Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome

 ,
,
Abstract
:1. Introduction
2. Results
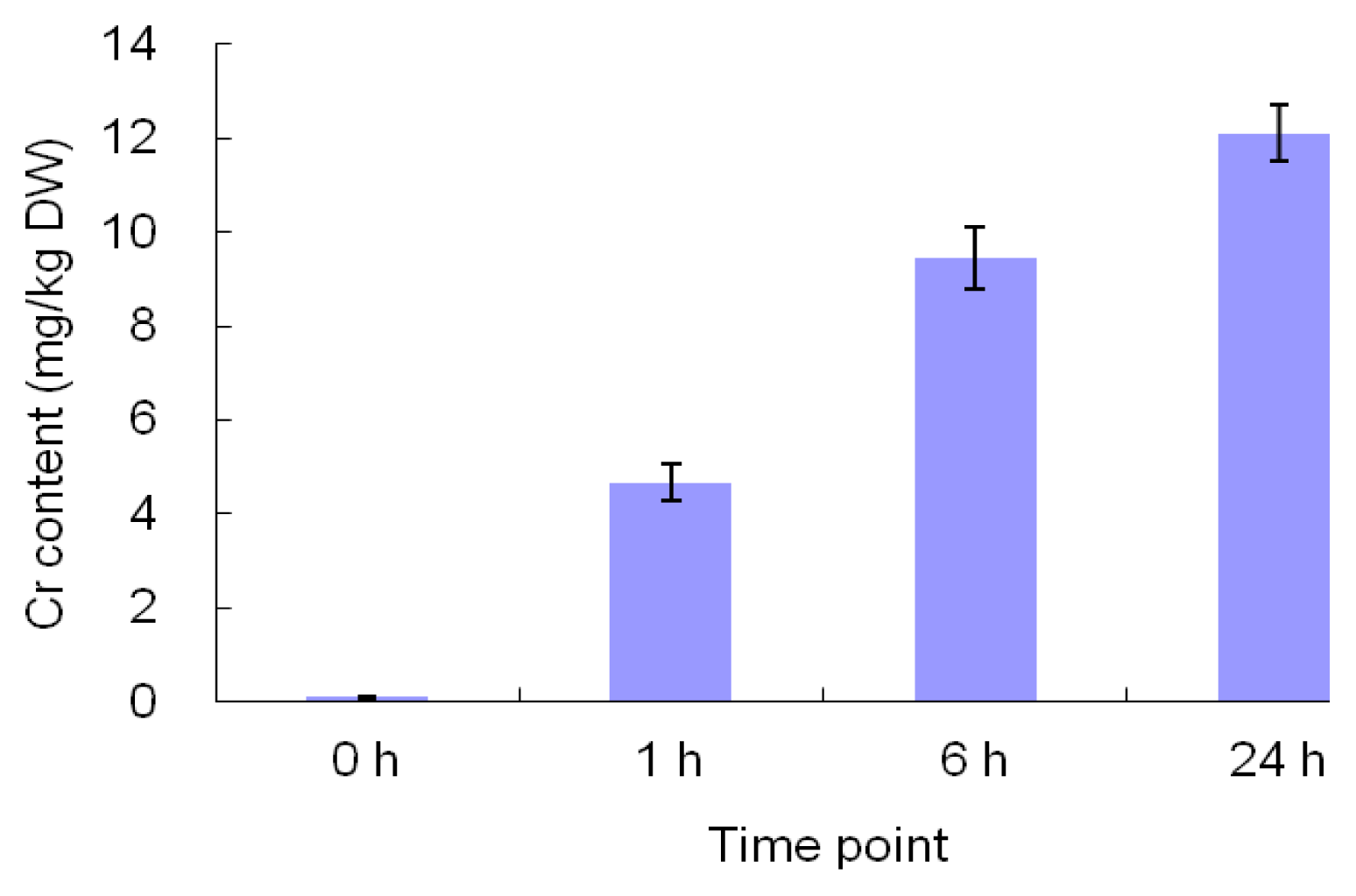
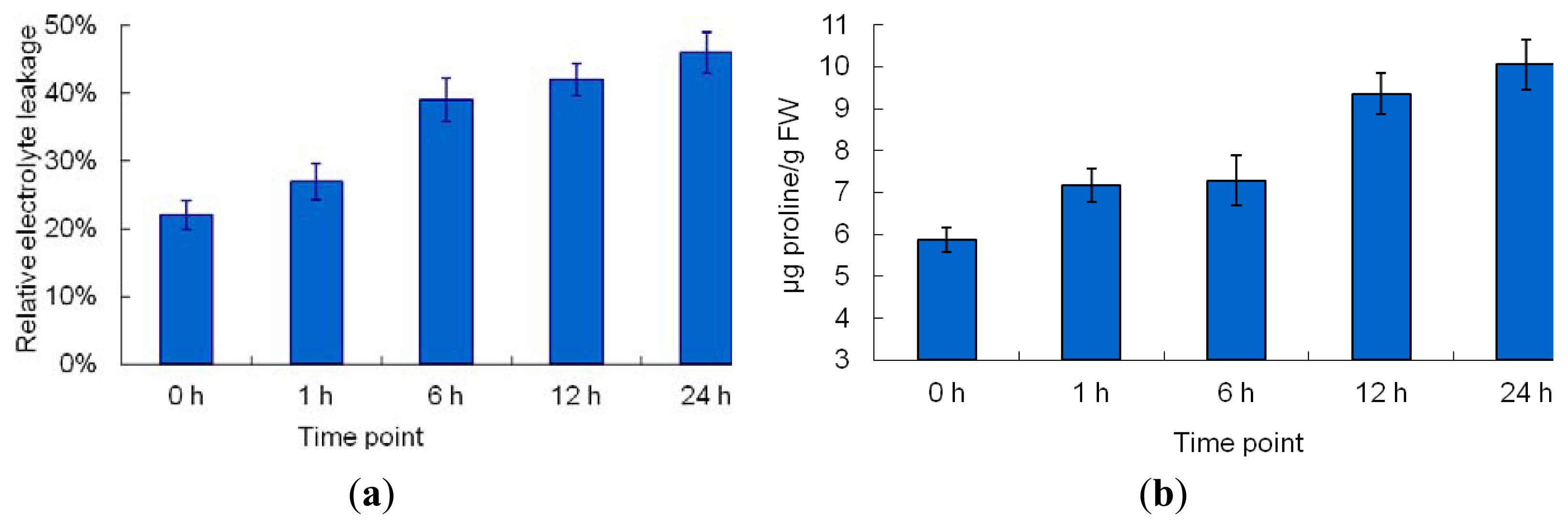
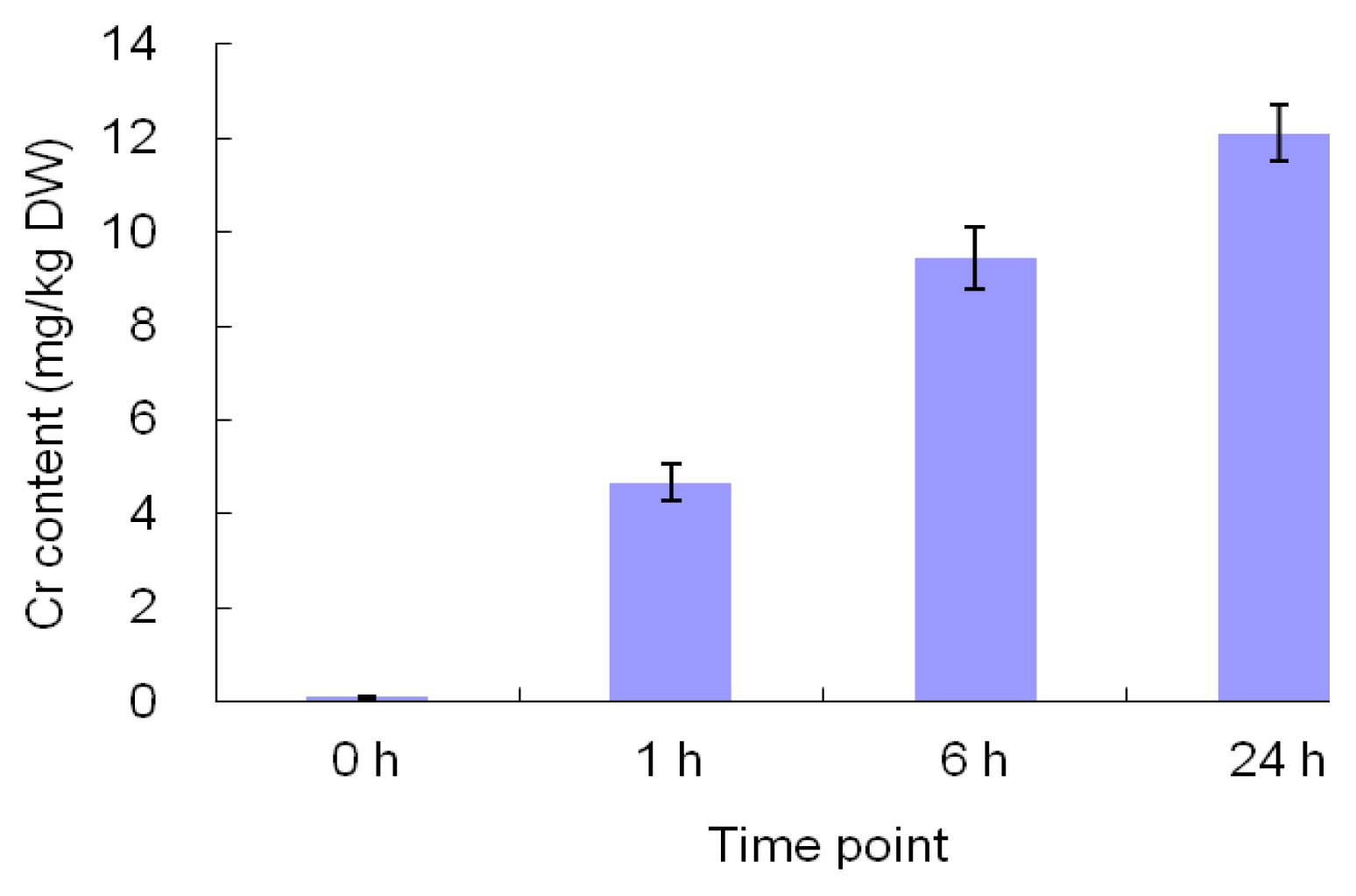
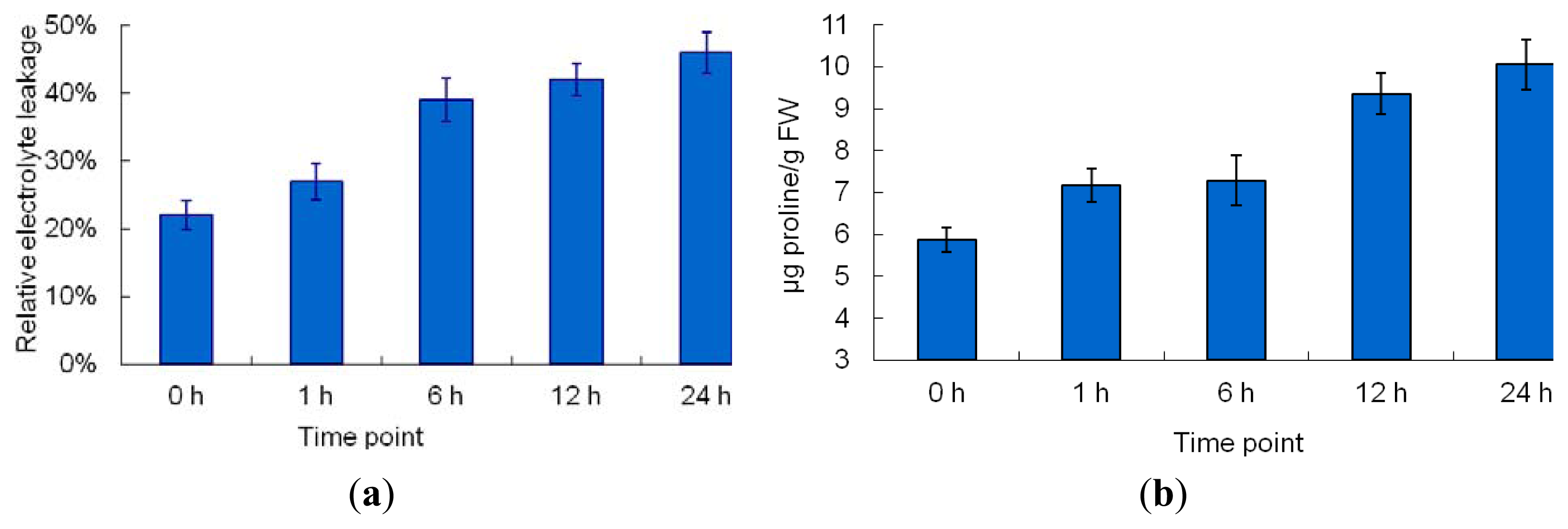
2.1. The Morphological and Physiological Responses in Maize Leaves Induced by Cr Treatment
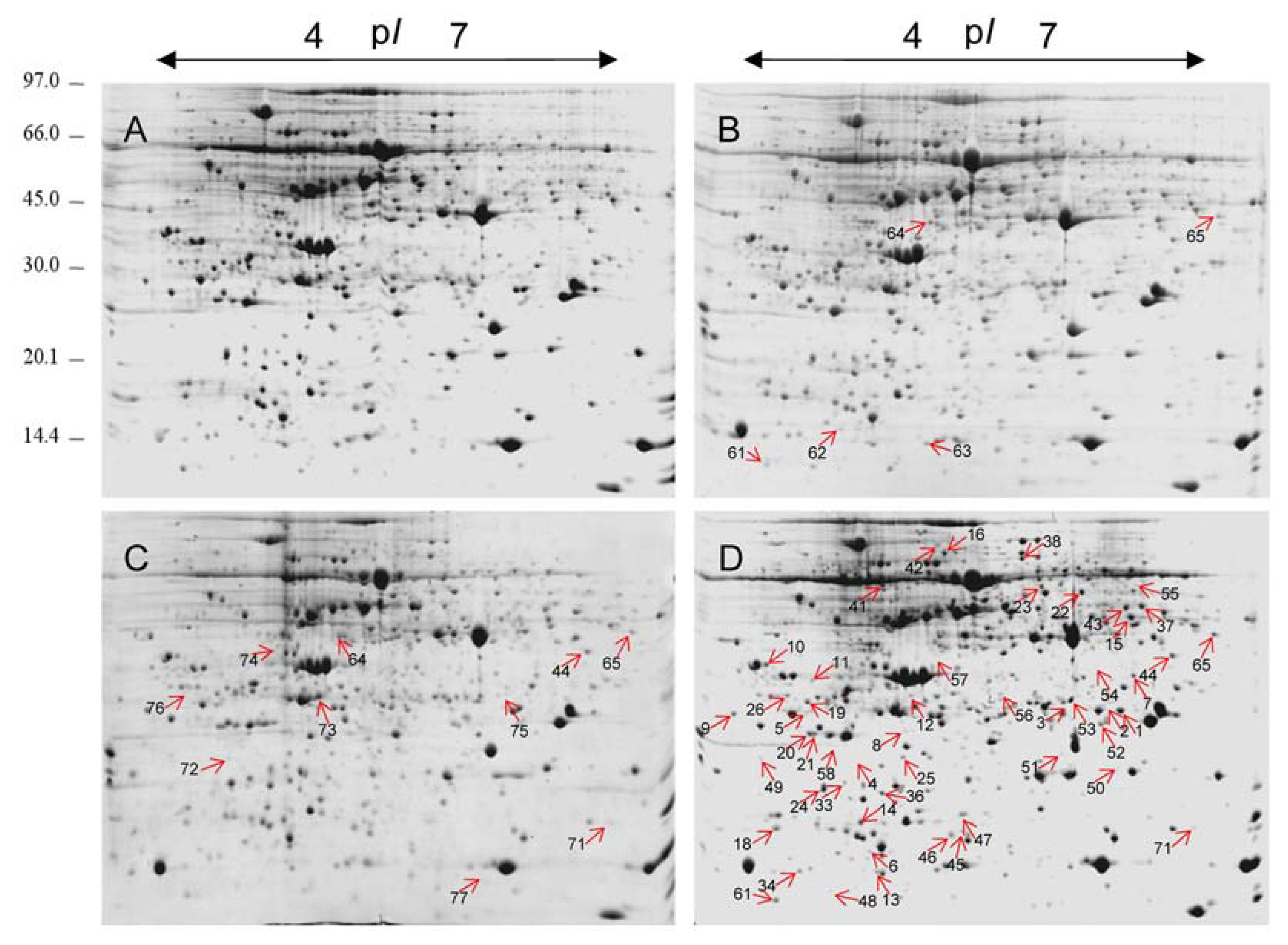
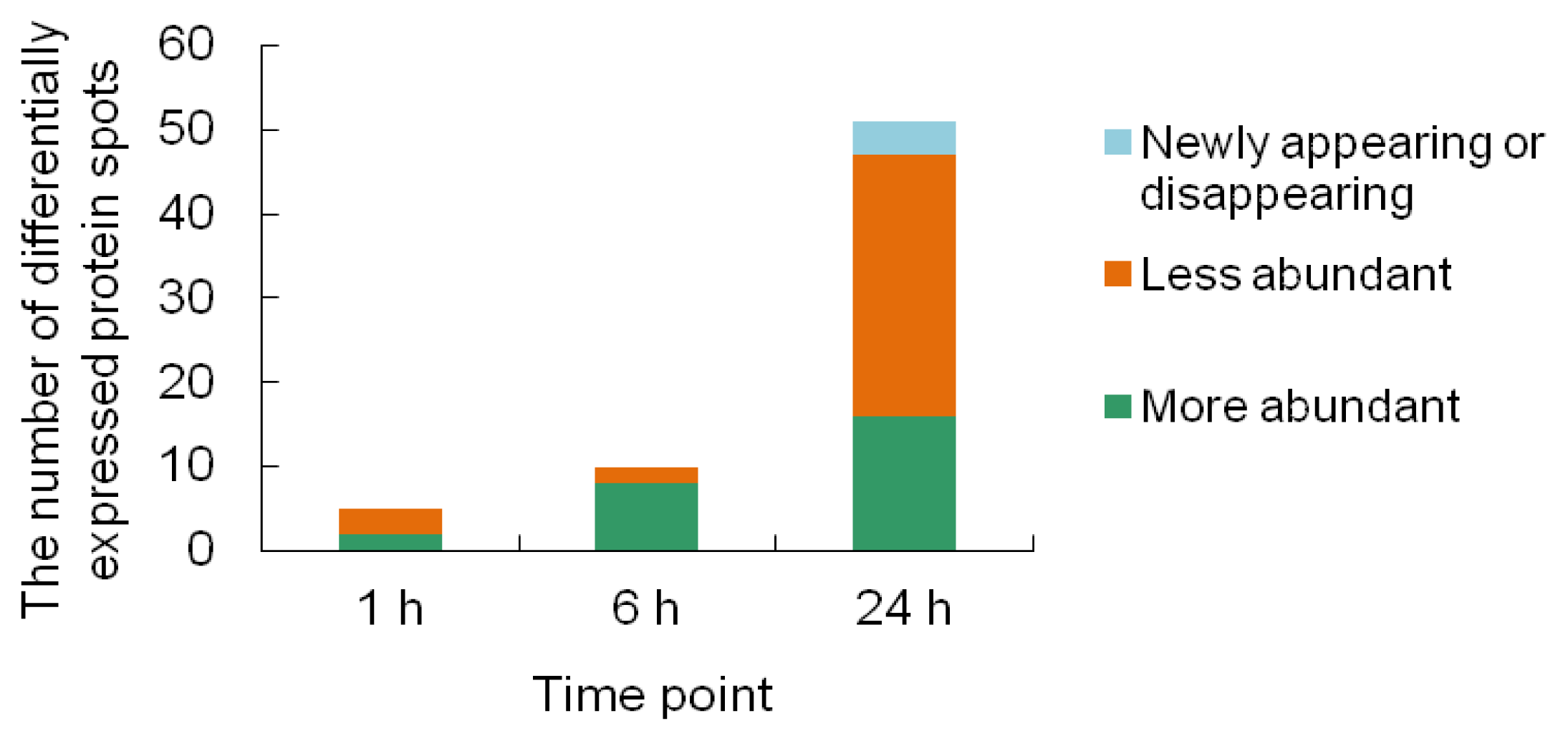
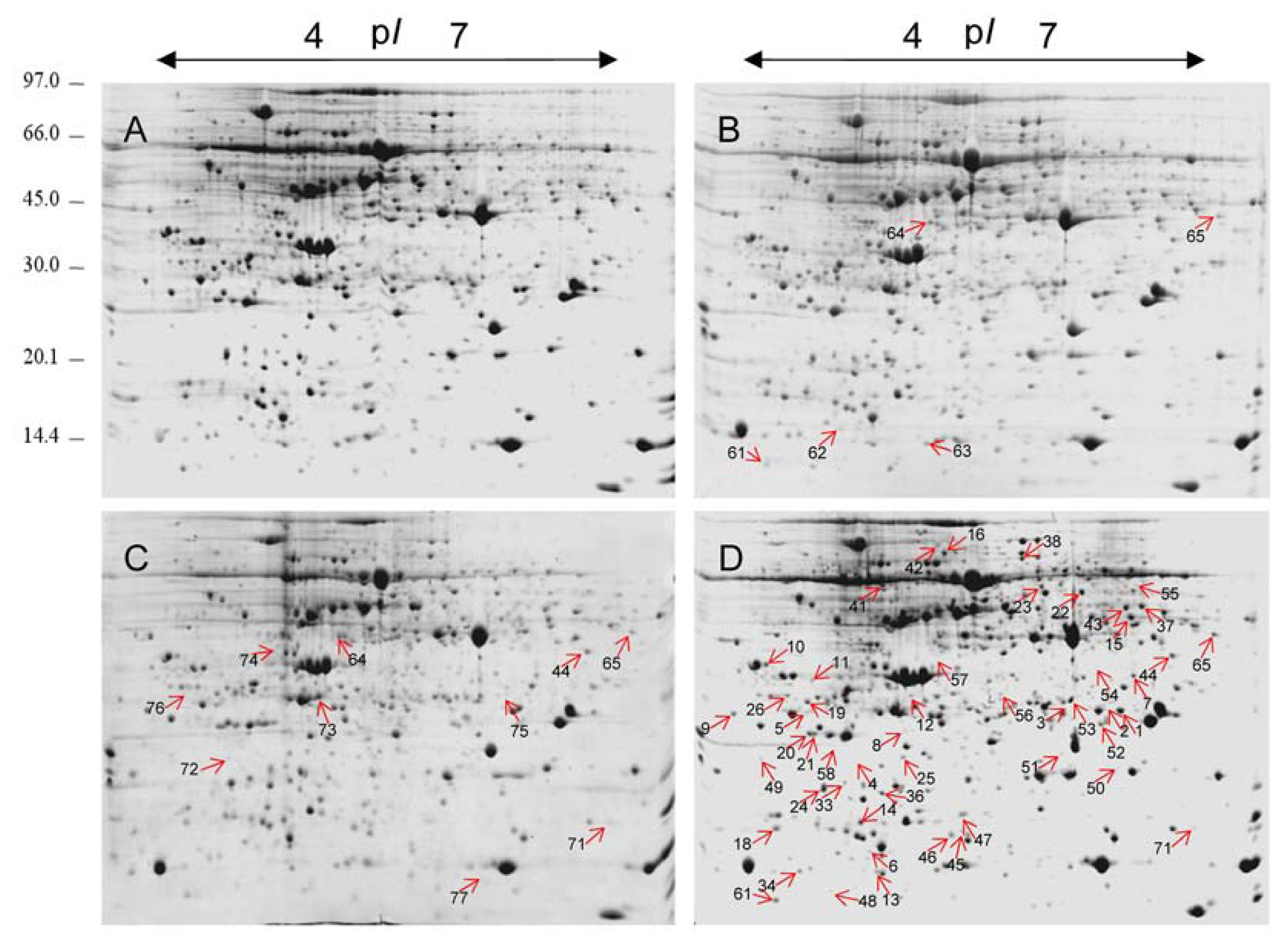
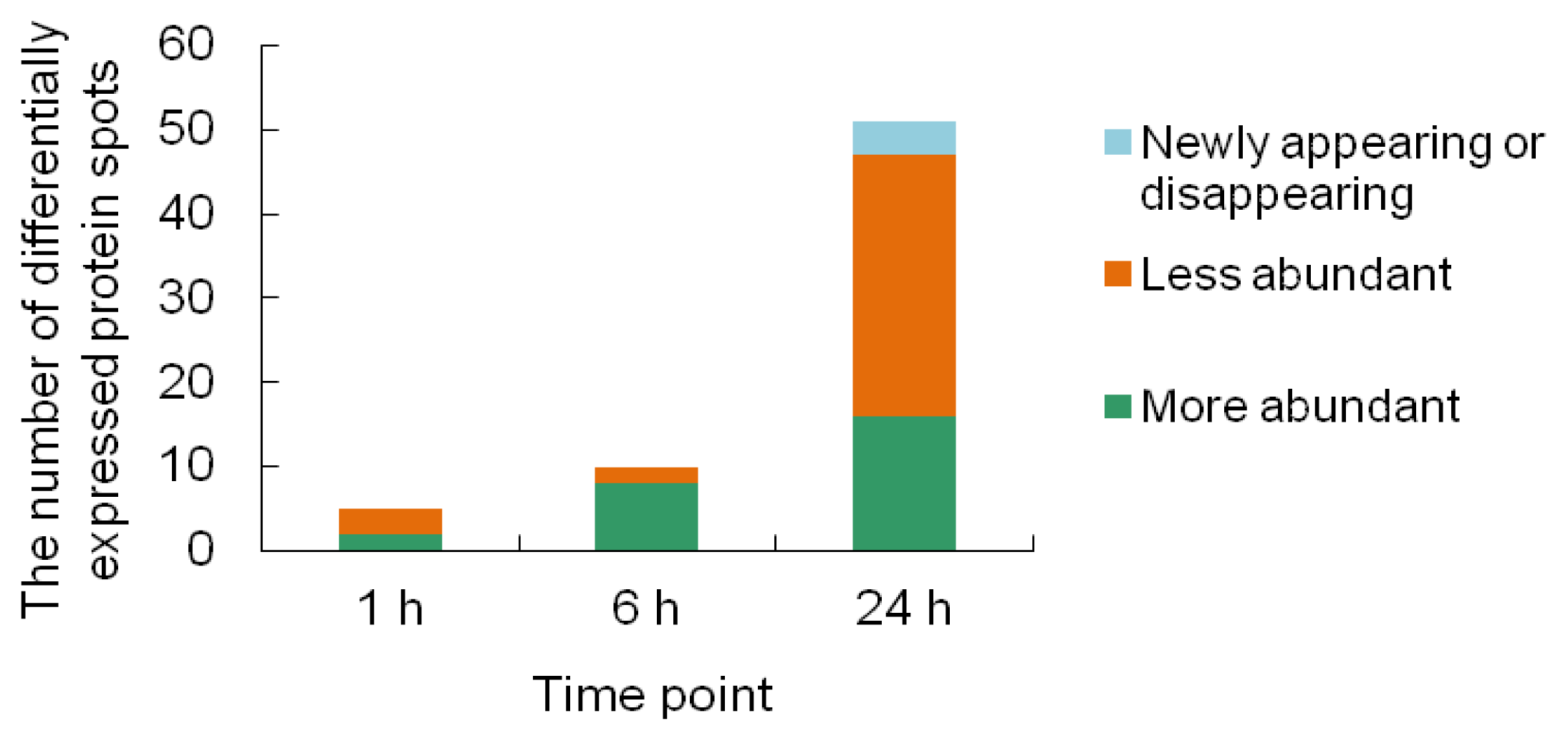
2.2. 2-DE Analysis of Cr-Treated Maize Leaf Proteins
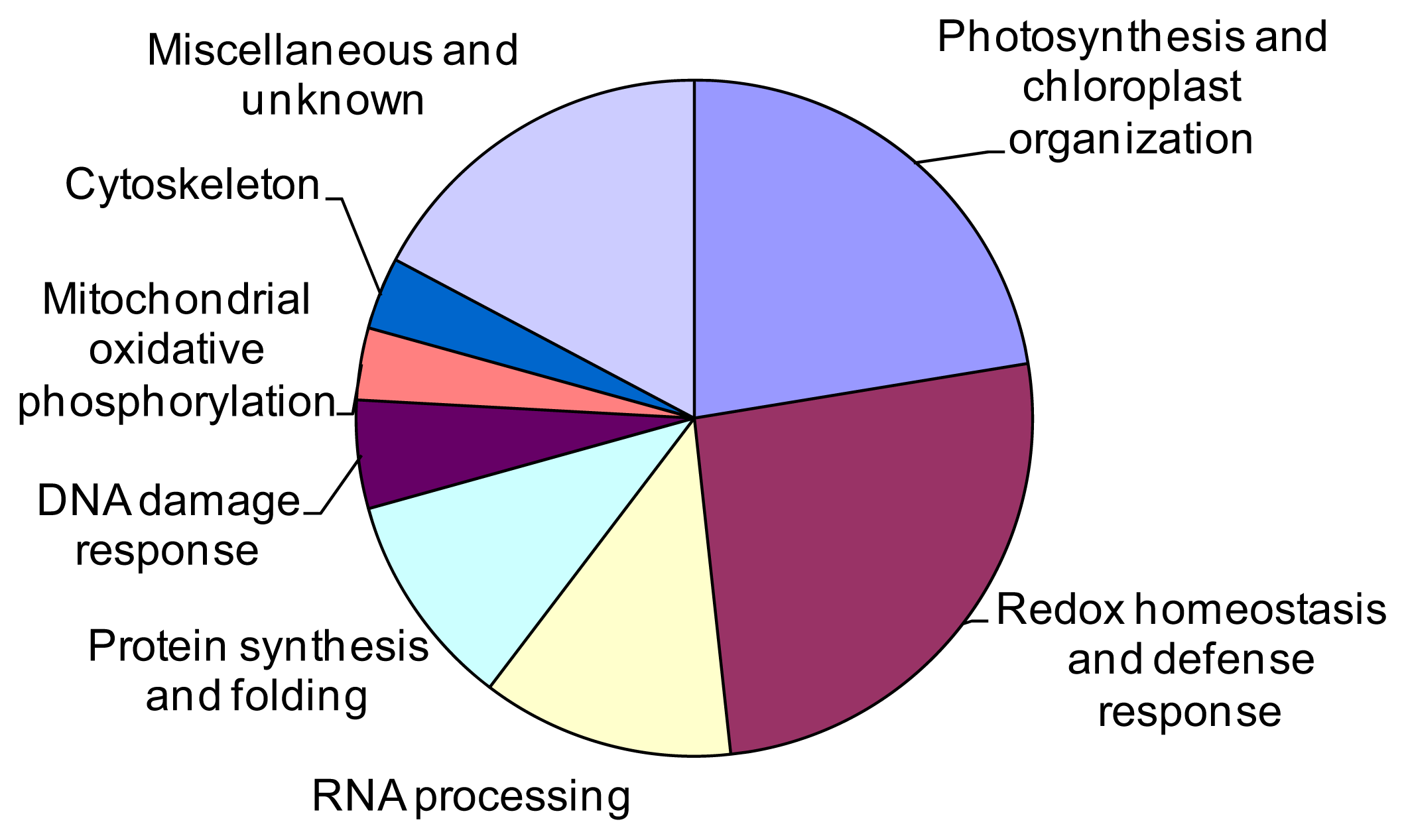
2.3. Identification of the Differentially Expressed Proteins
3. Discussion
3.1. Photosynthesis and Chloroplast Organization
3.2. Defense Response and ROS Detoxification
3.3. Post-Transcriptional Processing of RNA
3.4. Protein Synthesis and Folding
3.5. DNA Damage Response
3.6. Cytoskeleton
3.7. Miscellaneous and Unknown Proteins
4. Experimental Section
4.1. Plant Growth and Chromium Treatment
4.2. Determination of Cr Accumulation
4.3. Physiological Parameter Measurement
4.4. Protein Extraction and Two-Dimensional Electrophoresis
4.5. In-Gel Digestion and Mass Spectrometry Analysis
5. Conclusions
Supplementary Information
ijms-14-11125-s001.pdfAcknowledgments
Conflict of Interest
References
- Pilon-Smits, E. Phytoremediation. Annu. Rev. Plant Biol 2005, 56, 15–39. [Google Scholar]
- Faisal, M.; Hasnain, S. Chromate resistant Bacillus cereus augments sunflower growth by reducing toxicity Cr(VI). J. Plant Biol 2005, 48, 187–194. [Google Scholar]
- Gbaruko, B.C.; Friday, O.U. Bioaccumulation of heavy metals in some fauna and flora. Int. J. Environ. Sci. Technol 2007, 4, 197–202. [Google Scholar]
- Scoccianti, V.; Crinelli, R.; Tirillini, B.; Mancinelli, V.; Speranza, A. Uptake and toxicity of Cr (III) in celery seedlings. Chemosphere 2006, 64, 1695–1703. [Google Scholar]
- Gao, Y.; Xia, J. Chromium contamination accident in China: Viewing environment policy of China. Environ. Sci. Technol 2011, 45, 8605–8606. [Google Scholar]
- Wang, Y.T.; Xiao, C. Factors affecting hexavalent chromium reduction in pure cultures of bacteria. Water Res 1995, 29, 2467–2474. [Google Scholar]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar]
- Manara, A. Plant Responses to Heavy Metal Toxicity. In Plants and Heavy Metals; Furini, A., Ed.; Springer: Berlin, Germany, 2012; pp. 27–53. [Google Scholar]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int 2005, 31, 739–753. [Google Scholar]
- Shanker, A.K.; Djanaguiraman, M.; Venkateswarlu, B. Chromium interactions in plants: Current status and future strategies. Metallomics 2009, 1, 375–383. [Google Scholar]
- Quaggiotti, S.; Barcaccia, G.; Schiavon, M.; Nicolé, S.; Galla, G.; Rossignolo, V.; Soattin, M.; Malagoli, M. Phytoremediation of chromium using Salix species: Cloning ESTs and candidate genes involved in the Cr response. Gene 2007, 402, 68–80. [Google Scholar]
- Dubey, S.; Misra, P.; Dwivedi, S.; Chatterjee, S.; Bag, S.; Mantri, S.; Asif, M.; Rai, A.; Kumar, S.; Shri, M.; et al. Transcriptomic and metabolomic shifts in rice roots in response to Cr(VI) stress. BMC Genomics 2010, 11, 648. [Google Scholar]
- Sharmin, S.A.; Alam, I.; Kim, K.H.; Kim, Y.G.; Kim, P.J.; Bahk, J.D.; Lee, B.H. Chromium-induced physiological and proteomic alterations in roots of Miscanthus sinensis. Plant Sci 2012, 187, 113–126. [Google Scholar]
- Sharma, D.C.; Sharma, C.P.; Tripathi, R.D. Phytotoxic lesions of chromium in maize. Chemosphere 2003, 51, 63–68. [Google Scholar]
- Labra, M.; Gianazza, E.; Waitt, R.; Eberini, I.; Sozzi, A.; Regondi, S.; Grassi, F.; Agradi, E. Zea mays L. protein changes in response to potassium dichromate treatments. Chemosphere 2006, 62, 1234–1244. [Google Scholar]
- Prinsi, B.; Negri, A.S.; Pesaresi, P.; Cocucci, M.; Espen, L. Evaluation of protein pattern changes in roots and leaves of Zea mays plants in response to nitrate availability by two-dimensional gel electrophoresis analysis. BMC Plant Biol 2009, 9, 113. [Google Scholar]
- The UniProt Knowledgebase (UniProtKB). Available online: http://www.ebi.uniprot.org/ (accessed on 8 January 2013).
- The Arabidopsis Information Resource (TAIR). Available online: http://www.arabidopsis.org/ (accessed on 8 January 2013).
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Qian, W.; Yu, C.; Qin, H.; Liu, X.; Zhang, A.; Johansen, I.E.; Wang, D. Molecular and functional analysis of phosphomannomutase (PMM) from higher plants and genetic evidence for the involvement of PMM in ascorbic acid biosynthesis in Arabidopsis and Nicotiana benthamiana. Plant J 2007, 49, 399–413. [Google Scholar]
- Hou, F.Y.; Huang, J.; Yu, S.L.; Zhang, H.S. The 6-phosphogluconate dehydrogenase genes are responsive to abiotic stresses in rice. J. Integr. Plant Biol 2007, 49, 655–663. [Google Scholar]
- Gao, F.; Zhou, Y.; Zhu, W.; Li, X.; Fan, L.; Zhang, G. Proteomic analysis of cold stress-responsive proteins in Thellungiella rosette leaves. Planta 2009, 230, 1033–1046. [Google Scholar]
- Rocha, M.; Sodek, L.; Licausi, F.; Hameed, M.W.; Dornelas, M.C.; van Dongen, J.T. Analysis of alanine aminotransferase in various organs of soybean (Glycine max) and in dependence of different nitrogen fertilisers during hypoxic stress. Amino Acids 2010, 39, 1043–1053. [Google Scholar]
- Vieira Dos Santos, C.; Cuiné, S.; Rouhier, N.; Rey, P. The Arabidopsis plastidic methionine sulfoxide reductase B proteins. Sequence and activity characteristics, comparison of the expression with plastidic methionine sulfoxide reductase A, and induction by photooxidative stress. Plant Physiol 2005, 138, 909–922. [Google Scholar]
- Dietz, K.J.; Jacob, S.; Oelze, M.L.; Laxa, M.; Tognetti, V.; de Miranda, S.M.; Baier, M.; Finkemeier, I. The function of peroxiredoxins in plant organelle redox metabolism. J. Exp. Bot 2006, 57, 1697–1709. [Google Scholar]
- Sachetto-Martins, G.; Franco, L.O.; de Oliveira, D.E. Plant glycine-rich proteins: A family or just proteins with a common motif? Biochim. Biophys. Acta 2000, 1492, 1–14. [Google Scholar]
- Wen, R.; Newton, L.; Li, G.; Wang, H.; Xiao, W. Arabidopsis thaliana UBC13: Implication of error-free DNA damage tolerance and Lys63-linked polyubiquitylation in plants. Plant Mol. Biol 2006, 61, 241–253. [Google Scholar]
- Sarry, J.E.; Kuhn, L.; Ducruix, C.; Lafaye, A.; Junot, C.; Hugouvieux, V.; Jourdain, A.; Bastien, O.; Fievet, J.B.; Vailhen, D.; et al. The early responses of Arabidopsis thaliana cells to cadmium exposure explored by protein and metabolite profiling analyses. Proteomics 2006, 6, 2180–2198. [Google Scholar]
- Kato, Y.; Sun, X.; Zhang, L.; Sakamoto, W. Cooperative D1 degradation in the photosystem II repair mediated by chloroplastic proteases in Arabidopsis. Plant Physiol 2012, 159, 1428–1439. [Google Scholar]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci 2006, 11, 15–19. [Google Scholar]
- Rapala-Kozik, M.; Wolak, N.; Kujda, M.; Banas, A.K. The upregulation of thiamine (vitamin B1) biosynthesis in Arabidopsis thaliana seedlings under salt and osmotic stress conditions is mediated by abscisic acid at the early stages of this stress response. BMC Plant Biol 2012, 12, 2. [Google Scholar]
- Davies, F.T.; Puryear, J.D.; Newton, R.J.; Egilla, J.N.; Grossi, J.A.S. Mycorrhizal fungi increase chromium uptake by sunflower plants: Influence on tissue mineral concentration, growth, and gas exchange. J. Plant Nutr 2002, 25, 2389–2407. [Google Scholar]
- Zhang, L.; Kato, Y.; Otters, S.; Vothknecht, U.C.; Sakamoto, W. Essential role of VIPP1 in chloroplast envelope maintenance in Arabidopsis. Plant Cell 2012, 24, 3695–3707. [Google Scholar]
- Lange, P.R.; Geserick, C.; Tischendorf, G.; Zrenner, R. Functions of chloroplastic adenylate kinases in Arabidopsis. Plant Physiol 2008, 146, 492–504. [Google Scholar]
- Gong, P.; Zhang, J.; Li, H.; Yang, C.; Zhang, C.; Zhang, X.; Khurram, Z.; Zhang, Y.; Wang, T.; Fei, Z.; Ye, Z. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. J. Exp. Bot 2010, 61, 3563–3575. [Google Scholar]
- Wang, L.; Ouyang, M.; Li, Q.; Zou, M.; Guo, J.; Ma, J.; Lu, C.; Zhang, L. The Arabidopsis chloroplast ribosome recycling factor is essential for embryogenesis and chloroplast biogenesis. Plant Mol. Biol 2010, 74, 47–59. [Google Scholar]
- Keunen, E.; Remans, T.; Bohler, S.; Vangronsveld, J.; Cuypers, A. Metal-induced oxidative stress and pant mitochondria. Int. J. Mol. Sci 2011, 12, 6894–6918. [Google Scholar]
- Dixit, V.; Pandey, V.; Shyam, R.V. Chromium ions inactivate electron transport and enhance superoxide generation in vivo in pea (Pisum sativum L. cv. Azad) root mitochondria. Plant Cell Environ 2002, 25, 687–693. [Google Scholar]
- Koussevitzky, S.; Suzuki, N.; Huntington, S.; Armijo, L.; Sha, W.; Cortes, D.; Shulaev, V.; Mittler, R. Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination. J. Biol. Chem 2008, 283, 34197–34203. [Google Scholar]
- Dowdle, J.; Ishikawa, T.; Gatzek, S.; Rolinski, S.; Smirnoff, N. Two genes in Arabidopsis thaliana encoding GDP-l-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability. Plant J 2007, 52, 673–689. [Google Scholar]
- Xiong, Y.; DeFraia, C.; Williams, D.; Zhang, X.; Mou, Z. Characterization of Arabidopsis 6-phosphogluconolactonase T-DNA insertion mutants reveals an essential role for the oxidative section of the plastidic pentose phosphate pathway in plant growth and development. Plant Cell Physiol 2009, 50, 1277–1291. [Google Scholar]
- Belenghi, B.; Acconcia, F.; Trovato, M.; Perazzolli, M.; Bocedi, A.; Polticelli, F.; Ascenzi, P.; Delledonne, M. AtCYS1, a cystatin from Arabidopsis thaliana, suppresses hypersensitive cell death. Eur. J. Biochem 2003, 270, 2593–2604. [Google Scholar]
- Zhang, X.; Liu, S.; Takano, T. Two cysteine proteinase inhibitors from Arabidopsis thaliana, AtCYSa and AtCYSb, increasing the salt, drought, oxidation and cold tolerance. Plant Mol. Biol 2008, 68, 131–143. [Google Scholar]
- Wei, Z.M.; Laby, R.J.; Zumoff, C.H.; Bauer, D.W.; He, S.Y.; Collmer, A.; Beer, S.V. Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science 1992, 257, 85–88. [Google Scholar]
- Mazzucotelli, E.; Mastrangelo, A.M.; Crosatti, C.; Guerra, D.; Stanca, A.M.; Cattivelli, L. Abiotic stress response in plants: When post-transcriptional and post-translational regulations control transcription. Plant Sci 2008, 174, 420–431. [Google Scholar]
- Burd, C.G.; Dreyfuss, G. Conserved structures and diversity of functions of RNA-binding proteins. Science 1994, 265, 615–621. [Google Scholar]
- Gao, F. Minzu University of China, Beijing, China; Unpublished work; 2008. [Google Scholar]
- Kim, J.S.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Goh, C.H.; Woo, Y.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding protein 7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J 2008, 55, 455–466. [Google Scholar]
- Kwak, K.J.; Park, S.J.; Han, J.H.; Kim, M.K.; Oh, S.H.; Han, Y.S.; Kang, H. Structural determinants crucial to the RNA chaperone activity of glycine-rich RNA-binding proteins 4 and 7 in Arabidopsis thaliana during the cold adaptation process. J. Exp. Bot 2011, 62, 4003–4011. [Google Scholar]
- Kwak, K.J.; Kim, Y.O.; Kang, H. Characterization of transgenic Arabidopsis plants overexpressing GR-RBP4 under high salinity, dehydration, or cold stress. J. Exp. Bot 2005, 56, 3007–3016. [Google Scholar]
- Fu, Z.Q.; Guo, M.; Jeong, B.R.; Tian, F.; Elthon, T.E.; Cerny, R.L.; Staiger, D.; Alfano, J.R. A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 2007, 447, 284–288. [Google Scholar]
- Gerashchenko, M.V.; Lobanov, A.V.; Gladyshev, V.N. Genome-wide ribosome profiling reveals complex translational regulation in response to oxidative stress. Proc. Natl. Acad. Sci. USA 2012, 109, 17394–17399. [Google Scholar]
- Salnikow, K.; Zhitkovich, A. Genetic and epigenetic mechanisms in metal carcinogenesis and cocarcinogenesis: Nickel, arsenic, and chromium. Chem. Res. Toxicol 2008, 21, 28–44. [Google Scholar]
- Nickens, K.P.; Patierno, S.R.; Ceryak, S. Chromium genotoxicity: A double-edged sword. Chem. Biol. Interact 2010, 188, 276–288. [Google Scholar]
- Machado, C.R.; Praekelt, U.M.; de Oliveira, R.C.; Barbosa, A.C.; Byrne, K.L.; Meacock, P.A.; Menck, C.F. Dual role for the yeast THI4 gene in thiamine biosynthesis and DNA damage tolerance. J. Mol. Biol 1997, 273, 114–121. [Google Scholar]
- Ruzicka, D.R.; Kandasamy, M.K.; McKinney, E.C.; Burgos-Rivera, B.; Meagher, R.B. The ancient subclasses of Arabidopsis Actin Depolymerizing Factor genes exhibit novel and differential expression. Plant J 2007, 52, 460–672. [Google Scholar]
- Jiang, C.J.; Weeds, A.G.; Hussey, P.J. The maize actin-depolymerizing factor, ZmADF3, redistributes to the growing tip of elongating root hairs and can be induced to translocate into the nucleus with actin. Plant J 1997, 12, 1035–1043. [Google Scholar]
- Fan, T.; Zhai, H.; Shi, W.; Wang, J.; Jia, H.; Xiang, Y.; An, L. Overexpression of profilin 3 affects cell elongation and F-actin organization in Arabidopsis thaliana. Plant Cell. Rep 2013, 32, 149–160. [Google Scholar]
- Taketani, S.; Adachi, Y.; Kohno, H.; Ikehara, S.; Tokunaga, R.; Ishii, T. Molecular characterization of a newly identified heme-binding protein induced during differentiation of urine erythroleukemia cells. J. Biol. Chem 1998, 273, 31388–31394. [Google Scholar]
- Song, L.; Shi, Q.M.; Yang, X.H.; Xu, Z.H.; Xue, H.W. Membrane steroid-binding protein 1 (MSBP1) negatively regulates brassinosteroid signaling by enhancing the endocytosis of BAK1. Cell. Res 2009, 19, 864–876. [Google Scholar]
- Divi, U.K.; Rahman, T.; Krishna, P. Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol 2010, 10, 151. [Google Scholar]
- Majeran, W.; Friso, G.; Ponnala, L.; Connolly, B.; Huang, M.; Reidel, E.; Zhang, C.; Asakura, Y.; Bhuiyan, N.H.; Sun, Q.; Turgeon, R.; van Wijk, K.J. Structural and metabolic transitions of C4 leaf development and differentiation defined by microscopy and quantitative proteomics in maize. Plant Cell 2010, 22, 3509–3542. [Google Scholar]
- Yan, S.P.; Zhang, Q.Y.; Tang, Z.C.; Su, W.A.; Sun, W.N. Comparative proteomic analysis provides new insights into chilling stress responses in rice. Mol. Cell. Proteomics 2006, 5, 484–496. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Tears, I.D. Rapid determination of free proline in water stress studies. Plant Soil 1975, 39, 205–207. [Google Scholar]
- Katayama, H.; Nagasu, T.; Oda, Y. Improvement of in-gel digestion protocol for peptide mass fingerprinting by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom 2001, 15, 1416–1421. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot ID | Accession number a | Protein name and plant species | Score | Theor. Mr/pI b | Exp. Mr/pI c |
|---|---|---|---|---|---|
| Photosynthesis and chloroplast organization | |||||
| 23 | gi|226508728 | uncharacterized protein LOC100275158 [Zea mays] | 414 | 51.9/6.12 | 53/5.9 |
| 44 | gi|13096165 | Chain A, Crystal Structure Of The Complex Between Ferredoxin And Ferredoxin-Nadp+ Reductase [Zea mays] | 263 | 35.6/7.01 | 35/6.6 |
| 50 | gi|194697374 | unknown [Zea mays] | 337 | 23.1/9.07 | 23/6.2 |
| 26 | gi|226503027 | uncharacterized protein LOC100272863 [Zea mays] | 148 | 28.9/4.85 | 29/4.7 |
| 9 | gi|195613650 | ATP synthase delta chain [Zea mays] | 335 | 26.7/4.73 | 27/4.2 |
| 46 | gi|77554379 | ATP synthase epsilon chain, putative [Oryza sativa] | 151 | 15.3/5.46 | 16/5.4 |
| 38 | gi|30575690 | NADP-malic enzyme [Zea mays] | 373 | 70.4/6.2 | 70/5.8 |
| 73 | gi|1170606 | Adenylate kinase, chloroplastic [Zea mays] | 523 | 24.9/4.95 | 25/5.2 |
| 16 | gi|187830110 | filamentation temperature-sensitive H 2B [Zea mays] | 335 | 72.6/5.69 | 73/5.4 |
| 7 | gi|195622012 | membrane-associated 30 kDa protein [Zea mays] | 344 | 35.1/9.5 | 35/6.3 |
| 1 | gi|226497262 | ribosome recycling factor [Zea mays] | 373 | 29.3/9.22 | 29/6.3 |
| 2 | gi|226497262 | ribosome recycling factor [Zea mays] | 373 | 29.3/9.22 | 29/6.2 |
| 4 | gi|226508836 | uncharacterized protein LOC100277322 [Zea mays] | 131 | 23.4/6.31 | 23/4.8 |
| Redox homeostasis and defense response | |||||
| 53 | gi|226504576 | APx1-Cytosolic Ascorbate Peroxidase [Zea mays] | 764 | 27.5/5.65 | 27/5.9 |
| 52 | gi|168624 | manganese superoxide dismutase (SOD-3) [Zea mays] | 407 | 25.6/7.11 | 25/6.1 |
| 75 | gi|226530195 | uncharacterized protein LOC100272867 [Zea mays | 267 | 28.3/5.83 | 28/6.2 |
| 55 | gi|194702230 | unknown [Zea mays] | 363 | 53.1/5.93 | 53/6.2 |
| 12 | gi|414591366 | 6-phosphogluconolactonase isoform 1 [Zea mays] | 371 | 34.8/7.71 | 31/5.2 |
| 21 | gi|195626524 | 2-cys peroxiredoxin BAS1 [Zea mays] | 81 | 28.3/5.81 | 28/4.5 |
| 20 | gi|195626524 | 2-cys peroxiredoxin BAS1 [Zea mays] | 89 | 28.3/5.81 | 28/4.5 |
| 36 | gi|226505300 | LOC100283392 [Zea mays] | 165 | 17.3/4.85 | 18/4.9 |
| 8 | gi|223943539 | unknown [Zea mays] | 309 | 28.1/8.79 | 28/5.2 |
| 48 | gi|195624046 | thioredoxin X [Zea mays] | 213 | 19.3/8.75 | 14/4.8 |
| 63 | gi|66866417 | cysteine proteinase inhibitor [Zea mays subsp. parviglumis] | 133 | 14.8/6.3 | 15/5.4 |
| 56 | gi|293334301 | uncharacterized protein LOC100383635 [Zea mays] | 529 | 29.4/9.57 | 28/5.6 |
| 22 | gi|226492878 | bifunctional 3-phosphoadenosine 5-phosphosulfate synthetase 2 [Zea mays] | 354 | 52.5/8.30 | 52/6.2 |
| 37 | gi|226508814 | aspartate aminotransferase [Zea mays] | 306 | 50.5/8.15 | 50/6.4 |
| 51 | gi|226532399 | peptide methionine sulfoxide reductase [Zea mays] | 775 | 20.8/5.85 | 21/5.9 |
| RNA processing | |||||
| 3 | gi|363543235 | uncharacterized protein LOC100857032 [Zea mays] | 421 | 23.9/5.95 | 24/5.9 |
| 13 | gi|195642478 | glycine-rich RNA-binding protein 2 [Zea mays] | 115 | 15.6/9.00 | 14/4.8 |
| 15 | gi|414884012 | hypothetical protein ZEAMMB73_274910 [Zea mays] | 253 | 42.3/8.14 | 43/6.4 |
| 5 | gi|226502782 | ribonucleoprotein A [Zea mays] | 123 | 28.5/4.83 | 29/4.6 |
| 19 | gi|226502782 | ribonucleoprotein A [Zea mays] | 103 | 28.5/4.83 | 30/4.5 |
| 10 | gi|162463757 | nucleic acid binding protein1 [Zea mays] | 274 | 33.2/4.6 | 33/4.4 |
| 11 | gi|219363077 | uncharacterized protein LOC100217196 [Zea mays] | 284 | 31.5/5.13 | 31/4.6 |
| Protein synthesis and folding | |||||
| 1 | gi|413915954 | 40S ribosomal protein S16, mRNA [Zea mays] | 431 | 59.4/5.28 | 53/5.1 |
| 47 | gi|195604208 | 40S ribosomal protein S12 [Zea mays] | 180 | 15.4/5.33 | 18/5.4 |
| 61 | gi|195647902 | Glu-tRNAGln amidotransferase, C subunit family [Zea mays] | 304 | 15.9/5.76 | 14/4.6 |
| 6 | gi|414585580 | CHL-CPN10 [Zea mays] | 106 | 14.5/6.15 | 15/4.8 |
| 33 | gi|195610950 | 50S ribosomal protein L12-1 [Zea mays] | 147 | 19.1/5.4 | 19/4.9 |
| 24 | gi|195659273 | 50S ribosomal protein L12-1 [Zea mays] | 156 | 19.0/5.71 | 19/4.7 |
| DNA damage response | |||||
| 71 | gi|414881042 | putative ubiquitin-conjugating enzyme family [Zea mays] | 377 | 17.3/6.74 | 17/6.7 |
| 64 | gi|239985534 | thiamine thiazole synthase 2, chloroplastic precursor [Zea mays] | 432 | 37.4/5.59 | 38/5.3 |
| 57 | gi|239985530 | thiamine thiazole synthase 1, chloroplastic precursor [Zea mays] | 279 | 37.3/4.87 | 34/5.3 |
| Cytoskeleton | |||||
| 34 | gi|162461296 | profilin-5 [Zea mays] | 177 | 14.2/4.59 | 14/4.6 |
| 45 | gi|162459533 | actin-depolymerizing factor 3 [Zea mays] | 490 | 16.0/5.46 | 16/5.4 |
| Mitochondrial oxidative phosphorylation | |||||
| 25 | gi|226507194 | ATP synthase D chain, mitochondrial [Zea mays] | 178 | 19.9/5.19 | 23/5.2 |
| 72 | gi|223973939 | unknown [Zea mays] | 246 | 24.3/5.68 | 24/4.8 |
| Miscellaneous and unknown | |||||
| 42 | gi|414882068 | putative alpha-L-arabinofuranosidase family protein [Zea mays] | 303 | 73.0/5.1 | 73/5.2 |
| 76 | gi|195638660 | heme-binding protein 2 [Zea mays] | 448 | 23.8/4.75 | 24/4.5 |
| 54 | gi|226532343 | SOUL heme-binding protein [Zea mays] | 529 | 32.1/9.09 | 33/6.2 |
| 65 | gi|226507242 | uncharacterized protein LOC100274379 [Zea mays] | 776 | 38.8/6.3 | 38/6.7 |
| 18 | gi|226493727 | uncharacterized protein LOC100275650 [Zea mays] | 107 | 17.6/5.79 | 16/4.4 |
| 14 | gi|302819846 | Hypothetical protein SELMODRAFT_133757 [Selaginella moellendorffii] | 131 | 17.2/5.22 | 16/4.8 |
| 49 | gi|226508942 | Uncharacterized protein LOC100275367 [Zea mays] | 88 | 23.4/4.97 | 23/4.4 |
| 58 | gi|226528599 | Uncharacterized protein LOC100276423 [Zea mays] | 215 | 19.8/4.71 | 23/4.9 |
| 74 | gi|195635483 | Membrane steroid-binding protein 1 [Zea mays] | 425 | 27.9/5.45 | 28/5.0 |
| 62 | gi|223948417 | Unknown [Zea mays] | 115 | 20.4/7.66 | 15/4.8 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, R.; Gao, F.; Guo, B.-Q.; Huang, J.-C.; Wang, L.; Zhou, Y.-J. Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome. Int. J. Mol. Sci. 2013, 14, 11125-11144. https://doi.org/10.3390/ijms140611125
Wang R, Gao F, Guo B-Q, Huang J-C, Wang L, Zhou Y-J. Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome. International Journal of Molecular Sciences. 2013; 14(6):11125-11144. https://doi.org/10.3390/ijms140611125
Chicago/Turabian StyleWang, Rong, Fei Gao, Bing-Qian Guo, Ji-Chang Huang, Lei Wang, and Yi-Jun Zhou. 2013. "Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome" International Journal of Molecular Sciences 14, no. 6: 11125-11144. https://doi.org/10.3390/ijms140611125
APA StyleWang, R., Gao, F., Guo, B.-Q., Huang, J.-C., Wang, L., & Zhou, Y.-J. (2013). Short-Term Chromium-Stress-Induced Alterations in the Maize Leaf Proteome. International Journal of Molecular Sciences, 14(6), 11125-11144. https://doi.org/10.3390/ijms140611125