Role of Insulin-Transferrin-Selenium in Auricular Chondrocyte Proliferation and Engineered Cartilage Formation in Vitro


Abstract
:1. Introduction
2. Results and Discussion
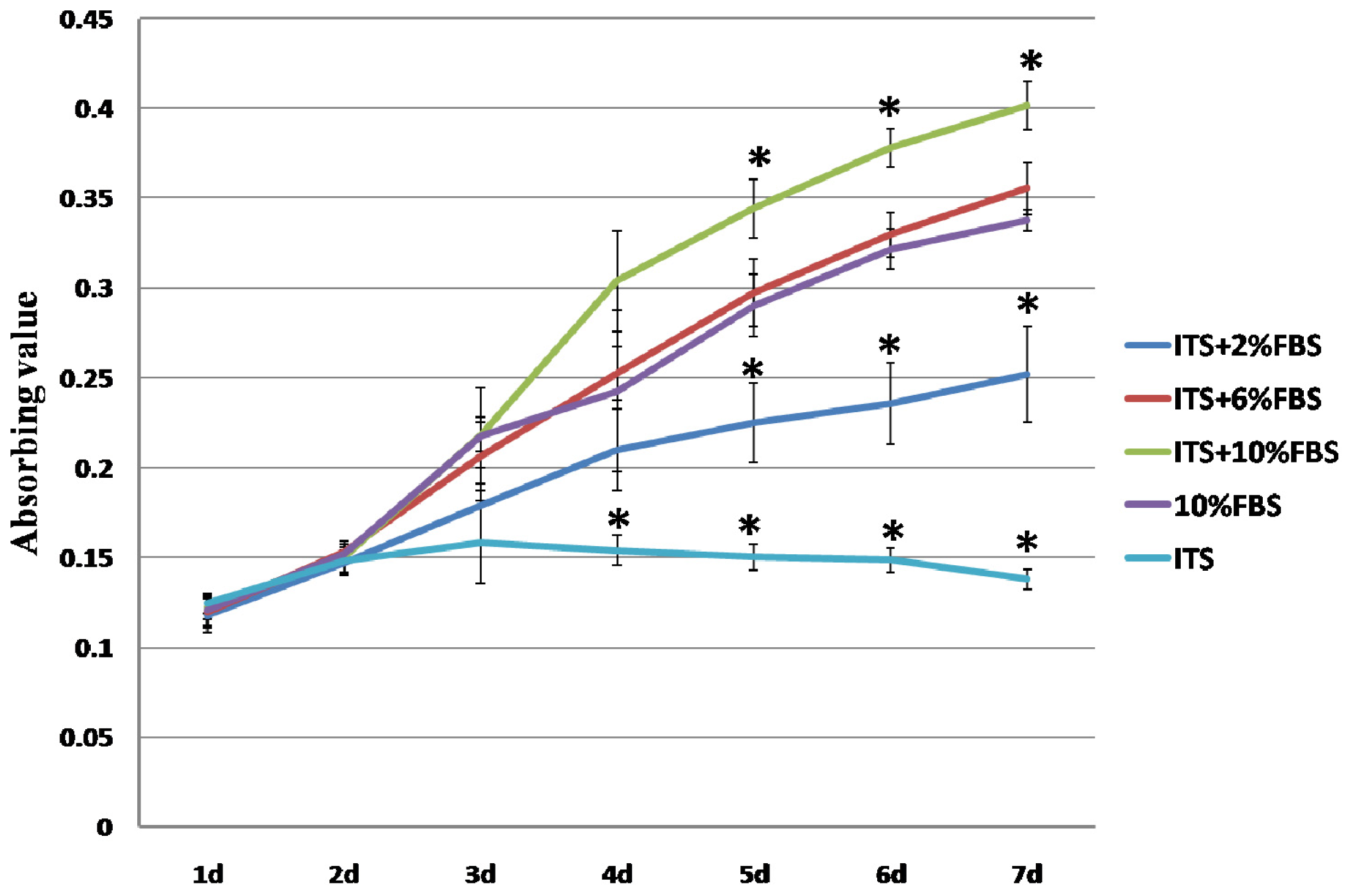
2.1. Expansion of Auricular Chondrocytes in Monolayer Culture
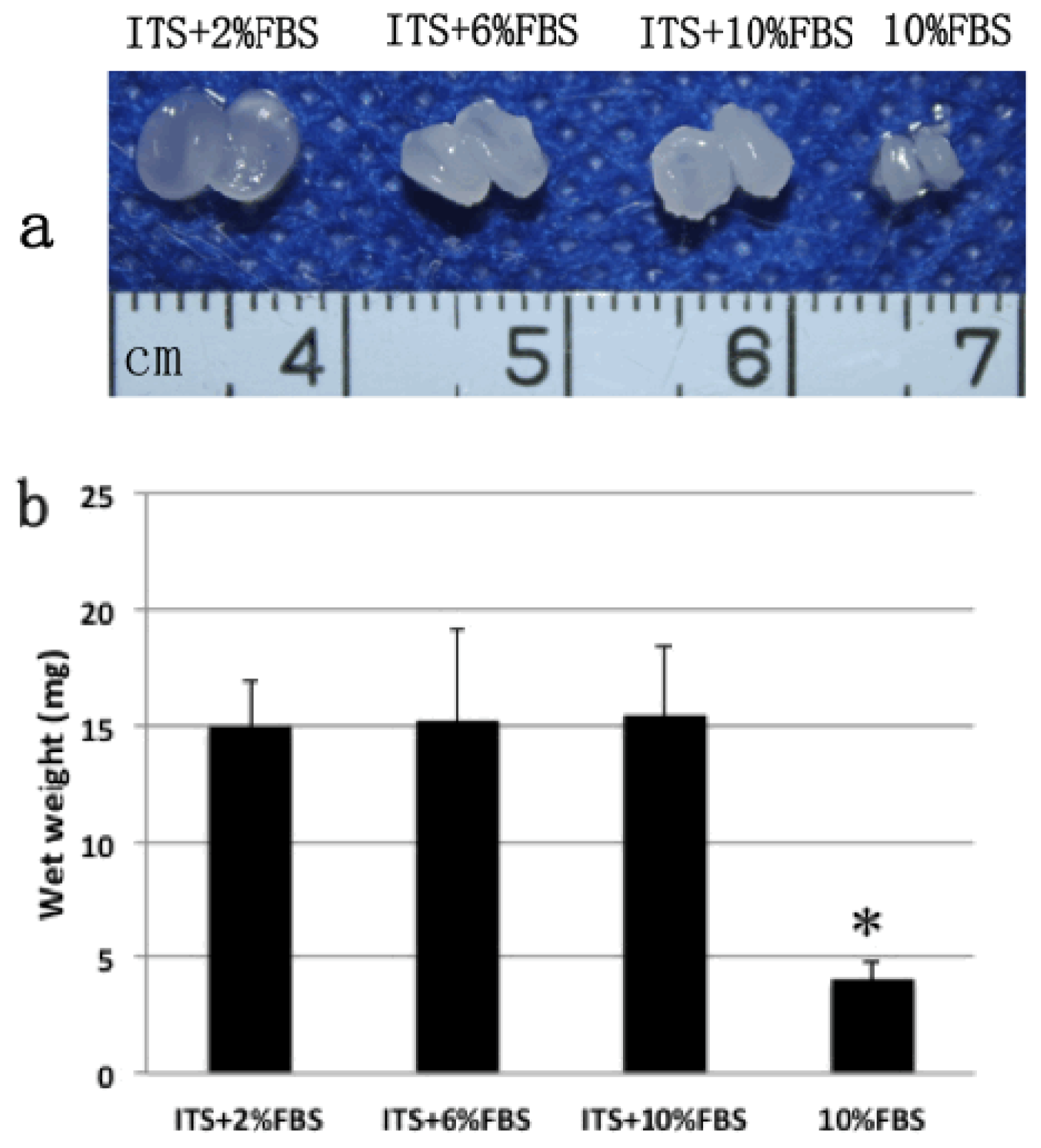
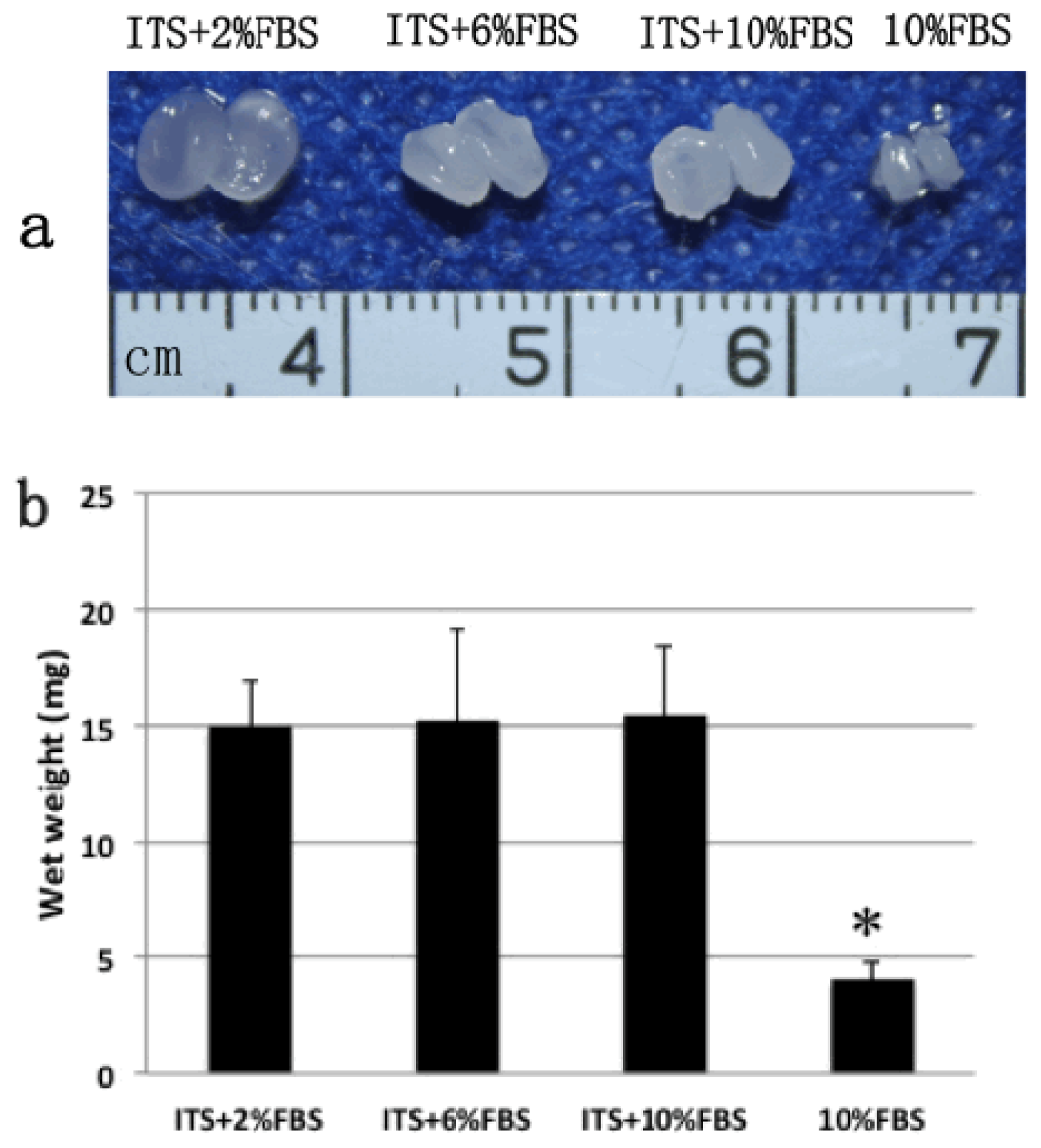
2.2. Gross View and Wet Weight of Auricular Chondrocyte Pellets
2.3. Histological and Immunohistochemical Analysis
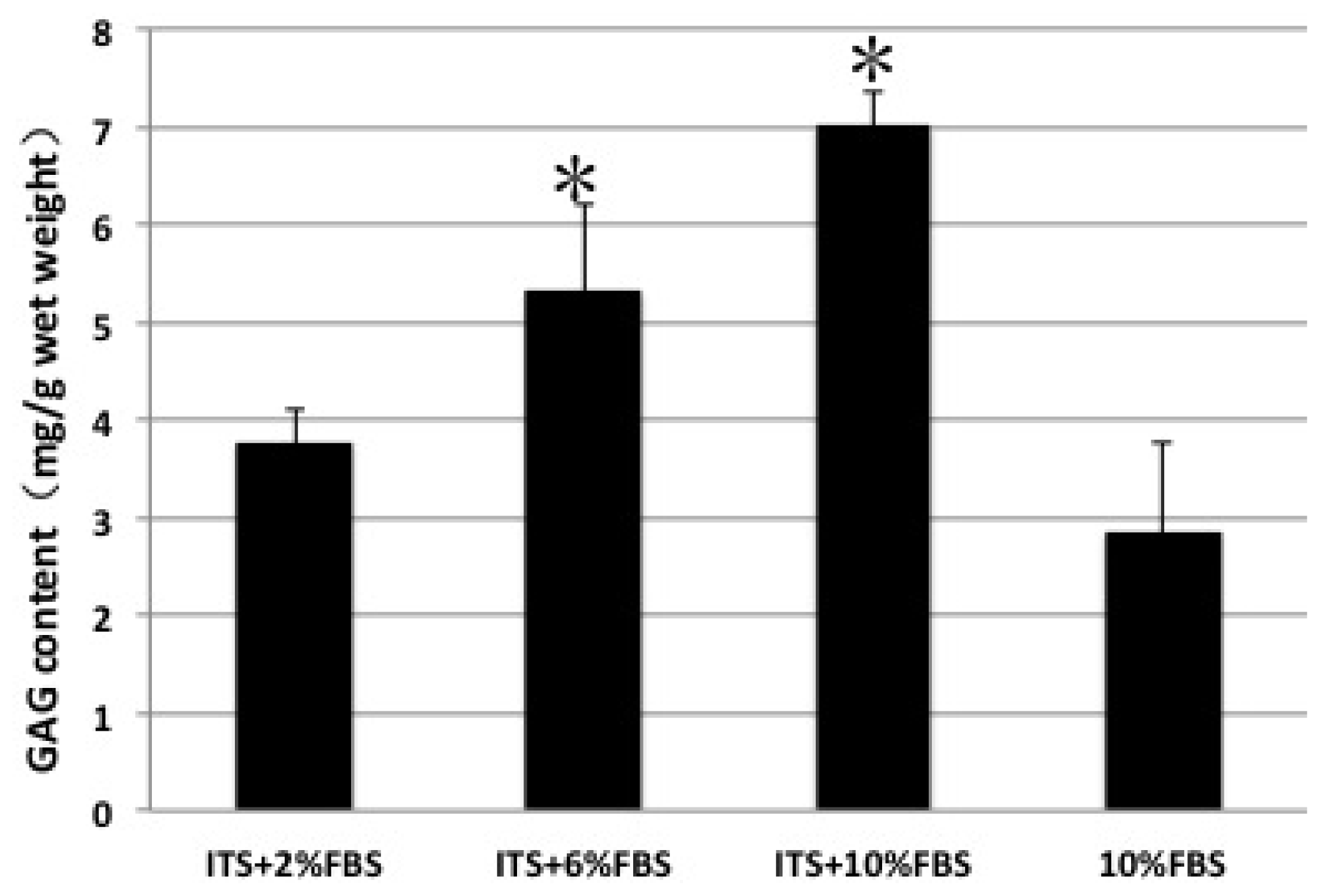
2.4. GAG Content
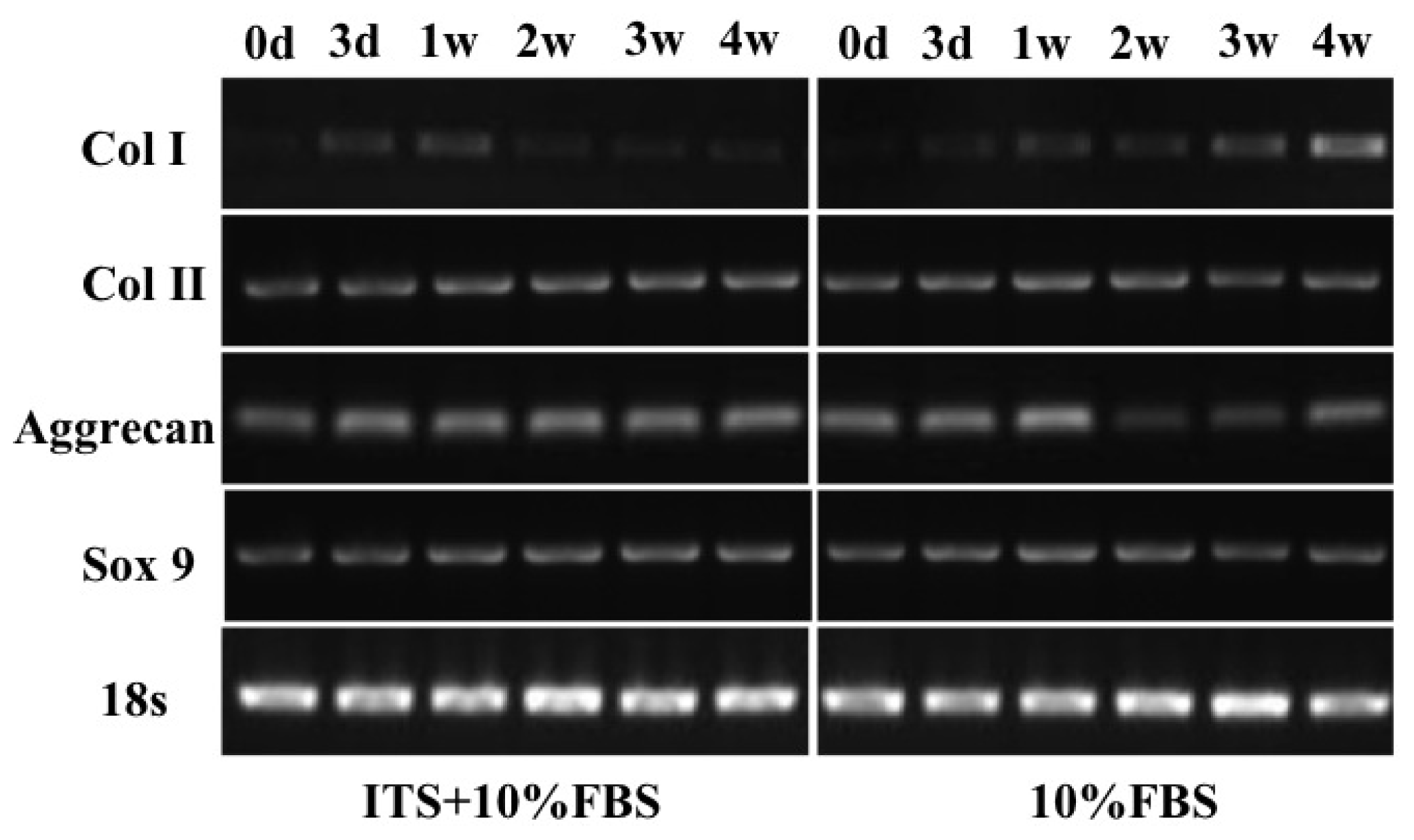
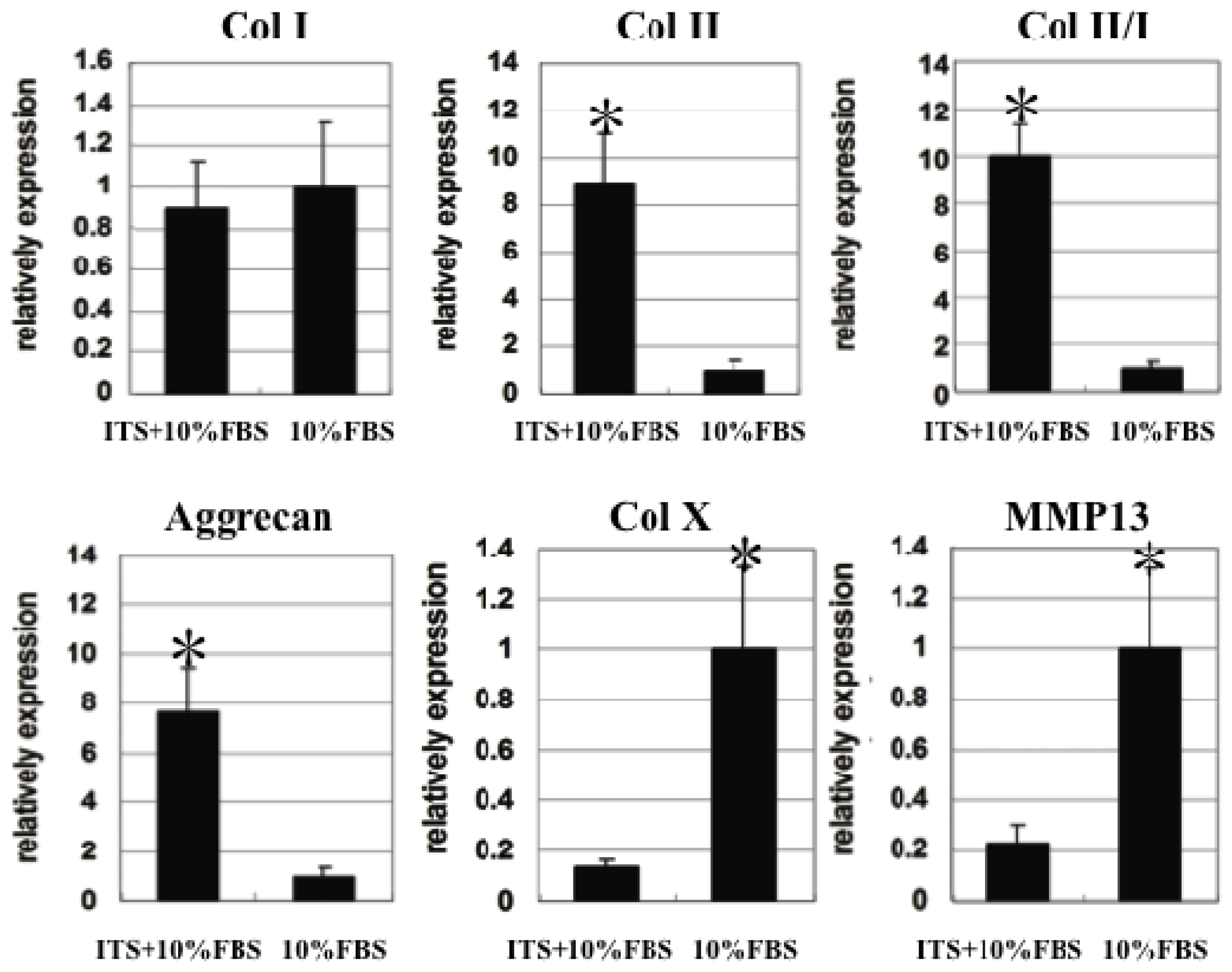
2.5. Quantitative Gene Expression of Chondrocyte Pellets
3. Experimental Section
3.1. Auricular Chondrocytes Isolation and Monolayer Culture
3.2. Chondrocyte Pellets Culture
3.3. Proliferation of Chondrocytes
3.4. Histology and Immunohistochemistry
3.5. GAG Quantification
3.6. RNA Extraction and Semi-Quantitative and Quantitative Real-Time Polymerase Chain Reaction (PCR)
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsStudy conception and design: Xia Liu, Jinchun Liu, Ran Xiao, Yilin Cao; acquisition of data: Jinchun Liu, Xia Liu, Ning Kang, Li Yan, Qian Wang, Xin Fu; analysis and interpretation of data: Xia Liu, Ran Xiao, Yilin Cao, Li Yan, Qian Wang, Xin Fu, Yuanyuan Zhang; drafting of manuscript: Xia Liu, Jinchun Liu; critical revision: Xia Liu, Ran Xiao, Yilin Cao, Yuanyuan Zhang.
References
- Nayyer, L.; Patel, K.H.; Esmaeili, A.; Rippel, R.A.; Birchall, M.; O’toole, G.; Butler, P.E.; Seifalian, A.M. Tissue engineering: Revolution and challenge in auricular cartilage reconstruction. Plast. Reconstr. Surg 2012, 129, 1123–1137. [Google Scholar]
- Isogai, N.; Kusuhara, H.; Ikada, Y.; Ohtani, H.; Jacquet, R.; Hillyer, J.; Lowder, E.; Landis, W.J. Comparison of different chondrocytes for use in tissue engineering of cartilage model structures. Tissue Eng 2006, 12, 691–703. [Google Scholar]
- Kisiday, J.D.; Kurz, B.; DiMicco, M.A.; Grodzinsky, A.J. Evaluation of medium supplemented with insulin-transferrin-selenium for culture of primary bovine calf chondrocytes in three-dimensional hydrogel scaffolds. Tissue Eng 2005, 11, 141–151. [Google Scholar]
- Loty, S.; Sautier, J.M.; Forest, N. Phenotypic modulation of nasal septal chondrocytes by cytoskeleton modification. Biorheology 2000, 37, 117–125. [Google Scholar]
- Van Osch, G.J.; Mandl, E.W.; Jahr, H.; Koevoet, W.; Nolst-Trenite, G.; Verhaar, J.A. Considerations on the use of ear chondrocytes as donor chondrocytes for cartilage tissue engineering. Biorheology 2004, 41, 411–421. [Google Scholar]
- Von der Mark, K.; Gauss, V.; von der Mark, H.; Müller, P. Relationship between cell shape and type of collagen synthesized as chondrocytes lose their cartilage phenotype in culture. Nature 1977, 267, 531–532. [Google Scholar]
- Elima, K.; Vuorio, E. Expression of mRNAs for collagens and other matrix components in dedifferentiating and redifferentiating human chondrocytes in culture. FEBS Lett 1989, 258, 195–198. [Google Scholar]
- Fujisawa, T.; Hattori, T.; Ono, M.; Uehara, J.; Kubota, S.; Kuboki, T.; Takigawa, M. CCN family 2/connective tissue growth factor (CCN2/CTGF) stimulates proliferation and differentiation of auricular chondrocytes. Osteoarthr. Cartil 2008, 16, 787–795. [Google Scholar]
- Chua, K.H.; Aminuddin, B.S.; Fuzina, N.H.; Ruszymah, B.H. Insulin-transferrin-selenium prevent human chondrocyte dedifferentiation and promote the formation of high quality tissue engineered human hyaline cartilage. Eur. Cell Mater 2005, 9, 58–67. [Google Scholar]
- Glaser, J.H.; Conrad, H.E. Properties of chick embryo chondrocytes grown in serum-free medium. J. Biol. Chem 1984, 259, 6766–6672. [Google Scholar]
- Webber, R.J.; Zitaglio, T.; Hough, A.J., Jr. Serum-free culture of rabbit meniscal fibrochondrocytes: Proliferative response. J. Orthop 1988, 6, 13–23. [Google Scholar]
- Kisiday, J.D.; Jin, M.; DiMicco, M.A.; Kurz, B.; Grodzinsky, A.J. Effects of dynamic compressive loading on chondrocyte biosynthesis in self-assembling peptide scaffolds. J. Biomech 2004, 37, 595–604. [Google Scholar]
- Li, W.J.; Danielson, K.G.; Alexander, P.G.; Tuan, R.S. Biological response of chondrocytes cultured in three-dimensional nanofibrous poly (epsilon-caprolactone) scaffolds. J. Biomed. Mater. Res 2003, 67, 1105–1114. [Google Scholar]
- Kelly, T.A.; Fisher, M.B.; Oswald, E.S.; Tai, T.; Mauck, R.L.; Ateshian, G.A.; Hung, C.T. Low-serum media and dynamic deformational loading in tissue engineering of articular cartilage. Ann. Biomed. Eng 2008, 36, 769–779. [Google Scholar]
- Yang, Y.H.; Barabino, G.A. Requirement for serum in medium supplemented with insulin-transferrin-selenium for hydrodynamic cultivation of engineered cartilage. Tissue Eng 2011, 17, 2025–2035. [Google Scholar]
- Zhang, L.; Song, H.; Zhao, X. Optimum combination of insulin-transferrin-selenium and fetal bovine serum for culture of rabbit articular chondrocytes in three-dimensional alginate scaffolds. Int. J. Cell Biol 2009, 2009, 747016. [Google Scholar]
- Bernstein, P.; Dong, M.; Corbeil, D.; Gelinsky, M.; Günther, K.P.; Fickert, S. Pellet culture elicits superior chondrogenic redifferentiation than alginate-based systems. Biotechnol Prog 2009, 25, 1146–1152. [Google Scholar]
- Wu, Y.; Zhu, L.; Jiang, H.; Liu, W.; Liu, Y.; Cao, Y.; Zhou, G. Engineering cartilage substitute with a specific size and shape using porous high-density polyethylene (HDPE) as internal support. J. Plast. Reconstr. Aesthet. Surg 2010, 63, e370–e375. [Google Scholar]
- Gardiner, B.; Smith, D.; Pivonka, P.; Grodzinsky, A.; Frank, E.; Zhang, L. Solute transport in cartilage undergoing cyclic deformation. Comput. Methods Biomech. Biomed. Eng 2007, 10, 265–278. [Google Scholar]
- Zhang, Y.; Yang, F.; Liu, K.; Shen, H.; Zhu, Y.; Zhang, W.; Liu, W.; Wang, S.; Cao, Y.; Zhou, G. The impact of PLGA scaffold orientation on in vitro cartilage regeneration. Biomaterials 2012, 33, 2926–2935. [Google Scholar]
- Bian, L.; Lima, E.G.; Angione, S.L.; Ng, K.W.; Williams, D.Y.; Xu, D.; Stoker, A.M.; Cook, J.L.; Ateshian, G.A.; Hung, C.T. Mechanical and biochemical characterization of cartilage explants in serum-free culture. J. Biomech 2008, 41, 1153–1159. [Google Scholar]
- Fitzsimmons, J.S.; Sanyal, A.; Gonzalez, C.; Fukumoto, T.; Clemens, V.R.; O’Driscoll, S.W.; Reinholz, G.G. Serum-free media for periosteal chondrogenesis in vitro. J. Orthop. Res. 2004, 22, 716–725. [Google Scholar]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp. Cell Res 1998, 238, 265–272. [Google Scholar]
- Cheung, W.H.; Lee, K.M.; Fung, K.P.; Leung, K.S. Growth plate chondrocytes inhibit neo-angiogenesis—A possible mechanism for tumor control. Cancer Lett 2001, 163, 25–32. [Google Scholar]
- Farndale, R.W.; Buttle, D.J.; Barrett, A.J. Improved quantitation and discrimination of sulphated glycosaminoglycans by use of dimethylmethylene blue. Biochim. Biophys. Acta 1986, 883, 173–177. [Google Scholar]
- Spiller, K.L.; Liu, Y.; Holloway, J.L.; Maher, S.A.; Cao, Y.; Liu, W.; Zhou, G.; Lowman, A.M. A novel method for the direct fabrication of growth factor-loaded microspheres within porous nondegradable hydrogels: Controlled release for cartilage tissue engineering. J. Control. Release 2012, 157, 39–45. [Google Scholar]
- Ibold, Y.; Lübke, C.; Pelz, S.; Augst, H.; Kaps, C.; Ringe, J.; Sittinger, M. Effect of different ascorbate supplementations on in vitro cartilage formation in porcine high-density pellet cultures. Tissue Cell 2009, 41, 249–256. [Google Scholar]
- Jefferies, D.; Farquharson, C.; Thomson, J.; Smith, W.; Seawright, E.; McCormack, H.; Whitehead, C. Differences in metabolic parameters and gene expression related to osteochondrosis/osteoarthrosis in pigs fed 25-hydroxyvitamin D3. Vet. Res 2002, 33, 383–396. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Abbreviation of Groups | Culture Media |
|---|---|---|
| 1 | ITS + 2% FBS | DMEM + 1% ITS + 2% FBS |
| 2 | ITS + 6% FBS | DMEM + 1% ITS + 6% FBS |
| 3 | ITS + 10% FBS | DMEM + 1% ITS + 10% FBS |
| 4 | 10% FBS | DMEM +10% FBS |
| 5 | ITS | DMEM + 1% ITS |
| Gene | Primer 5′–3′ | Size (bp) |
|---|---|---|
| Col I [27] | F: CGATGGCTGCACGAGTCACAC R: CAGGTTGGGATGGAGGGAGTTTAC | 180 |
| Col II [27] | F: CCGGGCAGAGGGCAATAGCAGGTT R: CAATGATGGGGAGGCGTGAG | 128 |
| aggrecan [27] | F: CCAGAATCTAGCAGGGAGTCATC R: AGGCAGAGGTGGCTTCAGTC | 117 |
| Sox 9 | F: CACAGCTCACCAGACCTTGA R: GTGGGTTCGAGTTGCCTTTA | 176 |
| Col X [28] | F: GCTGCCACATTCTGACACAATC R: TGCGCTGAGCATCATTTGAGAC | 225 |
| MMP13 | F: CTTGTTTCTTGTTGCTGCCC R: GTTGGGGTCTTCATCTCCTG | 178 |
| GAPDH [27] | F: CTGCCCCTTCTGCTGATGC R: TCCACGATGCCGAAGTTGTC | 151 |
| 18S | F: TGAGAAACGGCTACCACATC R: TCCCAAGATCCAACTACGAG | 250 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.; Liu, J.; Kang, N.; Yan, L.; Wang, Q.; Fu, X.; Zhang, Y.; Xiao, R.; Cao, Y. Role of Insulin-Transferrin-Selenium in Auricular Chondrocyte Proliferation and Engineered Cartilage Formation in Vitro. Int. J. Mol. Sci. 2014, 15, 1525-1537. https://doi.org/10.3390/ijms15011525
Liu X, Liu J, Kang N, Yan L, Wang Q, Fu X, Zhang Y, Xiao R, Cao Y. Role of Insulin-Transferrin-Selenium in Auricular Chondrocyte Proliferation and Engineered Cartilage Formation in Vitro. International Journal of Molecular Sciences. 2014; 15(1):1525-1537. https://doi.org/10.3390/ijms15011525
Chicago/Turabian StyleLiu, Xia, Jinchun Liu, Ning Kang, Li Yan, Qian Wang, Xin Fu, Yuanyuan Zhang, Ran Xiao, and Yilin Cao. 2014. "Role of Insulin-Transferrin-Selenium in Auricular Chondrocyte Proliferation and Engineered Cartilage Formation in Vitro" International Journal of Molecular Sciences 15, no. 1: 1525-1537. https://doi.org/10.3390/ijms15011525