Cascading cis-Cleavage on Transcript from trans-Acting siRNA-Producing Locus 3

Abstract
:1. Introduction
2. Results and Discussion
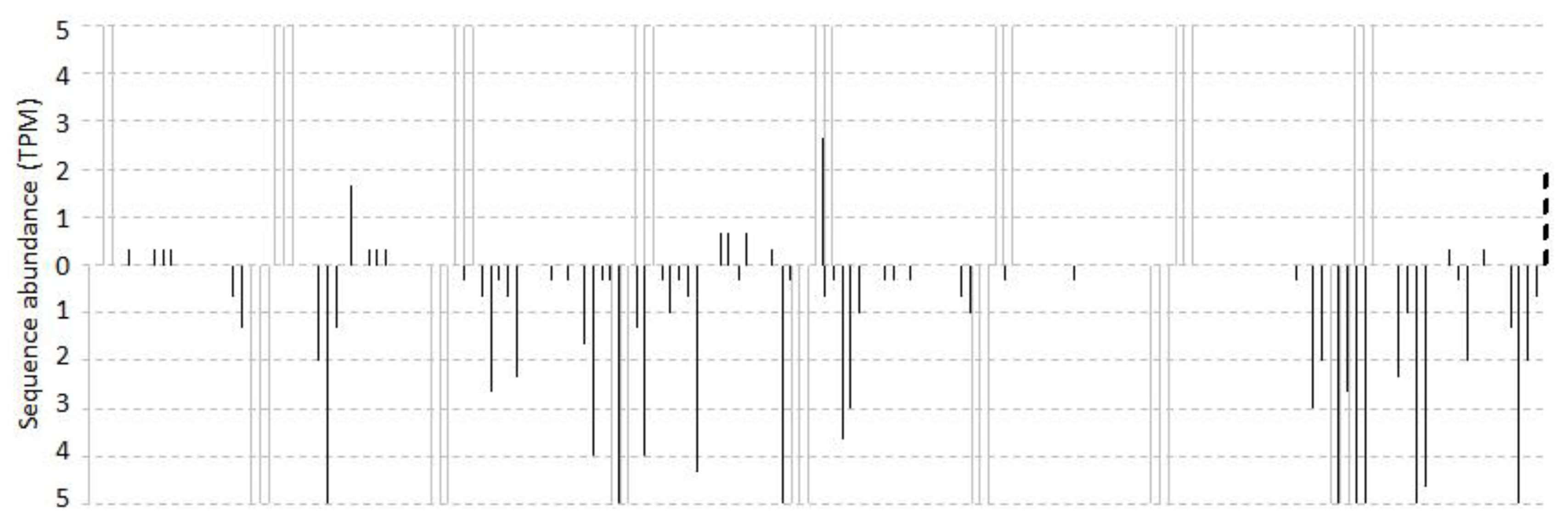
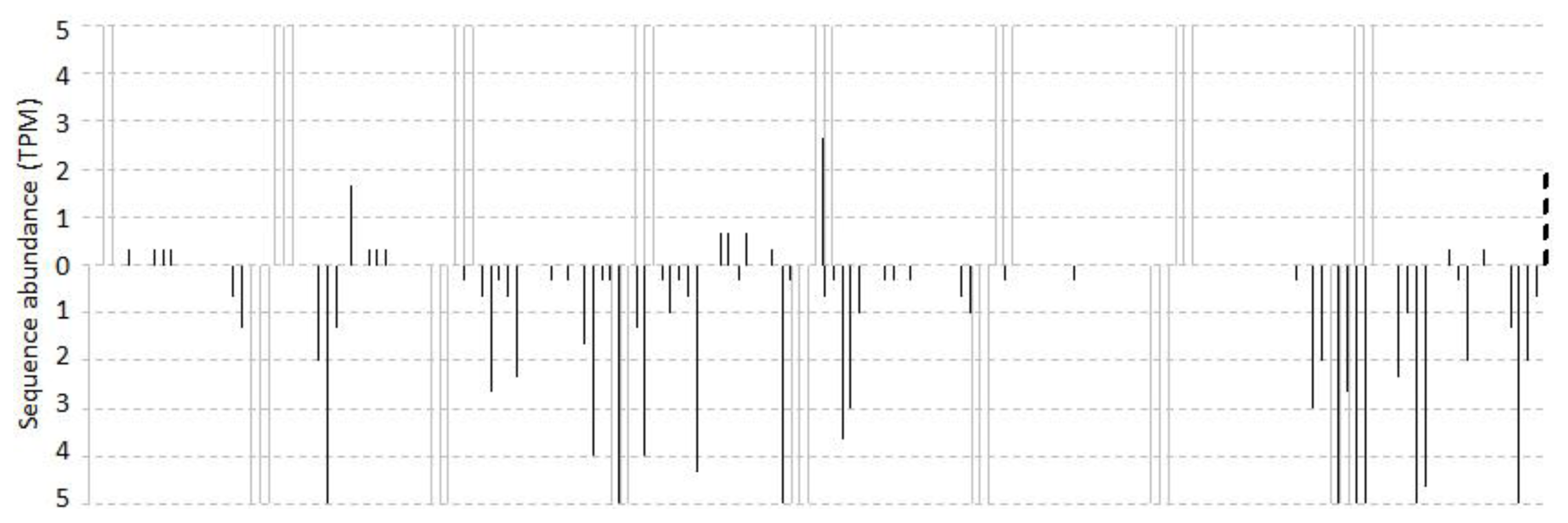
2.1. Overview of Small RNA Distribution on TAS3 from Vitis vinifera
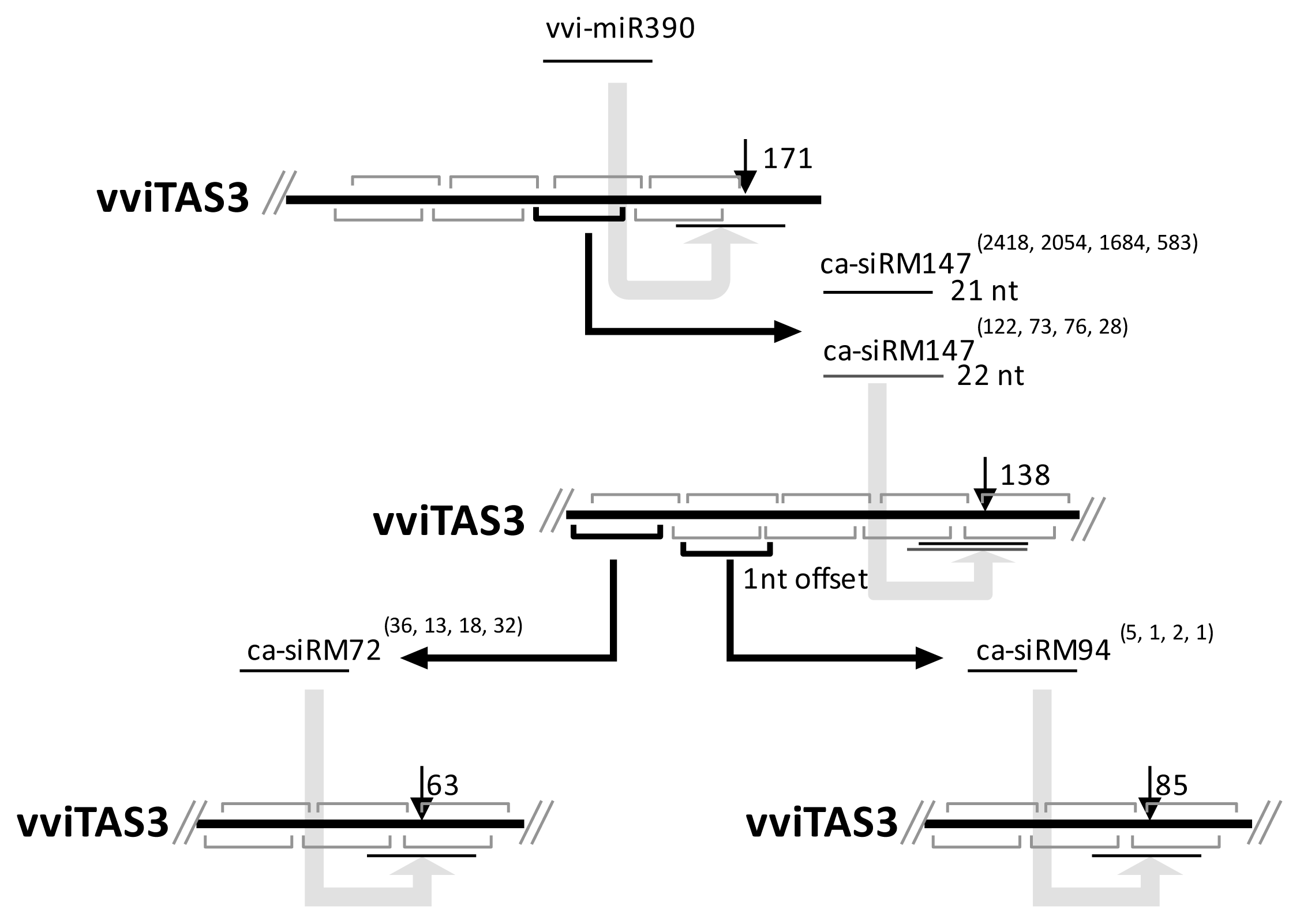
2.2. Computational Prediction and Validation of cis-Cleavages
2.3. Cascading cis-Cleavages
2.4. The Accumulated Levels of ca-siRNAs
2.5. cis-Cleavages Produced sRNAs in Increments of Approximately 21 nt
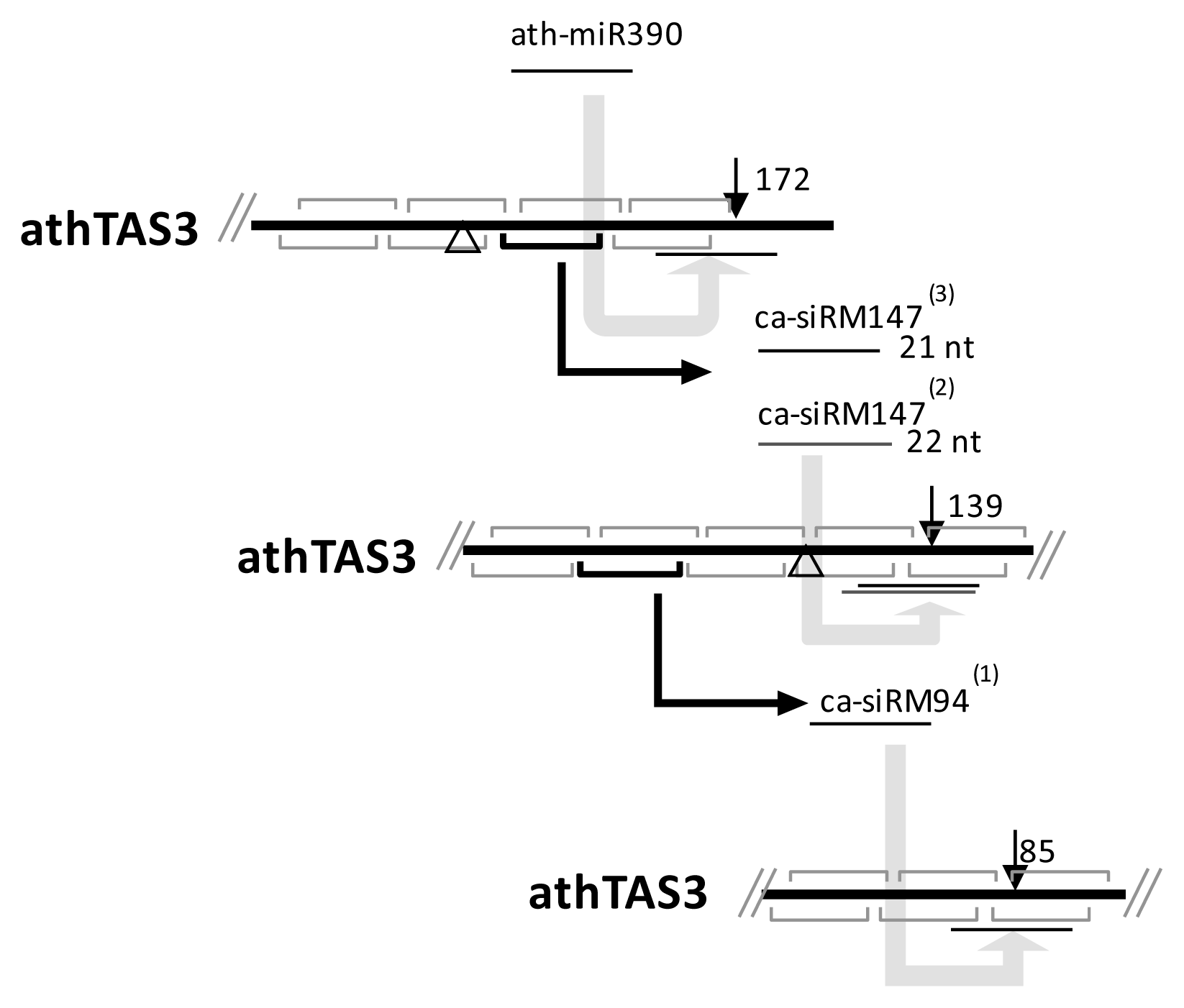
2.6. The Conservation of ca-siRNAs and Cascading cis-Cleavages
3. Experimental Section
3.1. Sources of sRNA Libraries
3.2. Evaluation of Phasing Patterns Set by cis-Cleavage
3.3. Expressional Conservation of ca-siRNAs
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Axtell, M.J. Classification and comparison of small RNAs from plants. Annu. Rev. Plant Biol 2013, 64, 137–159. [Google Scholar]
- Chen, X. Small RNAs and their roles in plant development. Annu. Rev. Cell Dev. Biol 2009, 25, 21–44. [Google Scholar]
- Peragine, A.; Yoshikawa, M.; Wu, G.; Albrecht, H.L.; Poethig, R.S. SGS3 and SGS2/SDE1/RDR6 are required for juvenile development and the production of trans-acting siRNAs in Arabidopsis. Genes Dev 2004, 18, 2368–2379. [Google Scholar]
- Vazquez, F.; Vaucheret, H.; Rajagopalan, R.; Lepers, C.; Gasciolli, V.; Mallory, A.C.; Hilbert, J.L.; Bartel, D.P.; Crete, P. Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol. Cell 2004, 16, 69–79. [Google Scholar]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar]
- Gasciolli, V.; Mallory, A.C.; Bartel, D.P.; Vaucheret, H. Partially redundant functions of Arabidopsis DICER-like enzymes and a role for DCL4 in producing trans-acting siRNAs. Curr. Biol 2005, 15, 1494–1500. [Google Scholar]
- Yoshikawa, M.; Peragine, A.; Park, M.Y.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes Dev 2005, 19, 2164–2175. [Google Scholar]
- Axtell, M.J.; Jan, C.; Rajagopalan, R.; Bartel, D.P. A two-hit trigger for siRNA biogenesis in plants. Cell 2006, 127, 565–577. [Google Scholar]
- Rajagopalan, R.; Vaucheret, H.; Trejo, J.; Bartel, D.P. A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev 2006, 20, 3407–3425. [Google Scholar]
- Li, F.; Orban, R.; Baker, B. SoMART: A web server for plant miRNA, tasiRNA and target gene analysis. Plant J 2012, 70, 891–901. [Google Scholar]
- Arif, M.A.; Fattash, I.; Ma, Z.; Cho, S.H.; Beike, A.K.; Reski, R.; Axtell, M.J.; Frank, W. DICER-LIKE3 activity in Physcomitrella patens DICER-LIKE4 mutants causes severe developmental dysfunction and sterility. Mol. Plant 2012, 5, 1281–1294. [Google Scholar]
- Zhang, C.; Li, G.; Wang, J.; Fang, J. Identification of trans-acting siRNAs and their regulatory cascades in grapevine. Bioinformatics 2012, 28, 2561–2568. [Google Scholar]
- Chen, H.M.; Li, Y.H.; Wu, S.H. Bioinformatic prediction and experimental validation of a microRNA-directed tandem trans-acting siRNA cascade in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 3318–3323. [Google Scholar]
- Rajeswaran, R.; Aregger, M.; Zvereva, A.S.; Borah, B.K.; Gubaeva, E.G.; Pooggin, M.M. Sequencing of RDR6-dependent double-stranded RNAs reveals novel features of plant siRNA biogenesis. Nucleic Acids Res 2012, 40, 6241–6254. [Google Scholar]
- Howell, M.D.; Fahlgren, N.; Chapman, E.J.; Cumbie, J.S.; Sullivan, C.M.; Givan, S.A.; Kasschau, K.D.; Carrington, J.C. Genome-wide analysis of the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 pathway in Arabidopsis reveals dependency on miRNA- and tasiRNA-directed targeting. Plant Cell 2007, 19, 926–942. [Google Scholar]
- German, M.A.; Pillay, M.; Jeong, D.H.; Hetawal, A.; Luo, S.; Janardhanan, P.; Kannan, V.; Rymarquis, L.A.; Nobuta, K.; German, R.; et al. Global identification of microRNA-target RNA pairs by parallel analysis of RNA ends. Nat. Biotechnol 2008, 26, 941–946. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- Pantaleo, V.; Szittya, G.; Moxon, S.; Miozzi, L.; Moulton, V.; Dalmay, T.; Burgyan, J. Identification of grapevine microRNAs and their targets using high-throughput sequencing and degradome analysis. Plant J. Cell Mol. Biol 2010, 62, 960–976. [Google Scholar]
- Dai, X.; Zhao, P.X. pssRNAMiner: A plant short small RNA regulatory cascade analysis server. Nucleic Acids Res 2008, 36, W114–W118. [Google Scholar]
- Zhang, C.; Wang, J.; Hua, X.; Fang, J.; Zhu, H.; Gao, X. A mutation degree model for the identification of transcriptional regulatory elements. BMC Bioinforma. 2011, 12. [Google Scholar] [CrossRef]
- Nakano, M.; Nobuta, K.; Vemaraju, K.; Tej, S.S.; Skogen, J.W.; Meyers, B.C. Plant MPSS databases: Signature-based transcriptional resources for analyses of mRNA and small RNA. Nucleic Acids Res 2006, 34, D731–D735. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cis-cleaved position | ca-siRNA | |
|---|---|---|
| Name | Sequence | |
| 63 | ca-siRM72 | UGGAAAACGAAGAAGAAAUGC |
| 85 | ca-siRM94 | UGGUAUAGAGUUCAUGACAAG |
| 138 | ca-siRM147 | UGAGUUGGGCGGAAACGGGGA UGAGUUGGGCGGAAACGGGGAA |
| cis-cleaved position | Upstream of cis-cleaved position | Downstream of cis-cleaved position | ||||
|---|---|---|---|---|---|---|
| Number of phased sRNAs | Number of non-phased sRNAs | p value * | Number of phased sRNAs | Number of non-phased sRNAs | p-value | |
| 63 | 2 | 12 | 1.11 × 10–1 | 3 | 47 | 4.92 × 10–1 |
| 85 | 5 | 27 | 7.54 × 10–3 | 4 | 30 | 1.00 × 10–1 |
| 138 | 8 | 41 | 4.71 × 10–4 | 3 | 16 | 7.43 × 10–2 |
| ca-siRNAs | Average of normalized abundance (TPM) | ||||
|---|---|---|---|---|---|
| Name | Length | Grapevine | Apple | Peach | Arabidopsis |
| ca-siRM72 | 21 nt | 6 | 2 | 1 | 1 |
| ca-siRM94 | 21 nt | 1 | 3 | 14 | 1 |
| ca-siRM147 | 2 1nt | 419 | 32 | 9 | 2 |
| 22 nt | 18 | 1 | 0 | 2 | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, C.; Li, G.; Wang, J.; Zhu, S.; Li, H. Cascading cis-Cleavage on Transcript from trans-Acting siRNA-Producing Locus 3. Int. J. Mol. Sci. 2013, 14, 14689-14699. https://doi.org/10.3390/ijms140714689
Zhang C, Li G, Wang J, Zhu S, Li H. Cascading cis-Cleavage on Transcript from trans-Acting siRNA-Producing Locus 3. International Journal of Molecular Sciences. 2013; 14(7):14689-14699. https://doi.org/10.3390/ijms140714689
Chicago/Turabian StyleZhang, Changqing, Guangping Li, Jin Wang, Shinong Zhu, and Hailing Li. 2013. "Cascading cis-Cleavage on Transcript from trans-Acting siRNA-Producing Locus 3" International Journal of Molecular Sciences 14, no. 7: 14689-14699. https://doi.org/10.3390/ijms140714689