Molecular Cloning, Characterization and mRNA Expression of a Chitin Synthase 2 Gene from the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae)


Abstract
:1. Introduction
2. Results and Discussion
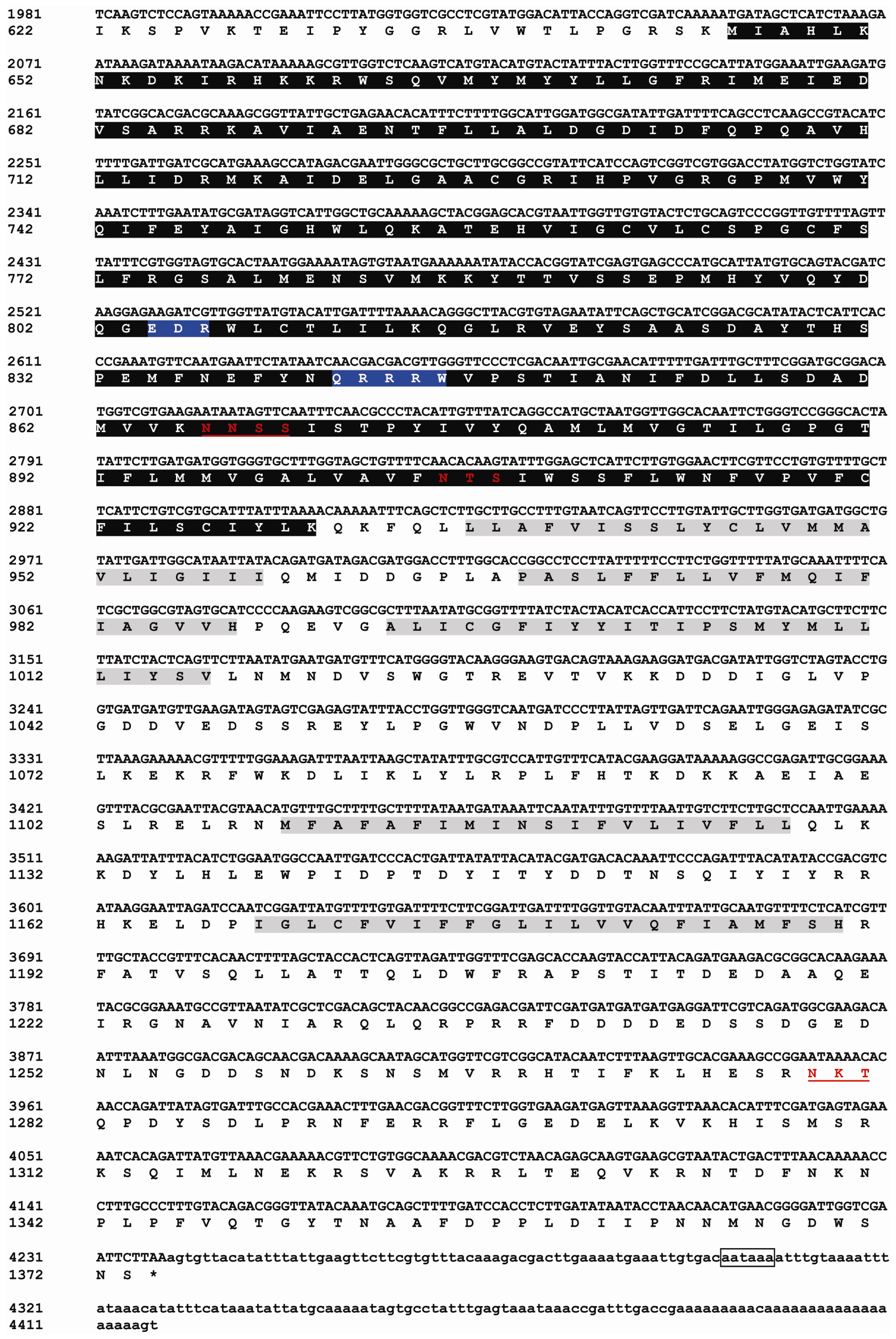
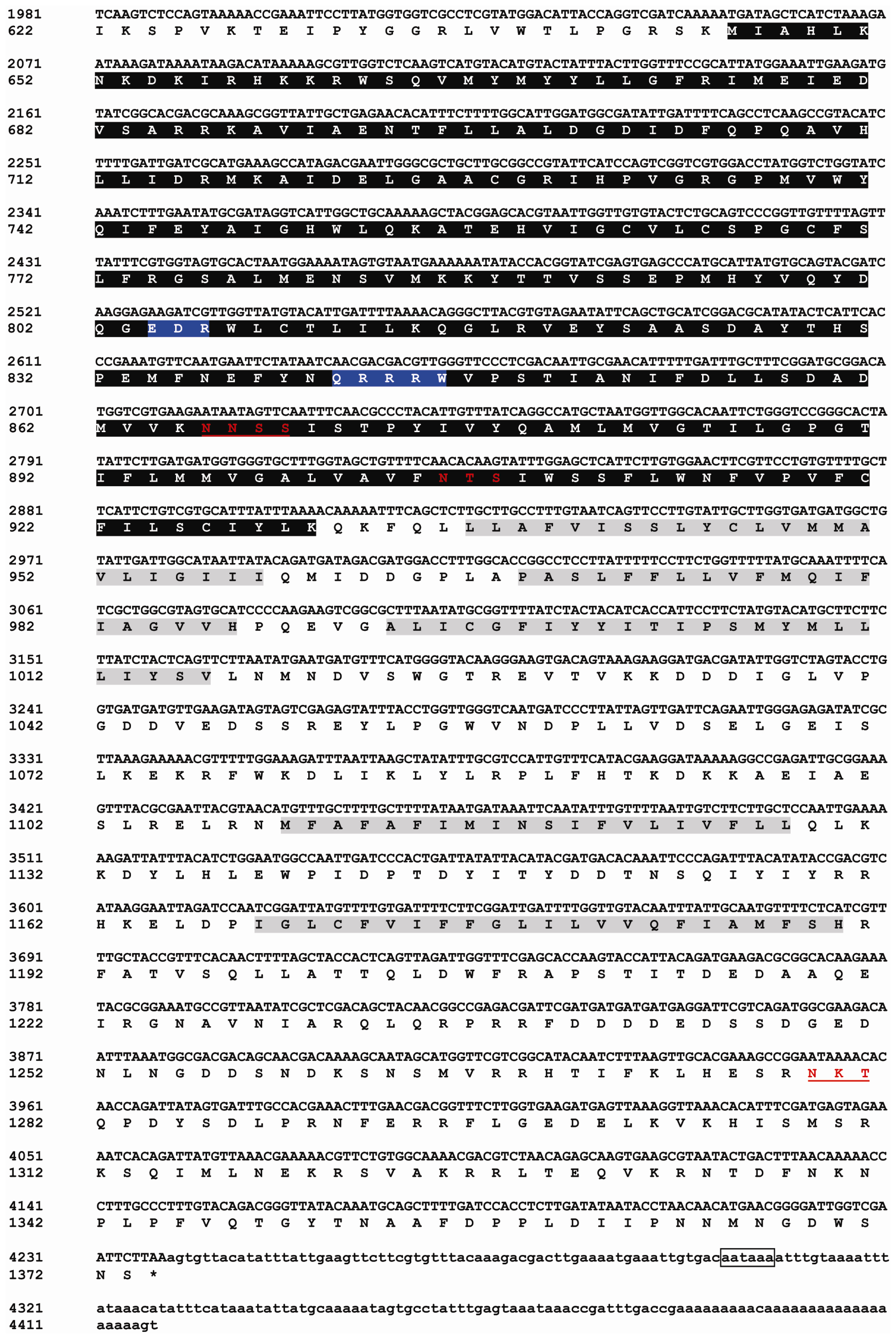
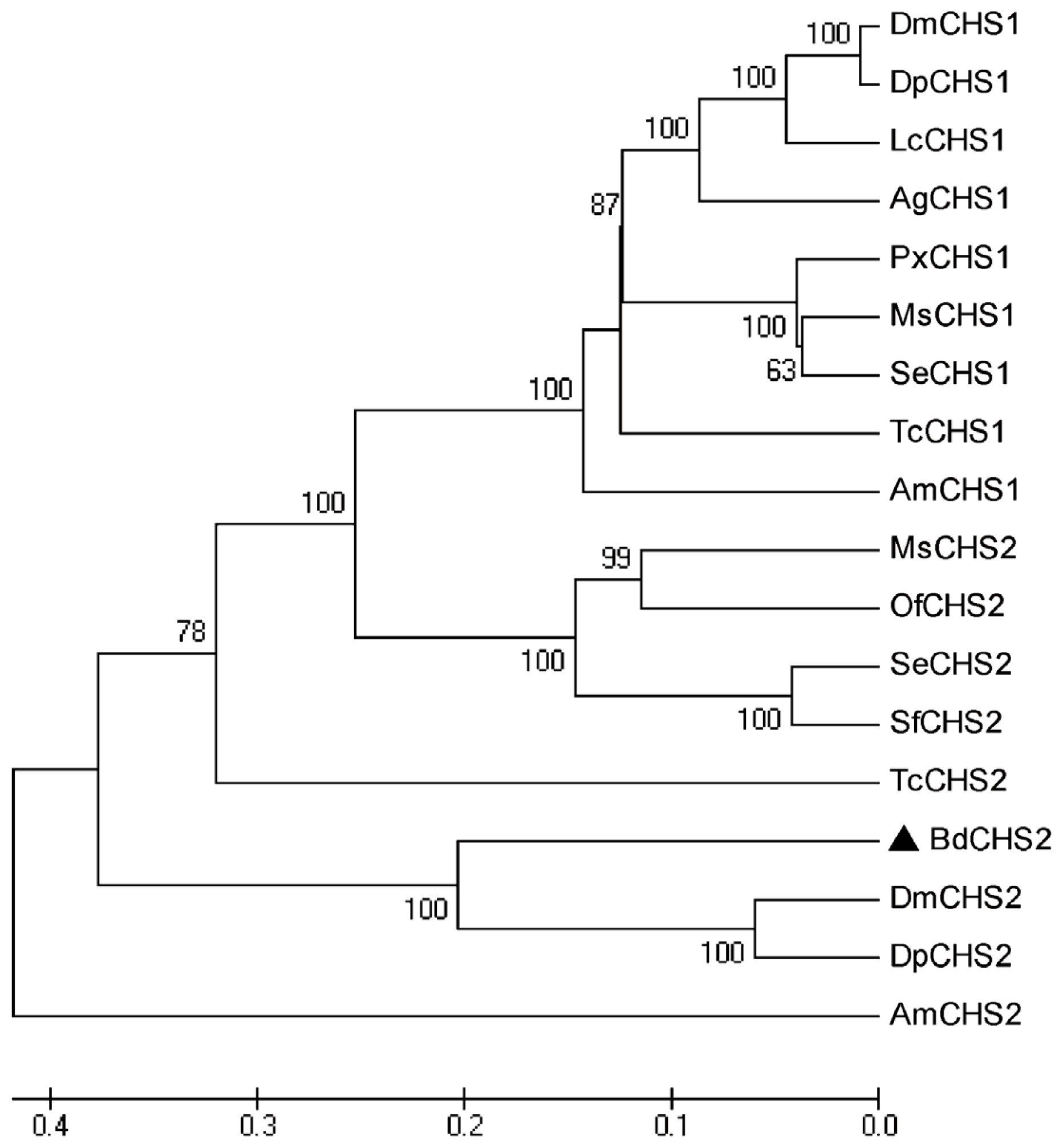
2.1. Identification and Characterization of BdCHS2
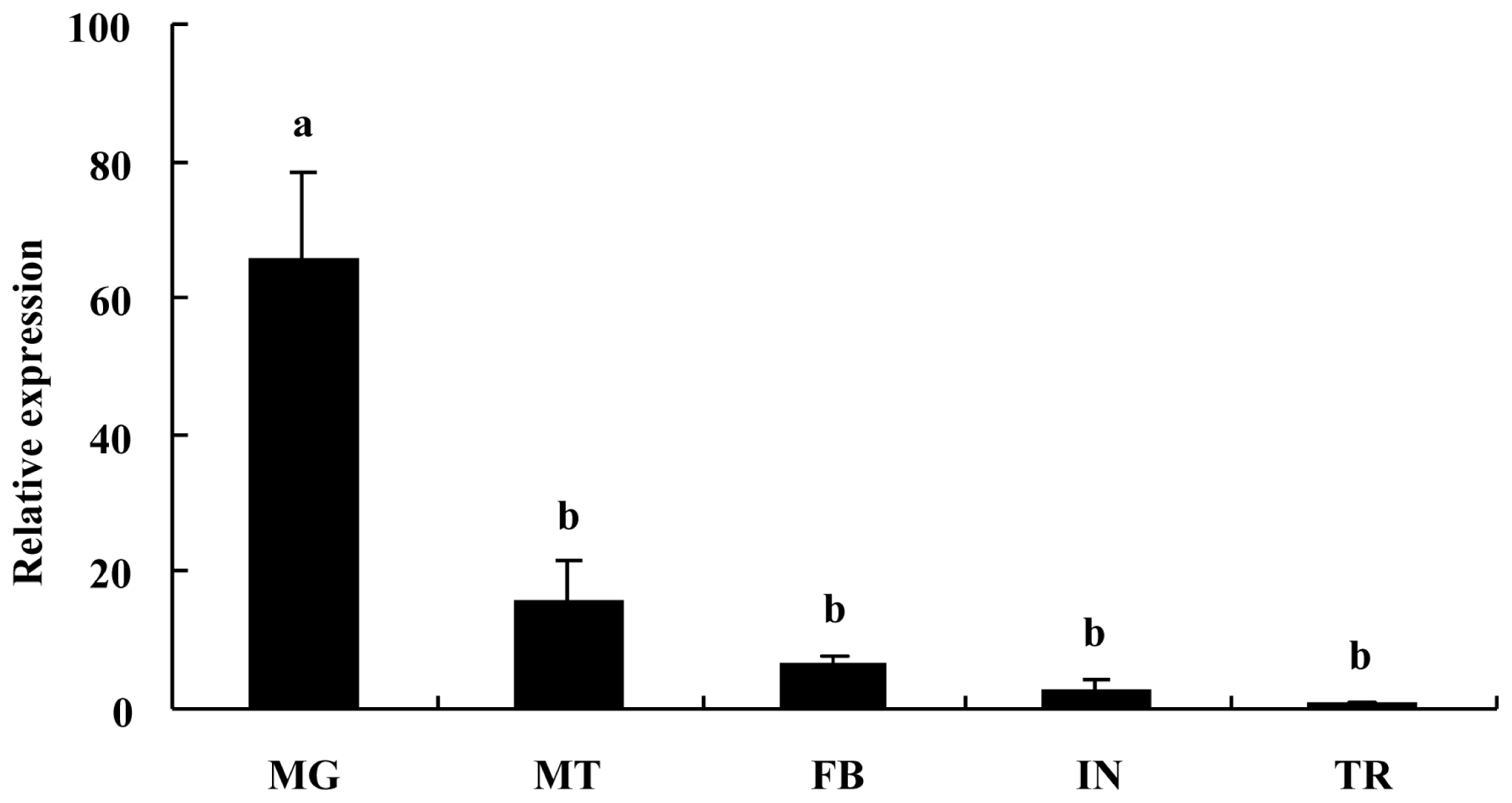
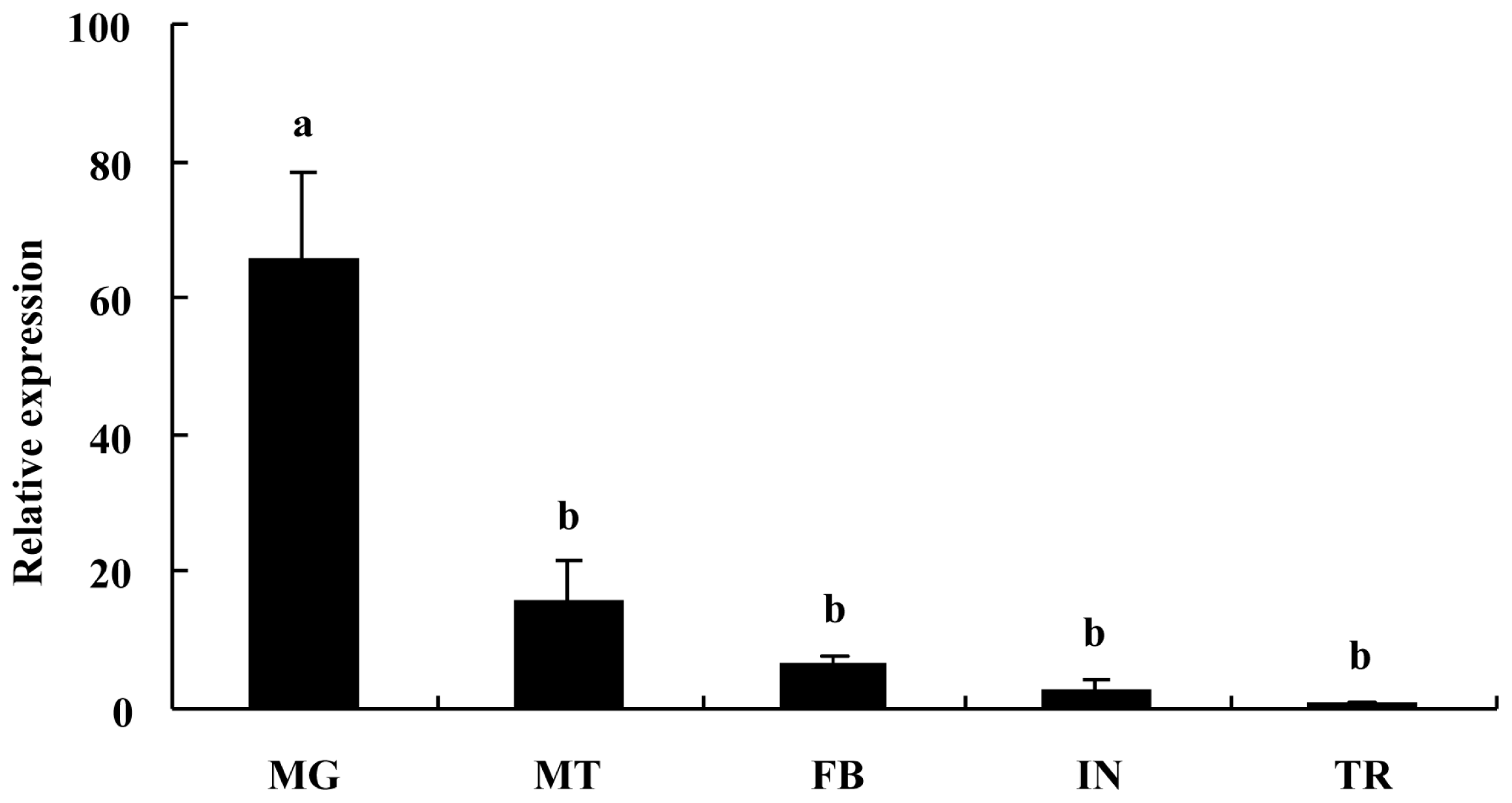
2.2. Tissue-Specific Expression Pattern of BdCHS2
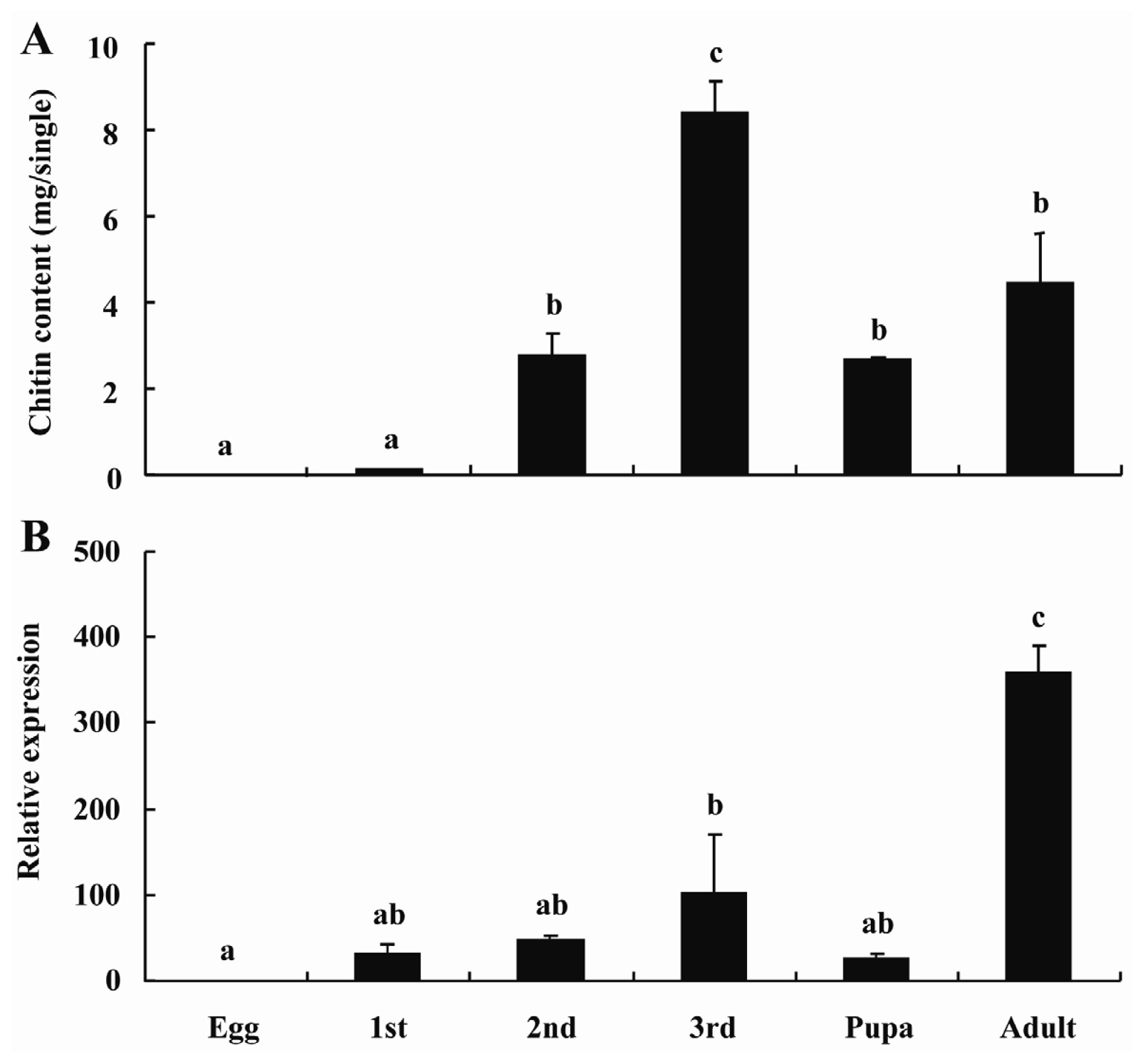
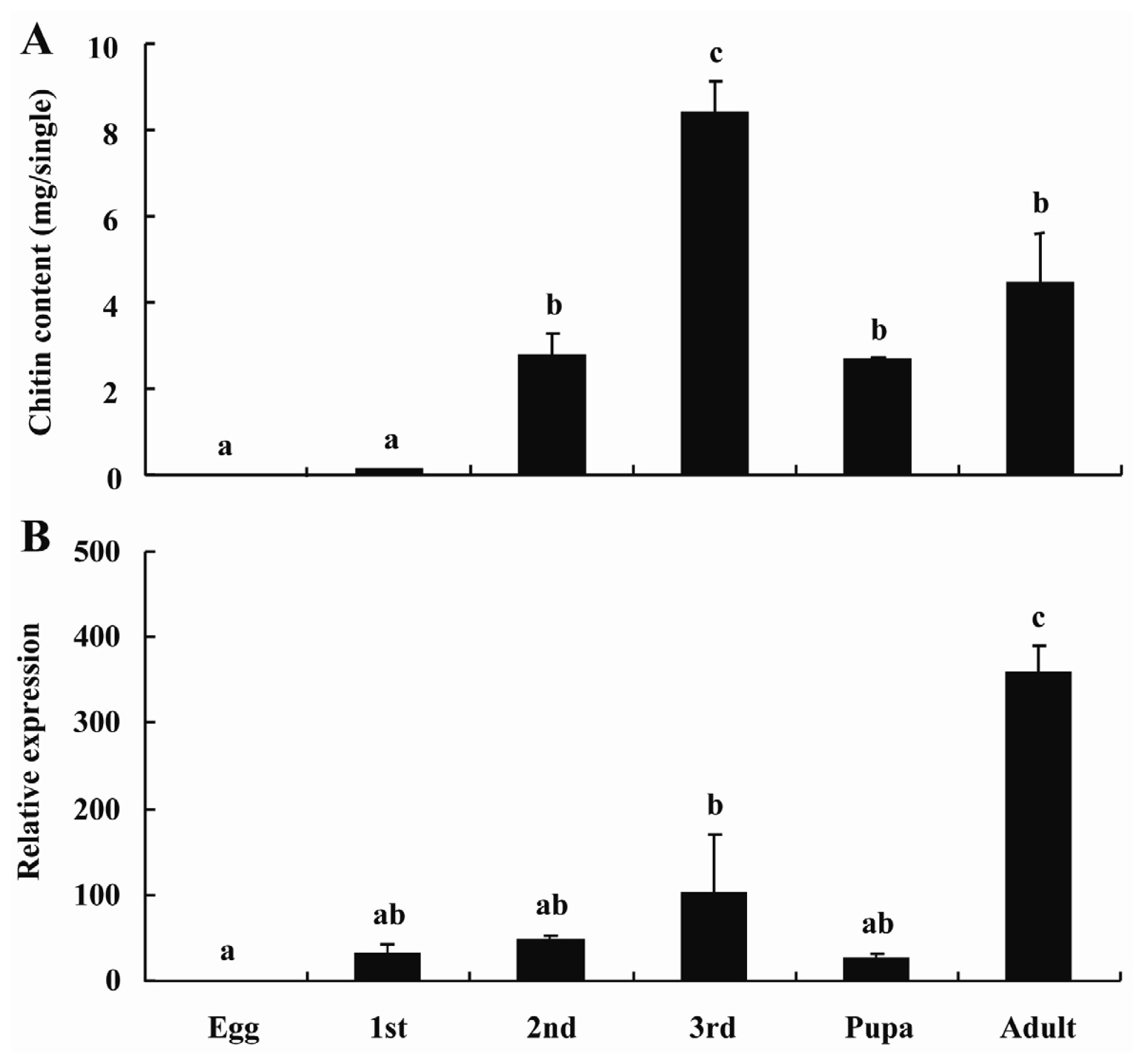
2.3. BdCHS2 Expression and Total Chitin Content during Development
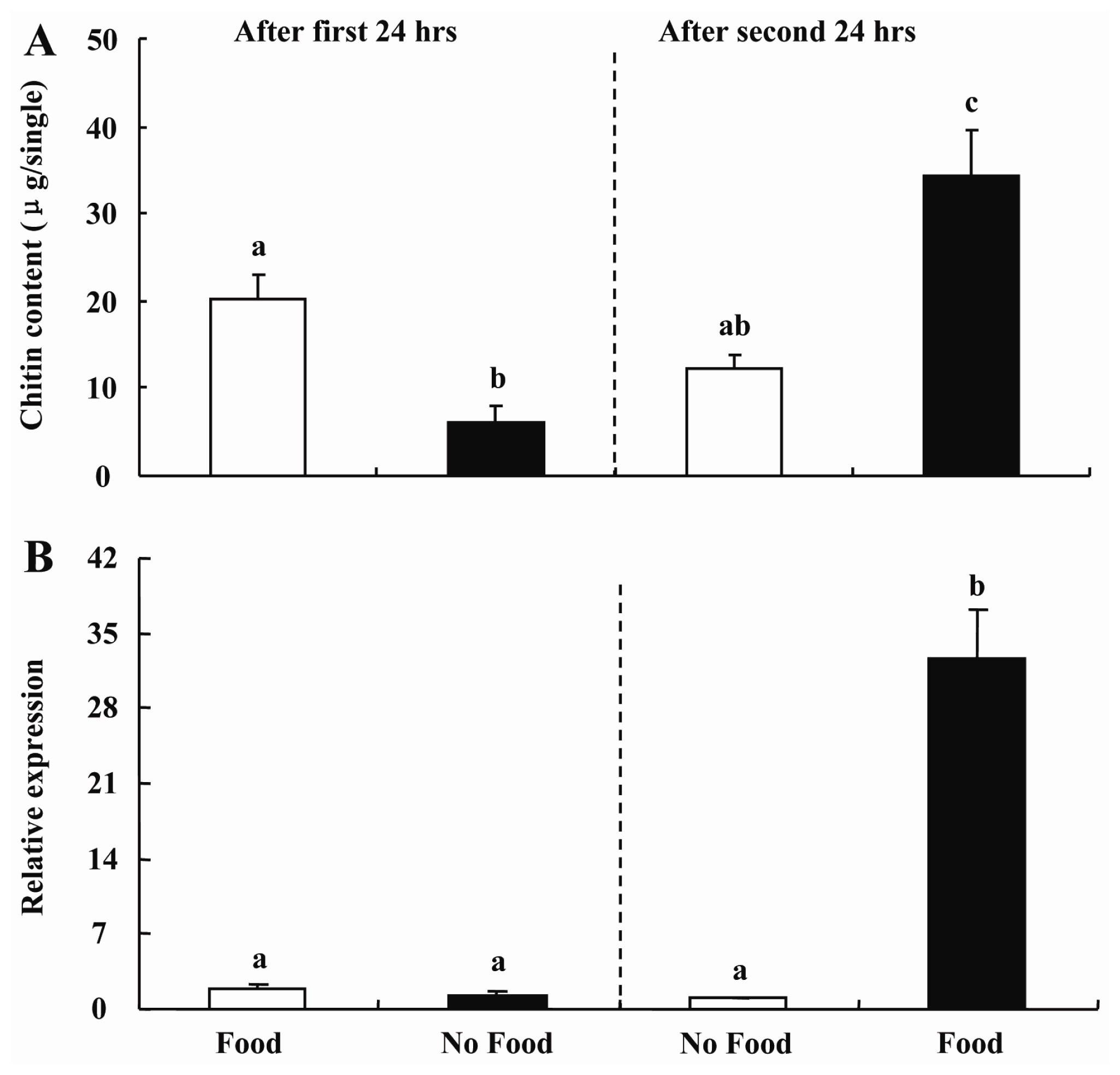
2.4. Feeding-Mediated Changes in Transcript Levels of BdCHS2 and Chitin Content in Midgut
2.5. Discussion
3. Experimental Section
3.1. Test Insect
3.2. cDNA Cloning of BdCHS2 and Sequence Analysis
3.2.1. RNA Extraction and cDNA Synthesis
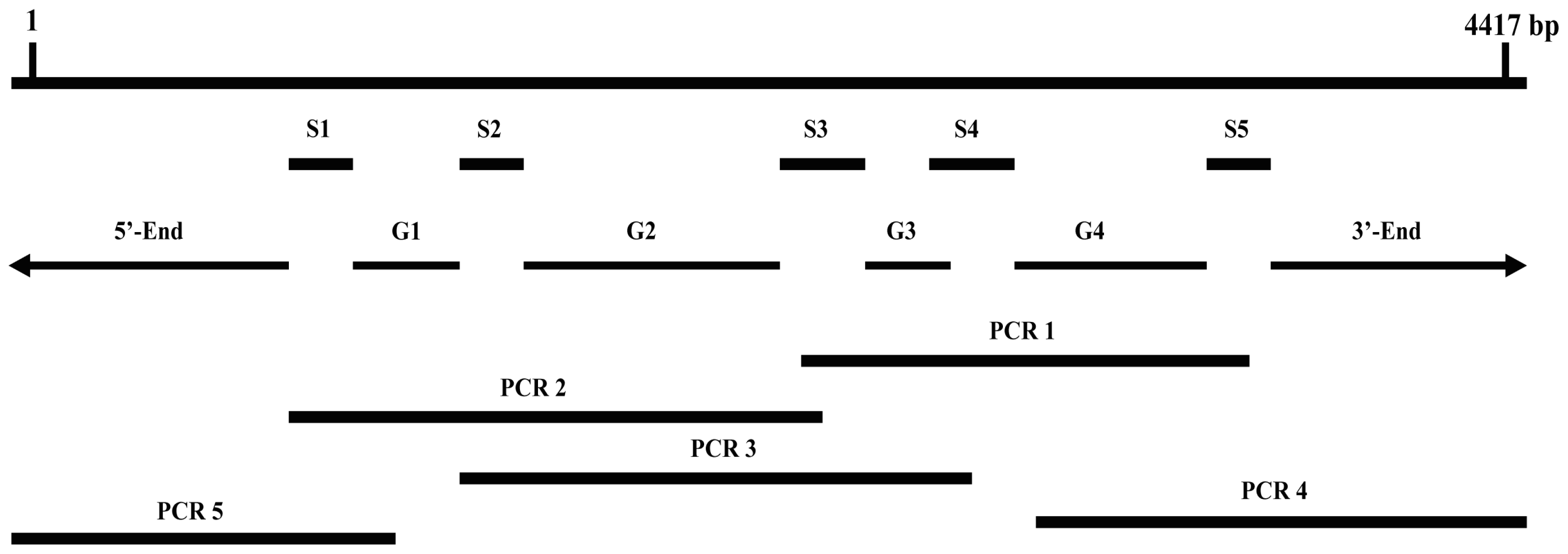
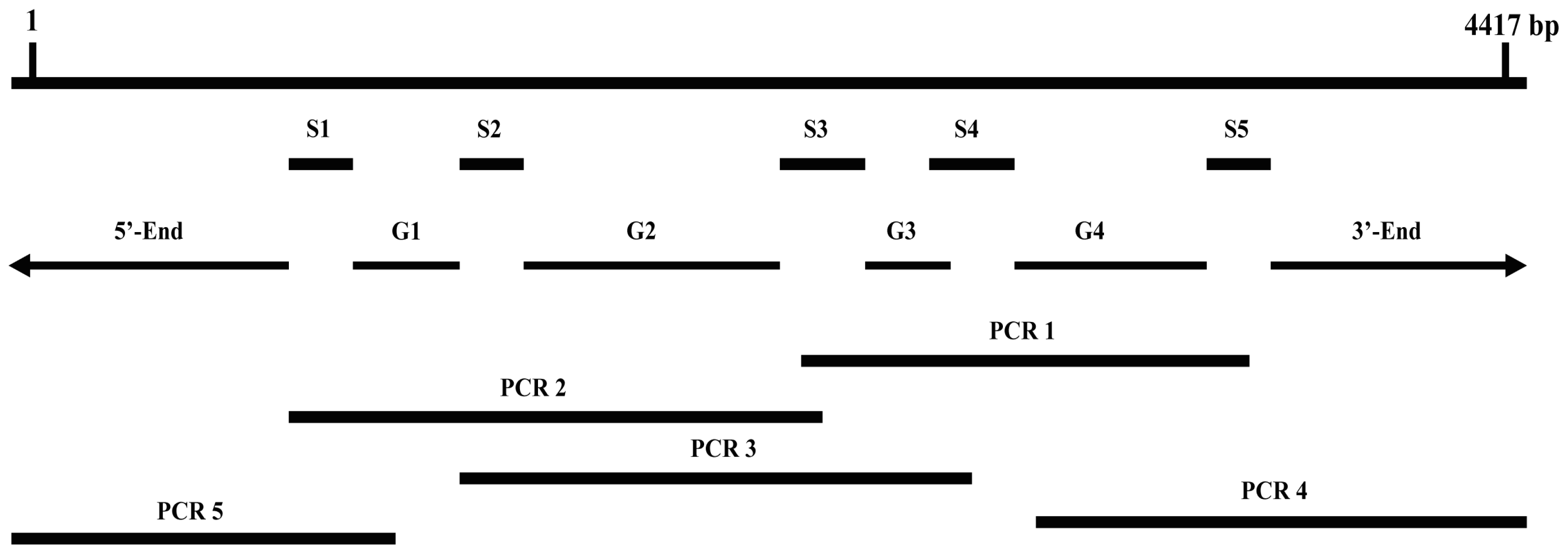
3.2.2. Obtaining Full-Length of BdCHS2 cDNA
3.2.3. Sequence Analysis and Phylogenetic Tree Construction
3.3. Tissue-Specific Expression of BdCHS2 Using Quantitative Real-Time PCR
3.4. Developmental Stages-Specific Expression of BdCHS2 and Total Chitin Content
3.5. Gene Expression Profiles and Chitin Content Assay under Feeding and Starvation Conditions
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Stephens, A.; Kriticos, D.; Leriche, A. The current and future potential geographical distribution of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Bull. Entomol. Res 2007, 97, 369–378. [Google Scholar]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol 2005, 50, 293–319. [Google Scholar]
- Hsu, J.C.; Feng, H.T.; Wu, W.J.; Geib, S.; Mao, C.H.; Vontas, J. Truncated transcripts of nicotinic acetylcholine subunit gene Bdα6 are associated with spinosad resistance in Bactrocera dorsalis. Insect Biochem. Mol. Biol 2012, 42, 805–816. [Google Scholar]
- Jin, T.; Zeng, L.; Lin, Y.; Lu, Y.; Liang, G. Insecticide resistance of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest Manag. Sci 2011, 67, 370–376. [Google Scholar]
- Kramer, K.; Muthukrishnan, S. Chitin Metabolism in Insects. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S., Eds.; Elsevier Press: Oxford, England, UK, 2005; Volume 4, pp. 111–144. [Google Scholar]
- Lehane, M. Peritrophic matrix structure and function. Annu. Rev. Entomol 1997, 42, 525–550. [Google Scholar]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol 2006, 176, 1–15. [Google Scholar]
- Arakane, Y.; Specht, C.A.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Chitin synthases are required for survival, fecundity and egg hatch in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol 2008, 38, 959–962. [Google Scholar]
- Bolognesi, R.; Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Terra, W.R.; Ferreira, C. Sequences of cDNAs and expression of genes encoding chitin synthase and chitinase in the midgut of Spodoptera frugiperda. Insect Biochem. Mol. Biol 2005, 35, 1249–1259. [Google Scholar]
- Kumar, N.S.; Tang, B.; Chen, X.; Tian, H.; Zhang, W. Molecular cloning, expression pattern and comparative analysis of chitin synthase gene B in Spodoptera exigua. J. Comp. Physiol. B 2008, 149, 447–453. [Google Scholar]
- Zhang, X.; Zhang, J.; Park, Y.; Zhu, K.Y. Identification and characterization of two chitin synthase genes in African malaria mosquito, Anopheles gambiae. Insect Biochem. Mol. Biol 2012, 42, 674–682. [Google Scholar]
- Ibrahim, G.H.; Smartt, C.T.; Kiley, L.M.; Christensen, B.M. Cloning and characterization of a chitin synthase cDNA from the mosquito Aedes aegypti. Insect Biochem. Mol. Biol 2000, 30, 1213–1222. [Google Scholar]
- Gagou, M.E.; Kapsetaki, M.; Turberg, A.; Kafetzopoulos, D. Stage-specific expression of the chitin synthase DmeChSA and DmeChSB genes during the onset of Drosophila metamorphosis. Insect Biochem. Mol. Biol 2002, 32, 141–146. [Google Scholar]
- Arakane, Y.; Hogenkamp, D.G.; Zhu, Y.C.; Kramer, K.J.; Specht, C.A.; Beeman, R.W.; Kanost, M.R.; Muthukrishnan, S. Characterization of two chitin synthase genes of the red flour beetle, Tribolium castaneum, and alternate exon usage in one of the genes during development. Insect Biochem. Mol. Biol 2004, 34, 291–304. [Google Scholar]
- Hogenkamp, D.G.; Arakane, Y.; Zimoch, L.; Merzendorfer, H.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Specht, C.A.; Muthukrishnan, S. Chitin synthase genes in Manduca sexta: Characterization of a gut-specific transcript and differential tissue expression of alternately spliced mRNAs during development. Insect Biochem. Mol. Biol 2005, 35, 529–540. [Google Scholar]
- Qu, M.; Liu, T.; Yang, J.; Yang, Q. The gene, expression pattern and subcellular localization of chitin synthase B from the insect Ostrinia furnacalis. Biochem. Biophys. Res. Commun 2011, 404, 302–307. [Google Scholar]
- Liu, X.; Zhang, H.; Li, S.; Zhu, K.Y.; Ma, E.; Zhang, J. Characterization of a midgut-specific chitin synthase gene (LmCHS2) responsible for biosynthesis of chitin of peritrophic matrix in Locusta migratoria. Insect Biochem. Mol. Biol 2012, 42, 902–910. [Google Scholar]
- Khajuria, C.; Buschman, L.L.; Chen, M.S.; Muthukrishnan, S.; Zhu, K.Y. A gut-specific chitinase gene essential for regulation of chitin content of peritrophic matrix and growth of Ostrinia nubilalis larvae. Insect Biochem. Mol. Biol 2010, 40, 621–629. [Google Scholar]
- Kato, N.; Mueller, C.R.; Fuchs, J.F.; Wessely, V.; Lan, Q.; Christensen, B.M. Regulatory mechanisms of chitin biosynthesis and roles of chitin in peritrophic matrix formation in the midgut of adult Aedes aegypti. Insect Biochem. Mol. Biol 2006, 36, 1–9. [Google Scholar]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Biochem. Mol. Biol 2005, 14, 453–463. [Google Scholar]
- Tellam, R.L.; Vuocolo, T.; Johnson, S.E.; Jarmey, J.; Pearson, R.D. Insect chitin synthase-cDNA sequence, gene organization and expression. Eur. J. Biochem 2000, 267, 6025–6042. [Google Scholar]
- Yang, W.J.; Xu, K.K.; Cong, L.; Wang, J.J. Identification, mRNA expression, and functional analysis of chitin synthase 1 gene and its two alternative splicing variants in oriental fruit fly, Bactrocera dorsalis. Int. J. Biol. Sci 2013, 9, 331–342. [Google Scholar]
- Wang, Y.; Fan, H.W.; Huang, H.J.; Xue, J.; Wu, W.J.; Bao, Y.Y.; Xu, H.J.; Zhu, Z.R.; Cheng, J.A.; Zhang, C.X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvatalugens and Laodelphaxstriatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol 2012, 42, 637–646. [Google Scholar]
- Qu, M.; Yang, Q. A novel alternative splicing site of class A chitin synthase from the insect Ostrinia furnacalis-Gene organization, expression pattern and physiological significance. Insect Biochem. Mol. Biol 2011, 41, 923–931. [Google Scholar]
- Ampasala, D.R.; Zheng, S.; Zhang, D.; Ladd, T.; Doucet, D.; Krell, P.J.; Retnakaran, A.; Feng, Q. An epidermis-specific chitin synthase cDNA in Choristoneura fumiferana: Cloning, characterization, developmental and hormonal-regulated expression. Arch. Insect Biochem. Physiol 2011, 76, 83–96. [Google Scholar]
- Zhang, J.; Liu, X.; Li, D.; Sun, Y.; Guo, Y.; Ma, E.; Zhu, K.Y. Silencing of two alternative splicing-derived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locust, Locusta migratoria manilensis (Meyen). Insect Biochem. Mol. Biol 2010, 40, 824–833. [Google Scholar]
- Terra, W.R. The origin and functions of the insect peritrophic membrane and peritrophic gel. Arch. Insect Biochem. Physiol 2001, 47, 47–61. [Google Scholar]
- Terra, W.; Ferreira, C. Biochemistry of digestion. Compr. Mol. Insect Sci 2005, 4, 171–224. [Google Scholar]
- Shen, Z.; Jacobs-Lorena, M. Characterization of a novel gut-specific chitinase gene from the human malaria vector Anopheles gambiae. J. Biol. Chem 1997, 272, 28895–28900. [Google Scholar]
- Ramalho-Ortigao, J.; Traub-Csekö, Y. Molecular characterization of Llchit1, a midgut chitinase cDNA from the leishmaniasis vector Lutzomyia longipalpis. Insect Biochem. Mol. Biol 2003, 33, 279–287. [Google Scholar]
- Kikuchi, E.; Takagi, S.; Shikano, S. Changes in carbon and nitrogen stable isotopes of chironomid larvae during growth, starvation and metamorphosis. Rapid Commun. Mass Spectrom 2007, 21, 997–1002. [Google Scholar]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol 2007, 25, 1322–1326. [Google Scholar]
- Cong, L.; Yang, W.J.; Shen, G.M.; Dou, W.; Wang, J.J. Molecular characterization of the cDNA encoding ecdysone receptor isoform B1 and its expression in the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol 2012, 95, 650–658. [Google Scholar]
- Shen, G.M.; Dou, W.; Niu, J.Z.; Jiang, H.B.; Yang, W.J.; Jia, F.X.; Hu, F.; Cong, L.; Wang, J.J. Transcriptome analysis of the oriental fruit fly (Bactrocera dorsalis). PLoS One 2011, 6, e29127. [Google Scholar]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 15 September 2012).
- Compute pI/Mw Tool. Available online: http://cn.expasy.org/tools/pi_tool.html (accessed on 15 September 2012).
- NetNGlyc 1.0 Server. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/ (accessed on 15 September 2012).
- PSORT WWW Server. http://psort.nibb.ac.jp/ (accessed on 15 September 2012).
- SignalP 3.0 Server. http://www.cbs.dtu.dk/services/SignalP-3.0/ (accessed on 15 September 2012).
- TMHMM Server 2.0. http://www.cbs.dtu.dk/services/TMHMM-2.0/ (accessed on 15 September 2012).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar]
- Lehmann, P.F.; White, L.O. Chitin assay used to demonstrate renal localization and cortisone-enhanced growth of Aspergillus fumigatus Mycelium in mice. Am. Soc. Microbiol 1975, 12, 987–992. [Google Scholar]
- Zhang, J.; Zhu, K.Y. Characterization of a chitin synthase cDNA and its increased mRNA level associated with decreased chitin synthesis in Anopheles quadrimaculatus exposed to diflubenzuron. Insect Biochem. Mol. Biol 2006, 36, 712–725. [Google Scholar]
- Ride, J.; Drysdale, R. A rapid method for the chemical estimation of filamentous fungi in plant tissue. Physiol. Plant Pathol 1972, 2, 7–15. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cDNA fragment | Length (bp) | Position in the coding area of BdCHS2 (bp) |
|---|---|---|
| S1 | 284 | 770–1,053 |
| S2 | 183 | 1,353–1,535 |
| S3 | 319 | 2,177–2,495 |
| S4 | 252 | 2,643–2,894 |
| S5 | 243 | 3,718–3,960 |
| Application of primers | cDNA fragment | Primer name | Primer sequence(5′-3′) | cDNA position in the coding area (bp) |
|---|---|---|---|---|
| cDNA cloning | PCR 1 | CHS2-1 | TACTCTGCAGTCCCGGTTGTT | 2404–2424 |
| CHS2-2 | CTTGTGCCGCGTCTTCATCTG | 3757–3777 | ||
| PCR 2 | CHS2-3 | TAGTCGTTCTAGATATCAGAC | 926–946 | |
| CHS2-4 | AGCAGCGCCCAATTCGTCTATG | 2273–2294 | ||
| PCR 3 | CHS2-5 | GGATAACTCGACATATTTGGC | 1465–1485 | |
| CHS2-6 | TGTAGGGCGTTGAAATTGAACTA | 2717–2739 | ||
| PCR 4 (3′-RACE) | CHS2-7 | GGAAGTGACAGTAAAGAAGGATG | 3197–3219 | |
| CHS2-8 | TAAATGGCGACGACAGCAACG | 3874–3894 | ||
| PCR 5 (5′-RACE) | CHS2-9 | CCACATAGCAACGGCAACAGAAGC | 1290–1313 | |
| CHS2-10 | TAATGGGAGGGCGATTATTTGTAAC | 821–845 | ||
| UPM | CTAATACGACTCACTATAGGGC | – | ||
| NUP | AAGCAGTGGTATCAACGCAGAGT | – | ||
| qPCR analysis | CHS2 | CHS2-Q-F | ATTTTCAGCCTCAAGCCGTA | 2227–2246 |
| CHS2-Q-R | CGGGACTGCAGAGTACACAA | 2399–2418 | ||
| A-tubulin | α-tub-F | CGCATTCATGGTTGATAACG | – | |
| α-tub-R | GGGCACCAAGTTAGTCTGGA | – | ||
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, L.; Yang, W.-J.; Cong, L.; Xu, K.-K.; Wang, J.-J. Molecular Cloning, Characterization and mRNA Expression of a Chitin Synthase 2 Gene from the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae). Int. J. Mol. Sci. 2013, 14, 17055-17072. https://doi.org/10.3390/ijms140817055
Chen L, Yang W-J, Cong L, Xu K-K, Wang J-J. Molecular Cloning, Characterization and mRNA Expression of a Chitin Synthase 2 Gene from the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae). International Journal of Molecular Sciences. 2013; 14(8):17055-17072. https://doi.org/10.3390/ijms140817055
Chicago/Turabian StyleChen, Li, Wen-Jia Yang, Lin Cong, Kang-Kang Xu, and Jin-Jun Wang. 2013. "Molecular Cloning, Characterization and mRNA Expression of a Chitin Synthase 2 Gene from the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae)" International Journal of Molecular Sciences 14, no. 8: 17055-17072. https://doi.org/10.3390/ijms140817055
APA StyleChen, L., Yang, W.-J., Cong, L., Xu, K.-K., & Wang, J.-J. (2013). Molecular Cloning, Characterization and mRNA Expression of a Chitin Synthase 2 Gene from the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae). International Journal of Molecular Sciences, 14(8), 17055-17072. https://doi.org/10.3390/ijms140817055