Berry Phenolics of Grapevine under Challenging Environments


Abstract
:1. Introduction
2. Metabolism and Compartmentation of Phenolics in the Grape Berry
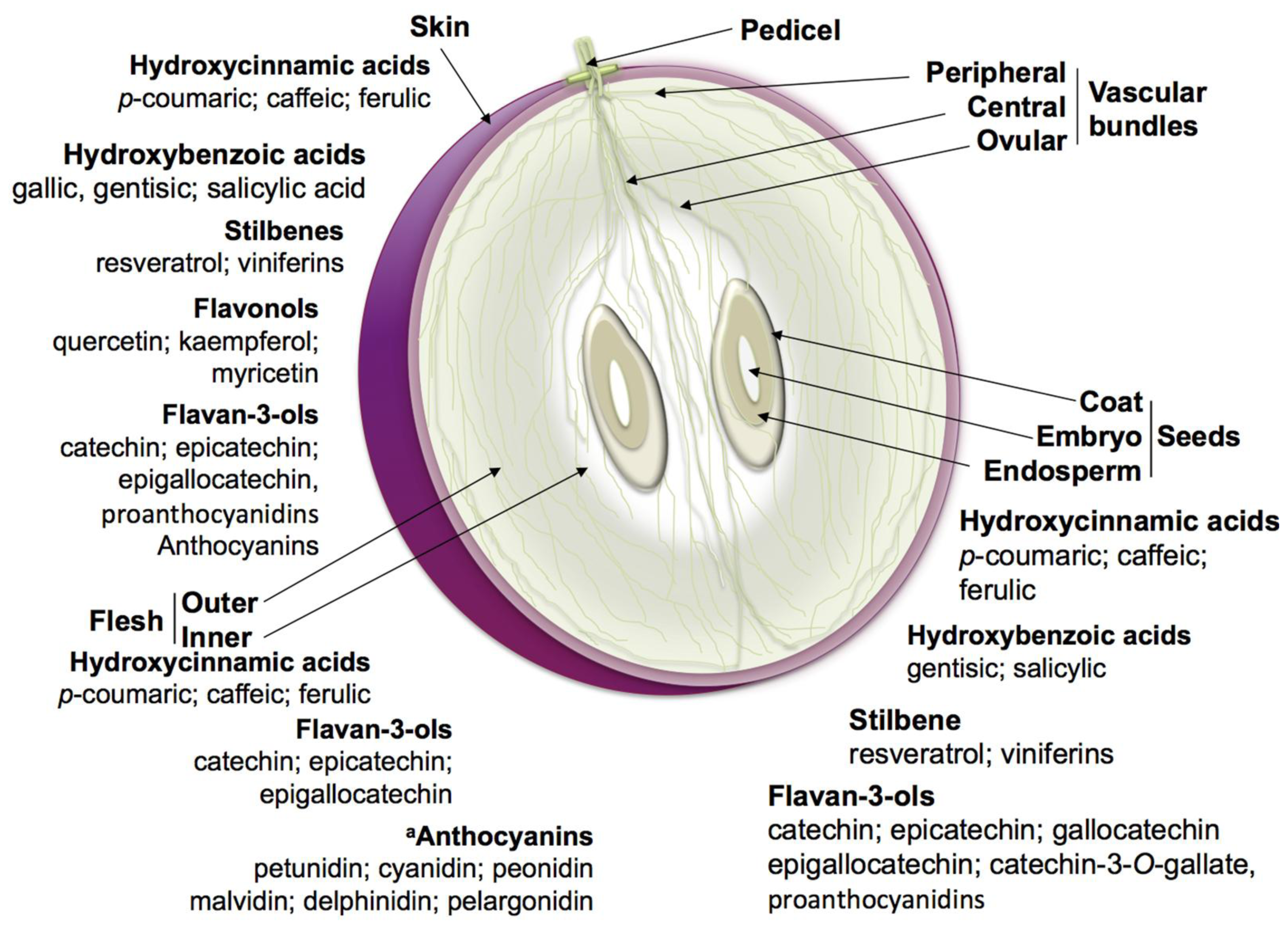
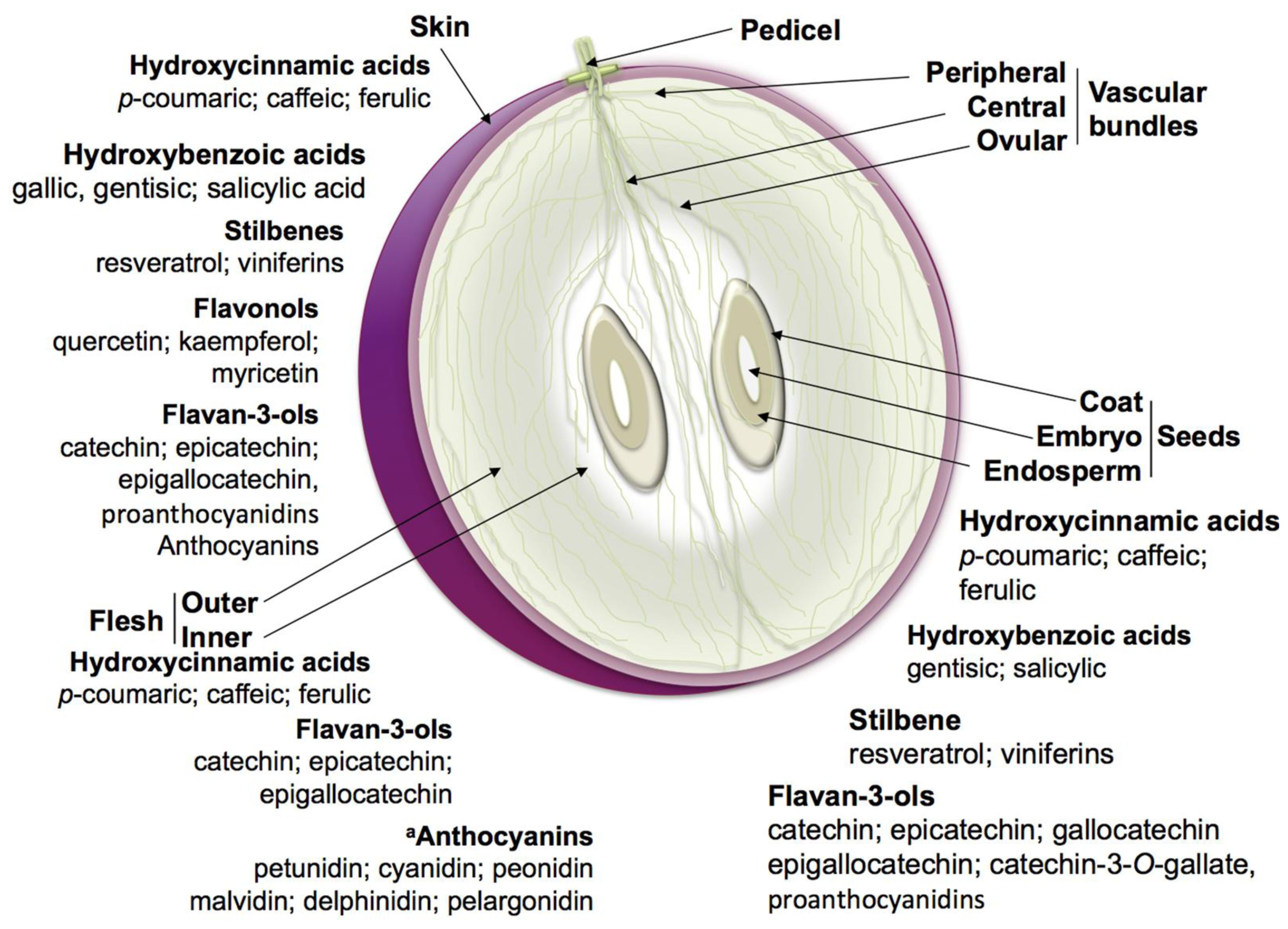
2.1. Nonflavonoid Phenolics
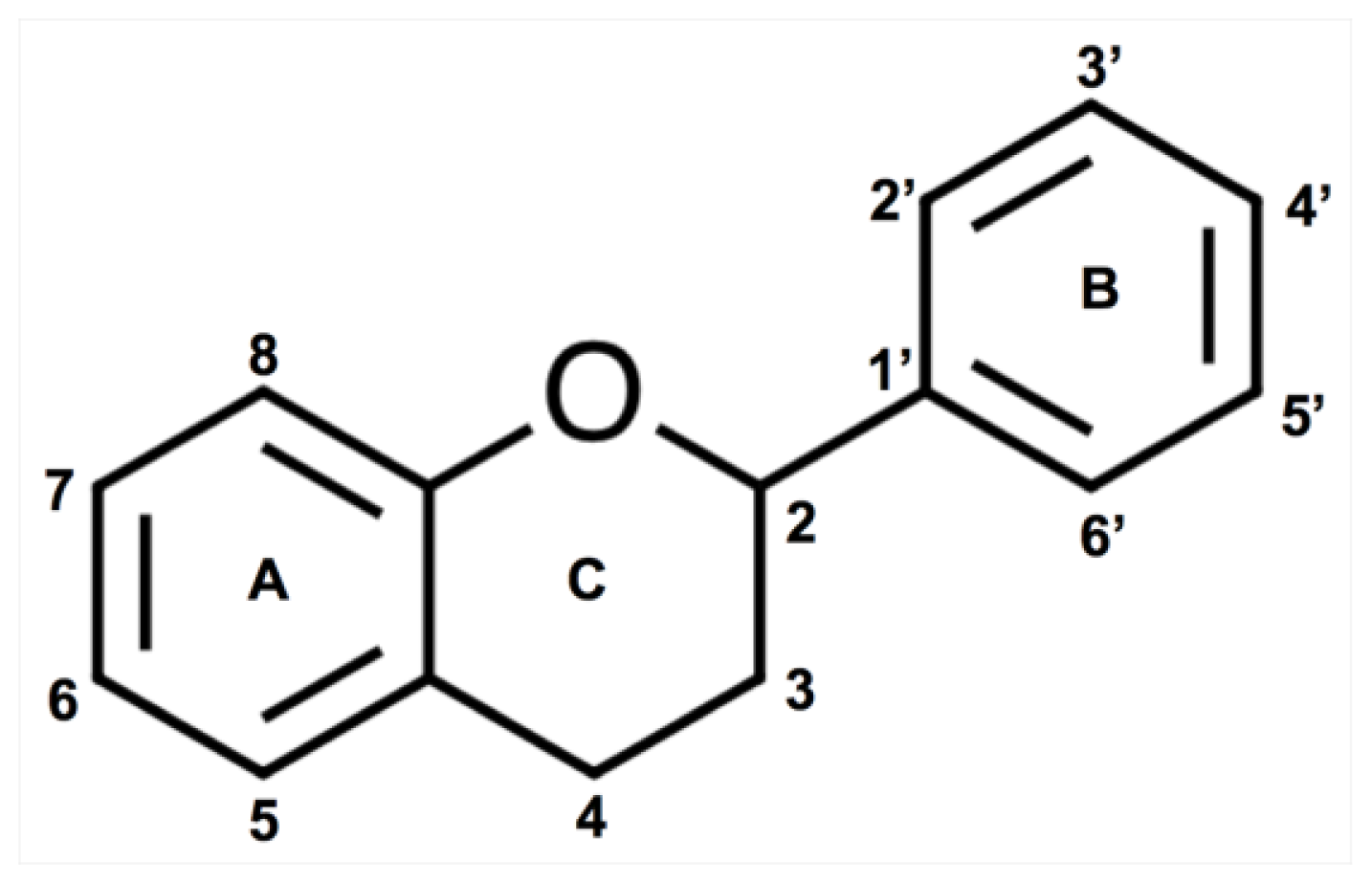
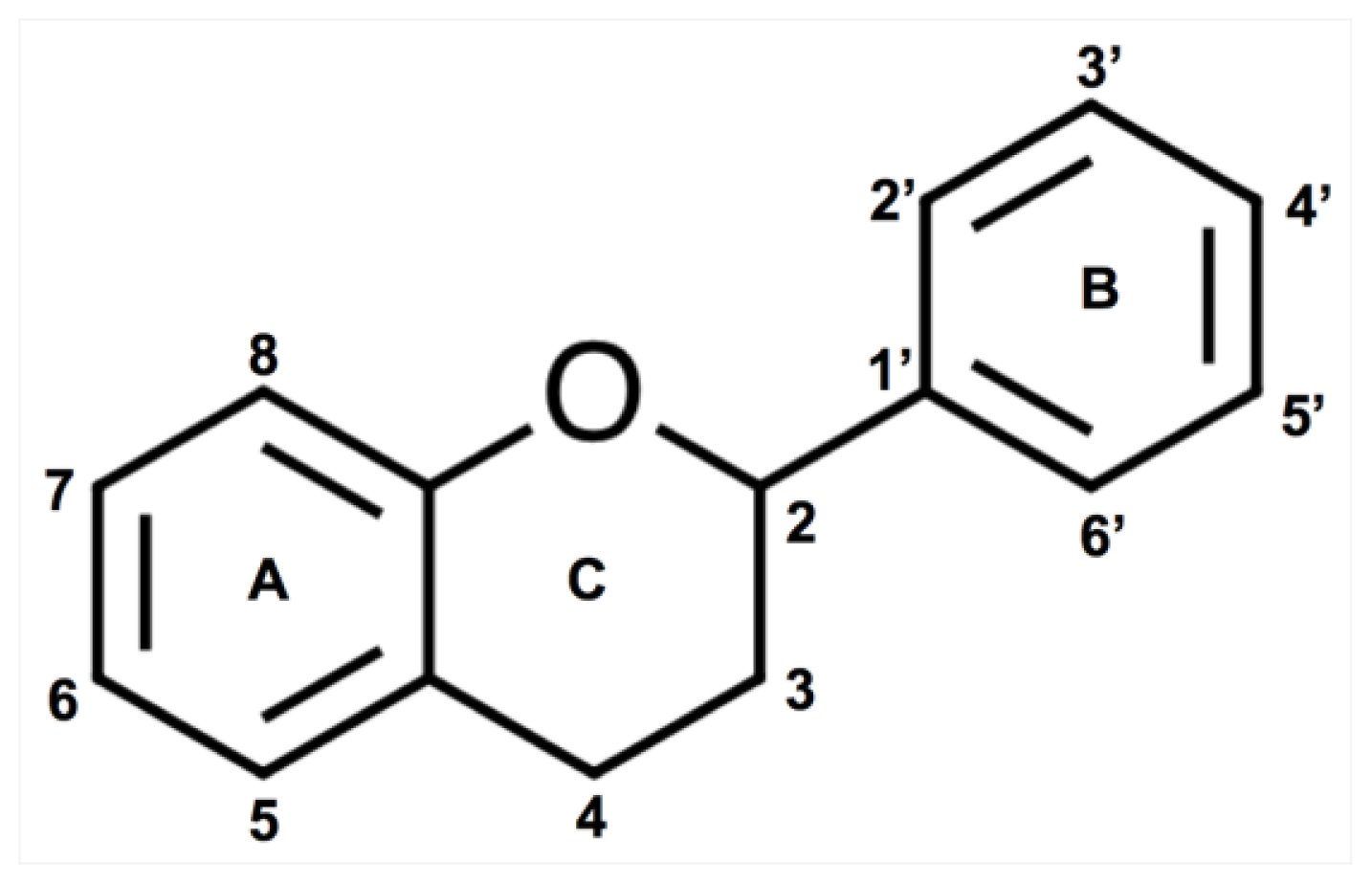
2.2. Flavonoids
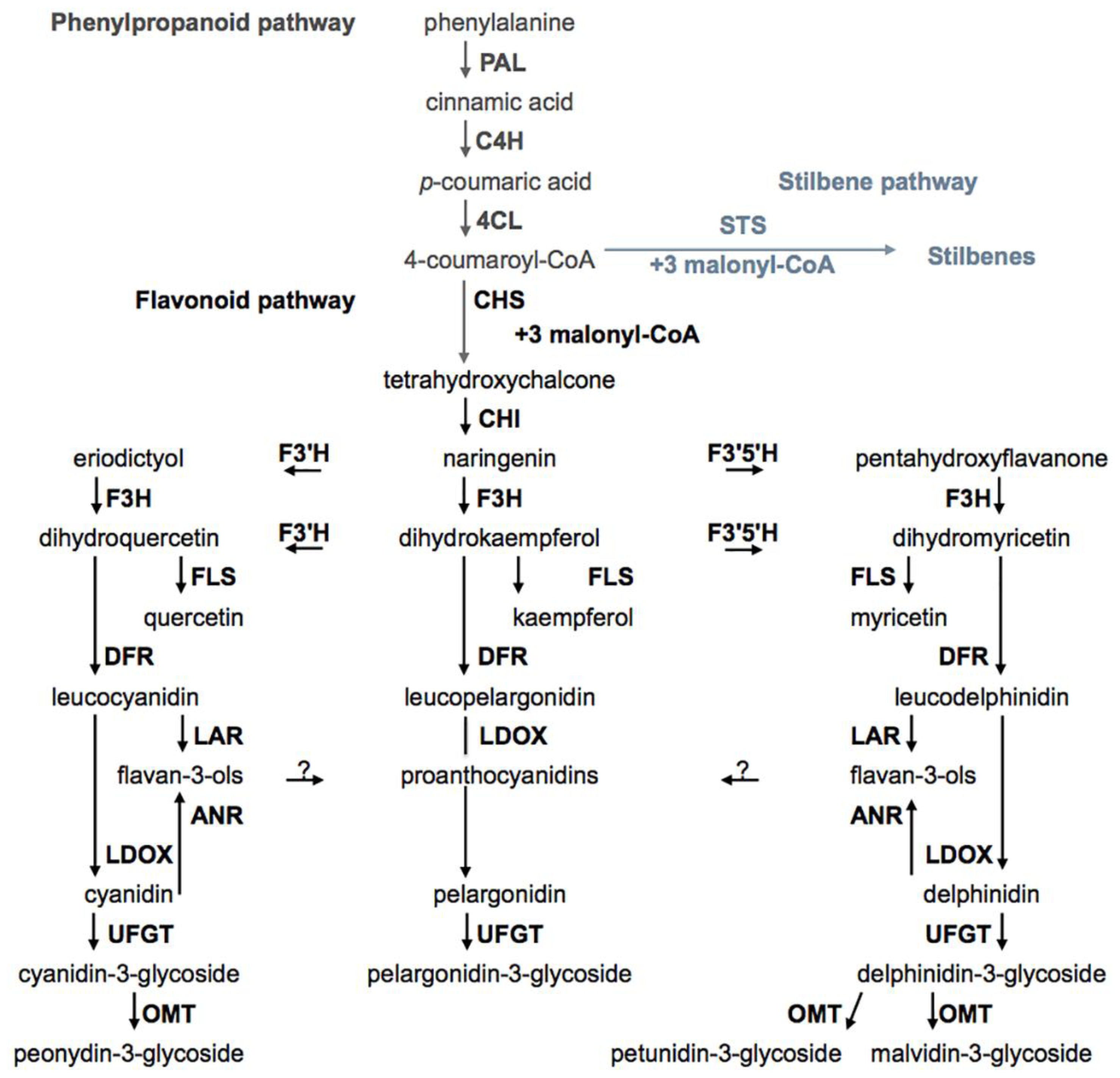
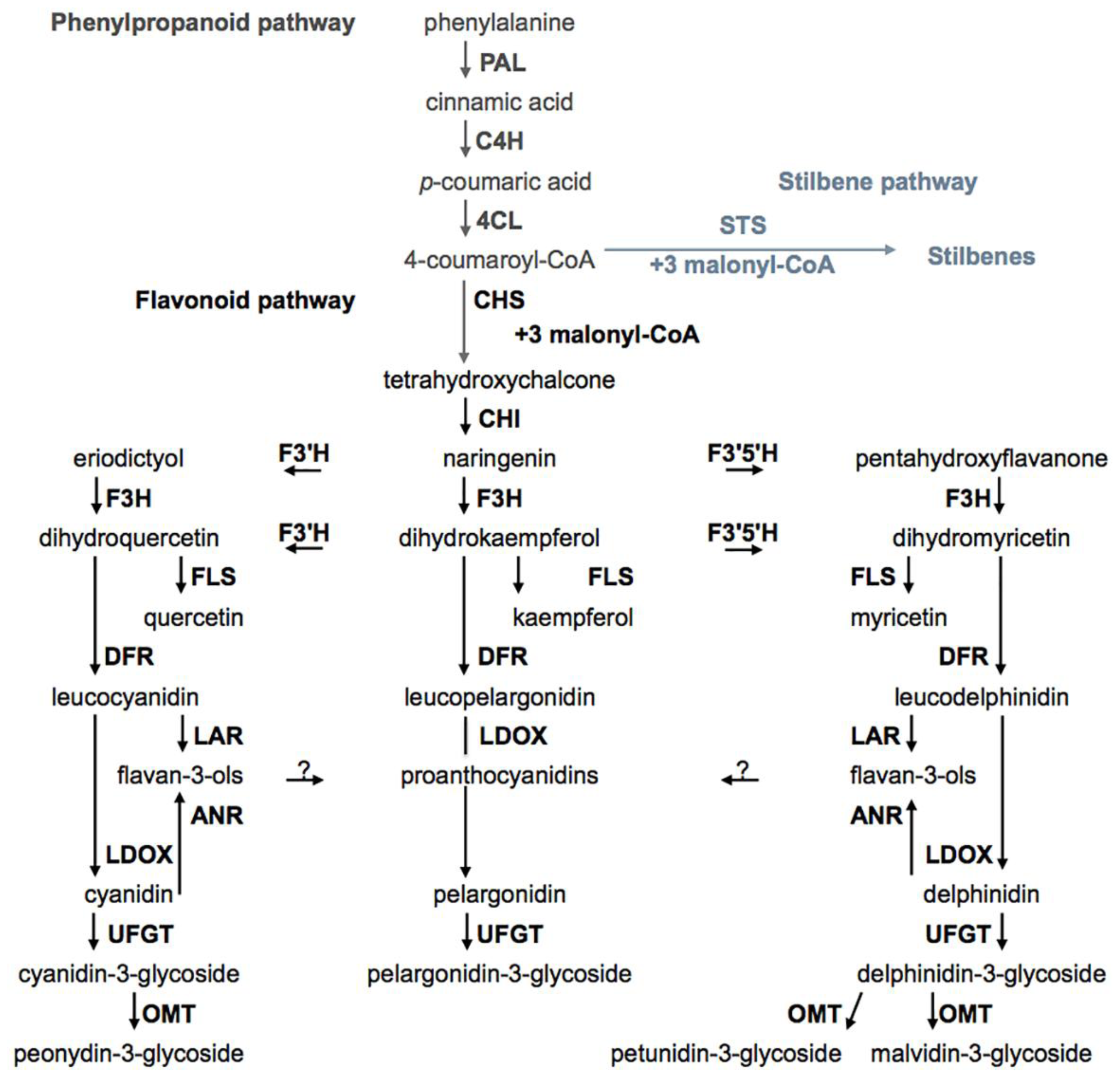
2.3. Biosynthesis Pathways of Phenolic Compounds in Wine Grape
3. Impact of Environment and Agricultural Practices in Grape Berry Phenolics
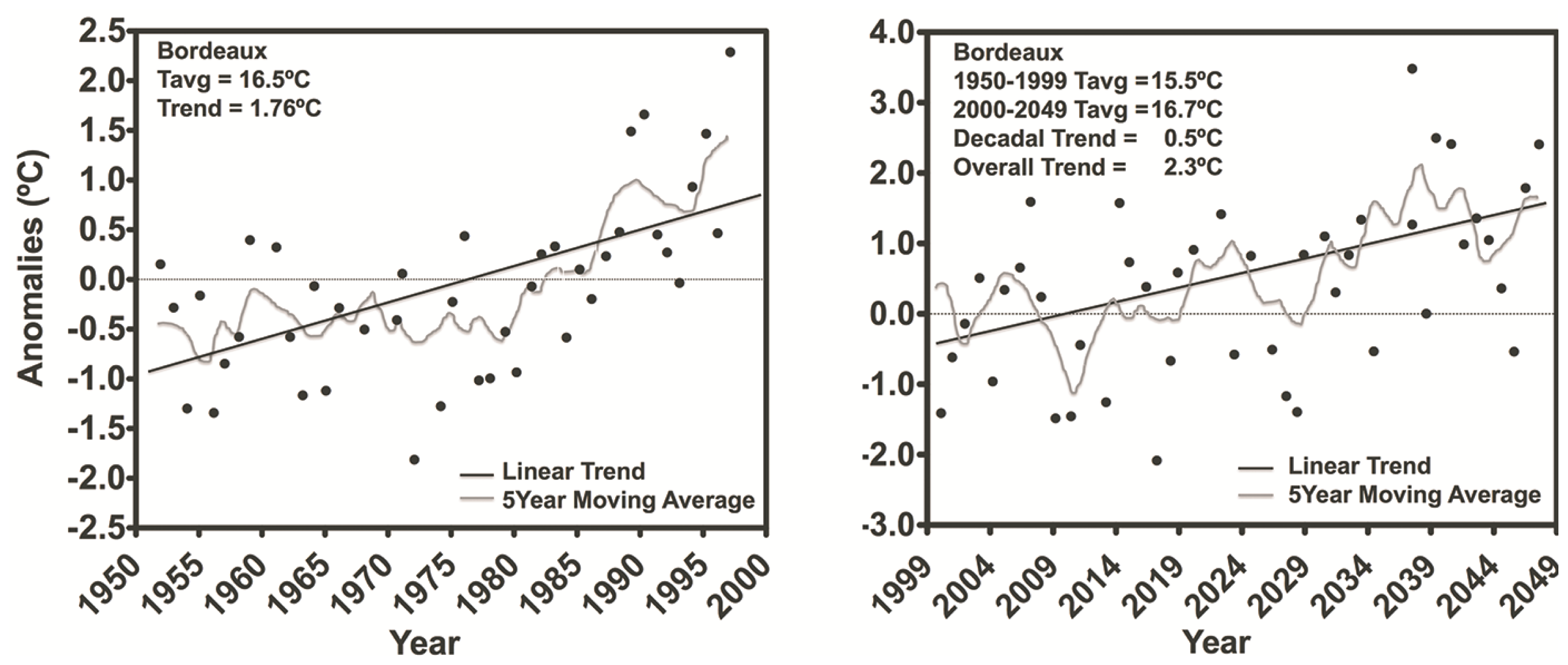
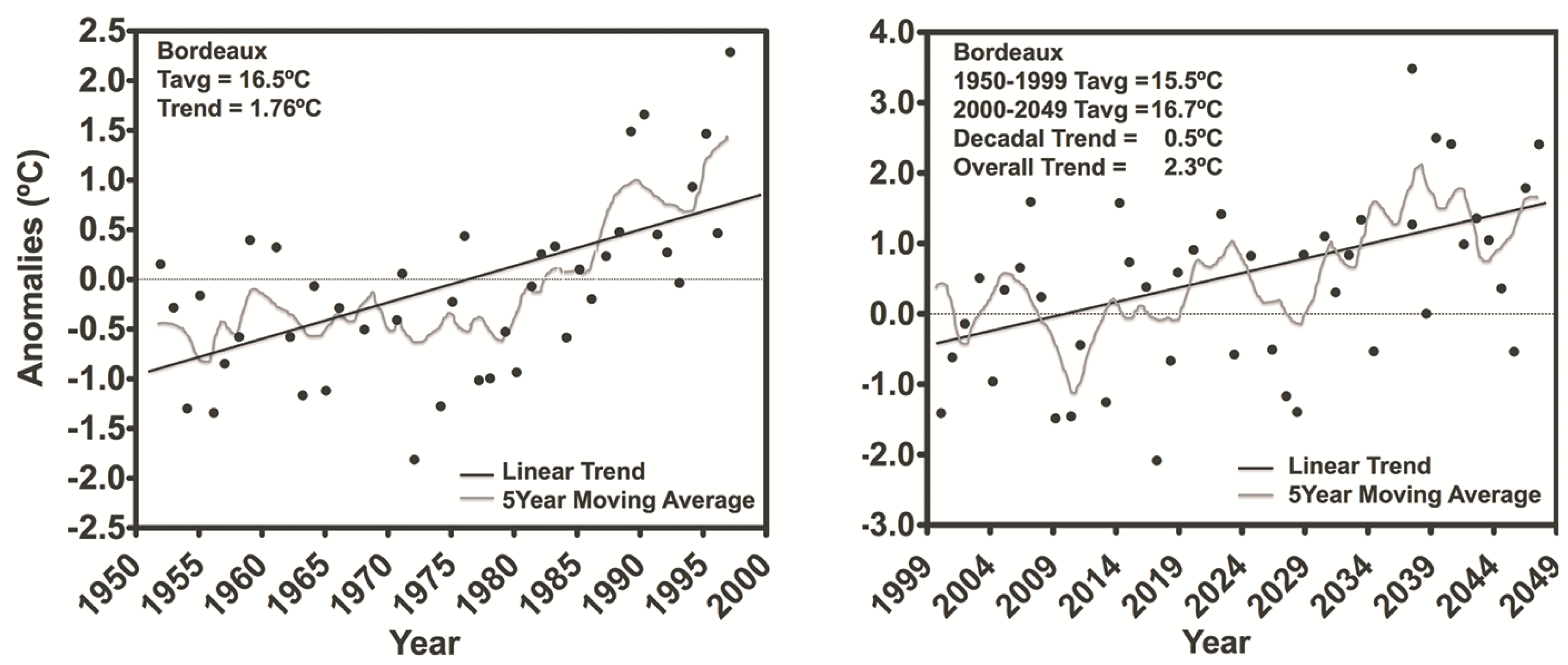
3.1. Temperature and Radiation
3.2. Agricultural Practices and the Levels of Synthesized Metabolites
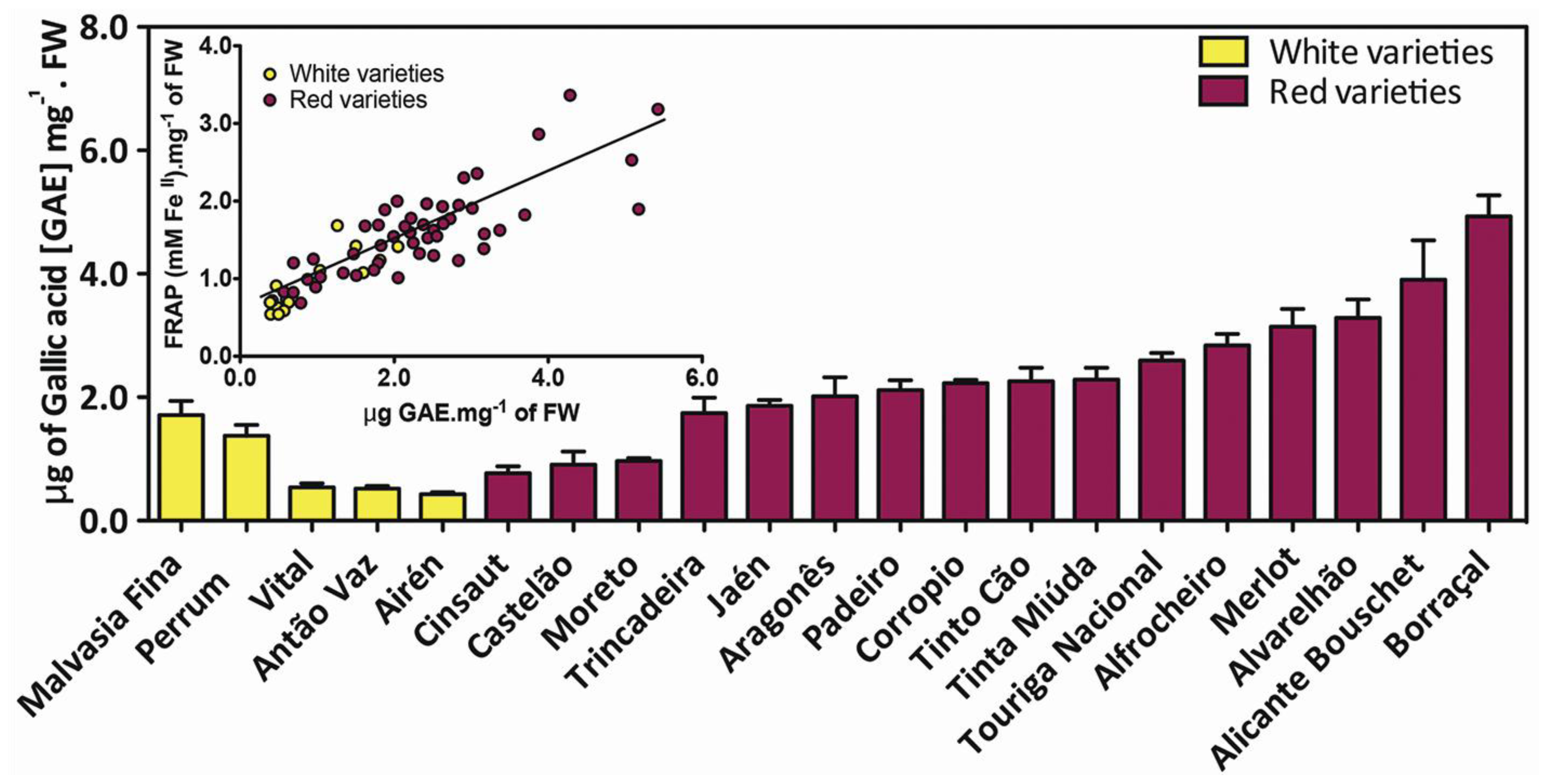
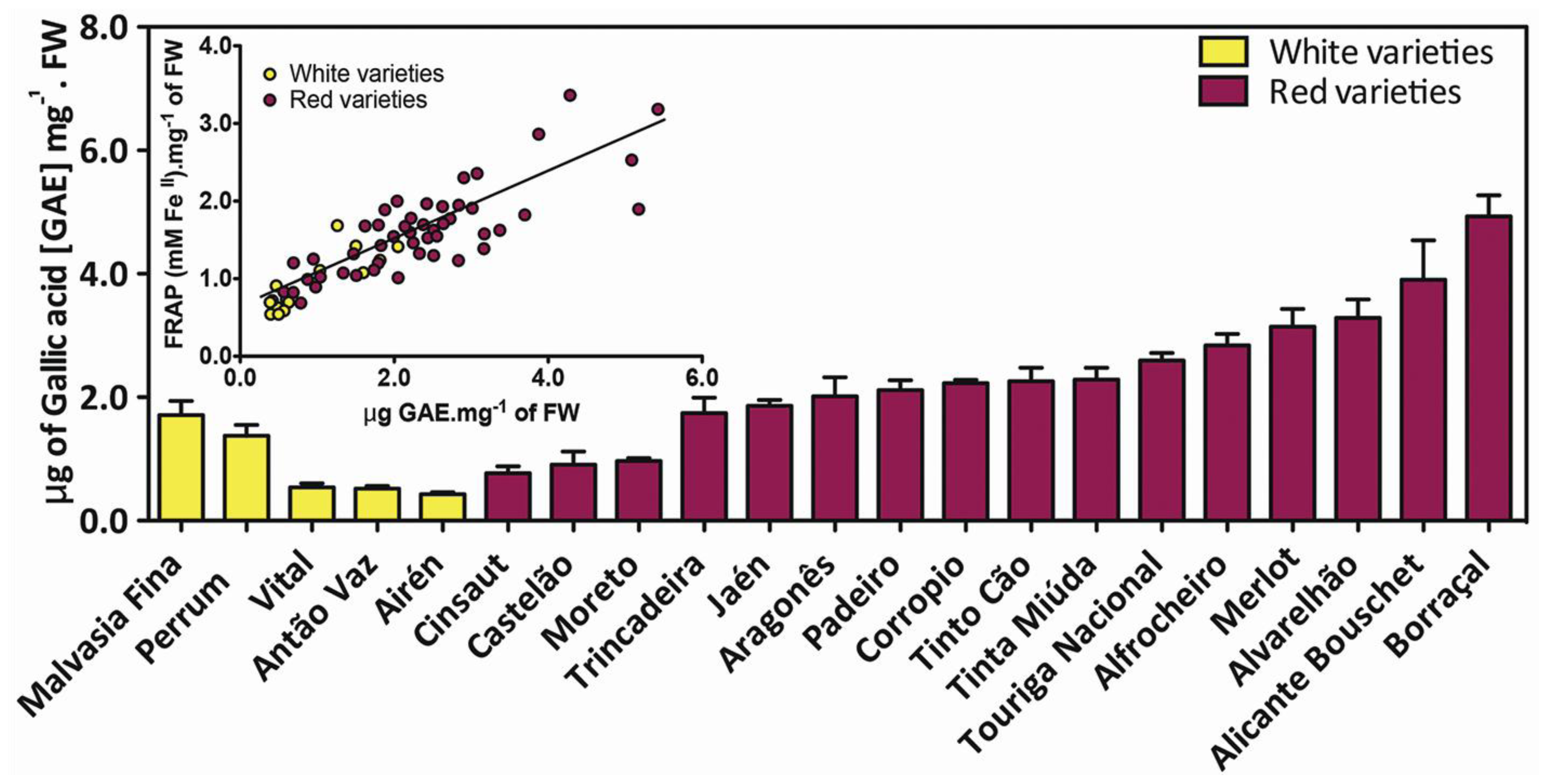
4. Varietal Dependence on Grape Berry Phenolics
5. Conclusions and Future Perspectives
Acknowledgements
Conflicts of Interest
References
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and wine phenolics: History and perspective. Am. J. Enol. Vitic 2006, 3, 20–21. [Google Scholar]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Castellarin, S.D.; Bavaresco, L.; Falginella, L.; Gonçalves, M.I.V.Z.; di Gaspero, G. Phenolics in Grape Berry and Key Antioxidants. In The Biochemistry of the grape berry; Gerós, H., Chaves, M., Delrot, S., Eds.; Bentham Science: Bussum, The Netherlands, 2012; pp. 89–110. [Google Scholar]
- Braidot, E.; Zancani, M.; Petrussa, E.; Peresson, C.; Bertolini, A.; Patui, S.; Macrì, F. Transport and accumulation of flavonoids in grapevine (Vitis vinifera L.). Plant Signal. Behav 2008, 3, 626–632. [Google Scholar]
- Martinoia, E.; Meyer, S.; de Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Ann. Rev. Plant Biol 2012, 63, 183–213. [Google Scholar]
- Adams, D. Phenolics and ripening in grape berries. Am. J. Enol. Vitic 2006, 3, 249–256. [Google Scholar]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and genotype control stilbene accumulation in healthy grapes. J. Agric. Food Chem 2008, 56, 11773–11785. [Google Scholar]
- Bavaresco, L.; Cantù, E.; Fregoni, M.T.M. Constitutive stilbene contents of grapevine cluster stems as potential source of resveratrol in wine. Vitis 1997, 36, 115–118. [Google Scholar]
- Fontes, N.; Gerós, H.; Delrot, S. Grape berry vacuole: A complex and heterogeneous membrane system specialized in the accumulation of solutes. Am. J. Enol. Vitic 2011, 62, 270–278. [Google Scholar]
- Castellarin, S.D.; Gambetta, G.A.; Wada, H.; Shackel, K.A.; Matthews, M.A. Fruit ripening in Vitis vinifera: Spatiotemporal relationships among turgor, sugar accumulation, and anthocyanin biosynthesis. J. Exp. Bot. 2011, 62, 4345–4354. [Google Scholar]
- Coombe, B.G. Influence of temperature on composition and quality of grapes. Acta Hort 1987, 206, 23–36. [Google Scholar]
- Cadot, Y.; Miñana-Castelló, M.T.; Chevalier, M. Anatomical, histological, and histochemical changes in grape seeds from Vitis vinifera L. cv Cabernet franc during fruit development. J. Agric. Food Chem 2006, 54, 9206, –9215... [Google Scholar]
- Easterling, D.; Evans, J.; Groisman, P.; Karl, T.; Kunkel, K.E.; Ambenje, P. Observed variability and trends in extreme climate events: A brief review. Bull. Am. Meteorol. Soc 2000, 81, 417–425. [Google Scholar]
- Chuine, I.; Yiou, P.; Seguin, B.; Daux, V.; Le, E.; Ladurie, R.; Parker, D.E. Historical phenology: Grape ripening as a past climate indicator. Nature 2004, 432, 289–290. [Google Scholar]
- Mann, M.E.; Woodruff, J.D.; Donnelly, J.P.; Zhang, Z. Atlantic hurricanes and climate over thepast 1,500 years. Nature 2009, 460, 880–883. [Google Scholar]
- Cohen, S.D.; Tarara, J.M.; Gambetta, G.A.; Matthews, M.A.; Kennedy, J.A. Impact of diurnal temperature variation on grape berry development, proanthocyanidin accumulation, and the expression of flavonoid pathway genes. J. Exp. Bot 2012, 63, 2655–2665. [Google Scholar]
- Kenny, G.J.; Harrison, P.A. The effects of climate variability and change on grape suitability in Europe. J. Wine Res 1992, 3, 163–183. [Google Scholar]
- Schultz, H.R.; Jones, G.V. Climate induced historic and future changes in Viticulture. J. Wine Res 2010, 21, 137–145. [Google Scholar]
- Owens, C.L. Grapes. Temperate Fruit Crop Breeding: Germplasm To Genomics; Hancock, J., Ed.; Springer: Heidelberg, Germany, 2008; pp. 197–233. [Google Scholar]
- Liang, Z.; Owens, C.L.; Zhong, G.Y.; Cheng, L. Polyphenolic profiles detected in the ripe berries of Vitis vinifera germplasm. Food Chem 2011, 129, 940–950. [Google Scholar]
- Singleton, V.; Zaya, J.; Trousdale, E. Caftaric and coutaric acids in fruit of Vitis. Phytochemistry 1986, 25, 2127–2133. [Google Scholar]
- Waterhouse, A.L. Wine phenolics. Ann. N. Y. Acad. Sci 2002, 957, 21–36. [Google Scholar]
- Vanhoenacker, G.; de Villiers, A.; Lazou, K.; de Keukeleire, D.; Sandra, P. Comparison of high-performance liquid chromatography—Mass spectroscopy and capillary electrophoresis—Mass spectroscopy for the analysis of phenolic compounds in diethyl ether extracts of red wines extraction of phenolic compounds LC-UV-MS high-per. Chromatographia 2001, 54, 309–315. [Google Scholar]
- Pozo-Bayón, M.A.; Hernández, M.T.; Martín-Alvarez, P.J.; Polo, M.C. Study of low molecular weight phenolic compounds during the aging of sparkling wines manufactured with red and white grape varieties. J. Agric. Food Chem 2003, 51, 2089–2095. [Google Scholar]
- Ali, K.; Maltese, F.; Choi, Y.H.; Verpoorte, R. Metabolic constituents of grapevine and grape-derived products. Phytochem. Rev 2010, 9, 357–378. [Google Scholar]
- Baderschneider, B.; Winterhalter, P. Isolation and characterization of novel benzoates, cinnamates, flavonoids, and lignans from Riesling wine and screening for antioxidant activity. J. Agric. Food Chem 2001, 49, 2788–2798. [Google Scholar]
- Harbertson, J.F.; Jennedy, J.A.; Adams, D. Tannin in skins and seeds of Cabernet Sauvignon, Syrah, and Pinot Noir berries during ripening. Am. J. Enol. Vitic 2002, 1, 54–59. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Simon, R. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine Res 2003, 9, 15–27. [Google Scholar]
- Pena-Neira, A.; Caceres, A.; Pastenes, C. Low molecular weight phenolic and anthocyanin composition of grape skins from cv. Syrah (Vitis vinifera L.) in the Maipo Valley (Chile): Effect of clusters thinning and vineyard yield. Food Sci. Tech. Int 2007, 13, 153–158. [Google Scholar]
- Montealegre, R.; Romero Peces, R.; Chacón Vozmediano, J.L.; Martínez Gascueña, J.; García Romero, E. Phenolic compounds in skins and seeds of ten grape Vitis vinifera varieties grown in a warm climate. J. Food Compos. Anal 2006, 19, 687–693. [Google Scholar]
- Langcake, P.; Pryce, R.J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Physiol. Plant Pathol 1976, 9, 77–86. [Google Scholar]
- Jackson, R. Grapevine Structure Function. In Wine Science—Principles and Aplications; Elsvier: San Diego, CA, USA, 2008; pp. 50–105. [Google Scholar]
- Mattivi, F.; Vrhovsek, U.; Masuero, D.; Trainotti, D. Differences in the amount and structure of extractable skin and seed tannins amongst red grape varieties. Aust. J. Grape Wine Res 2009, 15, 27–35. [Google Scholar]
- Hanlin, R.L.; Hrmova, M.; Harbertson, J.F.; Downey, M.O. Review: Condensed tannin and grape cell wall interactions and their impact on tannin extractability into wine. Aust. J. Grape Wine Res 2010, 16, 173–188. [Google Scholar]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci 2009, 177, 143–155. [Google Scholar]
- Mattivi, F.; Guzzon, R.; Vrhovsek, U.; Stefanini, M.; Velasco, R. Metabolite profiling of grape: Flavonols and anthocyanins. J. Agric. Food Chem 2006, 54, 7692–7702. [Google Scholar]
- Price, S.; Breen, P.; Valladao, M.; Watson, B. Cluster sun exposure and quercetin in Pinot noir grapes and wine. Am. J. Enol. Vitic 1995, 46, 187–194. [Google Scholar]
- Singleton, V.L. Tannins and the Qualities of Wines. In Plant Polyphenols; Springer: New York, NY, US, 1992; pp. 859–880. [Google Scholar]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, M.I. Quantification of catechins and proanthocyanidins in several Portuguese grapevine varieties and red wines. Ciência Téc. Vitiv 2001, 16, 23–34. [Google Scholar]
- Su, C.T.; Singleton, V. Identification of three flavan-3-ols from grapes. Phytochemistry 1969, 8, 1553–1558. [Google Scholar]
- Kennedy, J.A.; Hayasaka, Y.; Vidal, S.; Waters, E.J.; Jones, G.P. Composition of grape skin proanthocyanidins at different stages of berry development. J. Agric. Food Chem. 2001, 49, 5348–5355. [Google Scholar]
- Ribéreau-Gayon, P. Research on plant anthocyanins. In Applications to the genus Vitis; University of Bordeaux. Librairie de l’enseignement: Paris, France, 1959. [Google Scholar]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar]
- He, F.; He, J.J.; Pan, Q.H.; Duan, C.Q. Mass-spectrometry evidence confirming the presence of pelargonidin-3-O-glucoside in the berry skins of Cabernet Sauvignon and Pinot Noir (Vitis vinifera L.). Aust. J. Grape Wine Res 2010, 16, 464–468. [Google Scholar]
- Ford, C.M.; Boss, P.K.; Hoj, P.B. Cloning and characterization of Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase, a homologue of the enzyme encoded by the maize Bronze-1 locus that may primarily serve to glucosylate anthocyanidins in vivo. J. Biol. Chem 1998, 273, 9224–9233. [Google Scholar]
- Jánváry, L.; Hoffmann, T.; Pfeiffer, J.; Hausmann, L.; Töpfer, R.; Fischer, T.C.; Schwab, W. A double mutation in the anthocyanin 5-O-glucosyltransferase gene disrupts enzymatic activity in Vitis vinifera L. J. Agric. Food Chem 2009, 57, 3512–3518. [Google Scholar]
- Falginella, L.; di Gaspero, G.; Castellarin, S.D. Expression of flavonoid genes in the red grape berry of “Alicante Bouschet” varies with the histological distribution of anthocyanins and their chemical composition. Planta 2012, 236, 1037–1051. [Google Scholar]
- Hrazdina, G.; Parsons, G.F.; Mattick, L.R. Physiological and biochemical events during development and maturation of grape berries. Am. J. Enol. Vitic 1984, 35, 220–227. [Google Scholar]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M.; et al. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar]
- Conn, S.; Curtin, C.; Bézier, A. Cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J. Exp. Bot 2008, 59, 3621–3634. [Google Scholar]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verriès, C.; Souquet, J.M.; Mazauric, J.P.; Klein, M.; Cheynier, V.; et al. Grapevine MATE-type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. Plant Physiol 2009, 150, 402–415. [Google Scholar]
- Gomez, C.; Conejero, G.; Torregrosa, L.; Cheynier, V.; Terrier, N.; Ageorges, A. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST. Plant. J 2011, 67, 960–970. [Google Scholar]
- Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int 2010, 43, 1844–1855. [Google Scholar]
- Stock, M.; Gerstengarbe, F.W.; Kartschall, T.; Werner, P. Reliability of climate change impact assessments for viticulture. Acta Hortic 2004, 689, 29–39. [Google Scholar]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar]
- Jones, G.; White, M.; Cooper, O.; Storchmann, K. Climate change and global wine quality. Clim. Change 2005, 73, 319–343. [Google Scholar]
- Buttrose, M.; Hale, C. Effects of temperature on accumulation of starch or lipid in chloroplasts of grapevine. Planta 1971, 101, 166–170. [Google Scholar]
- Hale, C.R.; Buttrose, M. Effect of temperature on ontogeny of berries of Vitis vinifera L. cv. Cabernet Sauvignon. J. Am. Soc. Hortic. Sci 1973, 99, 390–394. [Google Scholar]
- Schultz, H. Climate change and viticulture: A European perspective on climatology, carbon dioxide and UV-B effects. Aust. J. Grape Wine Res 2000, 6, 2–12. [Google Scholar]
- Camps, J.O.; Ramos, M.C. Grape harvest and yield responses to inter-annual changes in temperature and precipitation in an area of north-east Spain with a Mediterranean climate. Int. J. Biometeorol 2012, 56, 853–864. [Google Scholar]
- Ganichot, B. Evolution of harvesting dates in meridional Rhône. Proceedings of 6th Rencontres rhodaiennes, Orange, France; 2002; pp. 38–41. [Google Scholar]
- Duchêne, E.; Schneider, C. Grapevine and climatic changes: A glance at the situation in Alsace. Agron. Sustain. Dev 2005, 25, 93–99. [Google Scholar]
- Laget, F.; Tondut, J.; Deloire, A.; Kelly, M. Climate trends in a specific Mediterranean viticultural area between 1950 and 2006 (in France). J. Int. Sci. Vigne 2008, 42, 113–123. [Google Scholar]
- Jones, G.; Davis, R. Climate influences on grapevine phenology, grape composition, and wine production and quality for Bordeaux, France. Am. J. Enol. Vitic 2000, 51, 249–261. [Google Scholar]
- Petrie, P.R.; Sadras, V.O. Advancement of grapevine maturity in Australia between 1993 and 2006: Putative causes, magnitude of trends and viticultural consequences. Aust. J. Grape Wine Res 2008, 14, 33–45. [Google Scholar]
- Webb, L.B.; Whetton, P.H.; Barlow, E.W.R. Observed trends in winegrape maturity in Australia. Glob. Change Biol 2011, 17, 2707–2719. [Google Scholar]
- Roubelakis-Angelakis, K.A.; Kliewer, W.M. Effects of exogenous factors on phenylalanine ammonia-lyase activity and accumulation of anthocyanins and total phenolics in grape berries. Am. J. Enol. Vitic 1986, 37, 275–280. [Google Scholar]
- Keller, M.; Torres-Martinez, N. Does UV radiation affect winegrape composition? Acta Hortic 2004, 640, 313–319. [Google Scholar]
- Schreiner, M.; Mewis, I.; Huyskens-Keil, S.; Jansen, M.A.K.; Zrenner, R.; Winkler, J.B.; O’Brien, N.; Krumbein, A. UV-B-induced secondary plant metabolites—Potential benefits for plant and human health. Crit. Rev. Plant Sci 2012, 31, 229–240. [Google Scholar]
- Berli, F.J.; Moreno, D.; Piccoli, P.; Hespanhol-Viana, L.; Silva, M.F.; Bressan-Smith, R.; Cavagnaro, J.B.; Bottini, R. Abscisic acid is involved in the response of grape (Vitis vinifera L.) cv. Malbec leaf tissues to ultraviolet-B radiation by enhancing ultraviolet-absorbing compounds, antioxidant enzymes and membrane sterols. Plant Cell Environ 2010, 33, 1–10. [Google Scholar]
- Jordan, B.; He, J.; Chow, W.S.; Anderson, J. Changes in mRNA levels and polypeptide subunits of ribulose 1,5-bisphosphate carboxylase in response to supplementary ultraviolet-B radiation. Plant Cell Environ 1992, 15, 91–98. [Google Scholar]
- Berli, F.; D’Angelo, J.; Cavagnaro, B.; Bottini, R.; Wuilloud, R.; Silva, M.F. Phenolic composition in grape (Vitis vinifera L. cv. Malbec) ripened with different solar UV-B radiation levels by capillary zone electrophoresis. J. Agric. Food Chem 2008, 56, 2892–2898. [Google Scholar]
- Kolb, C.A.; Käser, M.A.; Kopecký, J.; Zotz, G.; Riederer, M.; Pfündel, E.E. Effects of natural intensities of visible and ultraviolet radiation on epidermal ultraviolet screening and photosynthesis in grape leaves. Plant Physiol 2001, 127, 863–875. [Google Scholar]
- Keller, M. Managing grapevines to optimise fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res 2010, 16, 56–69. [Google Scholar]
- Petit, A-N.; Baillieul, F.; Vaillant-Gaveau, N.; Jacquens, L.; Conreux, A.; Jeandet, P.; Clément, C.; Fontaine, F. Low responsiveness of grapevine flowers and berries at fruit set to UV-C irradiation. J. Exp. Bot. 2009, 60, 1155–1162. [Google Scholar]
- Sbaghi, M.; Jeandet, P.; Faivre, B.; Bessis, R.; Fournioux, J. Development of methods using phytoalexin (resveratrol) assessment as a selection criterion to screen grapevine in vitro cultures for resistance to grey mould (Botrytis). Euphytica 1995, 86, 41–47. [Google Scholar]
- Spayd, S.; Tarara, J. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic 2002, 3, 171–182. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Simon, R. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Aust. J. Grape Wine Res 2004, 10, 55–73. [Google Scholar]
- Czemmel, S.; Stracke, R.; Weisshaar, B.; Cordon, N.; Harris, N.N.; Walker, A.R.; Robinson, S.P.; Bogs, J. The grapevine R2R3-MYB transcription factor VvMYBF1 regulates flavonol synthesis in developing grape berries. Plant Physiol 2009, 151, 1513–1530. [Google Scholar]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar]
- Ewart, A.; Kliewer, W. Effects of controlled day and night temperatures and nitrogen on fruit-set, ovule fertility, and fruit composition of several wine grape cultivars. Am. J. Enol. Vitic 1977, 28, 88–95. [Google Scholar]
- Del Rio, J.; Kennedy, J. Development of proanthocyanidins in Vitis vinifera L. cv. Pinot Noir grapes and extraction into wine. Am. J. Enol. Vitic 2006, 2, 125–132. [Google Scholar]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem 2006, 54, 8510–8520. [Google Scholar]
- Downey, M.O.; Dokoozlian, N.K.; Krstic, M. Cultural practice and environmental impacts on the flavonoid composition of grapes and wine: A review of recent research. Am. J. Enol. Vitic 2006, 3, 257–268. [Google Scholar]
- Guidoni, S.; Ferrandino, A.; Vittorino, N. Effects of seasonal and agronomical practices on skin anthocyanin profile of Nebbiolo grapes. Am. J. Enol. Vitic 2008, 1, 22–29. [Google Scholar]
- Tarara, J.; Lee, J.; Spayd, E.S. Berry temperature and solar radiation alter acylation, proportion, and concentration of anthocyanin in Merlot grapes. Am. J. Enol. Vitic 2008, 3, 235–247. [Google Scholar]
- Kliewer, W.M.; Torres, R.E. Effect of controlled day and night temperatures on grape coloration. Am. J. Enol. Vitic 1972, 23, 71–77. [Google Scholar]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot 2007, 58, 1935–1945. [Google Scholar]
- Cohen, S.D.; Tarara, J.M.; Kennedy, J.A. Assessing the impact of temperature on grape phenolic metabolism. Anal. Chim. Acta 2008, 621, 57–67. [Google Scholar]
- Kliewer, W. Influence of temperature, solar radiation and nitrogen on coloration and composition of Emperor grapes. Am. J. Enol. Vitic 1977, 28, 96–103. [Google Scholar]
- Delgado, R.; Martín, P.; Delálamo, M.; González, M. Changes in the phenolic composition of grape berries during ripening in relation to vineyard nitrogen and potassium fertilisation rates. J. Sci. Food Agr 2004, 84, 623–630. [Google Scholar]
- Russell, E.W. Soil conditions and plant growth. Soil Sci 1962, 93, 73. [Google Scholar]
- Northcote, K.H. Soils and Australian viticulture. In Viticulture; Winetitles: Adelaide, Australia, 1995; pp. 61–90. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK; p. 1995.
- Jackson, D.; Lombard, P. Environmental and management practices affecting grape composition and wine quality—A review. Am. J. Enol. Vitic 1993, 44, 409–430. [Google Scholar]
- McDonald, M.; Hughes, M. Survey of the free and conjugated myricetin and quercetin content of red wines of different geographical origins. J. Agric. Food Chem 1998, 8561, 368–375. [Google Scholar]
- Shaulis, N.; Amberg, H.; Crowe, D. Response of Concord grapes to light, exposure and Geneva double curtain training. J. Am. Soc. Hortic. Sci 1966, 89, 268–280. [Google Scholar]
- Pereira, G.E.; Gaudillere, J.P.; Pieri, P.; Hilbert, G.; Maucourt, M.; Deborde, C.; Moing, A.; Rolin, D. Microclimate influence on mineral and metabolic profiles of grape berries. J. Agric. Food Chem 2006, 54, 6765–6775. [Google Scholar]
- Smart, R.E.; Bobinson, J.B.; Due, G.R.; Brien, C. Canopy microclimate modification for the cultivar Shiraz. I. Definition of canopy microclimate. Vitis 1985, 24, 17–31. [Google Scholar]
- Haselgrove, L.; Botting, D.; Heeswijck, R.; Hog, P.B.; Dry, P.R.; Ford, C.; Iland, P. Canopy microclimate and berry composition: The effect of bunch exposure on the phenolic composition of Vitis vinifera L cv. Shiraz grape berries. Aust. J. Grape Wine Res 2000, 6, 141–149. [Google Scholar]
- Hunter, J.J.; de Villiers, O.T.; Watts, J.E. The effect of partial defoliation on quality characteristics of Vitis vinifera L. cv. Cabernet Sauvignon grapes. II. Skin color, skin sugar, and wine quality. Am. J. Enol. Vitic 1991, 42, 13–18. [Google Scholar]
- Dokoozlian, N.K.; Hirschfelt, D.J. The influence of cluster thinning at various stages of fruit development on Flame Seedless table grapes. Am. J. Enol. Vitic 1995, 46, 429–436. [Google Scholar]
- Castelan-Estrada, M.; Vivian, P.; Gaudillére, J. Allometric relationships to estimate seasonal above-ground vegetative and reproductive biomass of Vitis vinifera L. Ann. Bot 2002, 89, 401–408. [Google Scholar]
- Smart, R.; Dick, J.; Gravett, I.; Fisher, B. Canopy management to improve grape yield and wine quality-principles and practices. S. Afr. J. Enol. Vitic 1990, 11, 3–17. [Google Scholar]
- Cortell, J.M.; Halbleib, M.; Gallagher, A.V.; Righetti, T.L.; Kennedy, J.A. Influence of vine vigor on grape (Vitis vinifera L. cv. Pinot noir) and wine proanthocyanidins. J. Agric. Food Chem. 2005, 53, 5798–5808. [Google Scholar]
- Jeong, S.T.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci 2004, 167, 247–252. [Google Scholar]
- Koyama, K.; Goto-Yamamoto, N. Bunch shading during different developmental stages affects the phenolic biosynthesis in berry skins of “Cabernet Sauvignon” grapes. J. Am. Soc. Hortic. Sci 2008, 133, 743–753. [Google Scholar]
- Hardie, W.; Considine, J. Response of grapes to water-deficit stress in particular stages of development. Am. J. Enol. Vitic 1976, 27, 55–61. [Google Scholar]
- Mathews, M.A.; Anderson, M.M.; Schultz, H. Phenologic and growth responses to early and late season water deficits in Cabernet franc. Vitis 1987, 26, 147–160. [Google Scholar]
- McCarthy, M.G. The effect of transient water deficit on berry development of cv. Shiraz (Vitis vinifera L.). Aust. J. Grape Wine Res 1997, 3, 2–8. [Google Scholar]
- Ojeda, H.; Deloire, A.; Carbonneau, A. Influence of water deficits on grape berry growth. Vitis 2001, 40, 141–145. [Google Scholar]
- Chaves, M.M.; Santos, T.P.; Souza, C.R.; Ortuño, M.F.; Rodrigues, M.L.; Lopes, C.M.; Maroco, J.P.; Pereira, J.S. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol 2007, 150, 237–252. [Google Scholar]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; van Leeuwen, C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.). Effects on wine phenolic and aroma components. J. Agric. Food Chem 2006, 54, 5077–5086. [Google Scholar]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A. Effect of maturity and vine water status on grape skin and wine flavonoids. Am. J. Enol. Vitic 2002, 4, 268–274. [Google Scholar]
- Grimplet, J.; Deluc, L.G.; Tillett, R.L.; Wheatley, M.D.; Schlauch, K.A.; Cramer, G.R.; Cushman, J.C. Tissue-specific mRNA expression profiling in grape berry tissues. BMC Genomics 2007, 8, 187. [Google Scholar] [Green Version]
- Vezzulli, S.; Civardi, S.F.F.; Bavaresco, L. Methyl jasmonate treatment as a trigger of resveratrol synthesis in cultivated grapevine. Am. J. Enol. Vitic 2007, 4, 2–5. [Google Scholar]
- Deluc, L.G.; Decendit, A.; Papastamoulis, Y.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water deficit increases stilbene metabolism in Cabernet Sauvignon berries. J. Agric. Food Chem 2011, 59, 289–297. [Google Scholar]
- Deluc, L.G.; Quilici, D.R.; Decendit, A.; Grimplet, J.; Wheatley, M.D.; Schlauch, K.A.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water deficit alters differentially metabolic pathways affecting important flavor and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genomics 2009, 10, 212. [Google Scholar]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Changes in grape seed polyphenols during fruit ripening. Phytochemistry 2000, 55, 77–85. [Google Scholar]
- Geny, L.; Saucier, C.; Bracco, S.; Daviaud, F.; Glories, Y. Composition and cellular localization of tannins in grape seeds during maturation. J. Agric. Food Chem 2003, 51, 8051–8054. [Google Scholar]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res 2004, 10, 100–107. [Google Scholar]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: hints from physiological and molecular data. Ann. Bot 2010, 105, 661–676. [Google Scholar]
- Ollé, D.; Guiraud, J.L.; Souquet, J.M.; Terrier, N.; Ageorges, A.; Cheynier, V.; Verries, C. Effect of pre- and post-veraison water deficit on proanthocyanidin and anthocyanin accumulation during Shiraz berry development. Aust. J. Grape Wine Res 2011, 17, 90–100. [Google Scholar]
- Castellarin, S.D.; Matthews, M.A.; di Gaspero, G.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar]
- Bucchetti, B.; Matthews, M.A.; Falginella, L.; Peterlunger, E.; Castellarin, S.D. Effect of water deficit on Merlot grape tannins and anthocyanins across four seasons. Sci. Hortic 2011, 128, 297–305. [Google Scholar]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ 2007, 30, 1381–1399. [Google Scholar]
- Zarrouk, O.; Francisco, R.; Pinto-Marijuan, M.; Brossa, R.; Santos, R.R.; Pinheiro, C.; Costa, J.M.; Lopes, C.; Chaves, M.M. Impact of irrigation regime on berry development and flavonoids composition in Aragonez (Syn. Tempranillo) grapevine. Agric. Water Manag 2012, 114, 18–29. [Google Scholar]
- Chapman, D.; Roby, G. Sensory attributes of Cabernet Sauvignon wines made from vines with different water status. Aust. J. Grape Wine Res 2005, 11, 339–347. [Google Scholar]
- Pomar, F.; Novo, M.; Masa, A. Varietal differences among the anthocyanin profiles of 50 red table grape cultivars studied by high performance liquid chromatography. J. Chromatogr. A 2005, 1094, 34–41. [Google Scholar]
- Moncada, X.; Muñoz, L.; Castro, M.H.; Hinrichsen, P. Clonal polymorphism in the red wine cultivars “Carmenère” and “Cabernet Sauvignon”. Acta Hort. 2005, 513–520. [Google Scholar]
- Stajner, N.; Jakse, J.; Lavornik, B.; Masuelli, R.W.; Martínez, L.E. Highly variable AFLP and S-SAP markers for the identification of “Malbec” and “Syrah” clones. Vitis 2009, 48, 145–150. [Google Scholar]
- Anderson, M.M.; Smith, R.J.; Williams, M.A.; Wolpert, J.A. Viticultural evaluation of French and California pinot noir clones grown for production of sparkling wine. Am. J. Enol. Vitic 2008, 2, 188–193. [Google Scholar]
- Yang, J.; Martinson, T.E.; Liu, R.H. Phytochemical profiles and antioxidant activities of wine grapes. Food Chem 2009, 116, 332–339. [Google Scholar]
- Katalinić, V.; Možina, S.S.; Skroza, D.; Generalić, I.; Abramovič, H.; Miloš, M.; Ljubenkov, I.; Piskernik, S.; Pezo, I.; Terpinc, P. Polyphenolic profile, antioxidant properties and antimicrobial activity of grape skin extracts of 14 Vitis vinifera varieties grown in Dalmatia (Croatia). Food Chem 2010, 119, 715–723. [Google Scholar]
- Arozarena, I.; Ayestarán, B.; Cantalejo, M.; Navarro, M.; Vera, M.; Abril, I.; Casp, A. Anthocyanin composition of Tempranillo, Garnacha and Cabernet Sauvignon grapes from high- and low-quality vineyards over two years. Eur. Food Res. Technol 2002, 214, 303–309. [Google Scholar]
- Castillo-Muñoz, N. Flavonol profiles of Vitis vinifera red grapes and their single-cultivar wines. J. Agric. Food Chem 2007, 55, 992–1002. [Google Scholar]
- Mattivi, F.; Zulian, C.; Nicolini, G.; Valenti, L. Wine, biodiversity, technology, and antioxidants. Ann. N. Y. Acad. Sci 2002, 56, 37–56. [Google Scholar]
- Lafay, S.; Jan, C.; Nardon, K.; Lemaire, B. Grape extract improves antioxidant status and physical performance in elite male athletes. J. Sport Sci. Med 2009, 8, 468–480. [Google Scholar]
- Siret, R.; Boursiquot, J.M.; Merle, M.H.; Cabanis, J.C.; This, P. Toward the authentication of varietal wines by the analysis of grape (Vitis vinifera L.) residual DNA in must and wine using microsatellite markers. J. Agric. Food Chem 2000, 48, 5035–5040. [Google Scholar]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem 2002, 50, 2891–2899. [Google Scholar]
- Garde-Cerdán, T.; Lorenzo, C.; Lara, J.F.; Prado, F.; Ancín-Azplicueta, C.; Salinas, M. Study of the evolution of nitrogen compounds during grape ripening. Application to differentiate grape varieties and cultivated systems. J. Agric. Food Chem 2009, 57, 2410–2419. [Google Scholar]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Gómez-Cordovés, C.; Peña-Neira, A. Phenolic characterisation of red wines from different grape varieties cultivated in Mendoza province (Argentina). J. Sci. Food Agr 2011, 92, 704–718. [Google Scholar]
- Monagas, M.; Núñez, V. Anthocyanin-derived pigments in Graciano, Tempranillo, and Cabernet Sauvignon wines produced in Spain. Am. J. Enol. Vitic 2003, 3, 163–169. [Google Scholar]
- Guidoni, S.; Allara, P.; Schubert, A. Effect of cluster thinning on berry skin anthocyanin composition of Vitis vinifera cv. Nebbiolo. Am. J. Enol. Vitic 2002, 3, 2001–2003. [Google Scholar]
- Ryan, J.M.; Revilla, E. Anthocyanin composition of Cabernet Sauvignon and Tempranillo grapes at different stages of ripening. J. Agric. Food Chem 2003, 51, 3372–3378. [Google Scholar]
- Ortega-Regules, A.; Romero-Cascales, I.; López-Roca, J.; Ros-García, J.; Gómez-Plaza, E. Anthocyanin fingerprint of grapes: Environmental and genetic variations. J. Sci. Food Agric 2006, 86, 1460–1467. [Google Scholar]
- Hubbard, B.P.; Gomes, A.P.; Dai, H.; Li, J.; Case, A.W.; Considine, T.; Riera, T.V.; Lee, J.E.; Yen, S.; Lamming, D.W. Evidence for a common mechanism of SIRT1 regulation by allosteric activators. Science 2013, 339, 1216–1219. [Google Scholar]
- Delrot, S.; Medrano, H.; Or, E.; Bavaresco, L.; Grando, S. Methodologies and Results in Grapvine Research; Springer: New York, NY, USA, 2010; pp. 5–6. [Google Scholar]
- Cardoso, S.; Lau, W.; Eiras Dias, J.; Fevereiro, P.; Maniatis, N.A. Candidate-gene association study for berry colour and anthocyanin content in Vitis vinifera L. PLoS One 2012, 7, e46021. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Level of synthesis a | Location | Berry phenological scale b | |||||
|---|---|---|---|---|---|---|---|---|
| Skin | Flesh | Seed | Blooming | Green stage | Veraison | Ripening | ||
| Nonflavonoids | ||||||||
| Hydroxycinnamic acids | ++ | +++ | ++ | Hypodermal cells and placental cells of the pulp; primarily in the vacuoles of mesocarp cells. | +++ | +++ | + | + |
| Hydroxybenzoic acids | + | − | ++ | |||||
| Stilbenes | +++ | + | ++ | Berry skin and seeds. | − | + | ++ | +++ |
| Flavonoids | ||||||||
| Flavonols | ++ | − | − | Dermal cell vacuoles of the skin tissue and cell wall of skin and seeds. | ++ | + | +++ | ++ |
| Flavan-3-ols | ++ | + | +++ | Specific vacuoles of hypodermal skin cells and seed coat soft parenquima. | + | ++ | +++ | ++ |
| Anthocyanins | +++ | − * | − | Cell layers below the epidermis; storage confined to the vacuoles and cytoplasmic vesicles named anthocyanoplasts. | − | − | + | +++ |
| Variety | Compound | Effect of water deficit | References |
|---|---|---|---|
| Aragonez (Tempranillo) | Anthocyanins | Decreased concentration. | [128] |
| Barbera | Resveratrol | No effect. | [117] |
| Cabernet Sauvignon | Trans-piceid stilbene precursors | 5-fold increase in concentration. Increased transcript abundance of genes involved in the biosynthesis of stilbene precursors and phenylpropanoid metabolism in general. | [85,111,115,116,118,119,127,129] |
| Flavonols | Increased concentration in the skin and in the wine. No changes in seeds. | ||
| Anthocyanins | Increased of concentration in the skin and in the wine. | ||
| Increased expression of many genes responsible for their biosynthesis. | |||
| Chardonnay | Stilbene precursors | Increased concentration. | [119] |
| Flavonols | Decreased transcript abundance of biosynthetic genes. | ||
| Merlot | Anthocyanins | Increased concentration and biosynthesis; | [125,126] |
| Proanthocyanidins | Increased concentration in berry skin. | ||
| Shiraz | Anthocyanins | Increased concentration. | [124] |
| Varietiy | Nonflavonoids | Flavonoids | References | ||||
|---|---|---|---|---|---|---|---|
| Hydroxycinnamic acids mg·g−1 FW | Hydroxybenzoic acids mg·g−1 FW | Stilbenes mg·g−1 FW | Flavonols mg·g−1 FW | Flavan- 3-ols mg·g−1 FW | Anthocyanins mg·g−1 FW | ||
| Araclinos | 0.742 | 0.034 | 0.001 | 0.042 | 0.386 | 0.655 | [20] |
| Aragonez | 0.658 | [136] | |||||
| Cabernet | 0.103 | 0.011 | 0.003 | 0.039 | 1.830 | 1.830 | [8,136,137] |
| Sauvignon | 0.095 | 1.084 | |||||
| Chardonnay | 0.138 | 0.022 | 0.129 | [20] | |||
| Coudsi | 0.088 | 0.008 | 0.012 | 0.018 | 0.128 | [20] | |
| Garnacha | 0.474 | [137] | |||||
| Greco di Tufo | 0.0002 | [7] | |||||
| Melon | 0.822 | 0.049 | [20] | ||||
| Pinot Noir | 0.152 | 0.018 | 0.003 | 0.035 | 0.161 | 0.800 | [7,20] |
| Rofar Vidor | 0.402 | 0.081 | 0.053 | 0.440 | 0.655 | [20] | |
| Royalty | 0.002 | 0.148 | 0.734 | 5.123 | |||
| Sauvignon Blanc | 0.221 | 0.035 | 0.003 | 0.022 | 0.123 | ||
| Touriga Nacional | 0.754 | 0.024 | 0.006 | 0.176 | 0.33 | 2.632 | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Teixeira, A.; Eiras-Dias, J.; Castellarin, S.D.; Gerós, H. Berry Phenolics of Grapevine under Challenging Environments. Int. J. Mol. Sci. 2013, 14, 18711-18739. https://doi.org/10.3390/ijms140918711
Teixeira A, Eiras-Dias J, Castellarin SD, Gerós H. Berry Phenolics of Grapevine under Challenging Environments. International Journal of Molecular Sciences. 2013; 14(9):18711-18739. https://doi.org/10.3390/ijms140918711
Chicago/Turabian StyleTeixeira, António, José Eiras-Dias, Simone D. Castellarin, and Hernâni Gerós. 2013. "Berry Phenolics of Grapevine under Challenging Environments" International Journal of Molecular Sciences 14, no. 9: 18711-18739. https://doi.org/10.3390/ijms140918711
APA StyleTeixeira, A., Eiras-Dias, J., Castellarin, S. D., & Gerós, H. (2013). Berry Phenolics of Grapevine under Challenging Environments. International Journal of Molecular Sciences, 14(9), 18711-18739. https://doi.org/10.3390/ijms140918711