Artesunate Reduces Serum Lipopolysaccharide in Cecal Ligation/Puncture Mice via Enhanced LPS Internalization by Macrophages through Increased mRNA Expression of Scavenger Receptors

Abstract
:1. Introduction
2. Results and Discussion
2.1. AS Protection on CLP Mice Is Related to Decreasing Serum Cytokines and LPS Levels
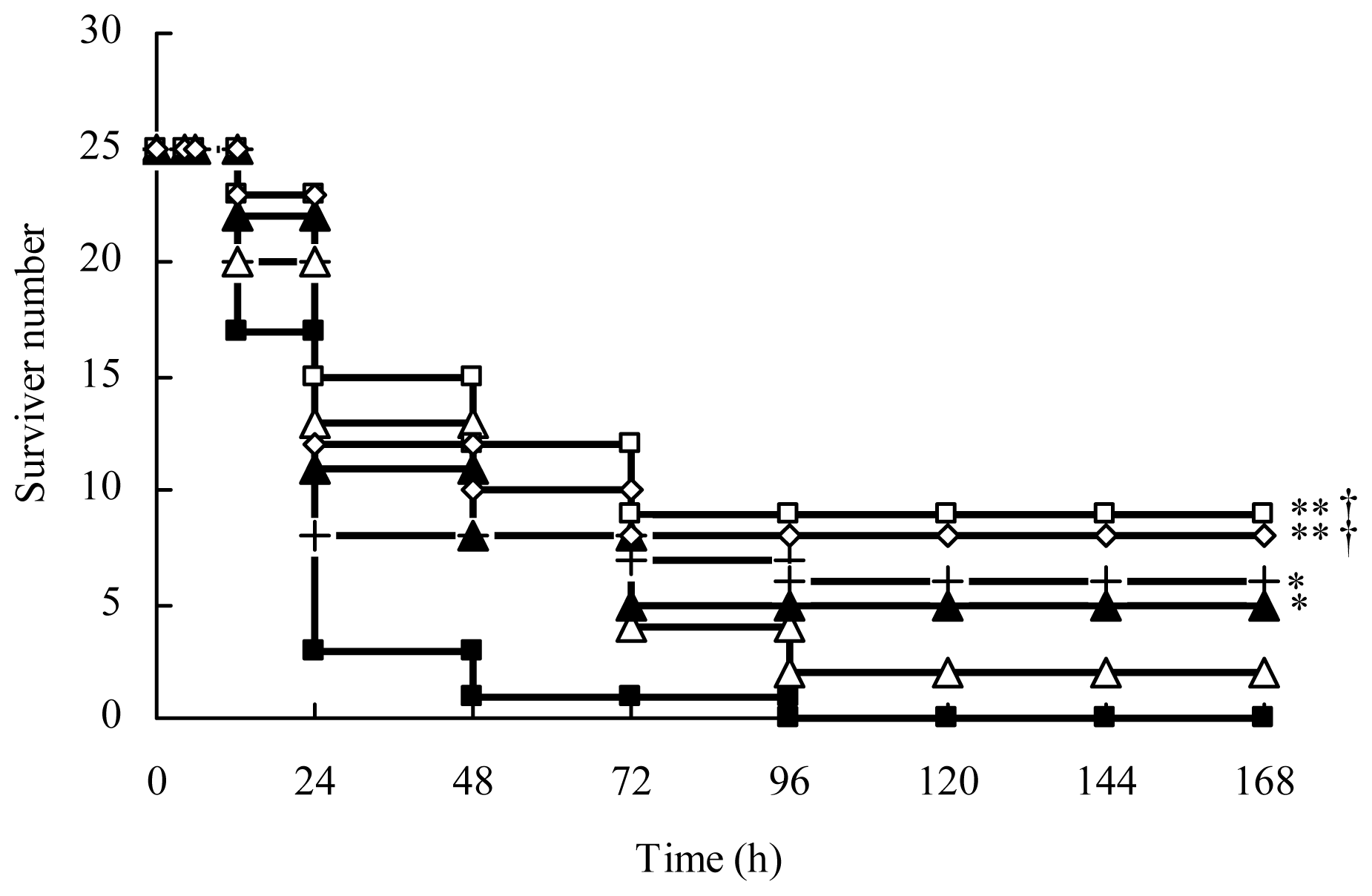
2.1.1. AS Improves the Survival of CLP Mice
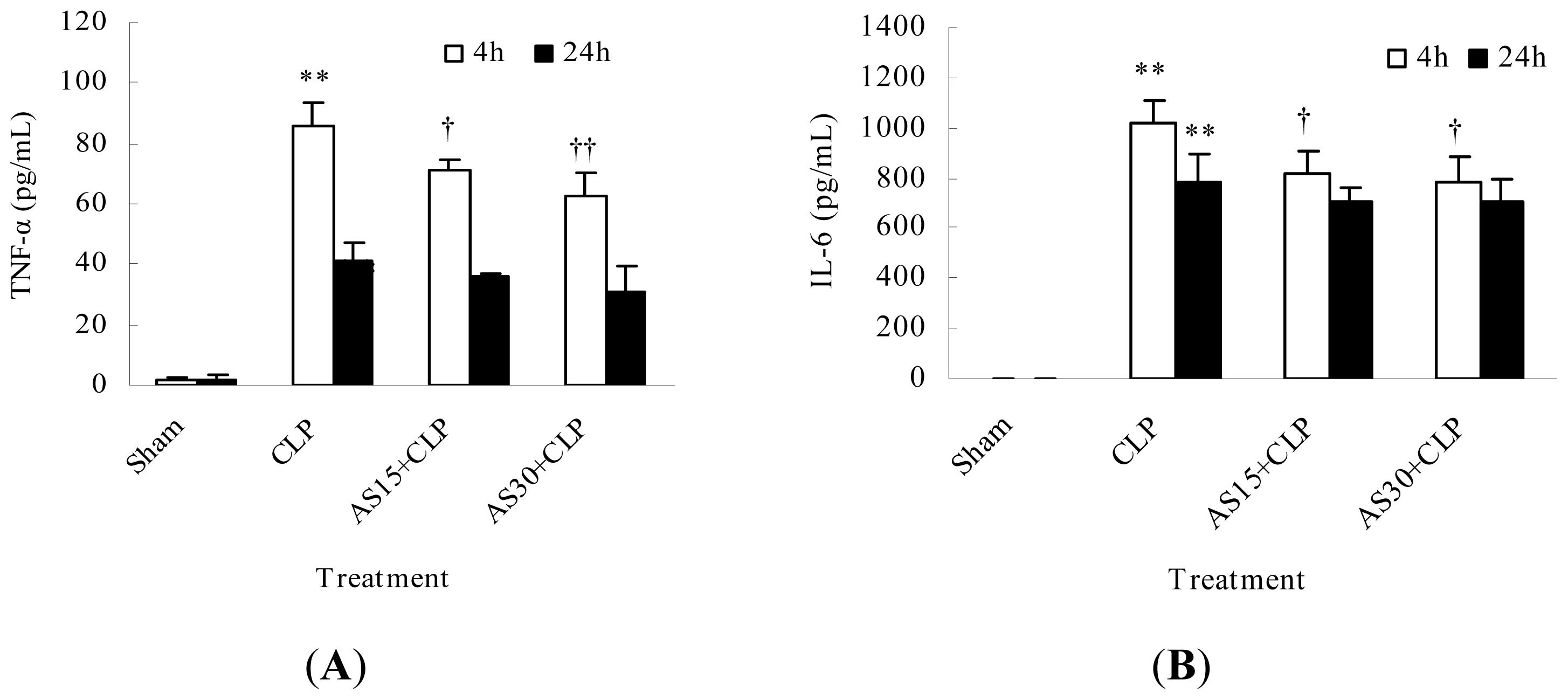
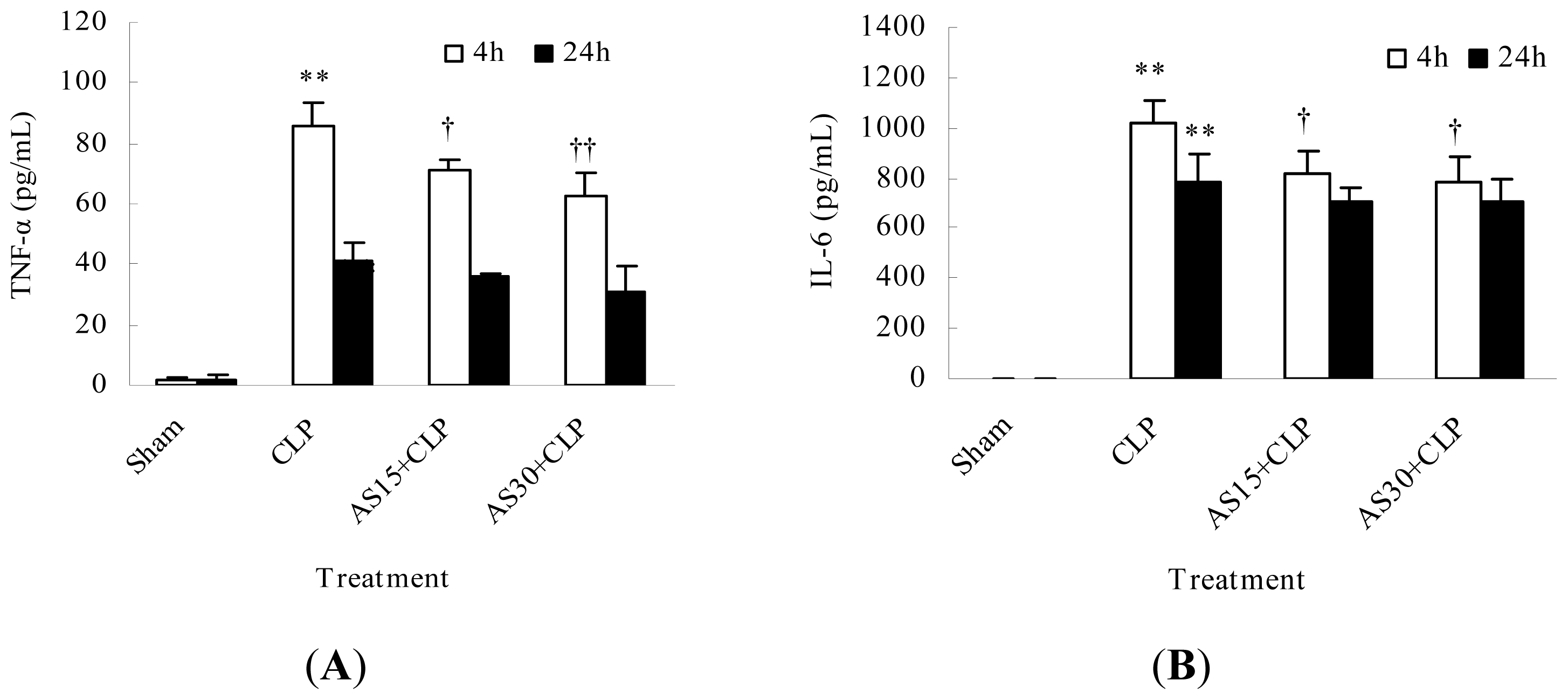
2.1.2. AS Decreases the Serum Pro-Inflammatory Cytokine Levels of CLP Mice
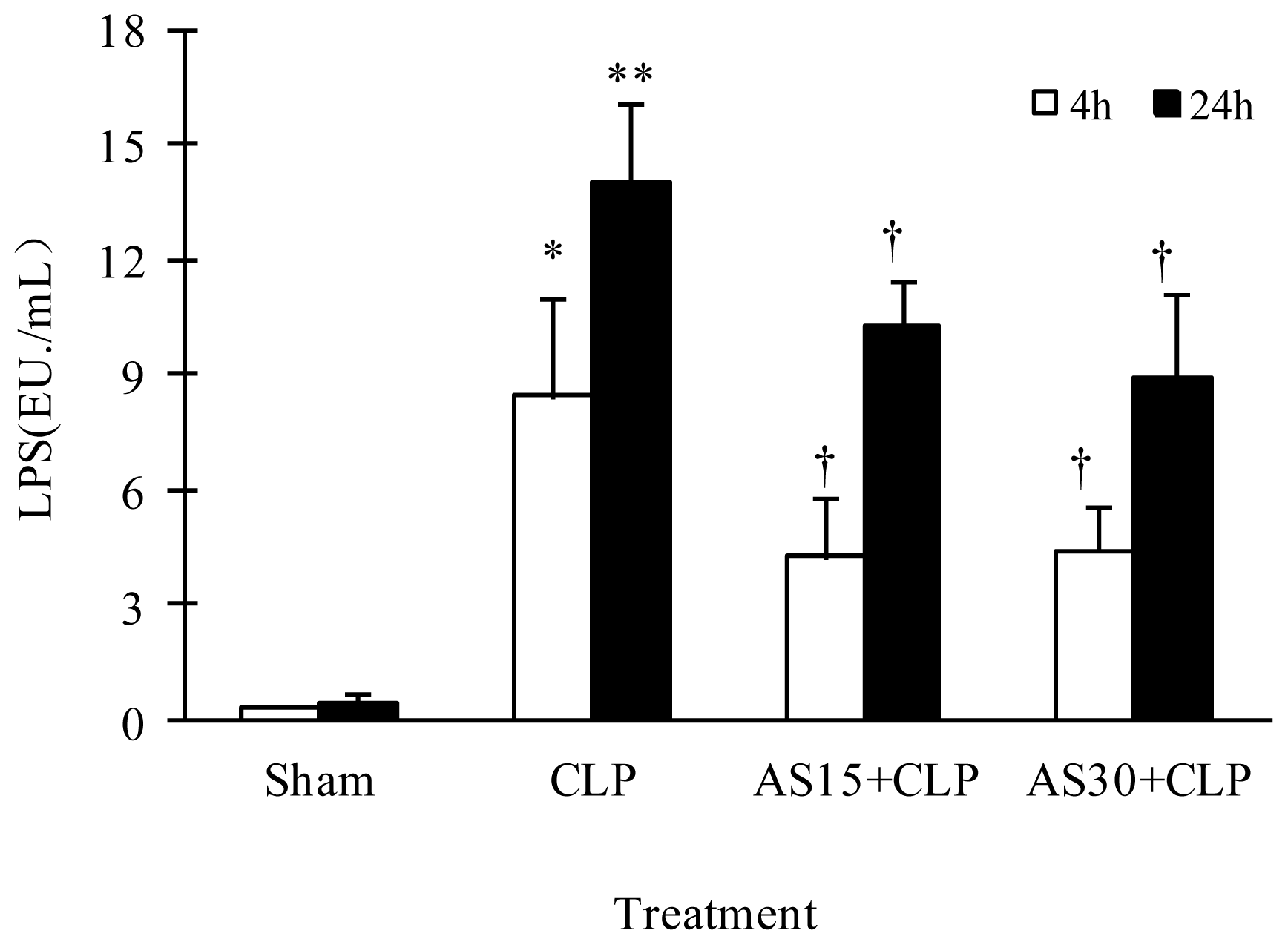
2.1.3. AS Reduces the Serum LPS Level of CLP Mice
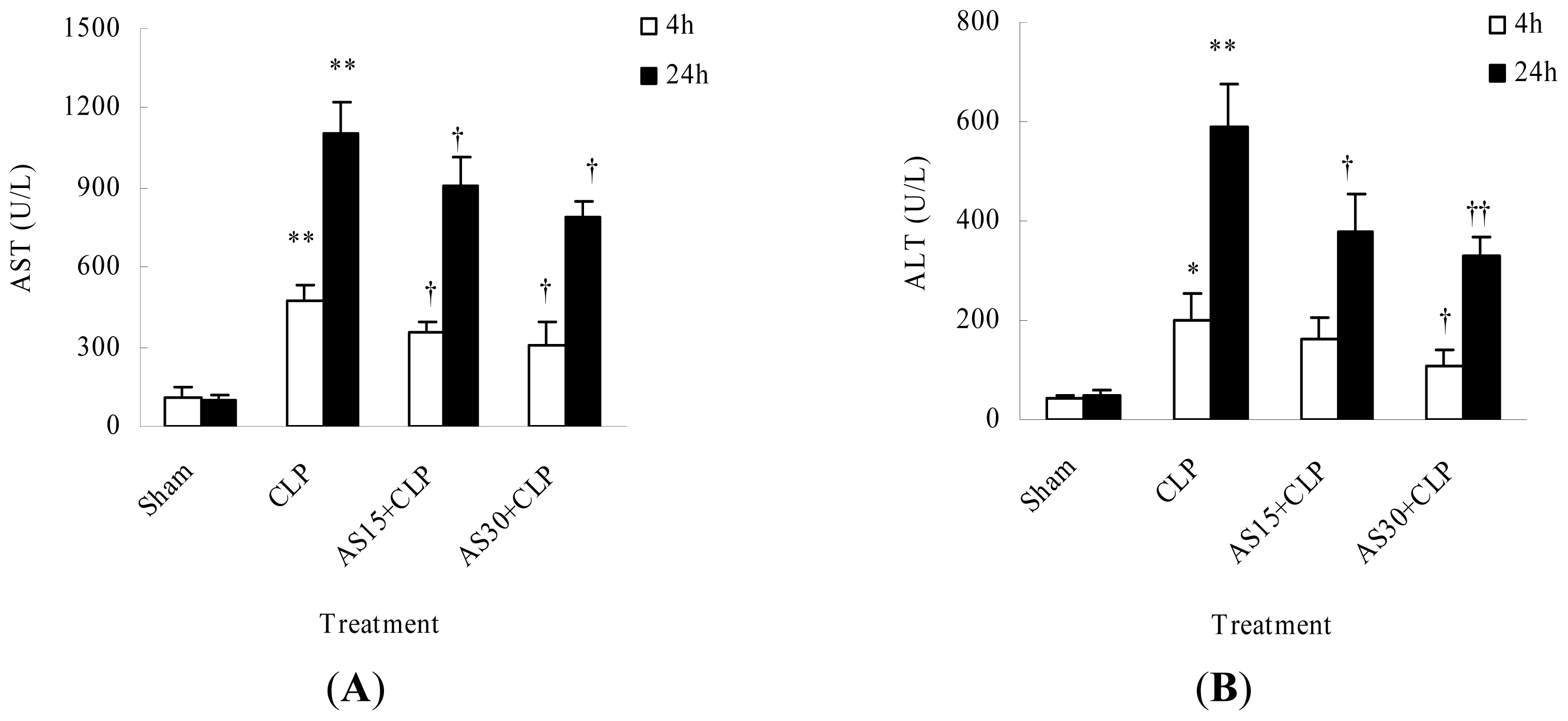
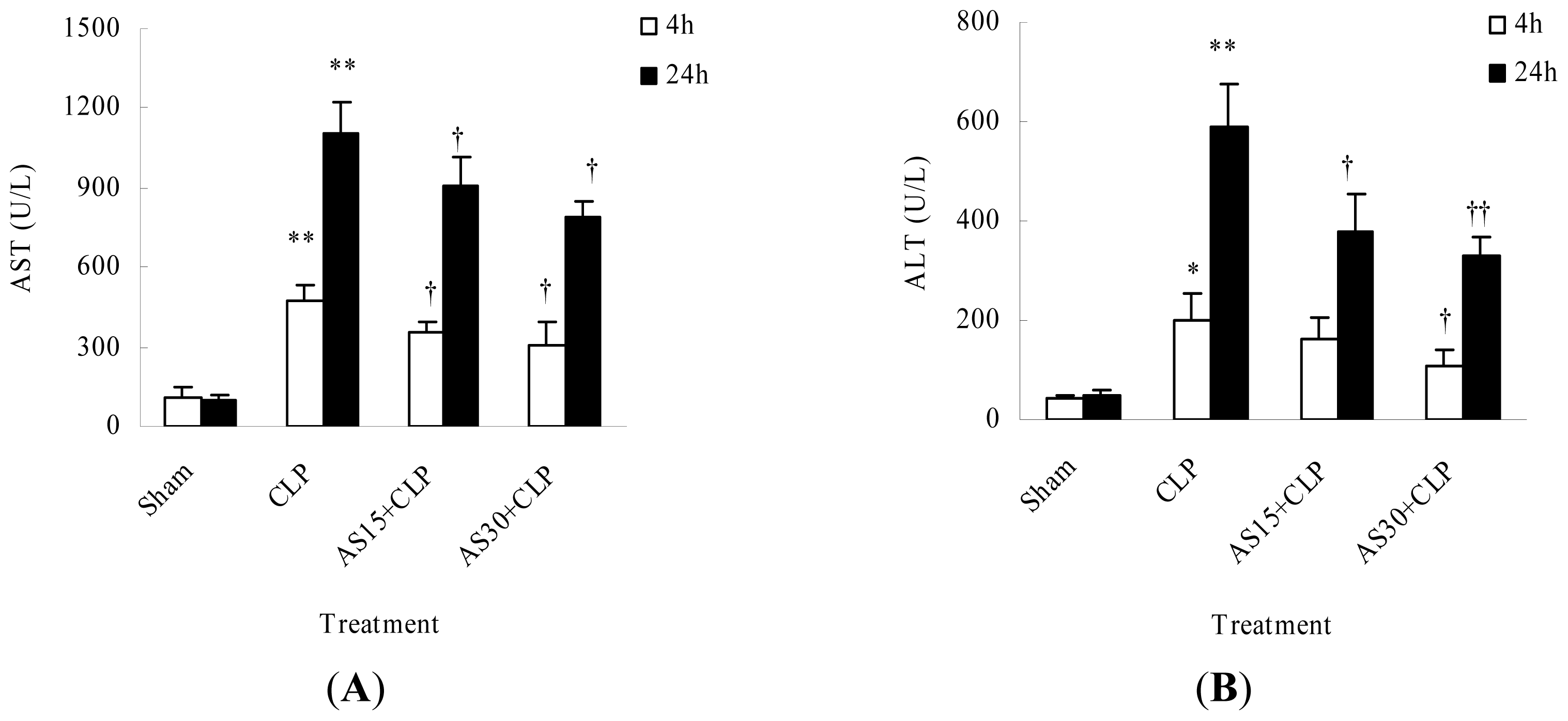
2.1.4. AS Protects Liver Function of CLP Mice
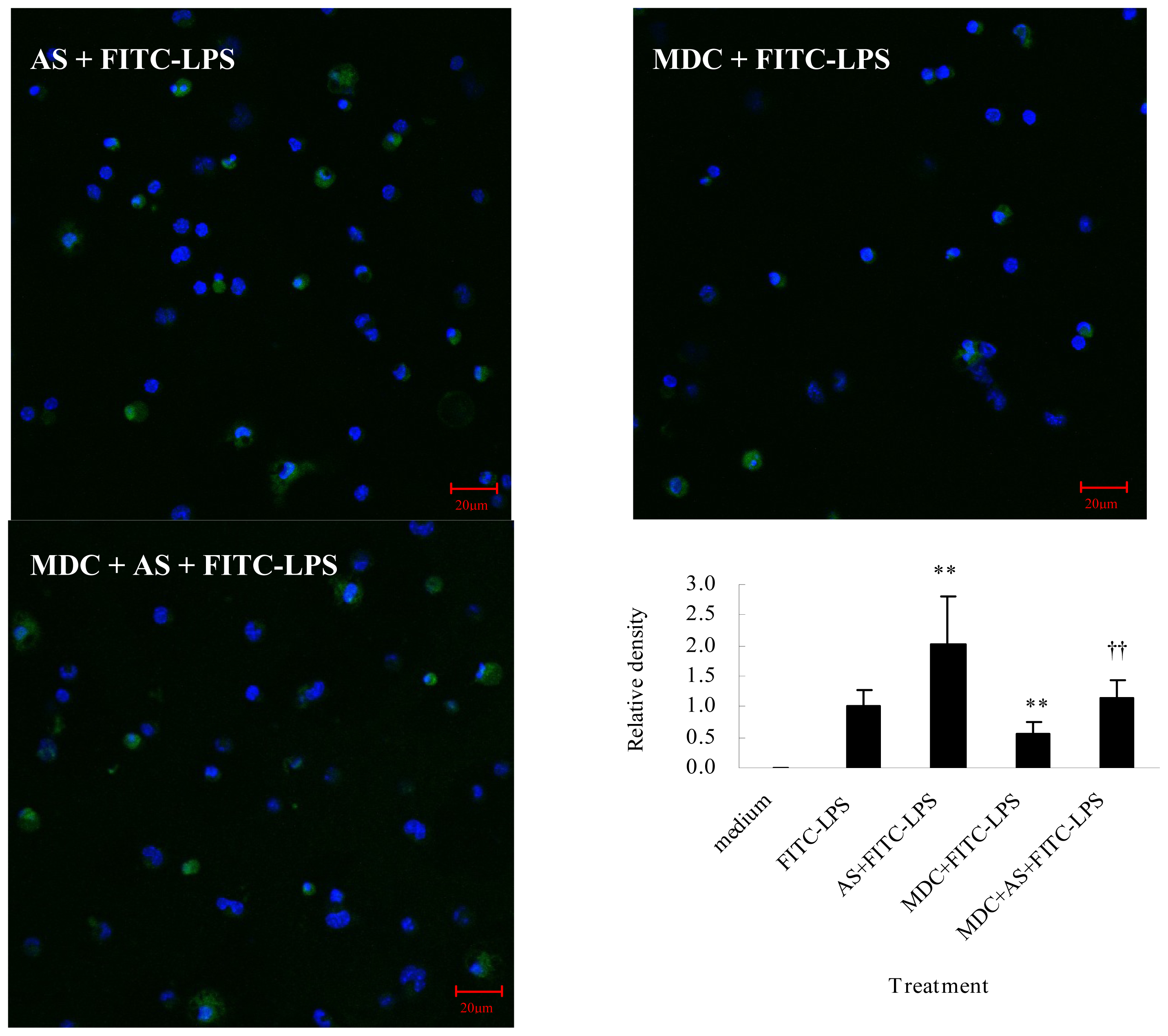
2.2. AS Enhancement on LPS Internalization Is not Related to the Clathrin-Dependent Pathway
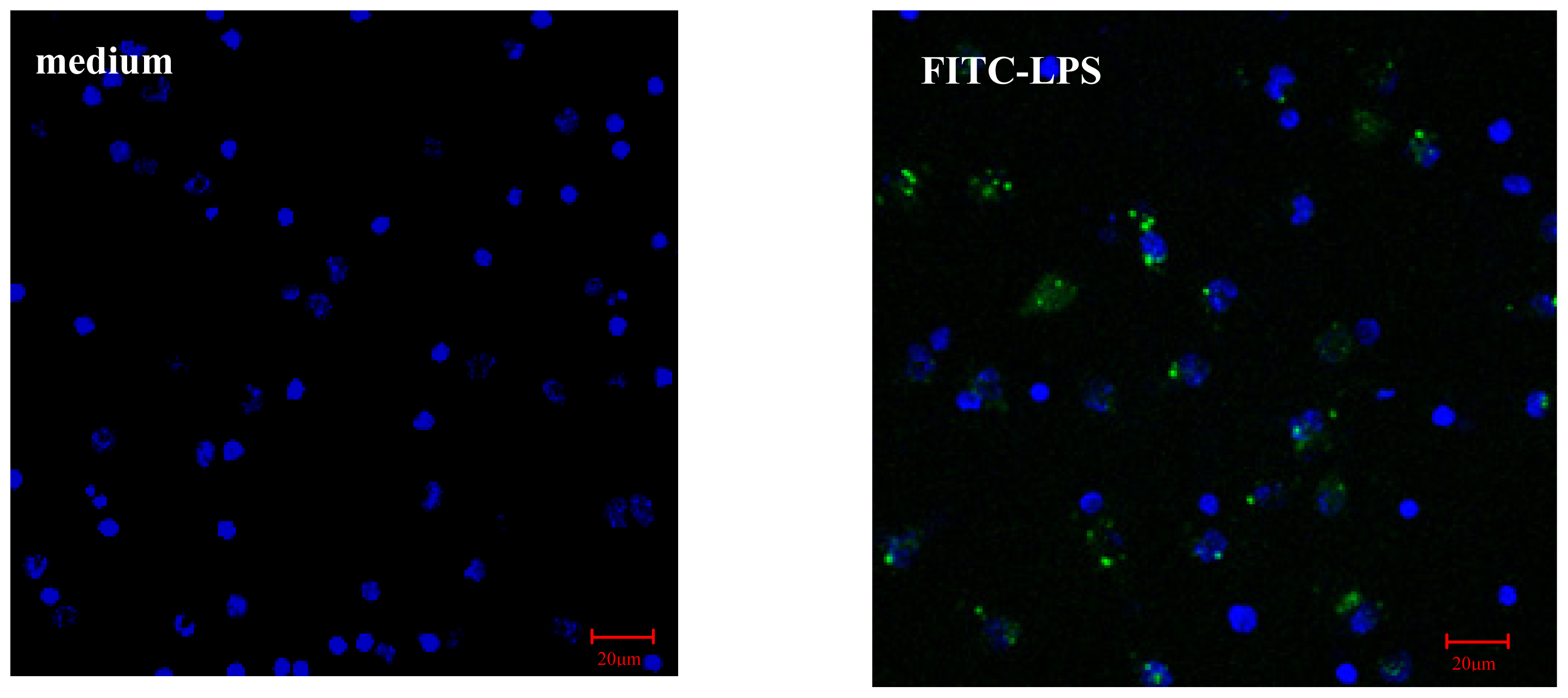
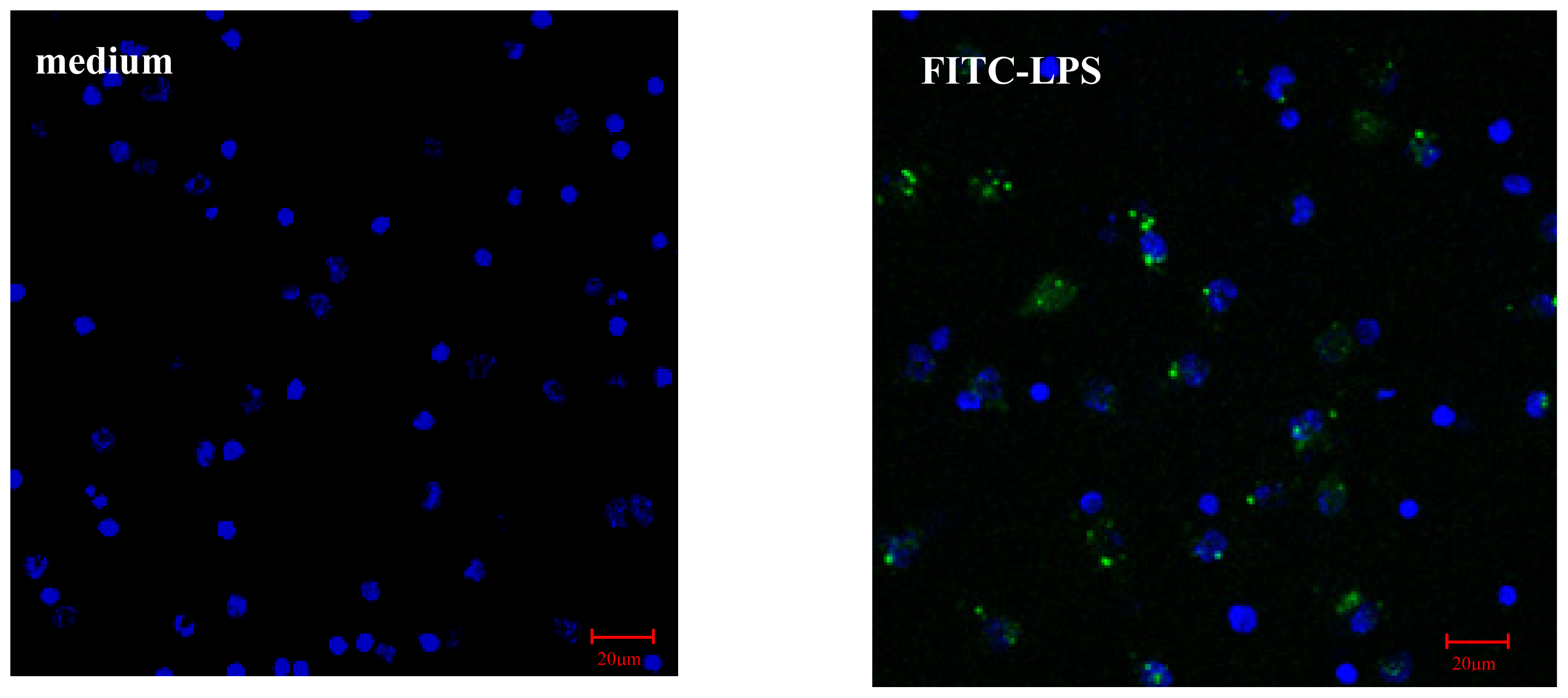
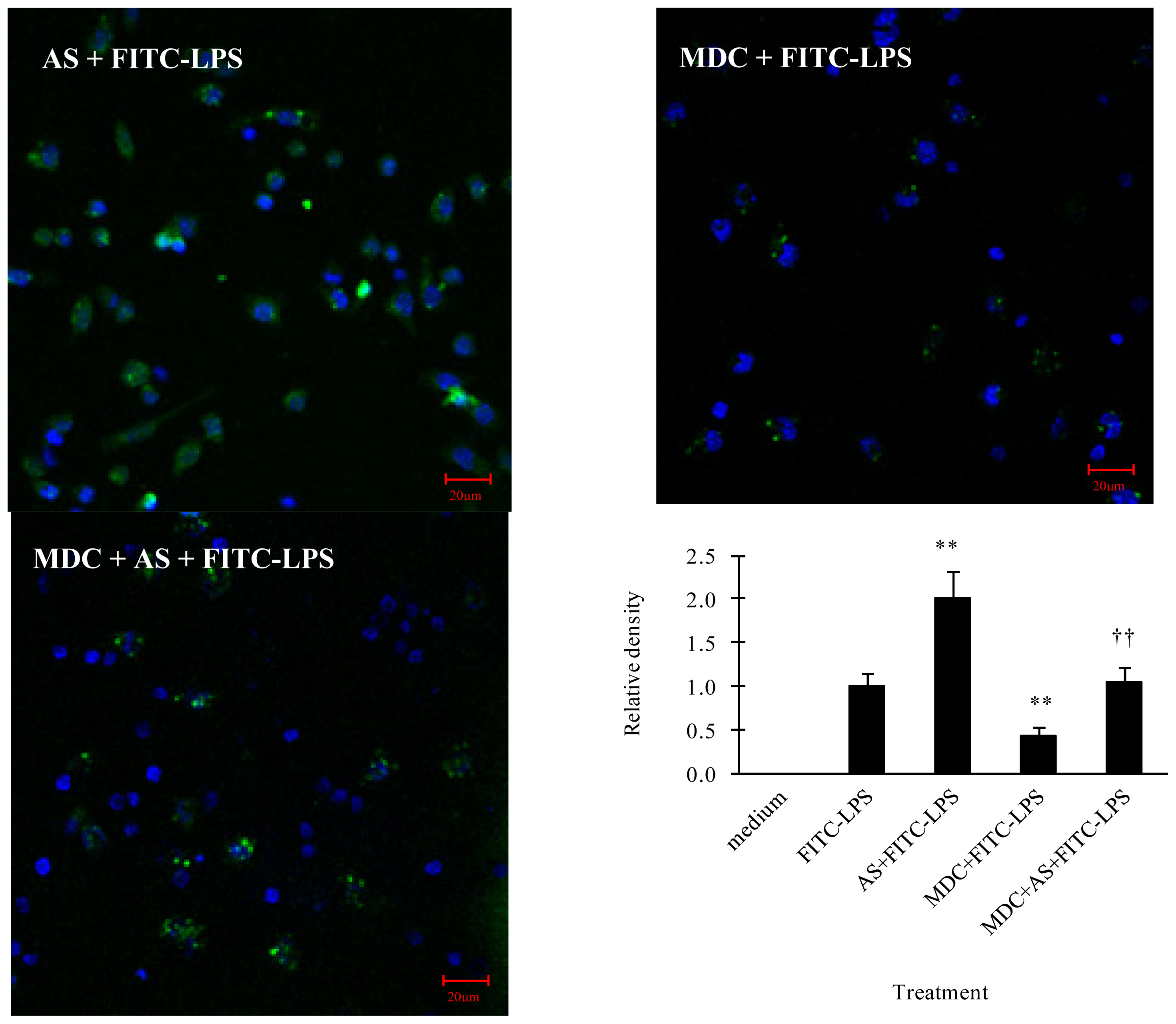
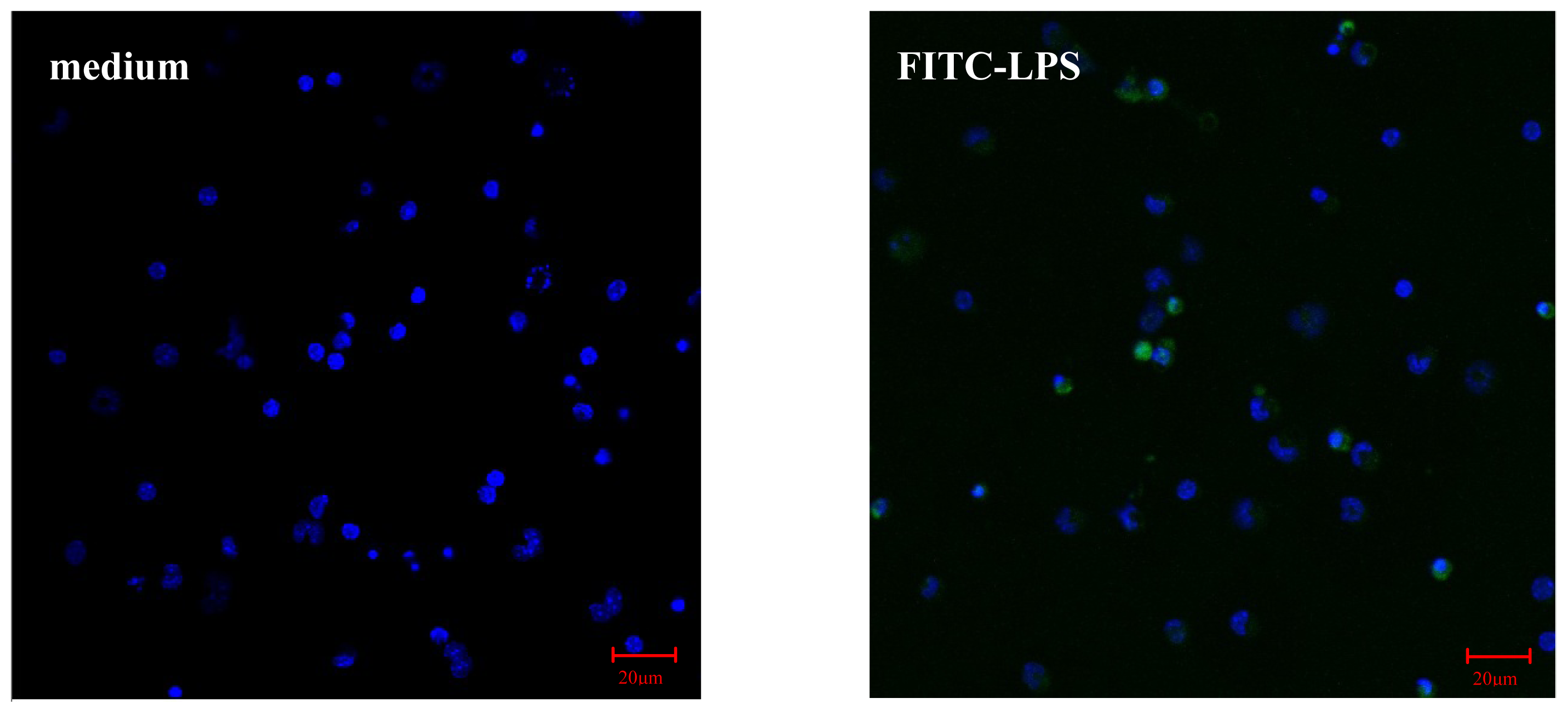
2.2.1. AS Increases FITC-LPS Internalization by Peritoneal Macrophages and Liver Kupffer Cells
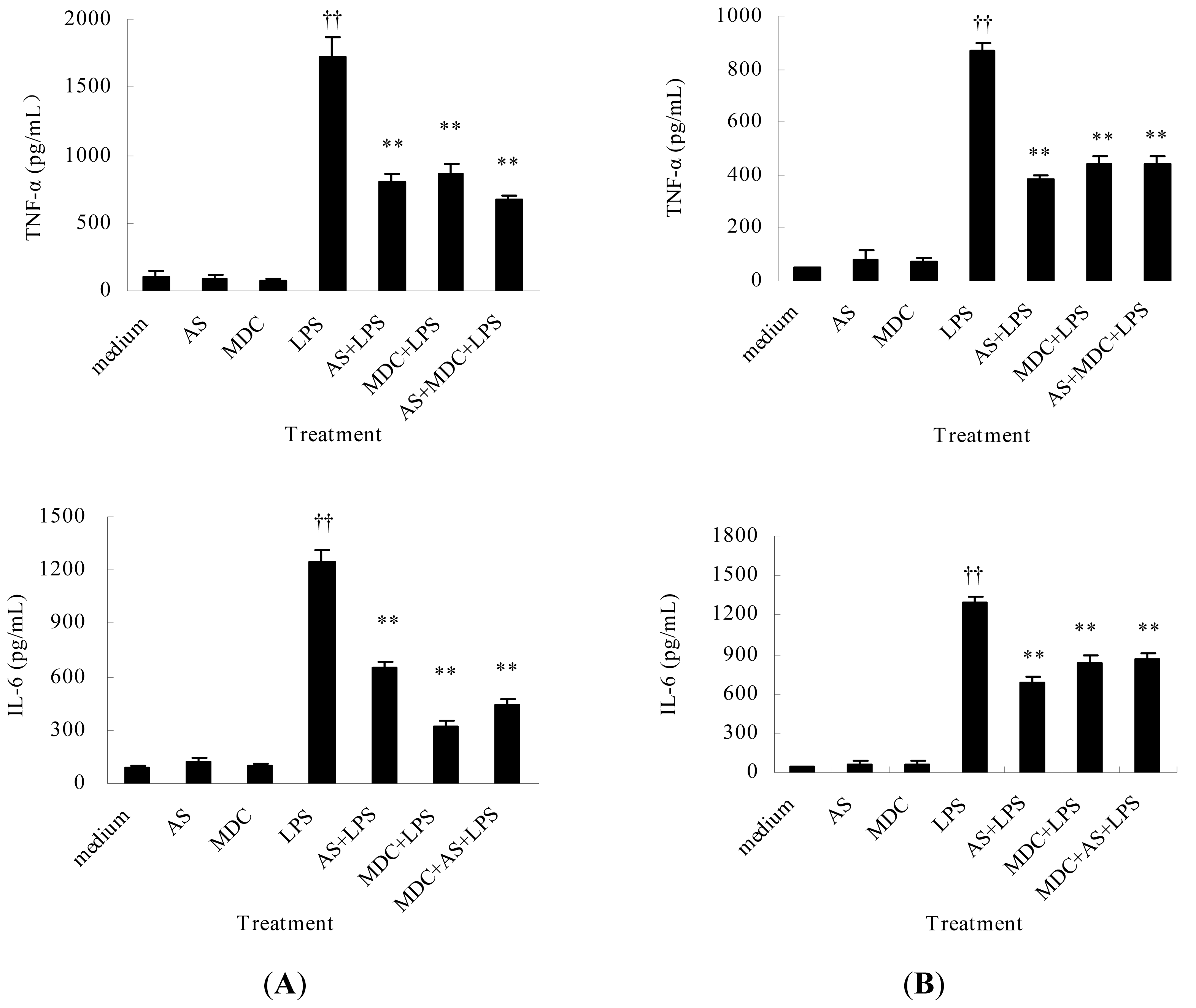
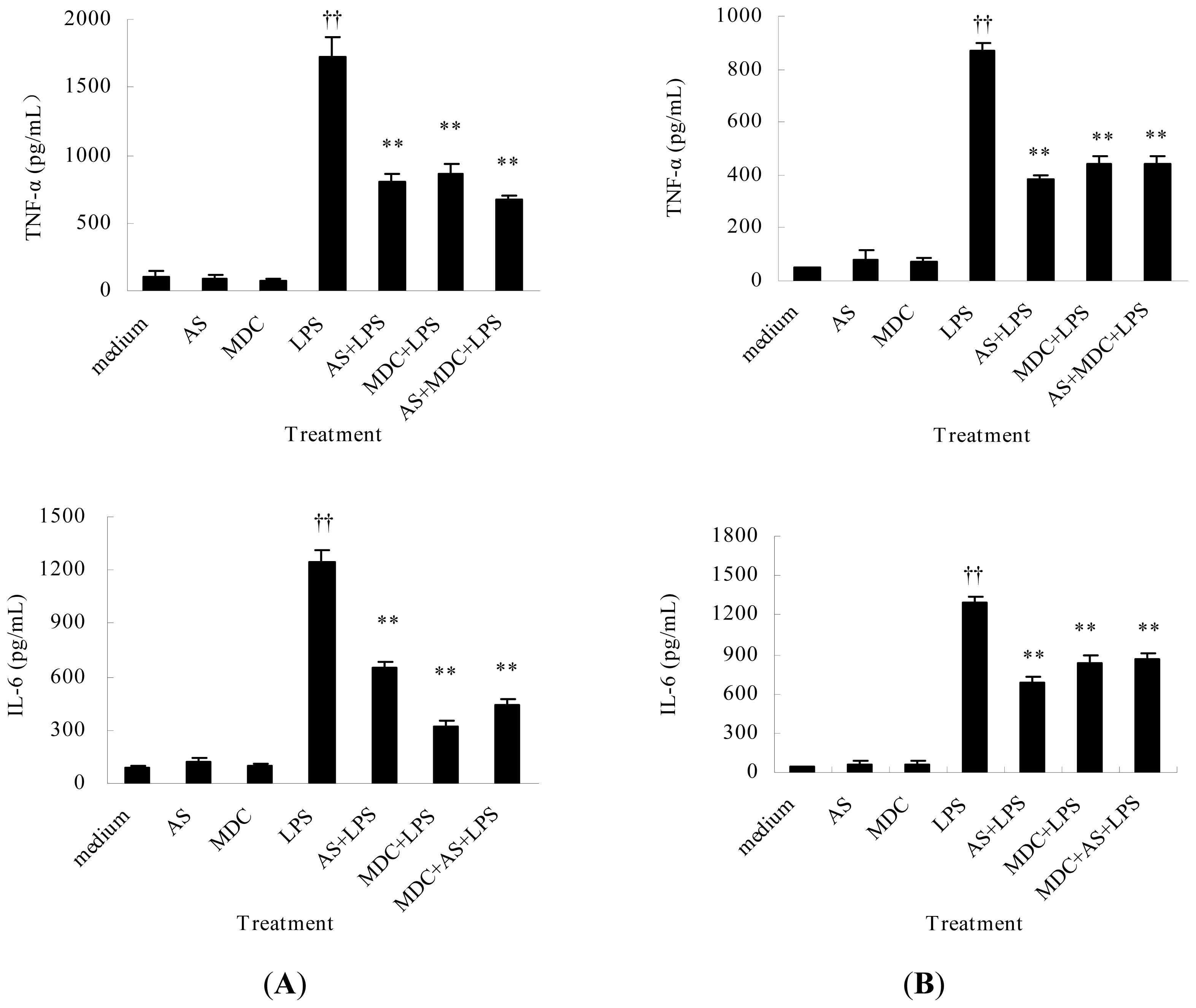
2.2.2. AS Inhibits TNF-α and IL-6 Release from Peritoneal Macrophages and Liver Kupffer Cells Induced by LPS
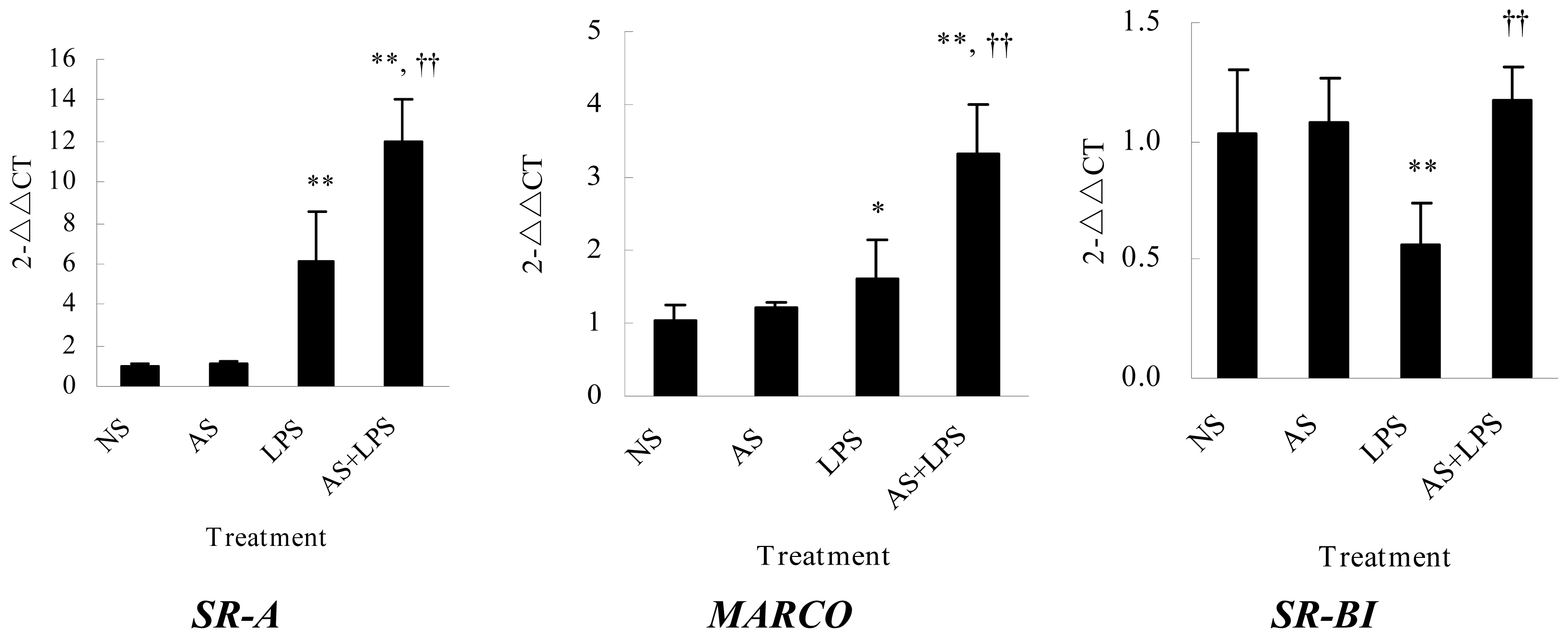
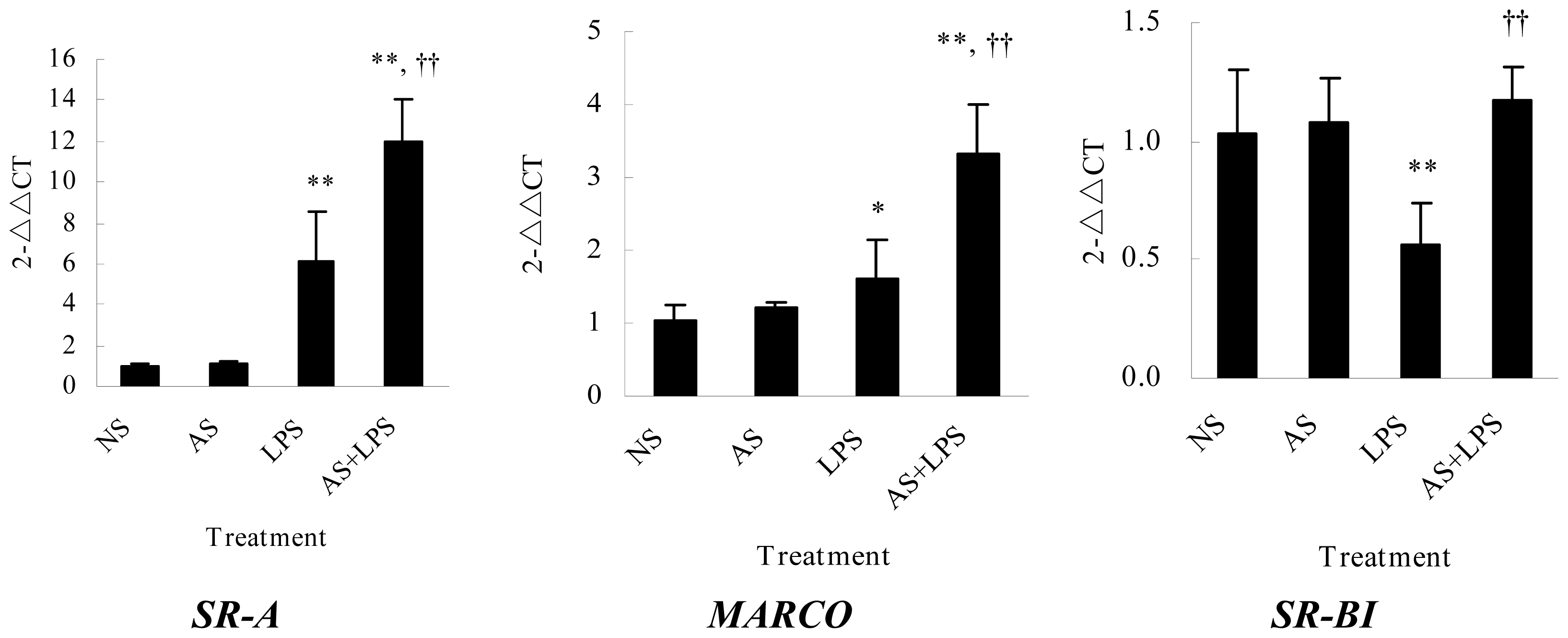
2.3. AS Increases Scavenger Receptors mRNA Expression in Peritoneal Macrophages
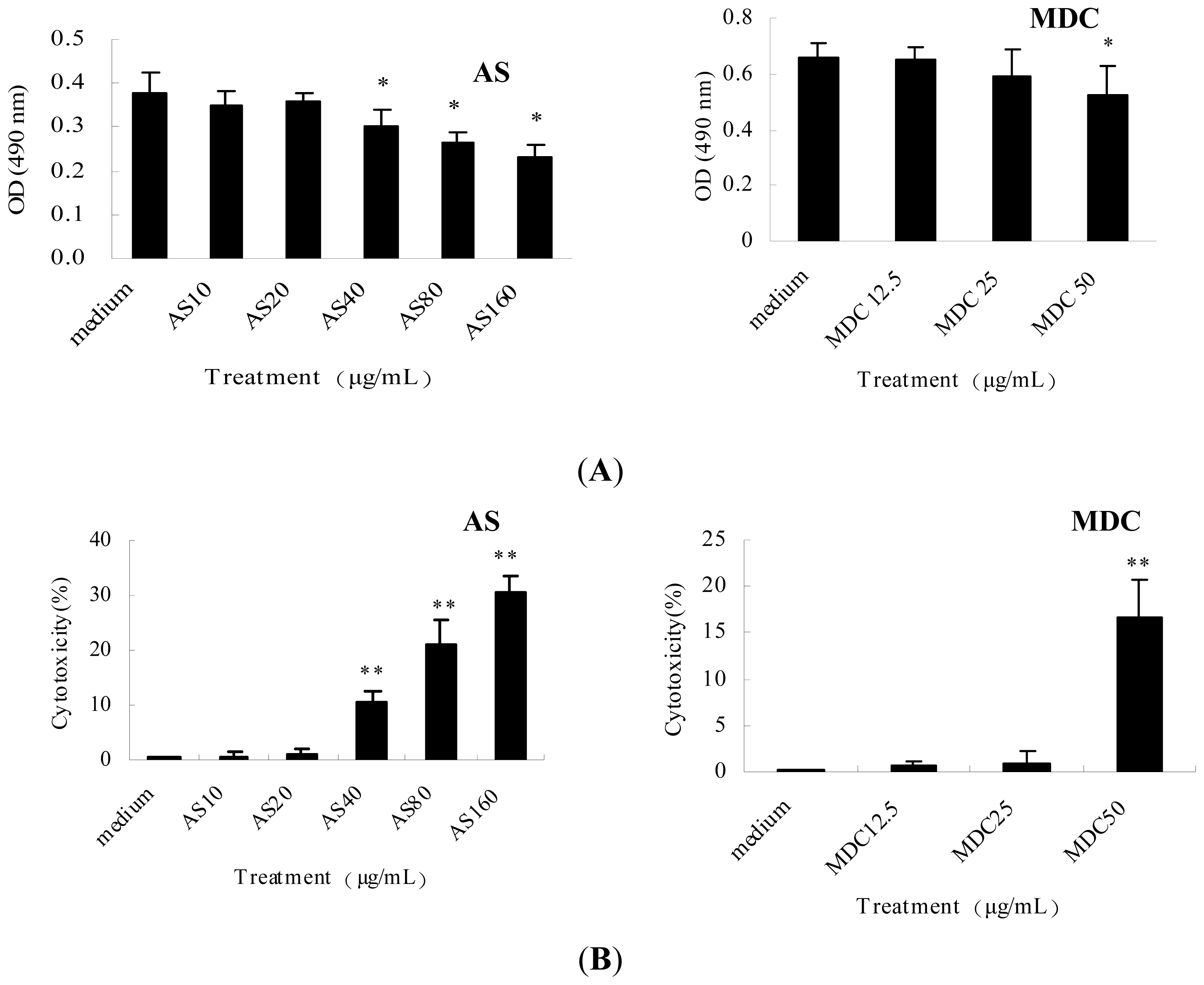
2.4. AS Used in Indicated Concentrations Exhibits no Cellular Toxicity in Vitro
2.5. Discussion
3. Experimental Section
3.1. Materials
3.2. Experimental Sepsis Induced by CLP and Survival Rate
3.3. Experimental Sepsis Induced by CLP and Serum Collection
3.4. Isolation of Peritoneal Macrophages and Liver Kupffer Cells from Mice and Cell Culture
3.5. Immunofluorescence Imaging of LPS Internalization
3.6. Cytokine Quantification
3.7. Quantitative PCR Analysis
3.8. Cytotoxicity Assays
3.9. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-15-01143-s001.pdfAcknowledgments
Conflicts of Interest
References
- McPherson, D.; Griffiths, C.; Williams, M.; Baker, A.; Klodawski, E.; Jacobson, B.; Donaldson, L. Sepsis-associated mortality in England: An analysis of multiple cause of death data from 2001 to 2010. BMJ Open 2013, 3, e002586. [Google Scholar]
- Wang, Y.; Mao, M.; Xu, J.C. Cell-surface nucleolin is involved in lipopolysaccharide internalization and signalling in alveolar macrophages. Cell. Biol. Int 2011, 35, 677–685. [Google Scholar]
- Lentschat, A.; El-Samalouti, V.T.; Schletter, J.; Kusumoto, S.; Brade, L.; Rietschel, E.T.; Gerdes, J.; Ernst, M.; Flad, H.; Ulmer, A.J. The internalization time course of a given lipopolysaccharide chemotype does not correspond to its, activation kinetics in monocytes. Infect. Immun 1999, 67, 2515–2521. [Google Scholar]
- Huang, X.; Venet, F.; Wang, Y.L.; Lepape, A.; Yuan, Z.; Chen, Y.; Swan, R.; Kherouf, H.; Monneret, G.; Chung, C.S.; et al. PD-1 expression by macrophages plays a pathologic role in altering microbial clearance and the innate inflammatory response to sepsis. Proc. Natl. Acad. Sci. USA 2009, 106, 6303–6308. [Google Scholar]
- Li, B.; Zhang, R.; Li, J.; Zhang, L.Z.; Ding, G.F.; Luo, P.; He, S.Q.; Dong, Y.; Jiang, W.W.; Lu, Y.L.; et al. Antimalarial artesunate protects sepsis model mice against heat-killed Escherichia coli challenge by decreasing TLR4, TLR9 mRNA expressions and transcription factor NF-κB activation. Int. Immunopharmacol 2008, 8, 379–389. [Google Scholar]
- Kakita, H.; Hussein, M.H.; Daoud, G.A.; Kato, T.; Murai, H.; Sugiura, T.; Mizuno, K.; Yamada, Y.; Ito, T.; Fukuda, S.; et al. Total hydroperoxide and biological antioxidant potentials in a neonatal sepsis model. Pediatr. Res 2006, 60, 675–679. [Google Scholar]
- Dellinger, R.P.; Levy, M.M.; Rhodes, A.; Annane, D.; Gerlach, H.; Opal, S.M.; Sevransky, J.E.; Sprung, C.L.; Douglas, I.S.; Jaeschke, R.; et al. Surviving Sepsis Campaign. International guidelines for management of severe sepsis and septic shock, 2012. Intensive Care Med 2012, 39, 165–228. [Google Scholar]
- Scott, M.J.; Liu, S.; Shapiro, R.A.; Vodovotz, Y.; Billiar, T.R. Endotoxin uptake in mouse liver is blocked by endotoxin pretreatment through a suppressor of cytokine signaling-1-dependent mechanism. Hepatology 2009, 49, 1695–1708. [Google Scholar]
- Li, B.; Yao, Q.; Pan, X.C.; Wang, N.; Zhang, R.; Li, J.; Ding, G.F.; Liu, X.; Wu, C.; Ran, D.Z.; et al. Artesunate enhances the antibacterial effect of β-lactam antibiotics against Escherichia coli by increasing antibiotic accumulation via inhibition of the multidrug efflux pump system AcrAB-TolC. J. Antimicrob. Chemother 2011, 66, 769–777. [Google Scholar]
- Kagan, J.C.; Su, T.; Horng, T.; Chow, A.; Akira, S.; Medzhitov, R. TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-β. Nat. Immunol 2008, 9, 361–368. [Google Scholar]
- Zhou, H.; Zheng, J.; Wang, L.X.; Ding, G.F.; Luo, P.; Lu, Y.L.; Pan, W.D.; Wang, M.H. Chloroquine protects mice from challenge with CpG ODN and LPS by decreasing proinflammatory cytokine release. Int. Immunopharmacol 2004, 4, 223–234. [Google Scholar]
- Weber, S.M.; Levitz, S.M. Chloroquine antagonizes the proinflammatory cytokine response to opportunistic fungi by alkalizing the fungal phagolysosome. J. Infect. Dis 2001, 183, 935–942. [Google Scholar]
- Ohnishi, K.; Komohara, Y.; Fujiwara, Y.; Takemura, K.; Lei, X.; Nakagawa, T.; Sakashita, N.; Takeya, M. Suppression of TLR4-mediated inflammatory response by macrophage class A scavenger receptor (CD204). Biochem. Biophys. Res. Commun 2011, 411, 516–522. [Google Scholar]
- Mukhopadhyay, S.; Varin, A.; Chen, Y.; Liu, B.; Tryggvason, K.; Gordon, S. SR-A/MARCO-mediated ligand delivery enhances intracellular TLR and NLR function, but ligand scavenging from cell surface limits TLR4 response to pathogens. Blood 2011, 117, 1319–1328. [Google Scholar]
- Vishnyakova, T.G.; Kurlander, R.; Bocharov, A.V.; Baranova, I.N.; Chen, Z.; Abu-Asab, M.S.; Tsokos, M.; Malide, D.; Basso, F.; Remaley, A.; et al. CLA-1 and its splicing variant CLA-2 mediate bacterial adhesion and cytosolic bacterial invasion in mammalian cells. Proc. Natl. Acad. Sci. USA 2006, 103, 16888–16893. [Google Scholar]
- Li, B.; Li, J.; Pan, X.C.; Ding, G.F.; Cao, H.W.; Jiang, W.W.; Zheng, J.; Zhou, H. Artesunate protects sepsis model mice challenged with Staphylococcus aureus by decreasing TNF-α release via inhibition TLR2 and Nod2 mRNA expressions and transcription factor NF-κB activation. Int. Immunopharmacol 2010, 10, 344–350. [Google Scholar]
- Karki, R.; Park, C.H.; Kim, D.W. Extract of buckwheat sprouts scavenges oxidation and inhibits pro-inflammatory mediators in lipopolysaccharide-stimulated macrophages (RAW264.7). J. Integr. Med 2013, 11, 246–252. [Google Scholar]
- Scott, M.J.; Billiar, T.R. β2-integrin-induced p38 MAPK activation is a key mediator in the CD14/TLR4/MD2-dependent uptake of lipopolysaccharide by hepatocytes. J. Biol. Chem 2008, 283, 29433–29446. [Google Scholar]
- Trinchieri, G.; Sher, A. Cooperation of Toll-like receptor signals in innate immune defence. Nat. Rev. Immunol 2007, 7, 179–190. [Google Scholar]
- Deng, M.; Scott, M.J.; Loughran, P.; Gibson, G.; Sodhi, C.; Watkins, S.; Hackam, D.; Billiar, T.R. Lipopolysaccharide clearance, bacterial clearance, and systemic inflammatory responses are regulated by cell type-specific functions of TLR4 during sepsis. J. Immunol 2013, 190, 5152–5160. [Google Scholar]
- Wang, Y.; Yang, Y.; Liu, X.; Wang, N.; Cao, H.; Lu, Y.; Zhou, H.; Zheng, J. Inhibition of clathrin/dynamin-dependent internalization interferes with LPS-mediated TRAM-TRIF-dependent signaling pathway. Cell Immunol 2012, 274, 121–129. [Google Scholar]
- Dunzendorfer, S.; Lee, H.K.; Soldau, K.; Tobias, P.S. TLR4 is the signaling but not the lipopolysaccharide uptake receptor. J. Immunol 2004, 173, 1166–1170. [Google Scholar]
- Areschoug, T.; Waldemarsson, J.; Gordon, S. Evasion of macrophage scavenger receptor A-mediated recognition by pathogenic streptococci. Eur. J. Immunol 2008, 38, 3068–3079. [Google Scholar]
- Hampton, R.Y.; Golenbock, D.T.; Penman, M.; Krieger, M.; Raetz, C.R. Recognition and plasma clearance of endotoxin by scavenger receptors. Nature 1991, 352, 342–344. [Google Scholar]
- Krieger, M. The other side of scavenger receptors: Pattern recognition for host defense. Curr. Opin. Lipidol 1997, 8, 275–280. [Google Scholar]
- Tsujita, K.; Kaikita, K.; Hayasaki, T.; Honda, T.; Kobayashi, H.; Sakashita, N.; Suzuki, H.; Kodama, T.; Ogawa, H.; Takeya, M. Targeted deletion of class A macrophage scavenger receptor increases the risk of cardiac rupture after experimental myocardial infarction. Circulation 2007, 115, 1904–1911. [Google Scholar]
- Mukhopadhyay, S.; Chen, Y.; Sankala, M.; Peiser, L.; Pikkarainen, T.; Kraal, G.; Tryggvason, K.; Gordon, S. MARCO, an innate activation marker of macrophages, is a class A scavenger receptor for Neisseria meningitidis. Eur. J. Immunol 2006, 36, 940–949. [Google Scholar]
- Guo, L.; Song, Z.; Li, M.; Wu, Q.; Wang, D.; Feng, H.; Bernard, P.; Daugherty, A.; Huang, B.; Li, X.A. Scavenger receptor BI protects against septic death through its role in modulating inflammatory response. J. Biol. Chem 2009, 284, 19826–19834. [Google Scholar]
- Baranova, I.; Vishnyakova, T.; Bocharov, A.; Chen, Z.; Remaley, A.T.; Stonik, J.; Eggerman, T.L.; Patterson, A.P. Lipopolysaccharide down regulates both scavenger receptor B1 and ATP binding cassette transporter A1 in RAW cells. Infect. Immun 2002, 70, 2995–3003. [Google Scholar]
- Guo, Y.B.; Zheng, J.; Zhou, H.; Lv, G.F.; Wang, L.X.; Wei, Guo; Lu, Y.L. A synthesized cationic tetradecapeptide from hornet venom kills bacteria and neutralizes lipopolysaccharide in vivo and in vitro. Biochem. Pharmacol. 2005, 70, 209–219. [Google Scholar]
- Kenneth, J.; Livak Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△CT methods. Methods 2001, 25, 402–408. [Google Scholar]
- Wu, S.; Liu, B.; Zhang, Q.; Liu, J.; Zhou, W.; Wang, C.; Li, M.; Bao, S.; Zhu, R. Dihydromyricetin reduced Bcl-2 expression via p53 in human hepatoma hepg2 cells. PLoS One 2013, 8, e76886. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Product size |
|---|---|---|
| SR-A | PF: 5′-ATTGGGAAATGAAGAACTGC-3′ PR:5′-GGACTGACGAAATCAAGGAA-3′ | 267 bp |
| SR-BI | PF: 5′-CCACCCAACGAAGGCTTCTGC-3′ PR: 5′-CTGAATGGCCTCCTTATCC-3′ | 190 bp |
| MARCO | PF: 5′-AGAAAGGGAGACACTGGAAGC-3′ PR: 5′-CCTCTGGAGTAACCGAGCAT-3′ | 274 bp |
| β-actin | PF: 5′-GAGACCTTCAACACCCCAGC-3′ PR: 5′-ATGTCACGCACGATTTCCC-3′ | 263 bp |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, B.; Yu, M.; Pan, X.; Ren, C.; Peng, W.; Li, X.; Jiang, W.; Zheng, J.; Zhou, H. Artesunate Reduces Serum Lipopolysaccharide in Cecal Ligation/Puncture Mice via Enhanced LPS Internalization by Macrophages through Increased mRNA Expression of Scavenger Receptors. Int. J. Mol. Sci. 2014, 15, 1143-1161. https://doi.org/10.3390/ijms15011143
Li B, Yu M, Pan X, Ren C, Peng W, Li X, Jiang W, Zheng J, Zhou H. Artesunate Reduces Serum Lipopolysaccharide in Cecal Ligation/Puncture Mice via Enhanced LPS Internalization by Macrophages through Increased mRNA Expression of Scavenger Receptors. International Journal of Molecular Sciences. 2014; 15(1):1143-1161. https://doi.org/10.3390/ijms15011143
Chicago/Turabian StyleLi, Bin, Mengchen Yu, Xichun Pan, Chuanliang Ren, Wei Peng, Xiaoli Li, Weiwei Jiang, Jiang Zheng, and Hong Zhou. 2014. "Artesunate Reduces Serum Lipopolysaccharide in Cecal Ligation/Puncture Mice via Enhanced LPS Internalization by Macrophages through Increased mRNA Expression of Scavenger Receptors" International Journal of Molecular Sciences 15, no. 1: 1143-1161. https://doi.org/10.3390/ijms15011143