Modulation by Melatonin of the Pathogenesis of Inflammatory Autoimmune Diseases


 ,
,
Abstract
:1. Introduction of Melatonin
1.1. The Discovery, Biosynthesis and Biological Function of Melatonin
1.2. The Effect of Melatonin on the Immune System
1.3. The Effect of Melatonin on the Production of Proinflammatory Cytokines
1.4. The Suppressive Effect of Melatonin on the Activation of NF-κB
1.5. The Modulatory Effect of Melatonin on the Th1/2 Balance
2. Overview of Autoimmune Diseases and Modulatory Effects of Melatonin on These Diseases
2.1. Multiple Sclerosis
2.1.1. The Pathogenesis and Animal Model of MS
2.1.2. The Role of Melatonin in MS
2.1.3. The Modulatory Role of Melatonin in EAE
2.2. Systemic Lupus Erythematosus
2.2.1. The Pathogenesis and Animal Model of SLE
2.2.2. The Dual Role of Melatonin in SLE Is Gender-Dependent in Animal Models
2.3. Rheumatoid Arthritis
2.3.1. Pathogenesis and Animal Model of RA
2.3.2. The Disease-Promoting Effect of Melatonin in RA
2.3.3. Investigation of Melatonin in an Animal Model of RA
2.4. Type 1 Diabetes Mellitus
2.4.1. Pathogenesis of T1D
2.4.2. Pathogenesis of Autoimmune Diabetes in NOD Mice
2.4.3. The Influence of Melatonin on the Insulin Production of Pancreatic β Cells
2.4.4. Modulatory Role of Melatonin in T1D
2.4.5. The Application of Melatonin on Islet Transplantation
2.5. Inflammatory Bowel Disease
2.5.1. Pathogenesis of IBD
2.5.2. The Therapeutic Potential of Melatonin Administration in IBD
2.5.3. Clinical Trial of Melatonin Treatment in IBD
3. The Future Application of Melatonin Treatment in Autoimmune Diseases
4. Concluding Remarks
Acknowledgments
Abbreviations
| EAE | experimental autoimmune encephalomyelitis |
| CIA | collagen-induced arthritis |
| AA | adjuvant-induced arthritis |
| i.p. | intraperitoneal injection |
| sc. | subcutaneous injection |
| dw | drinking water |
| SD | Sprague-Dawley |
| NOD | non-obese diabetic |
| DSS | dextran sodium sulphate |
| DNBS | dinitrobenzene sulfonic acid |
| TNBS | trinitrobenzene sulfonic acid |
Conflict of Interest
References
- Tan, D.X.; Manchester, L.C.; Hardeland, R.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Melatonin: A hormone, a tissue factor, an autocoid, a paracoid, and an antioxidant vitamin. J. Pineal Res. 2003, 34, 75–78. [Google Scholar]
- Reiter, R.J. The melatonin rhythm: Both a clock and a calendar. Experientia 1993, 49, 654–664. [Google Scholar]
- Malpaux, B.; Migaud, M.; Tricoire, H.; Chemineau, P. Biology of mammalian photoperiodism and the critical role of the pineal gland and melatonin. J. Biol. Rhythms 2001, 16, 336–347. [Google Scholar]
- Maldonado, M.D.; Murillo-Cabezas, F.; Terron, M.P.; Flores, L.J.; Tan, D.X.; Manchester, L.C.; Reiter, R.J. The potential of melatonin in reducing morbidity-mortality after craniocerebral trauma. J. Pineal Res 2007, 42, 1–11. [Google Scholar]
- Peyrot, F.; Ducrocq, C. Potential role of tryptophan derivatives in stress responses characterized by the generation of reactive oxygen and nitrogen species. J. Pineal Res 2008, 45, 235–246. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Terron, M.P.; Flores, L.J.; Czarnocki, Z. Melatonin and its metabolites: New findings regarding their production and their radical scavenging actions. Acta Biochim. Pol 2007, 54, 1–9. [Google Scholar]
- Reiter, R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev 1991, 12, 151–180. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Hardies, L.J.; Weintraub, S.T.; Vijayalaxmi Shepherd, A.M. A novel melatonin metabolite, cyclic 3-hydroxymelatonin: A biomarker of in vivo hydroxyl radical generation. Biochem. Biophys. Res. Commun. 1998, 253, 614–620. [Google Scholar]
- Bubenik, G.A. Thirty four years since the discovery of gastrointestinal melatonin. J. Physiol. Pharmacol 2008, 59, S33–S51. [Google Scholar]
- Jaworek, J.; Brzozowski, T.; Konturek, S.J. Melatonin as an organoprotector in the stomach and the pancreas. J. Pineal Res 2005, 38, 73–83. [Google Scholar]
- Stefulj, J.; Hortner, M.; Ghosh, M.; Schauenstein, K.; Rinner, I.; Wolfler, A.; Semmler, J.; Liebmann, P.M. Gene expression of the key enzymes of melatonin synthesis in extrapineal tissues of the rat. J. Pineal Res 2001, 30, 243–247. [Google Scholar]
- Gomez-Corvera, A.; Cerrillo, I.; Molinero, P.; Naranjo, M.C.; Lardone, P.J.; Sanchez-Hidalgo, M.; Carrascosa-Salmoral, M.P.; Medrano-Campillo, P.; Guerrero, J.M.; Rubio, A. Evidence of immune system melatonin production by two pineal melatonin deficient mice, C57BL/6 and Swiss strains. J. Pineal Res 2009, 47, 15–22. [Google Scholar]
- Carrillo-Vico, A.; Calvo, J.R.; Abreu, P.; Lardone, P.J.; Garcia-Maurino, S.; Reiter, R.J.; Guerrero, J.M. Evidence of melatonin synthesis by human lymphocytes and its physiological significance: Possible role as intracrine, autocrine, and/or paracrine substance. FASEB J 2004, 18, 537–539. [Google Scholar]
- Conti, A.; Conconi, S.; Hertens, E.; Skwarlo-Sonta, K.; Markowska, M.; Maestroni, J.M. Evidence for melatonin synthesis in mouse and human bone marrow cells. J. Pineal Res 2000, 28, 193–202. [Google Scholar]
- Naranjo, M.C.; Guerrero, J.M.; Rubio, A.; Lardone, P.J.; Carrillo-Vico, A.; Carrascosa-Salmoral, M.P.; Jimenez-Jorge, S.; Arellano, M.V.; Leal-Noval, S.R.; Leal, M.; et al. Melatonin biosynthesis in the thymus of humans and rats. Cell. Mol. Life Sci 2007, 64, 781–790. [Google Scholar]
- Carrillo-Vico, A.; Guerrero, J.M.; Lardone, P.J.; Reiter, R.J. A review of the multiple actions of melatonin on the immune system. Endocrine 2005, 27, 189–200. [Google Scholar]
- Garcia-Maurino, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Rafii-El-Idrissi, M.; Sanchez-Margalet, V.; Goberna, R.; Guerrero, J.M. Melatonin enhances IL-2, IL-6, and IFN-gamma production by human circulating CD4+ cells: A possible nuclear receptor-mediated mechanism involving T helper type 1 lymphocytes and monocytes. J. Immunol 1997, 159, 574–581. [Google Scholar]
- Pozo, D.; Delgado, M.; Fernandez-Santos, J.M.; Calvo, J.R.; Gomariz, R.P.; Martin-Lacave, I.; Ortiz, G.G.; Guerrero, J.M. Expression of the Mel1a-melatonin receptor mRNA in T and B subsets of lymphocytes from rat thymus and spleen. FASEB J 1997, 11, 466–473. [Google Scholar]
- Pioli, C.; Caroleo, M.C.; Nistico, G.; Doria, G. Melatonin increases antigen presentation and amplifies specific and non specific signals for T-cell proliferation. Int. J. Immunopharmacol 1993, 15, 463–468. [Google Scholar]
- Currier, N.L.; Sun, L.Z.; Miller, S.C. Exogenous melatonin: Quantitative enhancement in vivo of cells mediating non-specific immunity. J. Neuroimmunol 2000, 104, 101–108. [Google Scholar]
- Garcia-Maurino, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. Involvement of nuclear binding sites for melatonin in the regulation of IL-2 and IL-6 production by human blood mononuclear cells. J. Neuroimmunol 1998, 92, 76–84. [Google Scholar]
- Raghavendra, V.; Singh, V.; Kulkarni, S.K.; Agrewala, J.N. Melatonin enhances Th2 cell mediated immune responses: Lack of sensitivity to reversal by naltrexone or benzodiazepine receptor antagonists. Mol. Cell. Biochem 2001, 221, 57–62. [Google Scholar]
- Shaji, A.V.; Kulkarni, S.K.; Agrewala, J.N. Regulation of secretion of IL-4 and IgG1 isotype by melatonin-stimulated ovalbumin-specific T cells. Clin. Exp. Immunol 1998, 111, 181–185. [Google Scholar]
- Raghavendra, V.; Singh, V.; Shaji, A.V.; Vohra, H.; Kulkarni, S.K.; Agrewala, J.N. Melatonin provides signal 3 to unprimed CD4(+) T cells but failed to stimulate LPS primed B cells. Clin. Exp. Immunol 2001, 124, 414–422. [Google Scholar]
- Majewska, M.; Zajac, K.; Zemelka, M.; Szczepanik, M. Influence of melatonin and its precursor L-tryptophan on Th1 dependent contact hypersensitivity. J. Physiol. Pharmacol 2007, 58, S125–S132. [Google Scholar]
- Konakchieva, R.; Kyurkchiev, S.; Kehayov, I.; Taushanova, P.; Kanchev, L. Selective effect of methoxyindoles on the lymphocyte proliferation and melatonin binding to activated human lymphoid cells. J. Neuroimmunol 1995, 63, 125–132. [Google Scholar]
- Wang, H.; Wei, W.; Wang, N.P.; Gui, S.Y.; Wu, L.; Sun, W.Y.; Xu, S.Y. Melatonin ameliorates carbon tetrachloride-induced hepatic fibrogenesis in rats via inhibition of oxidative stress. Life Sci 2005, 77, 1902–1915. [Google Scholar]
- Lahiri, S.; Singh, P.; Singh, S.; Rasheed, N.; Palit, G.; Pant, K.K. Melatonin protects against experimental reflux esophagitis. J. Pineal Res 2009, 46, 207–213. [Google Scholar]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets 2005, 4, 471–479. [Google Scholar]
- Szczepanik, M. Melatonin and its influence on immune system. J. Physiol. Pharmacol 2007, 58, S115–S124. [Google Scholar]
- Crespo, E.; Macias, M.; Pozo, D.; Escames, G.; Martin, M.; Vives, F.; Guerrero, J.M.; Acuna-Castroviejo, D. Melatonin inhibits expression of the inducible NO synthase II in liver and lung and prevents endotoxemia in lipopolysaccharide-induced multiple organ dysfunction syndrome in rats. FASEB J 1999, 13, 1537–1546. [Google Scholar]
- Jung, K.H.; Hong, S.W.; Zheng, H.M.; Lee, D.H.; Hong, S.S. Melatonin downregulates nuclear erythroid 2-related factor 2 and nuclear factor-kappaB during prevention of oxidative liver injury in a dimethylnitrosamine model. J. Pineal Res 2009, 47, 173–183. [Google Scholar]
- Veneroso, C.; Tunon, M.J.; Gonzalez-Gallego, J.; Collado, P.S. Melatonin reduces cardiac inflammatory injury induced by acute exercise. J. Pineal Res 2009, 47, 184–191. [Google Scholar]
- Lopez, L.C.; Escames, G.; Tapias, V.; Utrilla, P.; Leon, J.; Acuna-Castroviejo, D. Identification of an inducible nitric oxide synthase in diaphragm mitochondria from septic mice: Its relation with mitochondrial dysfunction and prevention by melatonin. Int. J. Biochem. Cell. Biol 2006, 38, 267–278. [Google Scholar]
- Escames, G.; Lopez, L.C.; Ortiz, F.; Lopez, A.; Garcia, J.A.; Ros, E.; Acuna-Castroviejo, D. Attenuation of cardiac mitochondrial dysfunction by melatonin in septic mice. FEBS J 2007, 274, 2135–2147. [Google Scholar]
- Lowes, D.A.; Almawash, A.M.; Webster, N.R.; Reid, V.L.; Galley, H.F. Melatonin and structurally similar compounds have differing effects on inflammation and mitochondrial function in endothelial cells under conditions mimicking sepsis. Br. J. Anaesth 2011, 107, 193–201. [Google Scholar]
- Lowes, D.A.; Webster, N.R.; Murphy, M.P.; Galley, H.F. Antioxidants that protect mitochondria reduce interleukin-6 and oxidative stress, improve mitochondrial function, and reduce biochemical markers of organ dysfunction in a rat model of acute sepsis. Br. J. Anaesth 2013, 110, 472–480. [Google Scholar]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Invest 2001, 107, 7–11. [Google Scholar]
- Chuang, J.I.; Mohan, N.; Meltz, M.L.; Reiter, R.J. Effect of melatonin on NF-kappa-B DNA-binding activity in the rat spleen. Cell. Biol. Int 1996, 20, 687–692. [Google Scholar]
- Bertuglia, S.; Marchiafava, P.L.; Colantuoni, A. Melatonin prevents ischemia reperfusion injury in hamster cheek pouch microcirculation. Cardiovasc. Res 1996, 31, 947–952. [Google Scholar]
- Sewerynek, E.; Reiter, R.J.; Melchiorri, D.; Ortiz, G.G.; Lewinski, A. Oxidative damage in the liver induced by ischemia-reperfusion: Protection by melatonin. Hepatogastroenterology 1996, 43, 898–905. [Google Scholar]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and its relation to the immune system and inflammation. Ann. N. Y. Acad. Sci 2000, 917, 376–86. [Google Scholar]
- Gilad, E.; Wong, H.R.; Zingarelli, B.; Virag, L.; O’Connor, M.; Salzman, A.L.; Szabo, C. Melatonin inhibits expression of the inducible isoform of nitric oxide synthase in murine macrophages: Role of inhibition of NF-κB activation. FASEB J 1998, 12, 685–693. [Google Scholar]
- Li, Z.; Nickkholgh, A.; Yi, X.; Bruns, H.; Gross, M.L.; Hoffmann, K.; Mohr, E.; Zorn, M.; Buchler, M.W.; Schemmer, P. Melatonin protects kidney grafts from ischemia/reperfusion injury through inhibition of NF-κB and apoptosis after experimental kidney transplantation. J. Pineal Res 2009, 46, 365–372. [Google Scholar]
- Ozbek, E.; Ilbey, Y.O.; Ozbek, M.; Simsek, A.; Cekmen, M.; Somay, A. Melatonin attenuates unilateral ureteral obstruction-induced renal injury by reducing oxidative stress, iNOS, MAPK, and NF-κB expression. J. Endourol 2009, 23, 1165–1173. [Google Scholar]
- Pascua, P.; Camello-Almaraz, C.; Camello, P.J.; Martin-Cano, F.E.; Vara, E.; Fernandez-Tresguerres, J.A.; Pozo, M.J. Melatonin, and to a lesser extent growth hormone, restores colonic smooth muscle physiology in old rats. J. Pineal Res 2011, 51, 405–415. [Google Scholar]
- Yip, H.K.; Chang, Y.C.; Wallace, C.G.; Chang, L.T.; Tsai, T.H.; Chen, Y.L.; Chang, H.W.; Leu, S.; Zhen, Y.Y.; Tsai, C.Y.; et al. Melatonin treatment improves adipose-derived mesenchymal stem cell therapy for acute lung ischemia-reperfusion injury. J. Pineal Res 2013, 54, 207–221. [Google Scholar]
- Qin, W.; Lu, W.; Li, H.; Yuan, X.; Li, B.; Zhang, Q.; Xiu, R. Melatonin inhibits IL1beta-induced MMP9 expression and activity in human umbilical vein endothelial cells by suppressing NF-κB activation. J. Endocrinol 2012, 214, 145–153. [Google Scholar]
- Xia, M.Z.; Liang, Y.L.; Wang, H.; Chen, X.; Huang, Y.Y.; Zhang, Z.H.; Chen, Y.H.; Zhang, C.; Zhao, M.; Xu, D.X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88- and TRIF-dependent signaling pathways in lipopolysaccharide-stimulated RAW264.7 cells. J. Pineal Res 2012, 53, 325–334. [Google Scholar]
- Lim, H.D.; Kim, Y.S.; Ko, S.H.; Yoon, I.J.; Cho, S.G.; Chun, Y.H.; Choi, B.J.; Kim, E.C. Cytoprotective and anti-inflammatory effects of melatonin in hydrogen peroxide-stimulated CHON-001 human chondrocyte cell line and rabbit model of osteoarthritis via the SIRT1 pathway. J. Pineal Res 2012, 53, 225–237. [Google Scholar]
- Choi, E.Y.; Jin, J.Y.; Lee, J.Y.; Choi, J.I.; Choi, I.S.; Kim, S.J. Melatonin inhibits prevotella intermedia lipopolysaccharide-induced production of nitric oxide and interleukin-6 in murine macrophages by suppressing NF-κB and STAT1 activity. J. Pineal Res. 2011, 50, 197–206. [Google Scholar]
- Wang, J.; Xiao, X.; Zhang, Y.; Shi, D.; Chen, W.; Fu, L.; Liu, L.; Xie, F.; Kang, T.; Huang, W.; et al. Simultaneous modulation of COX-2, p300, Akt, and Apaf-1 signaling by melatonin to inhibit proliferation and induce apoptosis in breast cancer cells. J. Pineal Res 2012, 53, 77–90. [Google Scholar]
- Shi, D.; Xiao, X.; Wang, J.; Liu, L.; Chen, W.; Fu, L.; Xie, F.; Huang, W.; Deng, W. Melatonin suppresses proinflammatory mediators in lipopolysaccharide-stimulated CRL1999 cells via targeting MAPK, NF-κB, c/EBPbeta, and p300 signaling. J. Pineal Res 2012, 53, 154–165. [Google Scholar]
- Marino, A.; Di Paola, R.; Crisafulli, C.; Mazzon, E.; Morabito, R.; Paterniti, I.; Galuppo, M.; Genovese, T.; La Spada, G.; Cuzzocrea, S. Protective effect of melatonin against the inflammatory response elicited by crude venom from isolated nematocysts of Pelagia noctiluca (Cnidaria, Scyphozoa). J. Pineal Res 2009, 47, 56–69. [Google Scholar]
- Kang, J.W.; Koh, E.J.; Lee, S.M. Melatonin protects liver against ischemia and reperfusion injury through inhibition of toll-like receptor signaling pathway. J. Pineal Res 2011, 50, 403–411. [Google Scholar]
- Wang, J.; Hao, H.; Yao, L.; Zhang, X.; Zhao, S.; Ling, E.A.; Hao, A.; Li, G. Melatonin suppresses migration and invasion via inhibition of oxidative stress pathway in glioma cells. J. Pineal Res 2012, 53, 180–187. [Google Scholar]
- Petrovsky, N.; Harrison, L.C. Diurnal rhythmicity of human cytokine production: A dynamic disequilibrium in T helper cell type 1/T helper cell type 2 balance? J. Immunol 1997, 158, 5163–5168. [Google Scholar]
- Garcia-Maurino, S.; Pozo, D.; Carrillo-Vico, A.; Calvo, J.R.; Guerrero, J.M. Melatonin activates Th1 lymphocytes by increasing IL-12 production. Life Sci 1999, 65, 2143–2150. [Google Scholar]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. Pineal melatonin, its fundamental immunoregulatory role in aging and cancer. Ann. N. Y. Acad. Sci 1988, 521, 140–148. [Google Scholar]
- Inserra, P.; Zhang, Z.; Ardestani, S.K.; Araghi-Niknam, M.; Liang, B.; Jiang, S.; Shaw, D.; Molitor, M.; Elliott, K.; Watson, R.R. Modulation of cytokine production by dehydroepiandrosterone (DHEA) plus melatonin (MLT) supplementation of old mice. Proc. Soc. Exp. Biol. Med 1998, 218, 76–82. [Google Scholar]
- Jung, F.J.; Yang, L.; Harter, L.; Inci, I.; Schneiter, D.; Lardinois, D.; Keel, M.; Weder, W.; Korom, S. Melatonin in vivo prolongs cardiac allograft survival in rats. J. Pineal Res 2004, 37, 36–41. [Google Scholar]
- Davidson, A.; Diamond, B. Autoimmune diseases. N. Engl. J. Med 2001, 345, 340–350. [Google Scholar]
- Steinman, L. Multiple sclerosis: A coordinated immunological attack against myelin in the central nervous system. Cell 1996, 85, 299–302. [Google Scholar]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol 2005, 23, 683–747. [Google Scholar]
- Ercolini, A.M.; Miller, S.D. Mechanisms of immunopathology in murine models of central nervous system demyelinating disease. J. Immunol 2006, 176, 3293–3298. [Google Scholar]
- Martin, R.; McFarland, H.F.; McFarlin, D.E. Immunological aspects of demyelinating diseases. Annu. Rev. Immunol 1992, 10, 153–187. [Google Scholar]
- Sobel, R.A.; Blanchette, B.W.; Bhan, A.K.; Colvin, R.B. The immunopathology of experimental allergic encephalomyelitis. I. Quantitative analysis of inflammatory cells in situ. J. Immunol 1984, 132, 2393–2401. [Google Scholar]
- Besler, H.T.; Comoglu, S. Lipoprotein oxidation, plasma total antioxidant capacity and homocysteine level in patients with multiple sclerosis. Nutr. Neurosci 2003, 6, 189–196. [Google Scholar]
- Glabinski, A.; Tawsek, N.S.; Bartosz, G. Increased generation of superoxide radicals in the blood of MS patients. Acta Neurol. Scand 1993, 88, 174–177. [Google Scholar]
- Hunter, M.I.; Nlemadim, B.C.; Davidson, D.L. Lipid peroxidation products and antioxidant proteins in plasma and cerebrospinal fluid from multiple sclerosis patients. Neurochem. Res 1985, 10, 1645–1652. [Google Scholar]
- MacMicking, J.D.; Willenborg, D.O.; Weidemann, M.J.; Rockett, K.A.; Cowden, W.B. Elevated secretion of reactive nitrogen and oxygen intermediates by inflammatory leukocytes in hyperacute experimental autoimmune encephalomyelitis: Enhancement by the soluble products of encephalitogenic T cells. J. Exp. Med 1992, 176, 303–307. [Google Scholar]
- Brass, S.D.; Duquette, P.; Proulx-Therrien, J.; Auerbach, S. Sleep disorders in patients with multiple sclerosis. Sleep. Med. Rev 2010, 14, 121–129. [Google Scholar]
- Melamud, L.; Golan, D.; Luboshitzky, R.; Lavi, I.; Miller, A. Melatonin dysregulation, sleep disturbances and fatigue in multiple sclerosis. J. Neurol. Sci 2012, 314, 37–40. [Google Scholar]
- Hedstrom, A.K.; Akerstedt, T.; Hillert, J.; Olsson, T.; Alfredsson, L. Shift work at young age is associated with increased risk for multiple sclerosis. Ann. Neurol 2011, 70, 733–741. [Google Scholar]
- Kang, J.C.; Ahn, M.; Kim, Y.S.; Moon, C.; Lee, Y.; Wie, M.B.; Lee, Y.J.; Shin, T. Melatonin ameliorates autoimmune encephalomyelitis through suppression of intercellular adhesion molecule-1. J. Vet. Sci 2001, 2, 85–89. [Google Scholar]
- Constantinescu, C.S.; Hilliard, B.; Ventura, E.; Rostami, A. Luzindole, a melatonin receptor antagonist, suppresses experimental autoimmune encephalomyelitis. Pathobiology 1997, 65, 190–194. [Google Scholar]
- Dubocovich, M.L.; Yun, K.; Al-Ghoul, W.M.; Benloucif, S.; Masana, M.I. Selective MT2 melatonin receptor antagonists block melatonin-mediated phase advances of circadian rhythms. FASEB J 1998, 12, 1211–1220. [Google Scholar]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med 2011, 365, 2110–2121. [Google Scholar]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev 2003, 8, 223–246. [Google Scholar]
- Hayashi, T. Therapeutic strategies for SLE involving cytokines: Mechanism-oriented therapies especially IFN-gamma targeting gene therapy. J. Biomed. Biotechnol. 2010, 2010. [Google Scholar] [CrossRef]
- Wong, C.K.; Lit, L.C.; Tam, L.S.; Li, E.K.; Wong, P.T.; Lam, C.W. Hyperproduction of IL-23 and IL-17 in patients with systemic lupus erythematosus: Implications for Th17-mediated inflammation in auto-immunity. Clin. Immunol 2008, 127, 385–393. [Google Scholar]
- Lechner, O.; Dietrich, H.; Oliveira dos Santos, A.; Wiegers, G.J.; Schwarz, S.; Harbutz, M.; Herold, M.; Wick, G. Altered circadian rhythms of the stress hormone and melatonin response in lupus-prone MRL/MP-fas(Ipr) mice. J. Autoimmun 2000, 14, 325–333. [Google Scholar]
- Yin, Z.; Bahtiyar, G.; Zhang, N.; Liu, L.; Zhu, P.; Robert, M.E.; McNiff, J.; Madaio, M.P.; Craft, J. IL-10 regulates murine lupus. J. Immunol 2002, 169, 2148–2155. [Google Scholar]
- Jimenez-Caliani, A.J.; Jimenez-Jorge, S.; Molinero, P.; Fernandez-Santos, J.M.; Martin-Lacave, I.; Rubio, A.; Guerrero, J.M.; Osuna, C. Sex-dependent effect of melatonin on systemic erythematosus lupus developed in Mrl/Mpj-Faslpr mice: It ameliorates the disease course in females, whereas it exacerbates it in males. Endocrinology 2006, 147, 1717–1724. [Google Scholar]
- Jimenez-Caliani, A.J.; Jimenez-Jorge, S.; Molinero, P.; Rubio, A.; Guerrero, J.M.; Osuna, C. Treatment with testosterone or estradiol in melatonin treated females and males MRL/MpJ-Faslpr mice induces negative effects in developing systemic lupus erythematosus. J. Pineal Res 2008, 45, 204–211. [Google Scholar]
- Zhou, L.L.; Wei, W.; Si, J.F.; Yuan, D.P. Regulatory effect of melatonin on cytokine disturbances in the pristane-induced lupus mice. Mediators Inflamm. 2010. [Google Scholar] [CrossRef]
- Wu, C.C.; Lu, K.C.; Lin, G.J.; Hsieh, H.Y.; Chu, P.; Lin, S.H.; Sytwu, H.K. Melatonin enhances endogenous heme oxygenase-1 and represses immune responses to ameliorate experimental murine membranous nephropathy. J. Pineal Res 2012, 52, 460–469. [Google Scholar]
- Choy, E.H.; Panayi, G.S. Cytokine pathways and joint inflammation in rheumatoid arthritis. N. Engl. J. Med 2001, 344, 907–916. [Google Scholar]
- Sulli, A.; Maestroni, G.J.; Villaggio, B.; Hertens, E.; Craviotto, C.; Pizzorni, C.; Briata, M.; Seriolo, B.; Cutolo, M. Melatonin serum levels in rheumatoid arthritis. Ann. N. Y. Acad. Sci 2002, 966, 276–283. [Google Scholar]
- Cutolo, M.; Straub, R.H. Circadian rhythms in arthritis: Hormonal effects on the immune/inflammatory reaction. Autoimmun. Rev 2008, 7, 223–228. [Google Scholar]
- Cutolo, M.; Balleari, E.; Giusti, M.; Monachesi, M.; Accardo, S. Sex hormone status of male patients with rheumatoid arthritis: Evidence of low serum concentrations of testosterone at baseline and after human chorionic gonadotropin stimulation. Arthritis Rheum 1988, 31, 1314–1317. [Google Scholar]
- Valenti, S.; Giusti, M. Melatonin participates in the control of testosterone secretion from rat testis: An overview of our experience. Ann. N. Y. Acad. Sci 2002, 966, 284–289. [Google Scholar]
- Forrest, C.M.; Mackay, G.M.; Stoy, N.; Stone, T.W.; Darlington, L.G. Inflammatory status and kynurenine metabolism in rheumatoid arthritis treated with melatonin. Br. J. Clin. Pharmacol 2007, 64, 517–526. [Google Scholar]
- Hansson, I.; Holmdahl, R.; Mattsson, R. Constant darkness enhances autoimmunity to type II collagen and exaggerates development of collagen-induced arthritis in DBA/1 mice. J. Neuroimmunol 1990, 27, 79–84. [Google Scholar]
- Hansson, I.; Holmdahl, R.; Mattsson, R. The pineal hormone melatonin exaggerates development of collagen-induced arthritis in mice. J. Neuroimmunol. 1992, 39, 23–30. [Google Scholar]
- Hansson, I.; Holmdahl, R.; Mattsson, R. Pinealectomy ameliorates collagen II-induced arthritis in mice. Clin. Exp. Immunol 1993, 92, 432–436. [Google Scholar]
- Chen, Q.; Wei, W. Effects and mechanisms of melatonin on inflammatory and immune responses of adjuvant arthritis rat. Int. Immunopharmacol 2002, 2, 1443–1449. [Google Scholar]
- Bang, J.; Chang, H.W.; Jung, H.R.; Cho, C.H.; Hur, J.A.; Lee, S.I.; Choi, T.H.; Kim, S.H.; Ha, E. Melatonin attenuates clock gene cryptochrome1, which may aggravate mouse anti-type II collagen antibody-induced arthritis. Rheumatol. Int 2012, 32, 379–385. [Google Scholar]
- Jimenez-Caliani, A.J.; Jimenez-Jorge, S.; Molinero, P.; Guerrero, J.M.; Fernandez-Santos, J.M.; Martin-Lacave, I.; Osuna, C. Dual effect of melatonin as proinflammatory and antioxidant in collagen-induced arthritis in rats. J. Pineal Res 2005, 38, 93–99. [Google Scholar]
- Kawasaki, E.; Abiru, N.; Eguchi, K. Prevention of type 1 diabetes: From the view point of beta cell damage. Diabetes Res. Clin. Pract 2004, 66, S27–S32. [Google Scholar]
- Aoki, C.A.; Borchers, A.T.; Ridgway, W.M.; Keen, C.L.; Ansari, A.A.; Gershwin, M.E. NOD mice and autoimmunity. Autoimmun. Rev 2005, 4, 373–379. [Google Scholar]
- Wicker, L.S.; Todd, J.A.; Peterson, L.B. Genetic control of autoimmune diabetes in the NOD mouse. Annu. Rev. Immunol 1995, 13, 179–200. [Google Scholar]
- Fox, H.S. Androgen treatment prevents diabetes in nonobese diabetic mice. J. Exp. Med 1992, 175, 1409–1412. [Google Scholar]
- Liblau, R.S.; Singer, S.M.; McDevitt, H.O. Th1 and Th2 CD4+ T cells in the pathogenesis of organ-specific autoimmune diseases. Immunol. Today 1995, 16, 34–38. [Google Scholar]
- Hung, J.T.; Liao, J.H.; Lin, Y.C.; Chang, H.Y.; Wu, S.F.; Chang, T.H.; Kung, J.T.; Hsieh, S.L.; McDevitt, H.; Sytwu, H.K. Immunopathogenic role of TH1 cells in autoimmune diabetes: Evidence from a T1 and T2 doubly transgenic non-obese diabetic mouse model. J. Autoimmun 2005, 25, 181–192. [Google Scholar]
- Koarada, S.; Wu, Y.; Olshansky, G.; Ridgway, W.M. Increased nonobese diabetic Th1:Th2 (IFN-gamma:IL-4) ratio is CD4+ T cell intrinsic and independent of APC genetic background. J. Immunol 2002, 169, 6580–6587. [Google Scholar]
- Debray-Sachs, M.; Carnaud, C.; Boitard, C.; Cohen, H.; Gresser, I.; Bedossa, P.; Bach, J.F. Prevention of diabetes in NOD mice treated with antibody to murine IFN gamma. J. Autoimmun 1991, 4, 237–248. [Google Scholar]
- Campbell, I.L.; Kay, T.W.; Oxbrow, L.; Harrison, L.C. Essential role for interferon-gamma and interleukin-6 in autoimmune insulin-dependent diabetes in NOD/Wehi mice. J. Clin. Invest 1991, 87, 739–742. [Google Scholar]
- Cameron, M.J.; Arreaza, G.A.; Zucker, P.; Chensue, S.W.; Strieter, R.M.; Chakrabarti, S.; Delovitch, T.L. IL-4 prevents insulitis and insulin-dependent diabetes mellitus in nonobese diabetic mice by potentiation of regulatory T helper-2 cell function. J. Immunol 1997, 159, 4686–4692. [Google Scholar]
- Pennline, K.J.; Roque-Gaffney, E.; Monahan, M. Recombinant human IL-10 prevents the onset of diabetes in the nonobese diabetic mouse. Clin. Immunol. Immunopathol 1994, 71, 169–175. [Google Scholar]
- Peschke, E. Melatonin, endocrine pancreas and diabetes. J. Pineal Res 2008, 44, 26–40. [Google Scholar]
- Peschke, E.; Wolgast, S.; Bazwinsky, I.; Ponicke, K.; Muhlbauer, E. Increased melatonin synthesis in pineal glands of rats in streptozotocin induced type 1 diabetes. J. Pineal Res 2008, 45, 439–448. [Google Scholar]
- Stumpf, I.; Muhlbauer, E.; Peschke, E. Involvement of the cGMP pathway in mediating the insulin-inhibitory effect of melatonin in pancreatic beta-cells. J. Pineal Res 2008, 45, 318–327. [Google Scholar]
- Ramracheya, R.D.; Muller, D.S.; Squires, P.E.; Brereton, H.; Sugden, D.; Huang, G.C.; Amiel, S.A.; Jones, P.M.; Persaud, S.J. Function and expression of melatonin receptors on human pancreatic islets. J. Pineal Res 2008, 44, 273–279. [Google Scholar]
- Conti, A.; Maestroni, G.J. Role of the pineal gland and melatonin in the development of autoimmune diabetes in non-obese diabetic mice. J. Pineal Res 1996, 20, 164–172. [Google Scholar]
- Okitsu, T.; Bartlett, S.T.; Hadley, G.A.; Drachenberg, C.B.; Farney, A.C. Recurrent autoimmunity accelerates destruction of minor and major histoincompatible islet grafts in nonobese diabetic (NOD) mice. Am. J. Transplant 2001, 1, 138–145. [Google Scholar]
- Young, H.Y.; Zucker, P.; Flavell, R.A.; Jevnikar, A.M.; Singh, B. Characterization of the role of major histocompatibility complex in type 1 diabetes recurrence after islet transplantation. Transplantation 2004, 78, 509–515. [Google Scholar]
- Sibley, R.K.; Sutherland, D.E.; Goetz, F.; Michael, A.F. Recurrent diabetes mellitus in the pancreas iso- and allograft. A light and electron microscopic and immunohistochemical analysis of four cases. Lab. Invest 1985, 53, 132–144. [Google Scholar]
- Tyden, G.; Reinholt, F.P.; Sundkvist, G.; Bolinder, J. Recurrence of autoimmune diabetes mellitus in recipients of cadaveric pancreatic grafts. N. Engl. J. Med 1996, 335, 860–863. [Google Scholar]
- Gysemans, C.A.; Waer, M.; Valckx, D.; Laureys, J.M.; Mihkalsky, D.; Bouillon, R.; Mathieu, C. Early graft failure of xenogeneic islets in NOD mice is accompanied by high levels of interleukin-1 and low levels of transforming growth factor-beta mRNA in the grafts. Diabetes 2000, 49, 1992–1997. [Google Scholar]
- Sapmaz, E.; Ayar, A.; Celik, H.; Sapmaz, T.; Kilic, N.; Yasar, M.A. Effects of melatonin and oxytetracycline in autologous intraperitoneal ovary transplantation in rats. Neuro Endocrinol. Lett 2003, 24, 350–354. [Google Scholar]
- Lin, G.J.; Huang, S.H.; Chen, Y.W.; Hueng, D.Y.; Chien, M.W.; Chia, W.T.; Chang, D.M.; Sytwu, H.K. Melatonin prolongs islet graft survival in diabetic NOD mice. J. Pineal Res 2009, 47, 284–292. [Google Scholar]
- Zhang, Y.C.; Pileggi, A.; Agarwal, A.; Molano, R.D.; Powers, M.; Brusko, T.; Wasserfall, C.; Goudy, K.; Zahr, E.; Poggioli, R.; et al. Adeno-associated virus-mediated IL-10 gene therapy inhibits diabetes recurrence in syngeneic islet cell transplantation of NOD mice. Diabetes 2003, 52, 708–716. [Google Scholar]
- Kim, Y.H.; Lim, D.G.; Wee, Y.M.; Kim, J.H.; Yun, C.O.; Choi, M.Y.; Park, Y.H.; Kim, S.C.; Han, D.J. Viral IL-10 gene transfer prolongs rat islet allograft survival. Cell Transplant 2008, 17, 609–618. [Google Scholar]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med 2009, 361, 2066–2078. [Google Scholar]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The immunology of mucosal models of inflammation. Annu. Rev. Immunol 2002, 20, 495–549. [Google Scholar]
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol 2006, 3, 390–407. [Google Scholar]
- Neurath, M.F.; Finotto, S. The many roads to inflammatory bowel diseases. Immunity 2006, 25, 189–191. [Google Scholar]
- Fuss, I.J.; Neurath, M.; Boirivant, M.; Klein, J.S.; de la Motte, C.; Strong, S.A.; Fiocchi, C.; Strober, W. Disparate CD4+ lamina propria (LP) lymphokine secretion profiles in inflammatory bowel disease. Crohn’s disease LP cells manifest increased secretion of IFN-gamma, whereas ulcerative colitis LP cells manifest increased secretion of IL-5. J. Immunol 1996, 157, 1261–1270. [Google Scholar]
- Barrett, J.C.; Hansoul, S.; Nicolae, D.L.; Cho, J.H.; Duerr, R.H.; Rioux, J.D.; Brant, S.R.; Silverberg, M.S.; Taylor, K.D.; Barmada, M.M.; et al. Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat. Genet 2008, 40, 955–962. [Google Scholar]
- Fujino, S.; Andoh, A.; Bamba, S.; Ogawa, A.; Hata, K.; Araki, Y.; Bamba, T.; Fujiyama, Y. Increased expression of interleukin 17 in inflammatory bowel disease. Gut 2003, 52, 65–70. [Google Scholar]
- Elson, C.O.; Cong, Y.; Weaver, C.T.; Schoeb, T.R.; McClanahan, T.K.; Fick, R.B.; Kastelein, R.A. Monoclonal anti-interleukin 23 reverses active colitis in a T cell-mediated model in mice. Gastroenterology 2007, 132, 2359–2370. [Google Scholar]
- Hue, S.; Ahern, P.; Buonocore, S.; Kullberg, M.C.; Cua, D.J.; McKenzie, B.S.; Powrie, F.; Maloy, K.J. Interleukin-23 drives innate and T cell-mediated intestinal inflammation. J. Exp. Med 2006, 203, 2473–2483. [Google Scholar]
- Pentney, P.T.; Bubenik, G.A. Melatonin reduces the severity of dextran-induced colitis in mice. J. Pineal Res 1995, 19, 31–39. [Google Scholar]
- Nosal’ova, V.; Zeman, M.; Cerna, S.; Navarova, J.; Zakalova, M. Protective effect of melatonin in acetic acid induced colitis in rats. J. Pineal Res 2007, 42, 364–370. [Google Scholar]
- Dong, W.G.; Mei, Q.; Yu, J.P.; Xu, J.M.; Xiang, L.; Xu, Y. Effects of melatonin on the expression of iNOS and COX-2 in rat models of colitis. World J. Gastroenterol 2003, 9, 1307–1311. [Google Scholar]
- Cuzzocrea, S.; Mazzon, E.; Serraino, I.; Lepore, V.; Terranova, M.L.; Ciccolo, A.; Caputi, A.P. Melatonin reduces dinitrobenzene sulfonic acid-induced colitis. J. Pineal Res 2001, 30, 1–12. [Google Scholar]
- Mazzon, E.; Esposito, E.; Crisafulli, C.; Riccardi, L.; Muia, C.; di Bella, P.; Meli, R.; Cuzzocrea, S. Melatonin modulates signal transduction pathways and apoptosis in experimental colitis. J. Pineal Res. 2006, 41, 363–373. [Google Scholar]
- Esposito, E.; Mazzon, E.; Riccardi, L.; Caminiti, R.; Meli, R.; Cuzzocrea, S. Matrix metalloproteinase-9 and metalloproteinase-2 activity and expression is reduced by melatonin during experimental colitis. J. Pineal Res 2008, 45, 166–173. [Google Scholar]
- Necefli, A.; Tulumoglu, B.; Giris, M.; Barbaros, U.; Gunduz, M.; Olgac, V.; Guloglu, R.; Toker, G. The effect of melatonin on TNBS-induced colitis. Dig. Dis. Sci 2006, 51, 1538–1545. [Google Scholar]
- Li, J.H.; Yu, J.P.; Yu, H.G.; Xu, X.M.; Yu, L.L.; Liu, J.; Luo, H.S. Melatonin reduces inflammatory injury through inhibiting NF-κB activation in rats with colitis. Mediators Inflamm 2005, 2005, 185–193. [Google Scholar]
- Mei, Q.; Xu, J.M.; Xiang, L.; Hu, Y.M.; Hu, X.P.; Xu, Z.W. Change of nitric oxide in experimental colitis and its inhibition by melatonin in vivo and in vitro. Postgrad. Med. J 2005, 81, 667–672. [Google Scholar]
- Marquez, E.; Sanchez-Fidalgo, S.; Calvo, J.R.; la de Lastra, C.A.; Motilva, V. Acutely administered melatonin is beneficial while chronic melatonin treatment aggravates the evolution of TNBS-induced colitis. J. Pineal Res 2006, 40, 48–55. [Google Scholar]
- Chojnacki, C.; Wisniewska-Jarosinska, M.; Walecka-Kapica, E.; Klupinska, G.; Jaworek, J.; Chojnacki, J. Evaluation of melatonin effectiveness in the adjuvant treatment of ulcerative colitis. J. Physiol. Pharmacol 2011, 62, 327–334. [Google Scholar]
- Prud'homme, G.J. Gene therapy of autoimmune diseases with vectors encoding regulatory cytokines or inflammatory cytokine inhibitors. J. Gene Med 2000, 2, 222–232. [Google Scholar]
- Ruiz, P.A.; Shkoda, A.; Kim, S.C.; Sartor, R.B.; Haller, D. IL-10 gene-deficient mice lack TGF-beta/Smad-mediated TLR2 degradation and fail to inhibit proinflammatory gene expression in intestinal epithelial cells under conditions of chronic inflammation. Ann. N. Y. Acad. Sci 2006, 1072, 389–394. [Google Scholar]
- Balasa, B.; Van Gunst, K.; Jung, N.; Katz, J.D.; Sarvetnick, N. IL-10 deficiency does not inhibit insulitis and accelerates cyclophosphamide-induced diabetes in the nonobese diabetic mouse. Cell. Immunol 2000, 202, 97–102. [Google Scholar]
- Kuhn, R.; Lohler, J.; Rennick, D.; Rajewsky, K.; Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar]
- Groux, H.; O’Garra, A.; Bigler, M.; Rouleau, M.; Antonenko, S.; de Vries, J.E.; Roncarolo, M.G. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 1997, 389, 737–742. [Google Scholar]
- Hardeland, R.; Pandi-Perumal, S.R. Melatonin, a potent agent in antioxidative defense: Actions as a natural food constituent, gastrointestinal factor, drug and prodrug. Nutr. Metab 2005, 2, 22. [Google Scholar]
- Kucukakin, B.; Lykkesfeldt, J.; Nielsen, H.J.; Reiter, R.J.; Rosenberg, J.; Gogenur, I. Utility of melatonin to treat surgical stress after major vascular surgery—A safety study. J. Pineal Res 2008, 44, 426–431. [Google Scholar]
- Gitto, E.; Pellegrino, S.; Gitto, P.; Barberi, I.; Reiter, R.J. Oxidative stress of the newborn in the pre- and postnatal period and the clinical utility of melatonin. J. Pineal Res 2009, 46, 128–139. [Google Scholar]
- Kennaway, D.J.; Seamark, R.F. Circulating levels of melatonin following its oral administration or subcutaneous injection in sheep and goats. Aust. J. Biol. Sci 1980, 33, 349–353. [Google Scholar]
- DeMuro, R.L.; Nafziger, A.N.; Blask, D.E.; Menhinick, A.M.; Bertino, J.S., Jr. The absolute bioavailability of oral melatonin. J. Clin. Pharmacol. 2000, 40, 781–784. [Google Scholar]
- Jacob, S.; Poeggeler, B.; Weishaupt, J.H.; Siren, A.L.; Hardeland, R.; Bahr, M.; Ehrenreich, H. Melatonin as a candidate compound for neuroprotection in amyotrophic lateral sclerosis (ALS): High tolerability of daily oral melatonin administration in ALS patients. J. Pineal Res 2002, 33, 186–187. [Google Scholar]
- Weishaupt, J.H.; Bartels, C.; Polking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Huther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal Res 2006, 41, 313–323. [Google Scholar]
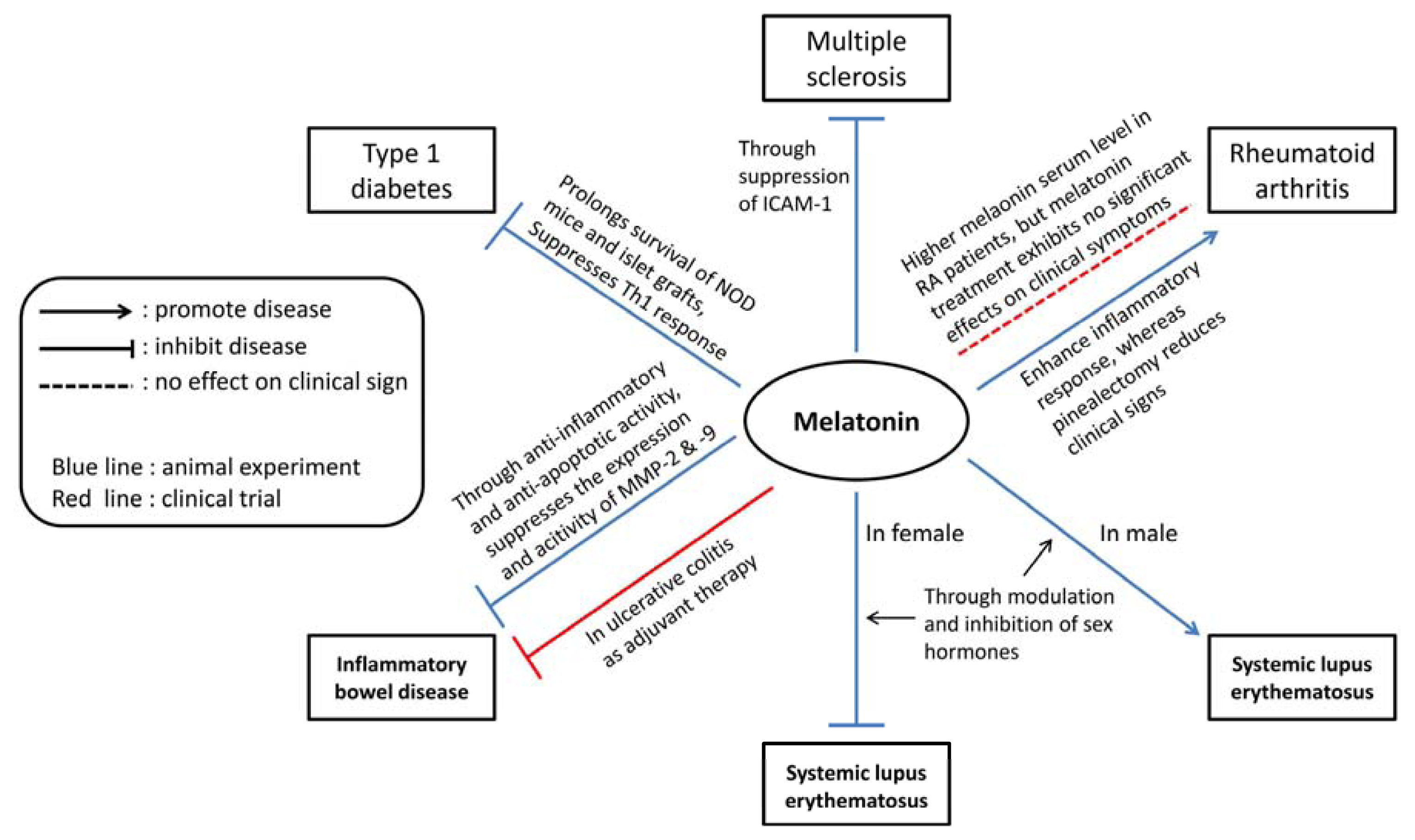

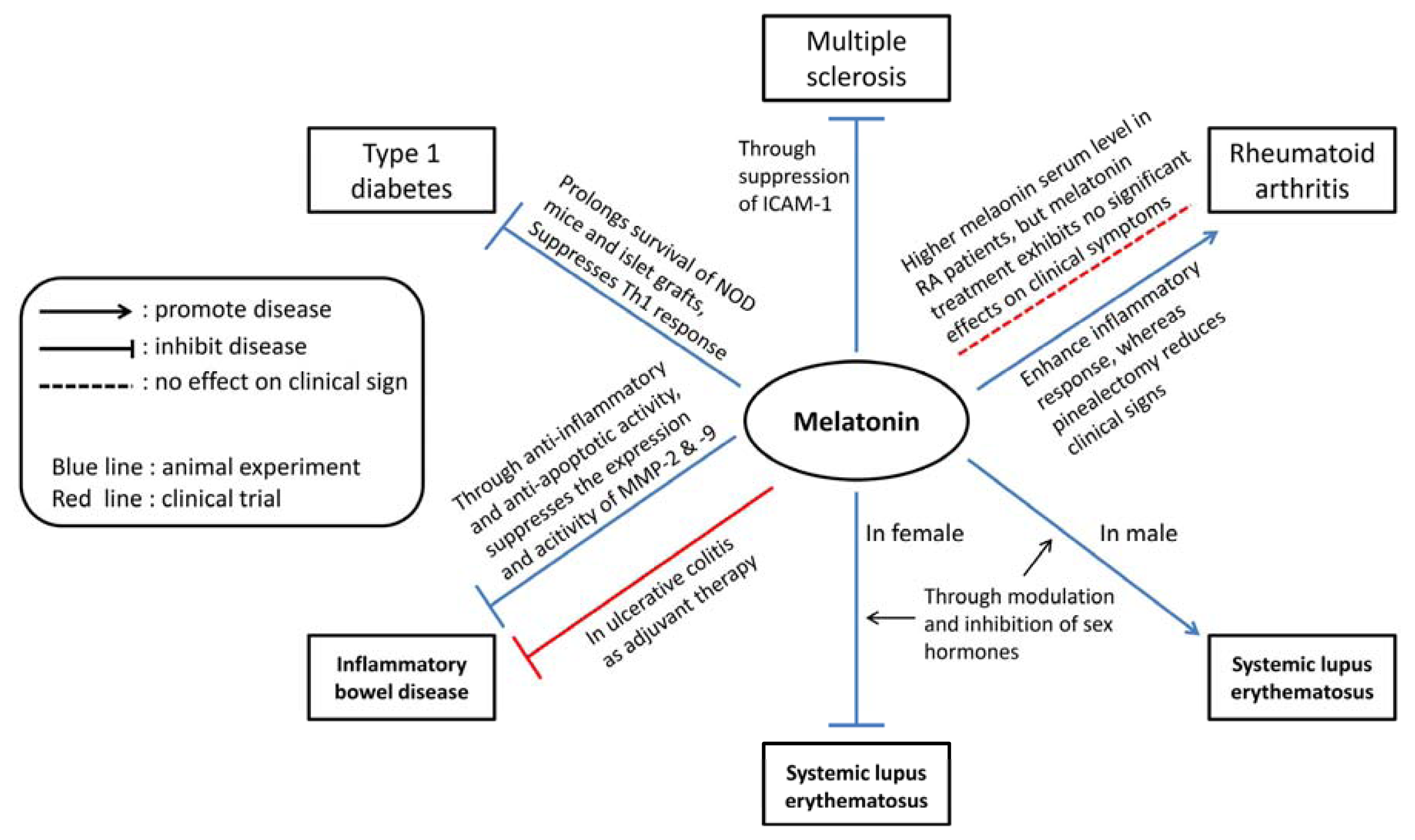{kind=link}
| Clinical diseases | Animal models | Melatonin treatment | Results | Reference |
|---|---|---|---|---|
| Multiple sclerosis | EAE | Receptor antagonist (luzindole) | Suppresses the onset of EAE | [76] |
| EAE | 5 mg/kg, orally | Reduces the incidence and severity of clinical signs | [75] | |
| Systemic lupus erythematosus | MRL/MpJ-lprFas mice | 30 mg/kg, dw | Ameliorates disease in females, whereas exacerbates disease in males | [84] |
| MRL/MpJ-lprFas mice | 30 mg/kg, dw, combined with testosterone in females, with estradiol in males | Decrease in total serum IgG, IgM, anti-ddsDNA, and anti-CII autoantibodies titers | [85] | |
| pristane-induced lupus in Balb/c mice | 0.01, 0.1, 1.0 mg/kg, intragastric | Display a beneficial effect on disease | [86] | |
| Membranous nephritis in Balb/c mice | 20 mg/kg, sc. | Suppresses the pathological injury of glomeruli and deposition of immune complexes | [87] | |
| Rheumatoid arthritis | CIA in DBA/1 mice | Constant darkness | Exaggerates severity and chronicity of arthritis | [94] |
| CIA in DBA/1 mice | 1 mg/kg, sc. | Increase arthritis severity | [95] | |
| CIA in DBA/1 and NFR/N mice | Pinealectomy | Ameliorates arthritis in both strain | [96] | |
| AA in SD rats | 1, 10, 100 μg/kg, intragastric | Ameliorates arthritis and inhibits inflammatory response | [97] | |
| CIA in Wistar rats | 30 μg/mouse, sc. | No significant effect in hitopathologic features, increases the levels of IL-1β and IL-6 in serum | [99] | |
| CIA in DBA/1 mice | 10 mg/kg, i.p. | Increases paw thickness and joint destruction | [98] | |
| Type 1 diabetes mellitus | NOD mice | pinealectomy | Promotes disease onset | [115] |
| NOD mice | 4 mg/kg, sc. | Protects from the development of disease | [115] | |
| NOD mice | 200 mg/kg, sc. | Prolongs islet grafts survival in NOD recipient | [122] | |
| Inflammatory bowel disease | DSS-induced colitis in mice | 150 μg/kg, i.p. | Reduces the severity of colitis | [134] |
| DNBS-colitis in SD rats | 15 mg/kg, i.p. | Reduces severity of colitis | [137] | |
| TNBS and acetic acid-colitis in SD rats | 5 and 10 mg/kg | Protects colonic injury in both colitis models | [136] | |
| TNBS-colitis in SD rats | 2.5, 5, 10 mg/kg | Reduces colonic inflammatory injury | [141] | |
| TNBS-colitis in SD rats | 2.5, 5, 10 mg/kg, intracolonic | Attenuates colonic injury | [142] | |
| TNBS-colitis in Wistar-albino rats | 10 mg/kg, i.p. | Decreases colitis scores | [140] | |
| DNBS-colitis in SD rats | 15 mg/kg, i.p. | Attenuates colonic injury | [138] | |
| TNBS-colitis in Wistar rats | acute administration: 0.5, 1, 2 mg/kg, i.p.; chronic administration: 1 and 2 mg/kg, i.p. | Short-term administration protects from colitis while chronic administration aggravates colitis | [143] | |
| Acetic acid-colitis in Wistar rats | 10 mg/kg, i.p. and intracolonic | Protects from colitis induced colonic damage | [135] | |
| DNBS-colitis in SD rats | 15 mg/kg, i.p. | Reduces colonic injury | [139] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lin, G.-J.; Huang, S.-H.; Chen, S.-J.; Wang, C.-H.; Chang, D.-M.; Sytwu, H.-K. Modulation by Melatonin of the Pathogenesis of Inflammatory Autoimmune Diseases. Int. J. Mol. Sci. 2013, 14, 11742-11766. https://doi.org/10.3390/ijms140611742
Lin G-J, Huang S-H, Chen S-J, Wang C-H, Chang D-M, Sytwu H-K. Modulation by Melatonin of the Pathogenesis of Inflammatory Autoimmune Diseases. International Journal of Molecular Sciences. 2013; 14(6):11742-11766. https://doi.org/10.3390/ijms140611742
Chicago/Turabian StyleLin, Gu-Jiun, Shing-Hwa Huang, Shyi-Jou Chen, Chih-Hung Wang, Deh-Ming Chang, and Huey-Kang Sytwu. 2013. "Modulation by Melatonin of the Pathogenesis of Inflammatory Autoimmune Diseases" International Journal of Molecular Sciences 14, no. 6: 11742-11766. https://doi.org/10.3390/ijms140611742
APA StyleLin, G.-J., Huang, S.-H., Chen, S.-J., Wang, C.-H., Chang, D.-M., & Sytwu, H.-K. (2013). Modulation by Melatonin of the Pathogenesis of Inflammatory Autoimmune Diseases. International Journal of Molecular Sciences, 14(6), 11742-11766. https://doi.org/10.3390/ijms140611742