Chemokine Receptors in Epithelial Ovarian Cancer

{kind=link}
{kind=link}
{kind=link}
Abstract
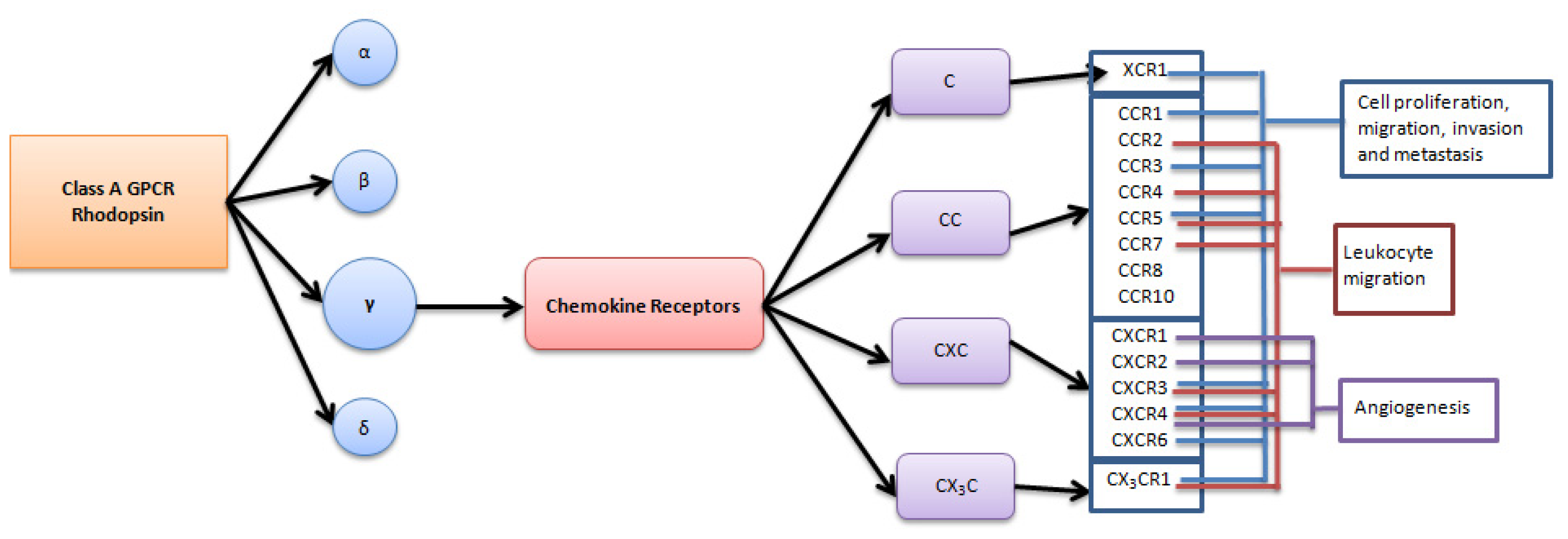
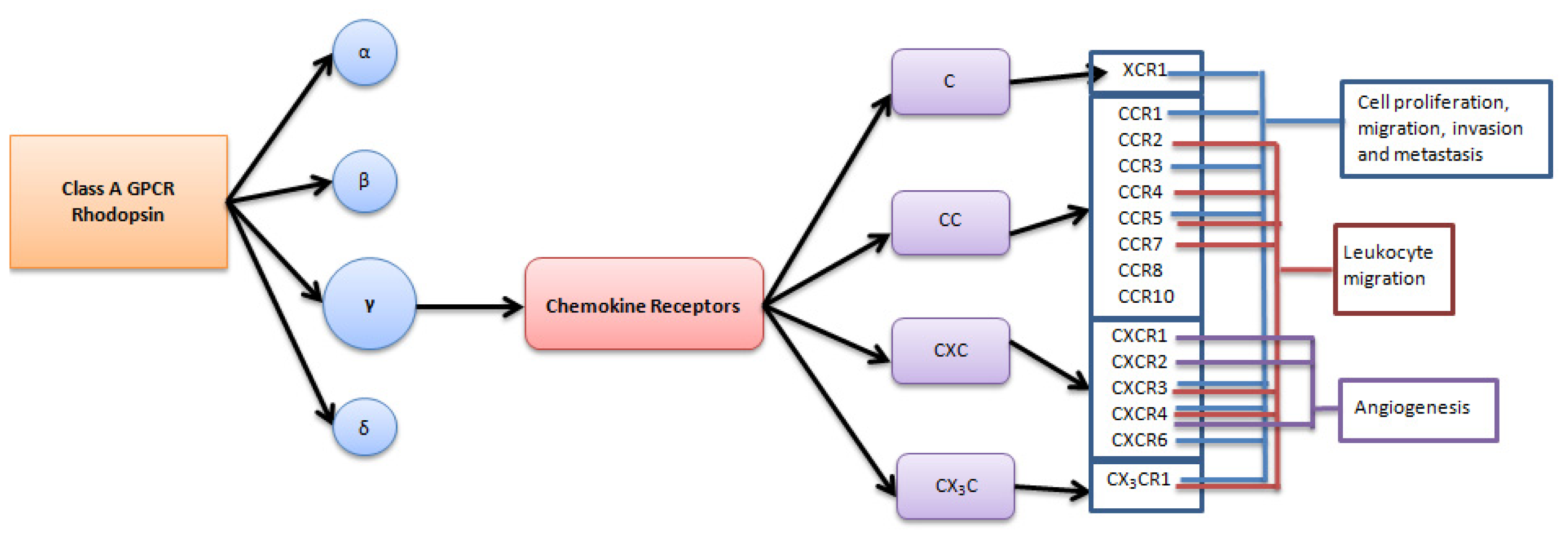
:1. Introduction
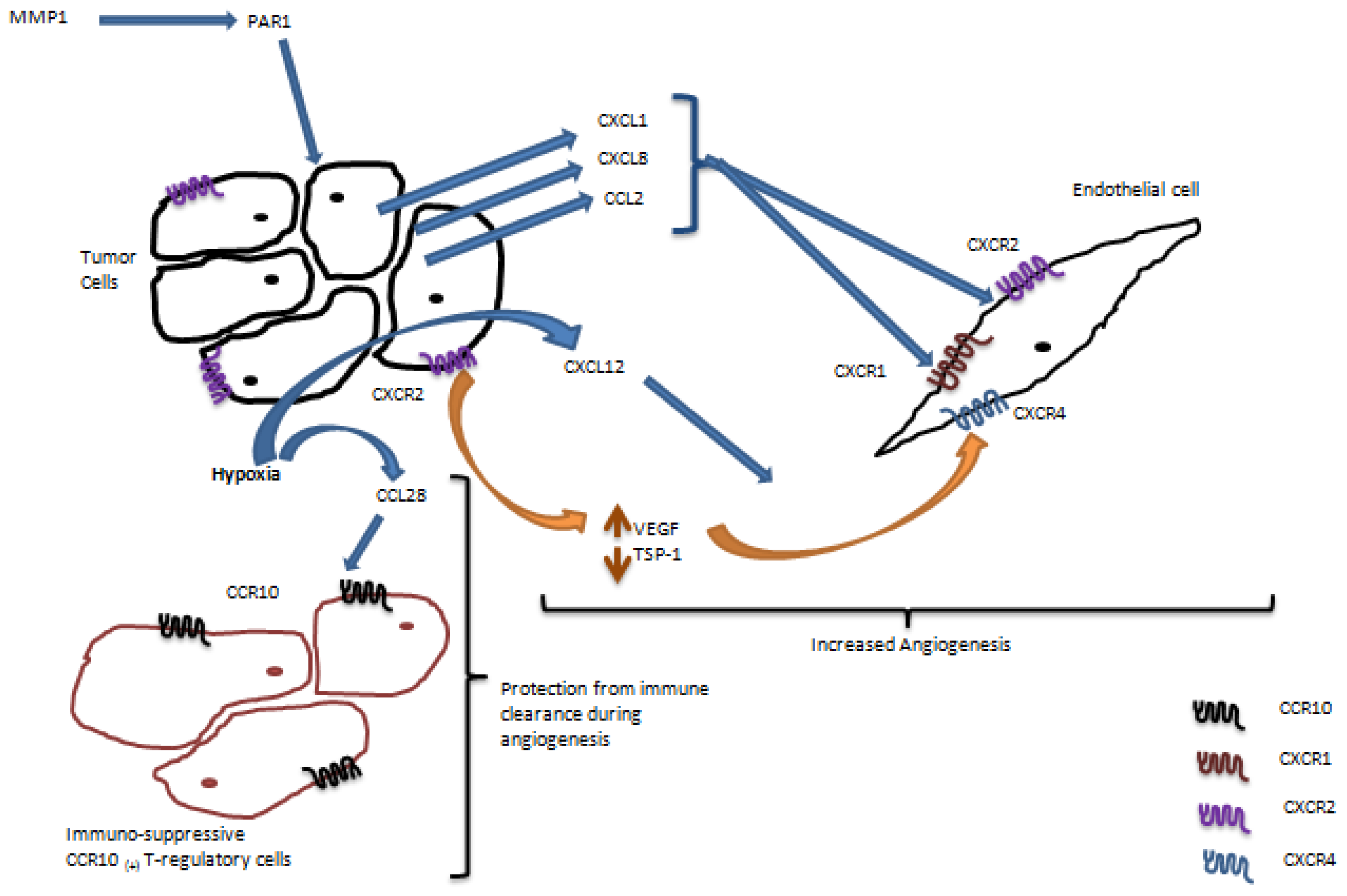
2. Role of Chemokine Receptors in Regulation of Angiogenesis in Ovarian Carcinoma
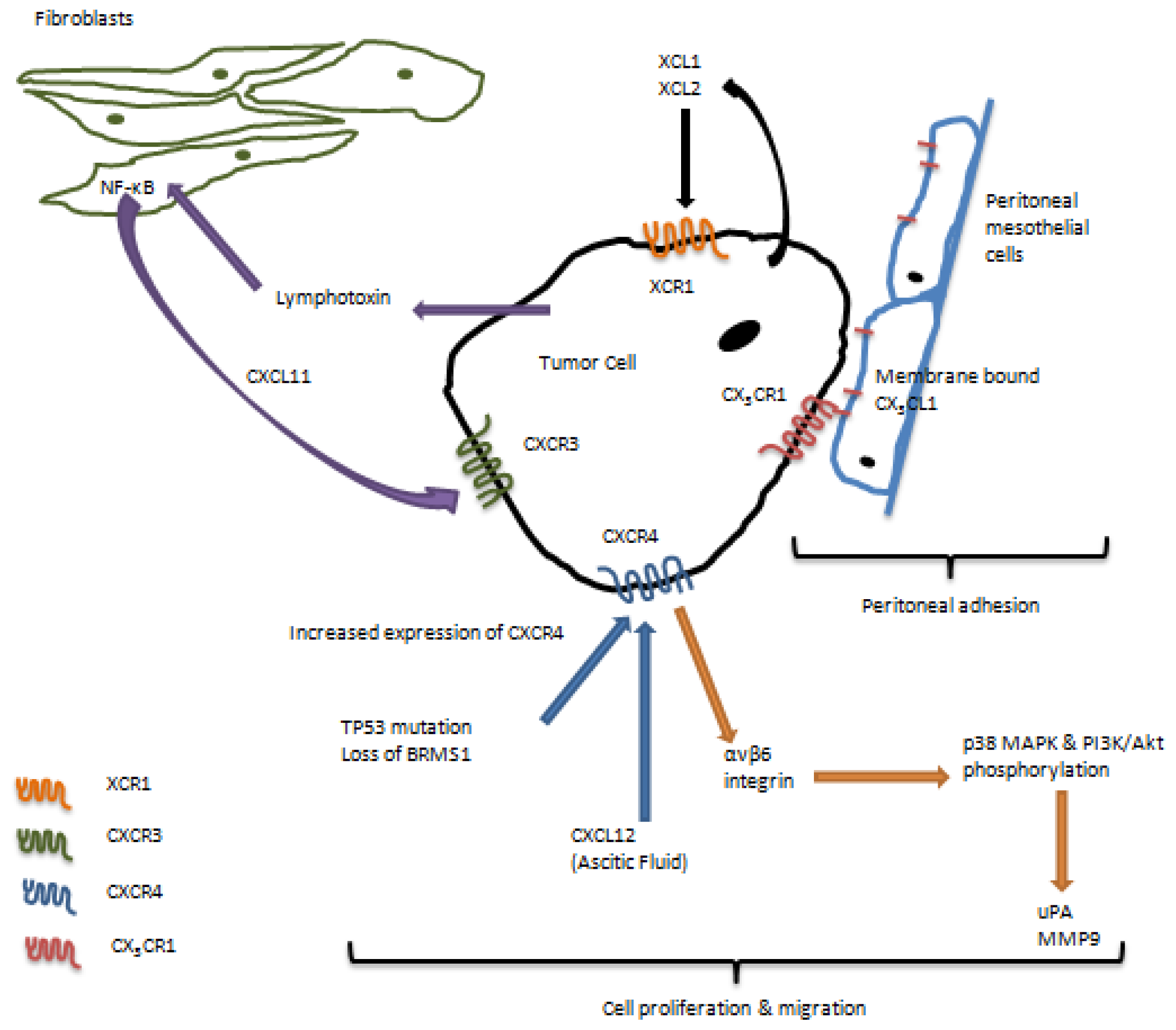
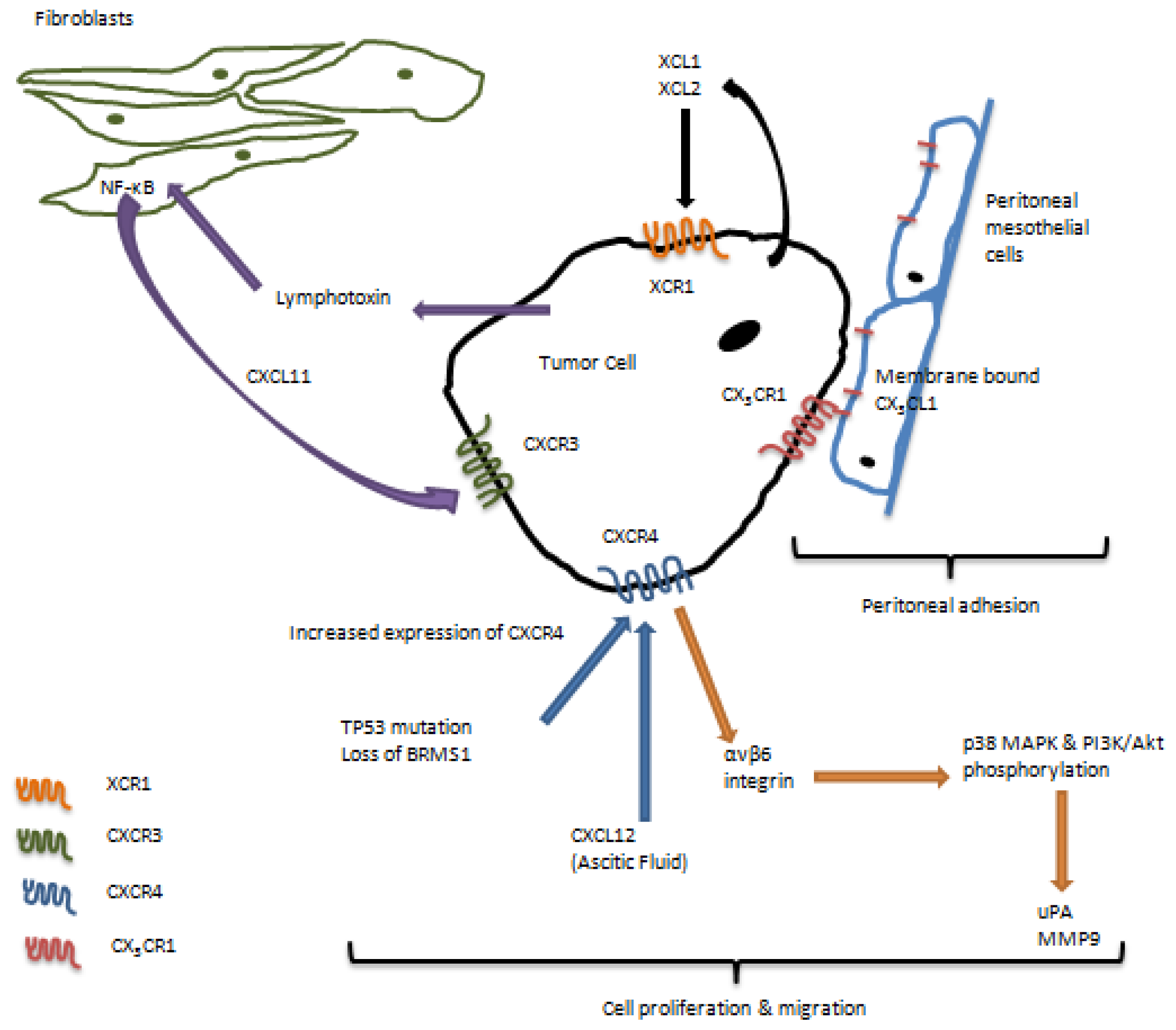
3. Role of Chemokine Receptors in Regulation of Ovarian Carcinoma Metastasis
4. Chemokine Receptors and the Immune System
5. Chemokine Receptor Inhibitors
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J. Clin 2013, 63, 11–30. [Google Scholar]
- American Cancer Society, American Cancer Society Cancer Facts & Figures 2013; American Cancer Society: Atlanta, GA, USA, 2013.
- Cannistra, S.A. Cancer of the ovary. N. Engl. J. Med 2004, 351, 2519–2529. [Google Scholar]
- Burleson, K.M.; Casey, R.C.; Skubitz, K.M.; Pambuccian, S.E.; Oegema, T.R., Jr.; Skubitz, A.P. Ovarian carcinoma ascites spheroids adhere to extracellular matrix components and mesothelial cell monolayers. Gynecol. Oncol 2004, 93, 170–181. [Google Scholar]
- Casey, R.C.; Skubitz, A.P. CD44 and beta1 integrins mediate ovarian carcinoma cell migration toward extracellular matrix proteins. Clin. Exp. Metastasis 2000, 18, 67–75. [Google Scholar]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol 2010, 177, 1053–1064. [Google Scholar]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol 2003, 63, 1256–1272. [Google Scholar]
- Fredriksson, R.; Schiöth, H.B. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol. Pharmacol 2005, 67, 1414–1425. [Google Scholar]
- Bjarnadóttir, T.K.; Gloriam, D.E.; Hellstrand, S.H.; Kristiansson, H.; Fredriksson, R.; Schiöth, H.B. Comprehensive repertoire and phylogenetic analysis of the G protein-coupled receptors in human and mouse. Genomics 2006, 88, 263–273. [Google Scholar]
- Lefkowitz, R.J. Seven transmembrane receptors: Something old, something new. Acta Physiol. (Oxf.) 2007, 190, 9–19. [Google Scholar]
- Attwood, T.K.; Findlay, J.B. Fingerprinting G-protein-coupled receptors. Protein Eng 1994, 7, 195–203. [Google Scholar]
- Kolakowski, L.F. GCRDb: A G-protein-coupled receptor database. Recept. Channels 1994, 2, 1–7. [Google Scholar]
- Foord, S.M.; Bonner, T.I.; Neubig, R.R.; Rosser, E.M.; Pin, J.P.; Davenport, A.P.; Spedding, M.; Harmar, A.J. International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol. Rev 2005, 57, 279–288. [Google Scholar]
- Joost, P.; Methner, A. Phylogenetic analysis of 277 human G-protein-coupled receptors as a tool for the prediction of orphan receptor ligands. Genome Biol 2002, 3. [Google Scholar] [CrossRef]
- Latek, D.; Modzelewska, A.; Trzaskowski, B.; Palczewski, K.; Filipek, S. G protein-coupled receptors—Recent advances. Acta Biochim. Pol 2012, 59, 515–529. [Google Scholar]
- Murphy, P.M.; Baggiolini, M.; Charo, I.F.; Hébert, C.A.; Horuk, R.; Matsushima, K.; Miller, L.H.; Oppenheim, J.J.; Power, C.A. International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol. Rev 2000, 52, 145–176. [Google Scholar]
- Szpakowska, M.; Fievez, V.; Arumugan, K.; van Nuland, N.; Schmit, J.C.; Chevigné, A. Function, diversity and therapeutic potential of the N-terminal domain of human chemokine receptors. Biochem. Pharmacol 2012, 84, 1366–1380. [Google Scholar]
- Clark-Lewis, I.; Kim, K.S.; Rajarathnam, K.; Gong, J.H.; Dewald, B.; Moser, B.; Baggiolini, M.; Sykes, B.D.; et al. Structure-activity relationships of chemokines. J. Leukoc Biol 1995, 57, 703–711. [Google Scholar]
- Fernandez, E.J.; Lolis, E. Structure, function, and inhibition of chemokines. Annu. Rev. Pharmacol. Toxicol 2002, 42, 469–499. [Google Scholar]
- Zlotnik, A.; Yoshie, O. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar]
- Kiefer, F.; Siekmann, A.F. The role of chemokines and their receptors in angiogenesis. Cell Mol. Life Sci 2011, 68, 2811–2830. [Google Scholar]
- Crump, M.P.; Gong, J.H.; Loetscher, P.; Rajarathnam, K.; Amara, A.; Arenzana-Seisdedos, F.; Virelizier, J.L.; Baggiolini, M.; Sykes, B.D.; Clark-Lewis, I. Solution structure and basis for functional activity of stromal cell-derived factor-1; dissociation of CXCR4 activation from binding and inhibition of HIV-1. EMBO J 1997, 16, 6996–7007. [Google Scholar]
- Monteclaro, F.S.; Charo, I.F. The amino-terminal extracellular domain of the MCP-1 receptor, but not the RANTES/MIP-1alpha receptor, confers chemokine selectivity. Evidence for a two-step mechanism for MCP-1 receptor activation. J. Biol. Chem 1996, 271, 19084–19092. [Google Scholar]
- Raman, D.; Sobolik-Delmaire, T.; Richmond, A. Chemokines in health and disease. Exp. Cell Res 2011, 317, 575–589. [Google Scholar]
- Neel, N.F.; Barzik, M.; Raman, D.; Sobolik-Delmaire, T.; Sai, J.; Ham, A.J.; Mernaugh, R.L.; Gertler, F.B.; Richmond, A. VASP is a CXCR2-interacting protein that regulates CXCR2-mediated polarization and chemotaxis. J. Cell Sci 2009, 122, 1882–1894. [Google Scholar]
- Raman, D.; Neel, N.F.; Sai, J.; Mernaugh, R.L.; Ham, A.J.; Richmond, A.J. Characterization of chemokine receptor CXCR2 interacting proteins using a proteomics approach to define the CXCR2 “chemosynapse”. Methods Enzymol 2009, 460, 315–330. [Google Scholar]
- Raman, D.; Sai, J.; Neel, N.F.; Chew, C.S.; Richmond, A. LIM and SH3 protein-1 modulates CXCR2-mediated cell migration. PLoS One 2010, 5, e10050. [Google Scholar]
- Neptune, E.R.; Bourne, H.R. Receptors induce chemotaxis by releasing the betagamma subunit of Gi, not by activating Gq or Gs. Proc. Natl. Acad. Sci. USA 1997, 94, 14489–14494. [Google Scholar]
- Schall, T.J.; Proudfoot, A.E. Overcoming hurdles in developing successful drugs targeting chemokine receptors. Nat. Rev. Immunol 2011, 11, 355–363. [Google Scholar]
- Springael, J.Y.; Urizar, E.; Parmentier, M. Dimerization of chemokine receptors and its functional consequences. Cytokine Growth Factor Rev 2005, 16, 611–623. [Google Scholar]
- Mellado, M.; Rodríguez-Frade, J.M.; Vila-Coro, A.J.; Fernández, S.; de Ana, A.M.; Jones, D.R.; Torán, J.L.; Martínez-A, C. Chemokine receptor homo- or heterodimerization activates distinct signaling pathways. EMBO J 2001, 20, 2497–2507. [Google Scholar]
- Chen, C.; Li, J.; Bot, G.; Szabo, I.; Rogers, T.J.; Liu-Chen, L.Y. Heterodimerization and cross-desensitization between the mu-opioid receptor and the chemokine CCR5 receptor. Eur. J. Pharmacol 2004, 483, 175–186. [Google Scholar]
- Nesmelova, I.V.; Sham, Y.; Dudek, A.Z.; van Eijk, L.I.; Wu, G.; Slungaard, A.; Mortari, F.; Griffioen, A.W.; Mayo, K.H. Platelet factor 4 and interleukin-8 CXC chemokine heterodimer formation modulates function at the quaternary structural level. J. Biol. Chem 2005, 280, 4948–4958. [Google Scholar]
- Porcile, C.; Bajetto, A.; Barbero, S.; Pirani, P.; Schettini, G. CXCR4 activation induces epidermal growth factor receptor transactivation in an ovarian cancer cell line. Ann. N. Y. Acad. Sci 2004, 1030, 162–169. [Google Scholar]
- Porcile, C.; Bajetto, A.; Barbieri, F.; Barbero, S.; Bonavia, R.; Biglieri, M.; Pirani, P.; Florio, T.; Schettini, G. Stromal cell-derived factor-1alpha (SDF-1alpha/CXCL12) stimulates ovarian cancer cell growth through the EGF receptor transactivation. Exp. Cell. Res 2005, 308, 241–253. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- Gavalas, N.G.; Liontos, M.; Trachana, S.P.; Bagratuni, T.; Arapinis, C.; Liacos, C.; Dimopoulos, M.A.; Bamias, A. Angiogenesis-related pathways in the pathogenesis of ovarian cancer. Int. J. Mol. Sci 2013, 14, 15885–15909. [Google Scholar]
- Musrap, N.; Diamandis, E.P. Revisiting the complexity of the ovarian cancer microenvironment—Clinical implications for treatment strategies. Mol. Cancer Res 2012, 10, 1254–1264. [Google Scholar]
- Bamias, A.; Pignata, S.; Pujade-Lauraine, E. Angiogenesis: A promising therapeutic target for ovarian cancer. Crit. Rev. Oncol. Hematol 2012, 84, 314–326. [Google Scholar]
- Salcedo, R.; Resau, J.H.; Halverson, D.; Hudson, E.A.; Dambach, M.; Powell, D.; Wasserman, K.; Oppenheim, J.J. Differential expression and responsiveness of chemokine receptors (CXCR1–3) by human microvascular endothelial cells and umbilical vein endothelial cells. FASEB J 2000, 14, 2055–2064. [Google Scholar]
- Coughlin, S.R. Thrombin signalling and protease-activated receptors. Nature 2000, 407, 258–264. [Google Scholar]
- Agarwal, A.; Tressel, S.L.; Kaimal, R.; Balla, M.; Lam, F.H.; Covic, L.; Kuliopulos, A. Identification of a metalloprotease-chemokine signaling system in the ovarian cancer microenvironment: Implications for antiangiogenic therapy. Cancer Res 2010, 70, 5880–5890. [Google Scholar]
- Lokshin, A.E.; Winans, M.; Landsittel, D.; Marrangoni, A.M.; Velikokhatnaya, L.; Modugno, F.; Nolen, B.M.; Gorelik, E. Circulating IL-8 and anti-IL-8 autoantibody in patients with ovarian cancer. Gynecol. Oncol 2006, 102, 244–251. [Google Scholar]
- Merritt, W.M.; Lin, Y.G.; Spannuth, W.A.; Fletcher, M.S.; Kamat, A.A.; Han, L.Y.; Landen, C.N.; Jennings, N.; De Geest, K.; Langley, R.R.; et al. Effect of interleukin-8 gene silencing with liposome-encapsulated small interfering RNA on ovarian cancer cell growth. J. Natl. Cancer Inst 2008, 100, 359–372. [Google Scholar]
- Yang, G.; Rosen, D.G.; Liu, G.; Yang, F.; Guo, X.; Xiao, X.; Xue, F.; Mercado-Uribe, I.; Huang, J.; Lin, S.H.; et al. CXCR2 promotes ovarian cancer growth through dysregulated cell cycle, diminished apoptosis, and enhanced angiogenesis. Clin. Cancer Res 2010, 16, 3875–3886. [Google Scholar]
- Kryczek, I.; Lange, A.; Mottram, P.; Alvarez, X.; Cheng, P.; Hogan, M.; Moons, L.; Wei, S.; Zou, L.; Machelon, V.; et al. CXCL12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers. Cancer Res 2005, 65, 465–472. [Google Scholar]
- Righi, E.; Kashiwagi, S.; Yuan, J.; Santosuosso, M.; Leblanc, P.; Ingraham, R.; Forbes, B.; Edelblute, B.; Collette, B.; Xing, D.; et al. CXCL12/CXCR4 blockade induces multimodal antitumor effects that prolong survival in an immunocompetent mouse model of ovarian cancer. Cancer Res 2011, 71, 5522–5534. [Google Scholar]
- Lotfi, R.; Lee, J.J.; Lotze, M.T. Eosinophilic granulocytes and damage-associated molecular pattern molecules (DAMPs, role in the inflammatory response within tumors. J. Immunother 2007, 30, 16–28. [Google Scholar]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.P.; Gimotty, P.A.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumour hypoxia promotes tolerance and angiogenesis via CCL28 and T(reg) cells. Nature 2011, 475, 226–230. [Google Scholar]
- Sakaguchi, S. Regulatory T cells: Key controllers of immunologic self-tolerance. Cell 2000, 101, 455–458. [Google Scholar]
- Burleson, K.M.; Hansen, L.K.; Skubitz, A.P. Ovarian carcinoma spheroids disaggregate on type I collagen and invade live human mesothelial cell monolayers. Clin. Exp. Metastasis 2004, 21, 685–697. [Google Scholar]
- Iwanicki, M.P.; Davidowitz, R.A.; Ng, M.R.; Besser, A.; Muranen, T.; Merritt, M.; Danuser, G.; Ince, T.A.; Brugge, J.S. Ovarian cancer spheroids use myosin-generated force to clear the mesothelium. Cancer Discov 2011, 1, 144–157. [Google Scholar]
- Davidowitz, R.A.; Iwanicki, M.P.; Brugge, J.S. In vitro mesothelial clearance assay that models the early steps of ovarian cancer metastasis. J. Vis. Exp 2012. [Google Scholar] [CrossRef]
- Kim, M.; Rooper, L.; Xie, J.; Kajdacsy-Balla, A.A.; Barbolina, M.V. Fractalkine receptor CX(3)CR1 is expressed in epithelial ovarian carcinoma cells and required for motility and adhesion to peritoneal mesothelial cells. Mol. Cancer Res 2012, 10, 11–24. [Google Scholar]
- Bazan, J.F.; Bacon, K.B.; Hardiman, G.; Wang, W.; Soo, K.; Rossi, D.; Greaves, D.R.; Zlotnik, A.; Schall, T.J. A new class of membrane-bound chemokine with a CX3C motif. Nature 1997, 385, 640–644. [Google Scholar]
- Gaudin, F.; Nasreddine, S.; Donnadieu, A.C.; Emilie, D.; Combadière, C.; Prévot, S.; Machelon, V.; Balabanian, K. Identification of the chemokine CX3CL1 as a new regulator of malignant cell proliferation in epithelial ovarian cancer. PLoS One 2011, 6, e21546. [Google Scholar]
- Kim, M.; Rooper, L.; Xie, J.; Rayahin, J.; Burdette, J.E.; Kajdacsy-Balla, A.A.; Barbolina, M.V. The lymphotactin receptor is expressed in epithelial ovarian carcinoma and contributes to cell migration and proliferation. Mol. Cancer Res 2012, 10, 1419–1429. [Google Scholar]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar]
- Junttila, M.R.; de Sauvage, F.J. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 2013, 501, 346–354. [Google Scholar]
- Schauer, I.G.; Sood, A.K.; Mok, S.; Liu, J. Cancer-associated fibroblasts and their putative role in potentiating the initiation and development of epithelial ovarian cancer. Neoplasia 2011, 13, 393–405. [Google Scholar]
- Lau, T.S.; Chung, T.K.; Cheung, T.H.; Chan, L.K.; Cheung, L.W.; Yim, S.F.; Siu, N.S.; Lo, K.W.; Yu, M.M.; Kulbe, H.; et al. Cancer cell-derived lymphotoxin mediates reciprocal tumour-stromal interactions in human ovarian cancer by inducing CXCL11 in fibroblasts. J. Pathol 2013, 232, 43–56. [Google Scholar]
- Cho, K.R.; Shih, I.M. Ovarian cancer. Annu. Rev. Pathol 2009, 4, 287–313. [Google Scholar]
- Archibald, K.M.; Kulbe, H.; Kwong, J.; Chakravarty, P.; Temple, J.; Chaplin, T.; Flak, M.B.; McNeish, I.A.; Deen, S.; Brenton, J.D.; et al. Sequential genetic change at the TP53 and chemokine receptor CXCR4 locus during transformation of human ovarian surface epithelium. Oncogene 2012, 31, 4987–4995. [Google Scholar]
- Guo, L.; Cui, Z.M.; Zhang, J.; Huang, Y. Chemokine axes CXCL12/CXCR4 and CXCL16/CXCR6 correlate with lymph node metastasis in epithelial ovarian carcinoma. Chin. J. Cancer 2011, 30, 336–343. [Google Scholar]
- Guo, Z.; Cai, S.; Fang, R.; Chen, H.; Du, J.; Tan, Y.; Ma, W.; Hu, H.; Liu, Y. The synergistic effects of CXCR4 and EGFR on promoting EGF-mediated metastasis in ovarian cancer cells. Colloids Surf. B 2007, 60, 1–6. [Google Scholar]
- Scotton, C.J.; Wilson, J.L.; Milliken, D.; Stamp, G.; Balkwill, F.R. Epithelial cancer cell migration: A role for chemokine receptors? Cancer Res 2001, 61, 4961–4965. [Google Scholar]
- Sheng, X.J.; Zhou, Y.Q.; Song, Q.Y.; Zhou, D.M.; Liu, Q.C. Loss of breast cancer metastasis suppressor 1 promotes ovarian cancer cell metastasis by increasing chemokine receptor 4 expression. Oncol. Rep 2012, 27, 1011–1018. [Google Scholar]
- Xue, B.; Wu, W.; Huang, K.; Xie, T.; Xu, X.; Zhang, H.; Qi, C.; Ge, J.; Yu, Y. Stromal cell-derived factor-1 (SDF-1) enhances cells invasion by αvβ6 integrin-mediated signaling in ovarian cancer. Mol. Cell Biochem 2013, 380, 177–184. [Google Scholar]
- Barbolina, M.V.; Kim, M.; Liu, Y.; Shepard, J.; Belmadani, A.; Miller, R.J.; Shea, L.D.; Stack, M.S. Microenvironmental regulation of chemokine (C-X-C-motif) receptor 4 in ovarian carcinoma. Mol. Cancer Res 2010, 8, 653–664. [Google Scholar]
- Scotton, C.J.; Wilson, J.L.; Scott, K.; Stamp, G.; Wilbanks, G.D.; Fricker, S.; Bridger, G.; Balkwill, F.R. Multiple actions of the chemokine CXCL12 on epithelial tumor cells in human ovarian cancer. Cancer Res 2002, 62, 5930–5938. [Google Scholar]
- Miyanishi, N.; Suzuki, Y.; Simizu, S.; Kuwabara, Y.; Banno, K.; Umezawa, K. Involvement of autocrine CXCL12/CXCR4 system in the regulation of ovarian carcinoma cell invasion. Biochem. Biophys. Res. Commun 2010, 403, 154–159. [Google Scholar]
- Long, H.; Xie, R.; Xiang, T.; Zhao, Z.; Lin, S.; Liang, Z.; Chen, Z.; Zhu, B. Autocrine CCL5 signaling promotes invasion and migration of CD133+ ovarian cancer stem-like cells via NF-κB-mediated MMP-9 upregulation. Stem Cells 2012, 30, 2309–2319. [Google Scholar]
- Kajiyama, H.; Shibata, K.; Terauchi, M.; Ino, K.; Nawa, A.; Kikkawa, F. Involvement of SDF-1alpha/CXCR4 axis in the enhanced peritoneal metastasis of epithelial ovarian carcinoma. Int. J. Cancer 2008, 122, 91–99. [Google Scholar]
- Kwong, J.; Kulbe, H.; Wong, D.; Chakravarty, P.; Balkwill, F. An antagonist of the chemokine receptor CXCR4 induces mitotic catastrophe in ovarian cancer cells. Mol. Cancer Ther 2009, 8, 1893–1905. [Google Scholar]
- Balabanian, K.; Lagane, B.; Infantino, S.; Chow, K.Y.; Harriague, J.; Moepps, B.; Arenzana-Seisdedos, F.; Thelen, M.; Bachelerie, F. The chemokine SDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 in T lymphocytes. J. Biol. Chem 2005, 280, 35760–35766. [Google Scholar]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T cells, recurrence, and survival in epithelial ovarian cancer. N. Engl. J. Med 2003, 348, 203–213. [Google Scholar]
- Mhawech-Fauceglia, P.; Wang, D.; Ali, L.; Lele, S.; Huba, M.A.; Liu, S.; Odunsi, K. Intraepithelial T cells and tumor-associated macrophages in ovarian cancer patients. Cancer Immun 2013, 13, 1. [Google Scholar]
- Kandalaft, L.E.; Motz, G.T.; Duraiswamy, J.; Coukos, G. Tumor immune surveillance and ovarian cancer: Lessons on immune mediated tumor rejection or tolerance. Cancer Metastasis Rev 2011, 30, 141–151. [Google Scholar]
- Scarlett, U.K.; Conejo-Garcia, J.R. Double agents in the war on cancer: Leukocytes govern ovarian cancer progression. Oncotarget 2012, 3, 226–227. [Google Scholar]
- Knutson, K.L.; Curiel, T.J.; Salazar, L.; Disis, M.L. Immunologic principles and immunotherapeutic approaches in ovarian cancer. Hematol. Oncol. Clin. N. Am 2003, 17, 1051–1073. [Google Scholar]
- Preston, C.C.; Goode, E.L.; Hartmann, L.C.; Kalli, K.R.; Knutson, K.L. Immunity and immune suppression in human ovarian cancer. Immunotherapy 2011, 3, 539–556. [Google Scholar]
- Yigit, R.; Massuger, L.F.; Figdor, C.G.; Torensma, R. Ovarian cancer creates a suppressive microenvironment to escape immune elimination. Gynecol. Oncol 2010, 117, 366–372. [Google Scholar]
- Fialová, A.; Partlová, S.; Sojka, L.; Hromádková, H.; Brtnický, T.; Fučíková, J.; Kocián, P.; Rob, L.; Bartůňková, J.; Spíšek, R. Dynamics of T-cell infiltration during the course of ovarian cancer: The gradual shift from a Th17 effector cell response to a predominant infiltration by regulatory T-cells. Int. J. Cancer 2013, 132, 1070–1079. [Google Scholar]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med 2004, 10, 942–949. [Google Scholar]
- Redjimi, N.; Raffin, C.; Raimbaud, I.; Pignon, P.; Matsuzaki, J.; Odunsi, K.; Valmori, D.; Ayyoub, M. CXCR3+ T regulatory cells selectively accumulate in human ovarian carcinomas to limit type I immunity. Cancer Res 2012, 72, 4351–4360. [Google Scholar]
- Wei, S.; Kryczek, I.; Zou, L.; Daniel, B.; Cheng, P.; Mottram, P.; Curiel, T.; Lange, A.; Zou, W. Plasmacytoid dendritic cells induce CD8+ regulatory T cells in human ovarian carcinoma. Cancer Res 2005, 65, 5020–5026. [Google Scholar]
- Burke, F.; Relf, M.; Negus, R.; Balkwill, F. A cytokine profile of normal and malignant ovary. Cytokine 1996, 8, 578–585. [Google Scholar]
- Negus, R.P.; Stamp, G.W.; Relf, M.G.; Burke, F.; Malik, S.T.; Bernasconi, S.; Allavena, P.; Sozzani, S.; Mantovani, A.; Balkwill, F.R. The detection and localization of monocyte chemoattractant protein-1 (MCP-1) in human ovarian cancer. J. Clin. Invest 1995, 95, 2391–2396. [Google Scholar]
- Sica, A.; Saccani, A.; Bottazzi, B.; Bernasconi, S.; Allavena, P.; Gaetano, B.; Fei, F.; LaRosa, G.; Scotton, C.; Balkwill, F.; et al. Defective expression of the monocyte chemotactic protein-1 receptor CCR2 in macrophages associated with human ovarian carcinoma. J. Immunol 2000, 164, 733–738. [Google Scholar]
- Furukawa, S.; Soeda, S.; Kiko, Y.; Suzuki, O.; Hashimoto, Y.; Watanabe, T.; Nishiyama, H.; Tasaki, K.; Hojo, H.; Abe, M.; et al. MCP-1 promotes invasion and adhesion of human ovarian cancer cells. Anticancer Res 2013, 33, 4785–4790. [Google Scholar]
- Szlosarek, P.W.; Balkwill, F.R. Tumour necrosis factor alpha: A potential target for the therapy of solid tumours. Lancet Oncol 2003, 4, 565–473. [Google Scholar]
- Kulbe, H.; Hagemann, T.; Szlosarek, P.W.; Balkwill, F.R.; Wilson, J.L. The inflammatory cytokine tumor necrosis factor-alpha regulates chemokine receptor expression on ovarian cancer cells. Cancer Res 2005, 65, 10355–10362. [Google Scholar]
- Obermajer, N.; Muthuswamy, R.; Odunsi, K.; Edwards, R.P.; Kalinski, P. PGE(2)-induced CXCL12 production and CXCR4 expression controls the accumulation of human MDSCs in ovarian cancer environment. Cancer Res 2011, 71, 7463–7470. [Google Scholar]
- Zou, W.; Machelon, V.; Coulomb-L’Hermin, A.; Borvak, J.; Nome, F.; Isaeva, T.; Wei, S.; Krzysiek, R.; Durand-Gasselin, I.; Gordon, A.; et al. Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat. Med 2001, 7, 1339–1346. [Google Scholar]
- Hart, K.M.; Bak, S.P.; Alonso, A.; Berwin, B. Phenotypic and functional delineation of murine CX(3)CR1 monocyte-derived cells in ovarian cancer. Neoplasia 2009, 11, 564–573, 1 p following 573. [Google Scholar]
- Wong, J.L.; Berk, E.; Edwards, R.P.; Kalinski, P. IL-18-primed helper NK cells collaborate with dendritic cells to promote recruitment of effector CD8+ T cells to the tumor microenvironment. Cancer Res 2013, 73, 4653–4662. [Google Scholar]
- Nesbeth, Y.; Scarlett, U.; Cubillos-Ruiz, J.; Martinez, D.; Engle, X.; Turk, M.J.; Conejo-Garcia, J.R. CCL5-mediated endogenous antitumor immunity elicited by adoptively transferred lymphocytes and dendritic cell depletion. Cancer Res 2009, 69, 6331–6338. [Google Scholar]
- Marchesi, F.; Grizzi, F.; Laghi, L.; Mantovani, A.; Allavena, P. Molecular mechanisms of pancreatic cancer dissemination: The role of the chemokine system. Curr. Pharm. Des 2012, 18, 2432–2438. [Google Scholar]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. Chemokine receptors as HIV-1 coreceptors: Roles in viral entry, tropism, and disease. Annu. Rev. Immunol 1999, 17, 657–700. [Google Scholar]
- Panzer, U.; Stahl, R.A. Chemokines and renal inflammation. Nephrologie 1999, 20, 335–241. [Google Scholar]
- Proudfoot, A.E.; Power, C.A.; Wells, T.N. The strategy of blocking the chemokine system to combat disease. Immunol. Rev 2000, 177, 246–256. [Google Scholar]
- Koelink, P.J.; Overbeek, S.A.; Braber, S.; de Kruijf, P.; Folkerts, G.; Smit, M.J.; Kraneveld, A.D. Targeting chemokine receptors in chronic inflammatory diseases: An extensive review. Pharmacol. Ther 2012, 133, 1–18. [Google Scholar]
- Pease, J.E. Asthma, allergy and chemokines. Curr. Drug Targets 2006, 7, 3–12. [Google Scholar]
- Slettenaar, V.I.; Wilson, J.L. The chemokine network: A target in cancer biology? Adv. Drug Deliv. Rev 2006, 58, 962–974. [Google Scholar]
- Wells, T.N.; Power, C.A.; Proudfoot, A.E. Definition, function and pathophysiological significance of chemokine receptors. Trends Pharmacol. Sci 1998, 19, 376–380. [Google Scholar]
- Pease, J.; Horuk, R. Chemokine receptor antagonists. J. Med. Chem 2012, 55, 9363–9392. [Google Scholar]
- MacArthur, R.D.; Novak, R.M. Reviews of anti-infective agents: Maraviroc: The first of a new class of antiretroviral agents. Clin. Infect. Dis 2008, 47, 236–241. [Google Scholar]
- De Clercq, E. The AMD3100 story: The path to the discovery of a stem cell mobilizer (Mozobil). Biochem. Pharmacol 2009, 77, 1655–1664. [Google Scholar]
- Brave, M.; Farrell, A.; Ching Lin, S.; Ocheltree, T.; Pope Miksinski, S.; Lee, S.L.; Saber, H.; Fourie, J.; Tornoe, C.; Booth, B.; et al. FDA review summary: Mozobil in combination with granulocyte colony-stimulating factor to mobilize hematopoietic stem cells to the peripheral blood for collection and subsequent autologous transplantation. Oncology 2010, 78, 282–288. [Google Scholar]
- Phase II Study of KW-0761 in Subjects With CCR4-positive Adult T-cell Leukemia-lymphoma. Available online: http://clinicaltrials.gov/show/NCT00920790 (accessed on 18 November 2013).
- Study of KW-0761 (Mogamulizumab) in Subjects With Previously Treated Peripheral T-cell Lymphoma (PTCL). Available online: http://clinicaltrials.gov/show/NCT01611142 (accessed on 18 November 2013).
- Safety Study of a Chemokine Receptor (CXCR4) Antagonist in Multiple Myeloma Patients. Available online: http://clinicaltrials.gov/show/NCT01010880 (accessed on 18 November 2013).
- Role of CXCR2 Ligands/CXCR2 Biological Axis in Pancreatic Cancer. Available online: http://clinicaltrials.gov/show/NCT00851955 (accessed on 18 November 2013).
- The Role of Stromal Cell-Derived Factor-1 (SDF-1)/CXC Chemokine Receptor 4 (CXCR4) in Metastasis of Laryngeal and Hypopharyngeal Squamous Cell Carcinoma. Available online: http://clinicaltrials.gov/show/NCT00174096 (accessed on 18 November 2013).

© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Muralidhar, G.G.; Barbolina, M.V. Chemokine Receptors in Epithelial Ovarian Cancer. Int. J. Mol. Sci. 2014, 15, 361-376. https://doi.org/10.3390/ijms15010361
Muralidhar GG, Barbolina MV. Chemokine Receptors in Epithelial Ovarian Cancer. International Journal of Molecular Sciences. 2014; 15(1):361-376. https://doi.org/10.3390/ijms15010361
Chicago/Turabian StyleMuralidhar, Goda G., and Maria V. Barbolina. 2014. "Chemokine Receptors in Epithelial Ovarian Cancer" International Journal of Molecular Sciences 15, no. 1: 361-376. https://doi.org/10.3390/ijms15010361