, U. Presence of an additional PstI fragment in intronhormone-encoding gene. Gene 1995, 160, 313–314. [Google Scholar]
- Harvey, S.; Scanes, C.G.; Daughaday, W.H. Growth Hormone; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Hua, G.H.; Chen, S.L.; Yu, J.N.; Cai, K.L.; Wua, C.J.; Li, Q.L.; Zhang, C.Y.; Liang, A.X.; Hana, L.; Geng, L.Y.; et al. Polymorphism of the growth hormone gene and its association with growth traits in Boer goat bucks. Meat Sci 2009, 81, 391–395. [Google Scholar]
- Balogh, O.; Kovács, K.; Kulcsár, M.; Gáspárdy, A.; Zsolnai, A.; Kátai, L.; Pécsi, A.; Fésüs, L.; Butler, W.R.; Huszenicza, G. AluI polymorphism of the bovine growth hormone (GH) gene, resumption of ovarian cyclicity, milk production and loss of body condition at the onset of lactation in dairy cows. Theriogenology 2009, 4, 553–559. [Google Scholar]
- Dettori, M.L.; Rocchigiani, A.M.; Luridiana, S.; Mura, M.C.; Carcangiu, V.; Pazzola, M. Growth hormone gene variability and its effects on milk traits in primiparous Sarda goats. J. Dairy Res 2013, 80, 255–262. [Google Scholar]
- Zhao, W.M.; Qiao, N.; Wang, X.B.; Chen, Q.; Cheng, J.H.; Xu, Q.; Chen, G.H. Comparative genomic analysis of growth hormone gene in geese. Anim. Sci. J 2011, 82, 62–66. [Google Scholar]
- Zhang, X.L.; Jiang, X.; Liu, Y.P.; Du, H.R.; Zhu, Q. Identification of Ava I polymorphisms in the third intron of GH gene and their associations with abdominal fat in chickens. Poult. Sci 2007, 86, 1079–1083. [Google Scholar]
- Ku, H.; Ni, L.; Wei, G.S. DNA polymorphisms in the chicken growth hormone gene: Response to selection for disease resistance and association with egg production. Anim. Genet 1997, 28, 116–123. [Google Scholar]
- Kansaku, N.; Zadworny, D.; Guemene, D. Genomic Cloning of Duck Growth Hormone; NCBI. GENEBANK: Bethesda, MD, USA, 2004. [Google Scholar]
- Zhan, K.; Yang, N. Effects of polymorphism in the coding region of GH gene on serum GH, T3 levels and body weight of ducks. Proceedings of the 3rd World Waterfowl Conference, Guangzhou, China; 2005; pp. 242–246. [Google Scholar]
- Dong, B.; Wang, J.; Duan, X.J.; Sun, G.B.; Zhu, S.Y.; Li, X.F. Polymorphism of the exons of growth hormone (GH) gene in goose. Jiangsu J. Agric. Sci 2010, 26, 1020–1025. [Google Scholar]
- Duan, X.J.; Dong, B.; Wang, J. Polymorphism in exon 2 of GH gene in six white geese. Anhui Nongxueyuan Xuebao 2010, 37, 41–46. [Google Scholar]
- Heidari, M.; Azari, M.A.; Hasani, S.; Khanahmadi, A.; Zerehdaran, S. Effect of polymorphic variants of GH, Pit-1, and β-LG genes on milk production of Holstein cows. Genetika 2012, 48, 503–507. [Google Scholar]
- Ding, X.Z.; Liang, C.N.; Guo, X.; Xing, C.F.; Bao, J.; Chu, M.; Pei, J.; Zhu, X.S.; Yan, P. A novel single nucleotide polymorphism in exon 7 of LPL gene and its association with carcass traits and visceral fat depositionin yak (Bos grunniens) steers. Mol. Biol. Rep 2012, 39, 669–673. [Google Scholar]
- Lemamy, G.J.; Guillaume, V.; Ndeboko, B.; Mouecoucou, J.; Oliver, C. Substance P stimulates Growth Hormone (GH) and GH-Releasing Hormone (GHRH) secretions through tachykinin NK2 receptors in sheep. Peptides 2012, 35, 60–64. [Google Scholar]
- Chang, M.T.; Cheng, Y.S.; Huang, M.C. The SNP genotypes of growth hormone gene associated with reproductive traits in Tsaiya ducks. Reprod. Domest. Anim 2012, 47, 568–573. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual; Science Press: Beijing, China, 2002; pp. 463–470. [Google Scholar]
- Zhou, Y.; Liu, Y.P.; Jiang, X.S.; Du, H.R.; Li, X.C.; Zhu, Q. Polymorphism of chicken myocyte-specific enhancer-binding factor 2A gene and its association with chicken carcass traits. Mol. Biol. Rep 2010, 37, 587–594. [Google Scholar]
- Ministry of Agriculture of the People’s Republic of China, Agricultural industry standard. In Poultry Production Terms and Measurement Method. NY/T 823-2004; China Agriculture Press: Beijing, China, 2005.
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar]
- Dai, J.J.; Yang, A. A modified chi-squared goodness-of-fit test. J. Math. Res. Appl 2009, 29, 113–123. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Populations (Sample Size) | Male (275) | Female (277) | Total Group (552) |
|---|---|---|---|
| Part A. Genotype | |||
| AA | 0.0182 (5) | 0.0361 (10) | 0.0272 (15) |
| AB | 0.0873 (24) | 0.0758 (21) | 0.0815 (45) |
| AC | 0.1382 (38) | 0.1300 (36) | 0.1341 (74) |
| AD | 0.1055 (29) | 0.1588 (44) | 0.1322 (73) |
| BB | 0.0255 (7) | 0.0397 (11) | 0.0326 (18) |
| BC | 0.1273 (35) | 0.0866 (24) | 0.1069 (59) |
| BD | 0.1709 (47) | 0.1227 (34) | 0.1467 (81) |
| CC | 0.0582 (16) | 0.0686 (19) | 0.0634 (35) |
| CD | 0.1345 (37) | 0.1805 (50) | 0.1576 (87) |
| DD | 0.1345 (37) | 0.1011 (28) | 0.1178 (65) |
| Part B. Allele | |||
| A | 0.1836 | 0.2184 | 0.2011 |
| B | 0.2182 | 0.1823 | 0.2002 |
| C | 0.2582 | 0.2671 | 0.2627 |
| D | 0.3400 | 0.3321 | 0.3361 |
| χ2 | 0.0600 | 0.0093 | 0.0170 |
| Traits | Genotypes (sample size) | |||||
|---|---|---|---|---|---|---|
| AA (♂, 59) | AB (♂, 125) | BB (♂, 91) | AA (♀, 65) | AB (♀, 139) | BB (♀, 73) | |
| BW (g) | 2815.27 ± 321.42 aB | 3066.61 ± 320.29 Aa | 2951.79 ± 322.01 b | 2560.49 ± 280.82 B | 2646.65 ± 251.18 B | 2761.82 ± 283.31 A |
| BIL (cm) | 29.35 ± 2.14 | 29.92 ± 1.99 | 29.51 ± 1.82 | 27.99 ± 1.83 Bb | 28.36 ± 1.72 b | 28.84 ± 1.73 Aa |
| DL (cm) | 66.62 ± 3.43 b | 67.99 ± 3.44 Aa | 66.63 ±3.41 Bb | 63.43 ± 3.34 Bb | 64.07 ± 3.04 b | 65.01 ± 2.61 Aa |
| SC (cm) | 4.63 ± 0.32 | 4.96 ± 0.67 | 4.79 ± 0.82 | 4.48 ± 0.25 b | 4.57 ± 0.24 ab | 4.58 ± 0.26 a |
| CW (cm) | 10.81 ± 0.83 b | 11.10 ± 0.83 a | 10.87 ± 0.78 b | 10.53 ± 0.83 | 10.59 ± 0.81 | 10.57 ± 0.84 |
| CD (cm) | 7.13 ± 0.52 b | 7.19 ± 0.65 a | 7.02 ± 0.54 ab | 6.96 ± 0.47 Bb | 7.06 ± 0.51 b | 7.20 ± 0.48 Aa |
| SL (cm) | 11.92 ± 1.15 Bb | 12.46 ± 1.30 Aa | 12.10 ± 1.20 b | 11.54 ± 1.27 Bb | 11.87 ± 0.99 a | 12.11 ± 0.79 Aa |
| SHL (cm) | 10.79 ± 0.52 b | 10.95 ± 0.45 a | 10.85 ± 0.54 ab | 10.40 ± 0.55 | 10.36 ± 0.41 | 10.47 ± 0.41 |
| Traits | Genotypes (sample size) | |||||
|---|---|---|---|---|---|---|
| AA (♂, 12) | AB (♂, 38) | BB (♂, 33) | AA (♀, 26) | AB (♀, 39) | BB (♀, 16) | |
| CW (g) | 2520.35 ± 81.88 b | 2754.11 ± 43.19 a | 2591.40 ± 63.50 b | 2270.86 ± 46.75 B | 2430.19 ± 36.83 A | 2518.40 ± 43.32 A |
| SEW (g) | 2222.61 ± 67.64 b | 2439.32 ± 40.87 a | 2343.92 ± 53.39 ab | 2033.60 ± 40.92 | 2331.60 ± 147.94 | 2230.59 ± 46.24 |
| EW (g) | 1974.14 ± 62.83 b | 2163.79 ± 39.57 a | 2074.64 ± 48.97 ab | 1805.63 ± 37.27 | 2080.31 ± 147.43 | 1989.69 ± 43.37 |
| BLW (g) | 279.34 ± 13.86 | 286.16 ± 6.87 | 285.10 ± 8.78 | 268.00 ± 10.90 | 296.37 ± 12.60 | 287.33 ± 12.61 |
| HW (g) | 21.37 ± 0.65 b | 23.64 ± 0.48 a | 23.19 ± 0.60 ab | 20.96 ± 0.83 | 21.500 ± 54 | 22.66 ± 0.92 |
| LW (g) | 78.99±3.66 | 82.07 ± 2.29 | 83.34 ± 3.38 | 68.13 ± 2.94B | 77.69 ± 2.18A | 76.04 ± 3.10 AB |
| GSW (g) | 99.87 ± 2.69 B | 115.56 ± 2.53 A | 102.17 ± 2.93 B | 95.93 ± 3.13 | 97.18 ± 2.37 | 102.73 ± 2.88 |
| AW (g) | 50.60 ± 6.48 | 54.24 ± 4.46 | 60.56 ± 4.65 | 42.92 ± 4.34 | 54.90 ± 4.97 | 39.45 ± 5.94 |
| Traits | Genotypes (sample size) | |||
|---|---|---|---|---|
| AA (31) | AB (63) | BB (49) | ||
| Breast muscle | pH | 5.91 ± 0.19 | 5.93 ± 0.21 | 5.88 ± 0.18 |
| TD | 45.36 ± 17.43 | 45.78 ± 13.68 | 43.67 ± 21.15 | |
| HW | 0.30 ± 0.06 ab | 0.26 ± 0.08 b | 0.30 ± 0.10 a | |
| Leg muscle | pH | 5.99 ± 0.20 | 6.01 ± 0.25 | 5.98 ± 0.32 |
| TD | 31.82 ± 9.65 | 30.04 ± 11.70 | 29.88 ± 9.33 | |
| HW | 0.29 ± 0.05 ab | 0.27 ± 0.06 b | 0.30 ± 0.06 a | |
| Traitsμm2 | Genotypes (sample size) | |||||
|---|---|---|---|---|---|---|
| AA (♂, 12) | AB (♂, 38) | BB (♂, 33) | AA (♀, 26) | AB (♀, 39) | BB (♀, 16) | |
| BM | 274.46 ± 52.13 | 330.56 ± 59.13 | 310.46 ± 57.43 | 235.57 ± 49.39 | 241.56 ± 65.83 | 236.94 ± 51.83 |
| LM | 620.97 ± 146.80 | 670.05 ± 197.46 | 650.84 ± 143.73 | 409.48 ± 75.86 | 607.14 ± 165.18 | 478.07 ± 92.54 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Y.; Zhu, Z.; Xu, Q.; Chen, G. Association of Polymorphisms of Exon 2 of the Growth Hormone Gene with Production Performance in Huoyan Goose. Int. J. Mol. Sci. 2014, 15, 670-683. https://doi.org/10.3390/ijms15010670
Zhang Y, Zhu Z, Xu Q, Chen G. Association of Polymorphisms of Exon 2 of the Growth Hormone Gene with Production Performance in Huoyan Goose. International Journal of Molecular Sciences. 2014; 15(1):670-683. https://doi.org/10.3390/ijms15010670
Chicago/Turabian StyleZhang, Yang, Zhen Zhu, Qi Xu, and Guohong Chen. 2014. "Association of Polymorphisms of Exon 2 of the Growth Hormone Gene with Production Performance in Huoyan Goose" International Journal of Molecular Sciences 15, no. 1: 670-683. https://doi.org/10.3390/ijms15010670