Generation of Magnetized Olfactory Ensheathing Cells for Regenerative Studies in the Central and Peripheral Nervous Tissue

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
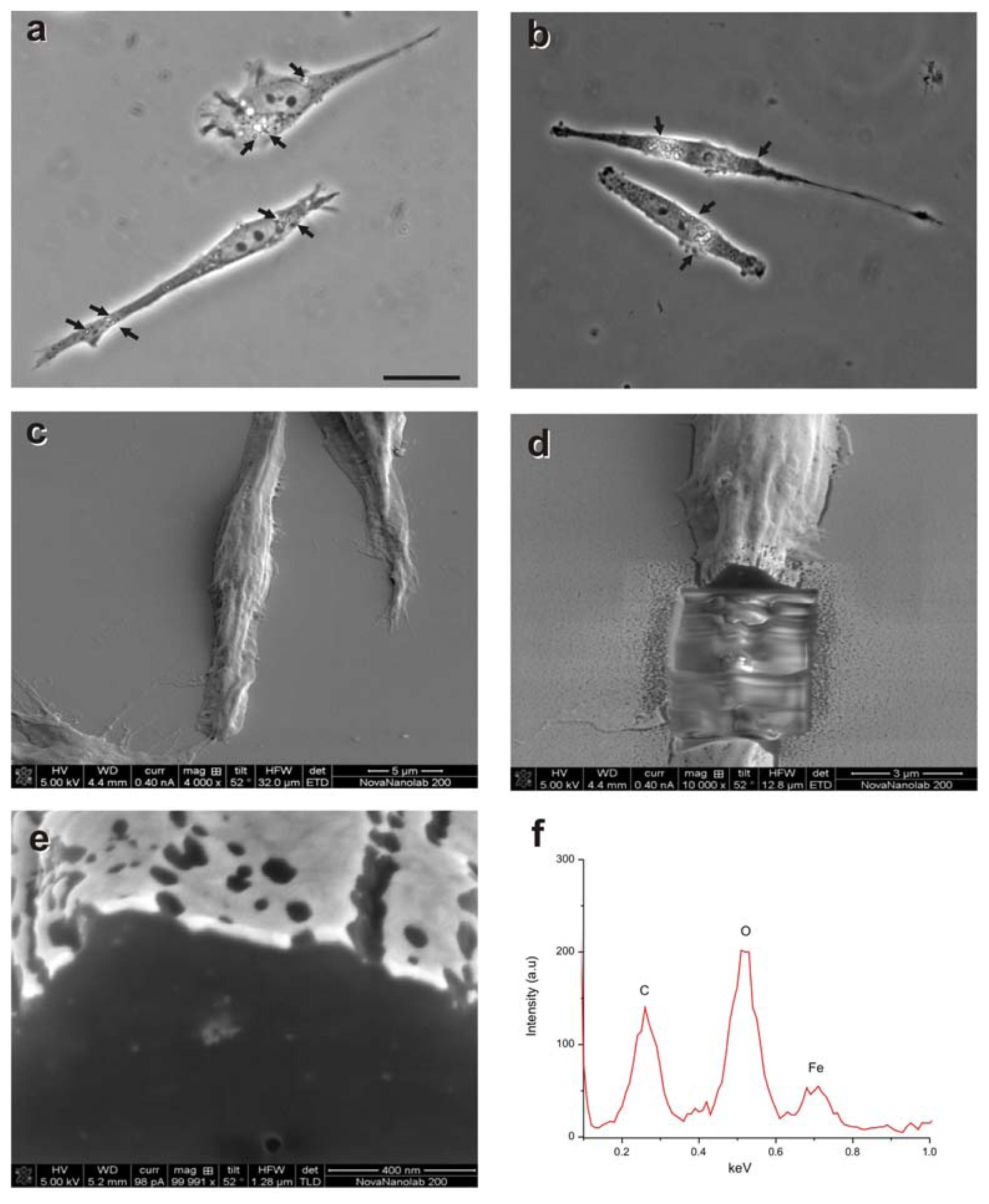
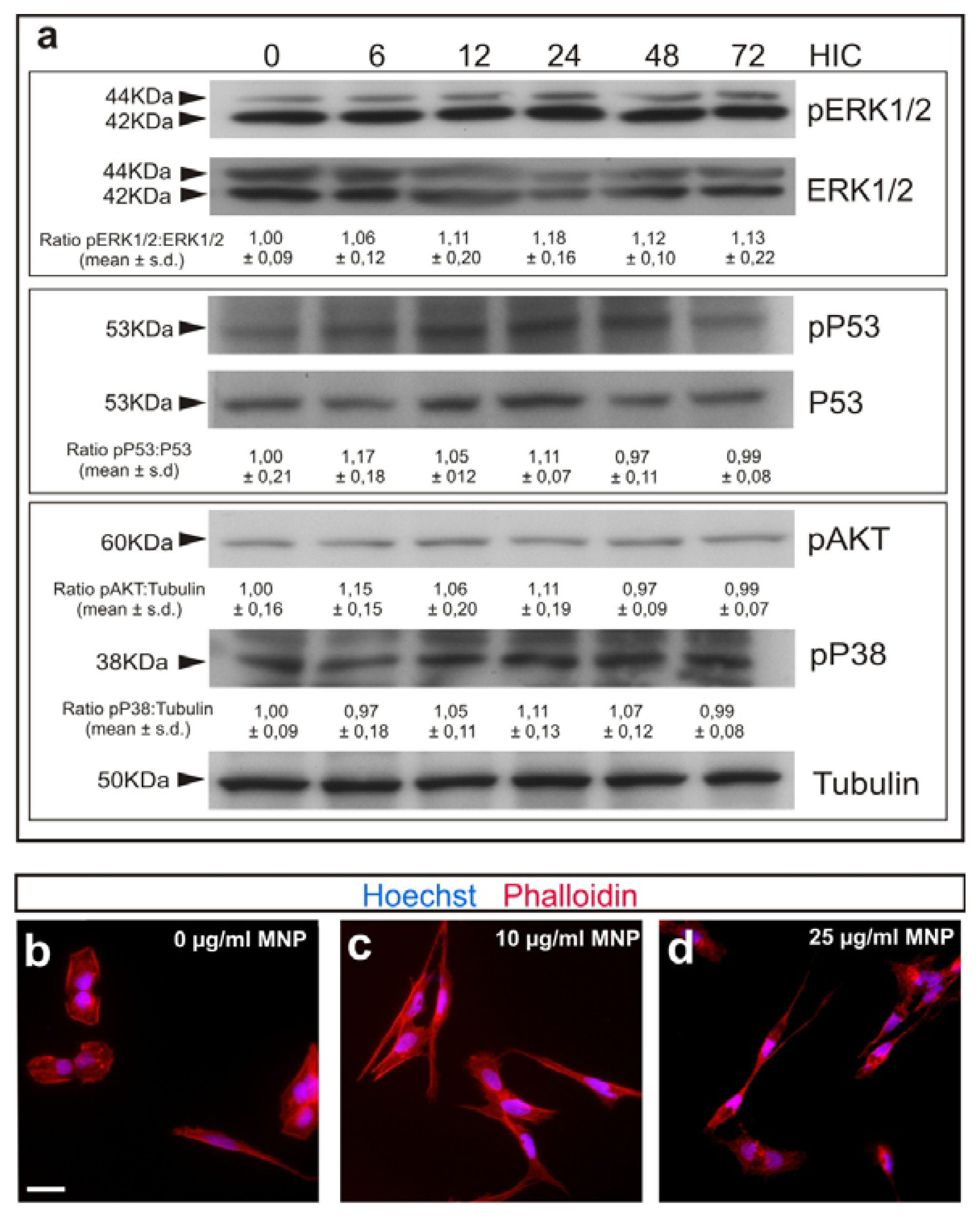
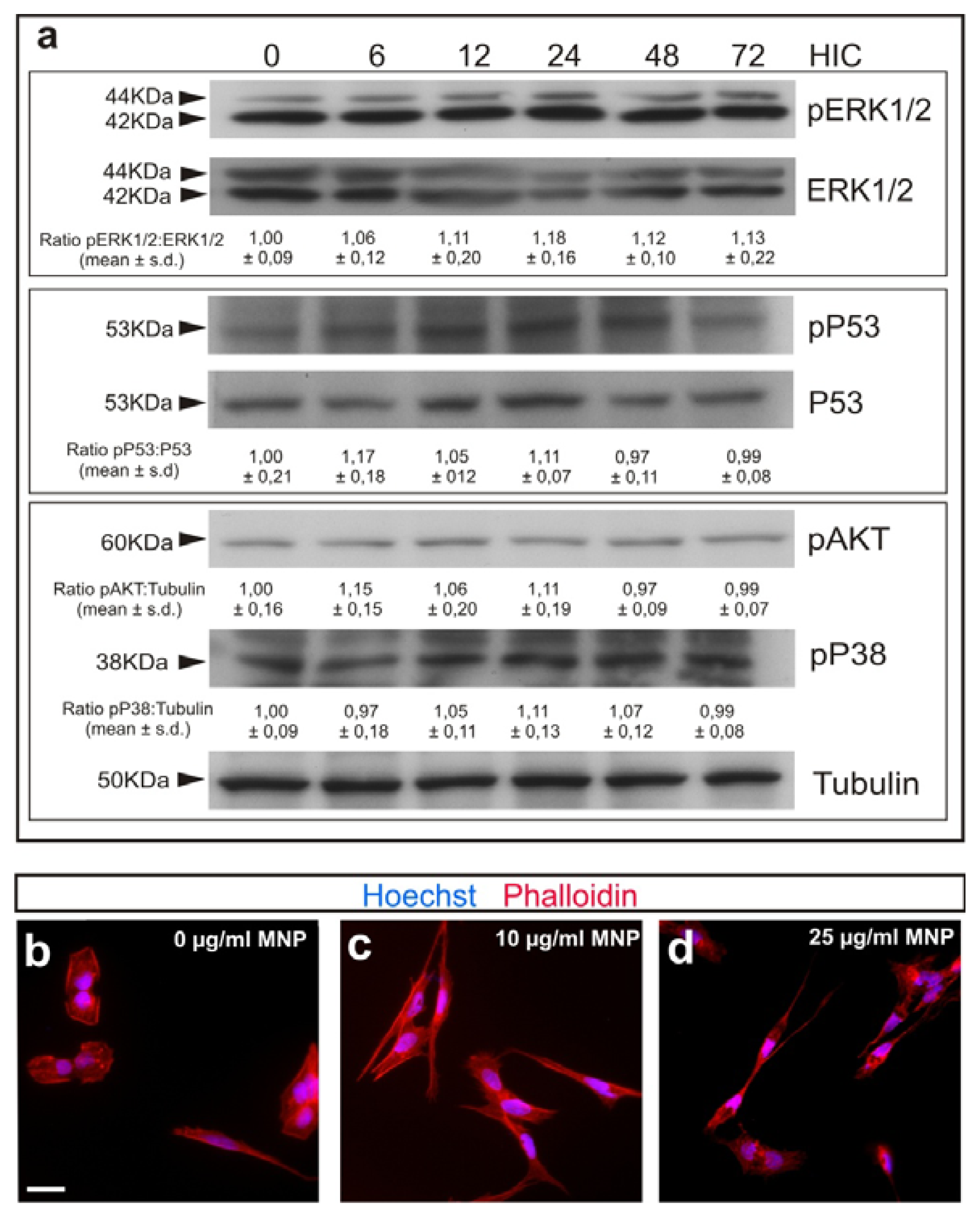
2.1. Magnetization of OECs via MNPs
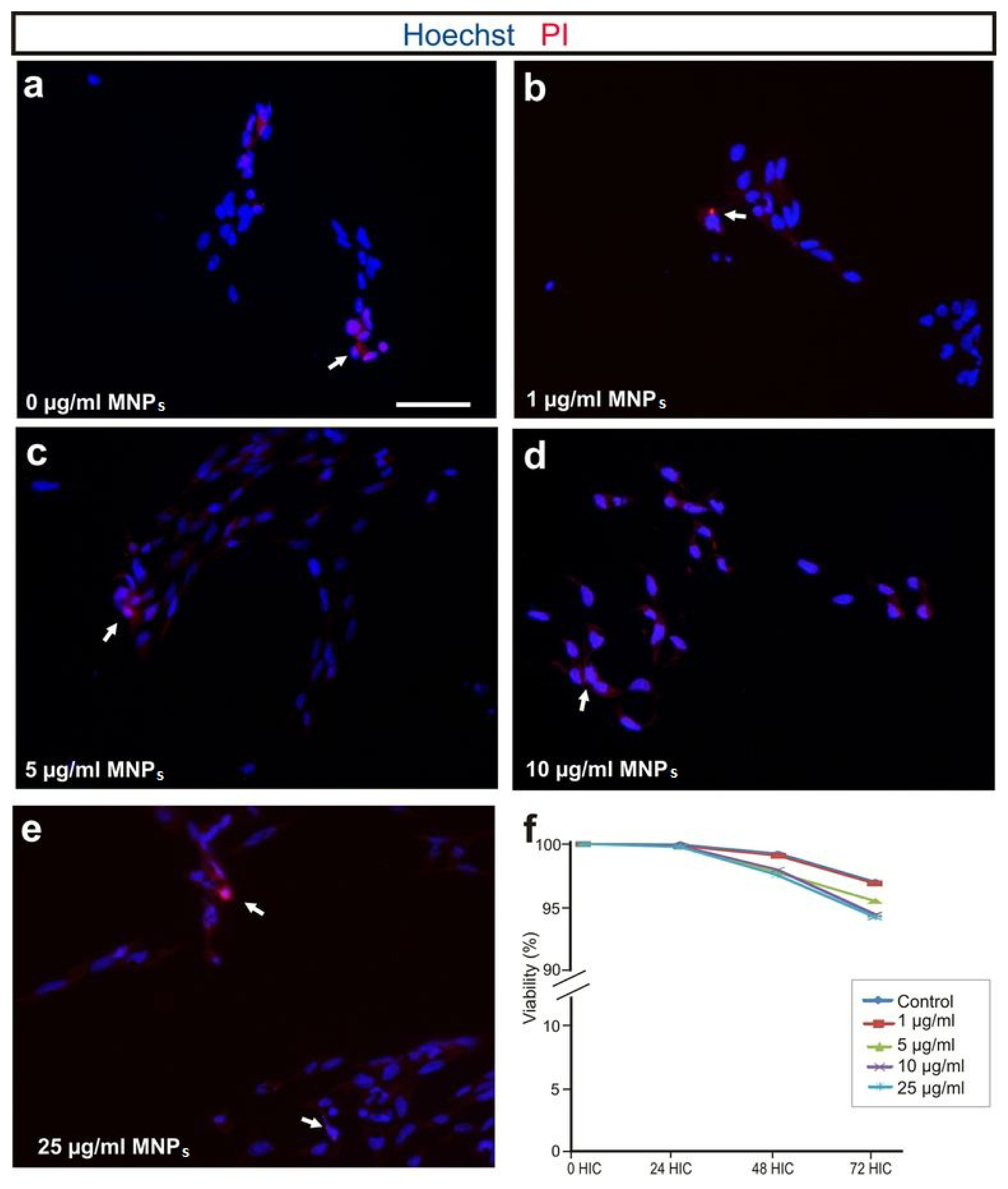
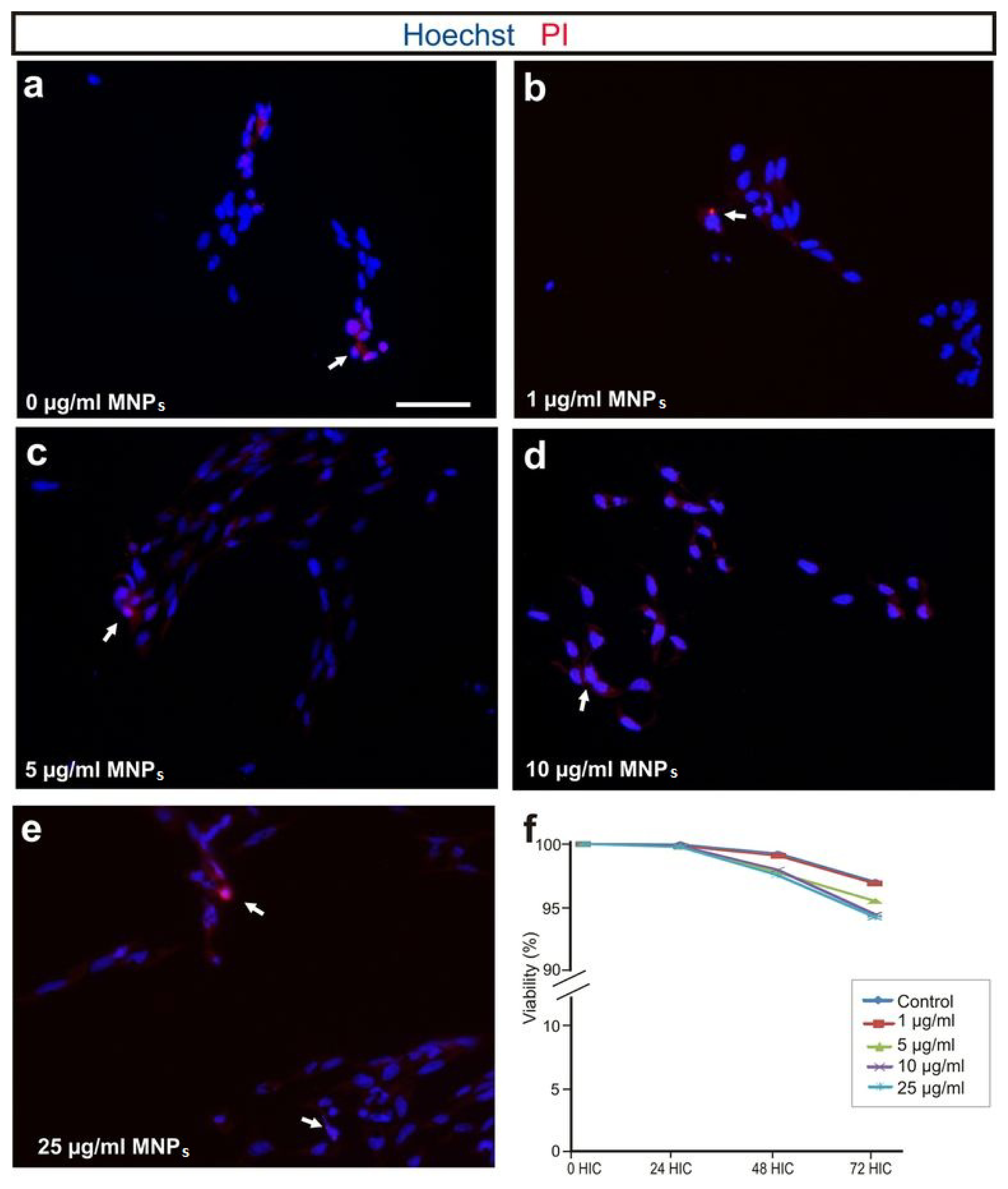
2.2. M-OECs Viability
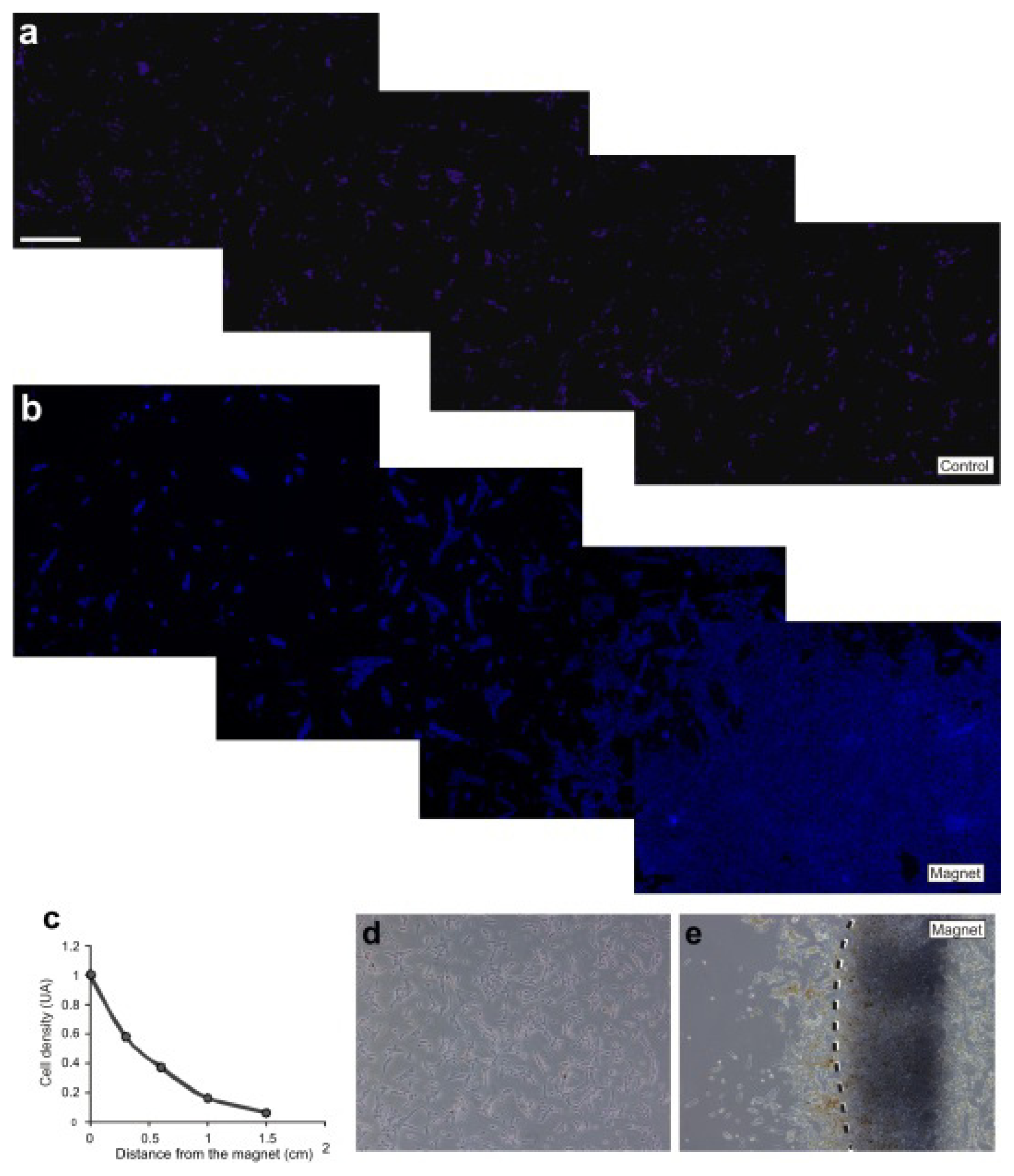
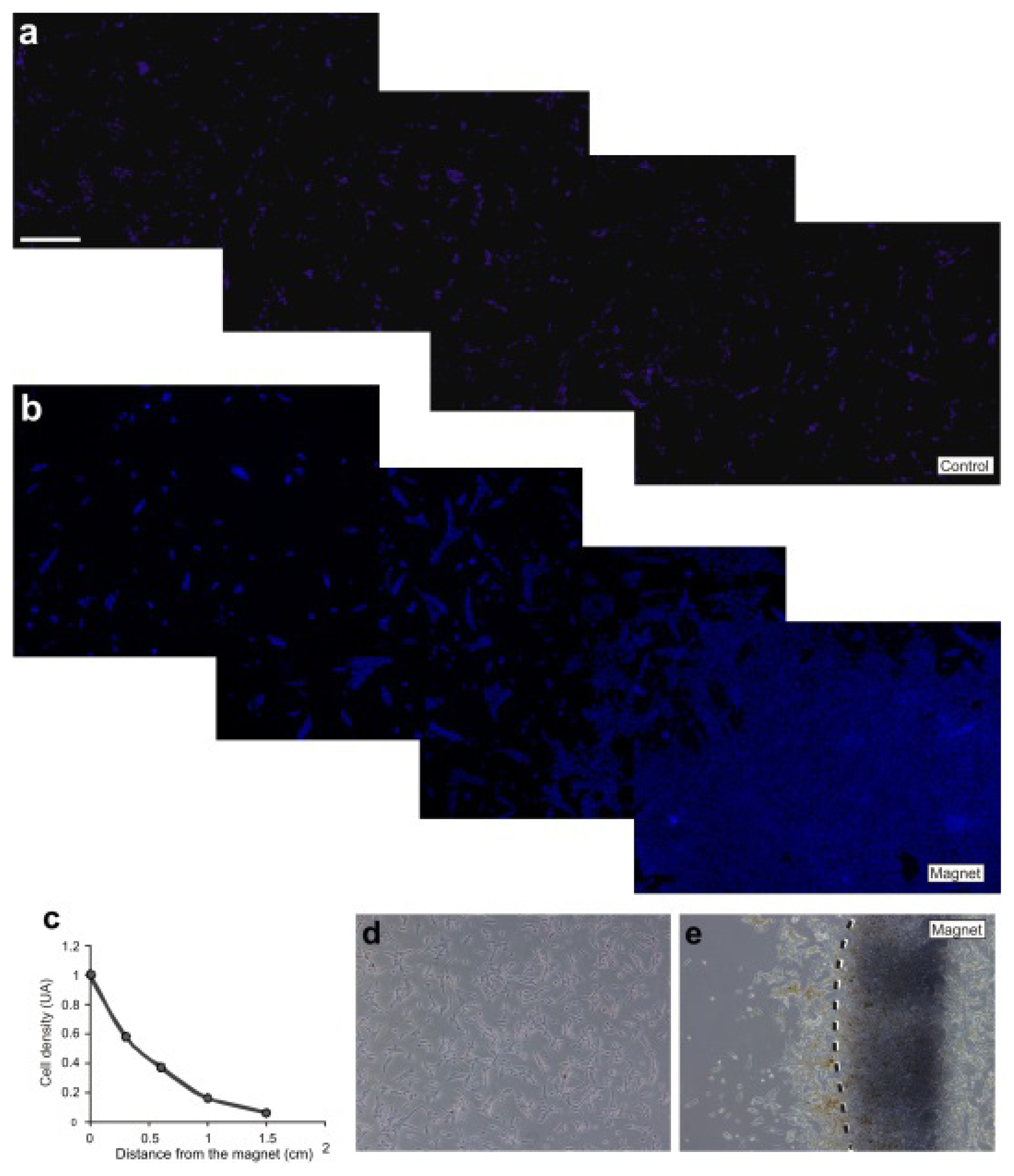
2.3. M-OECs Movement toward a Magnetic Source
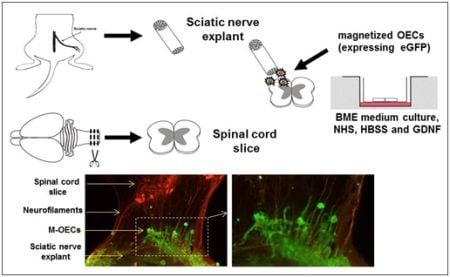
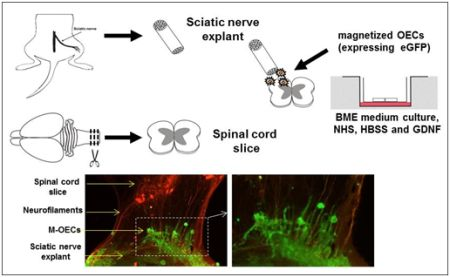
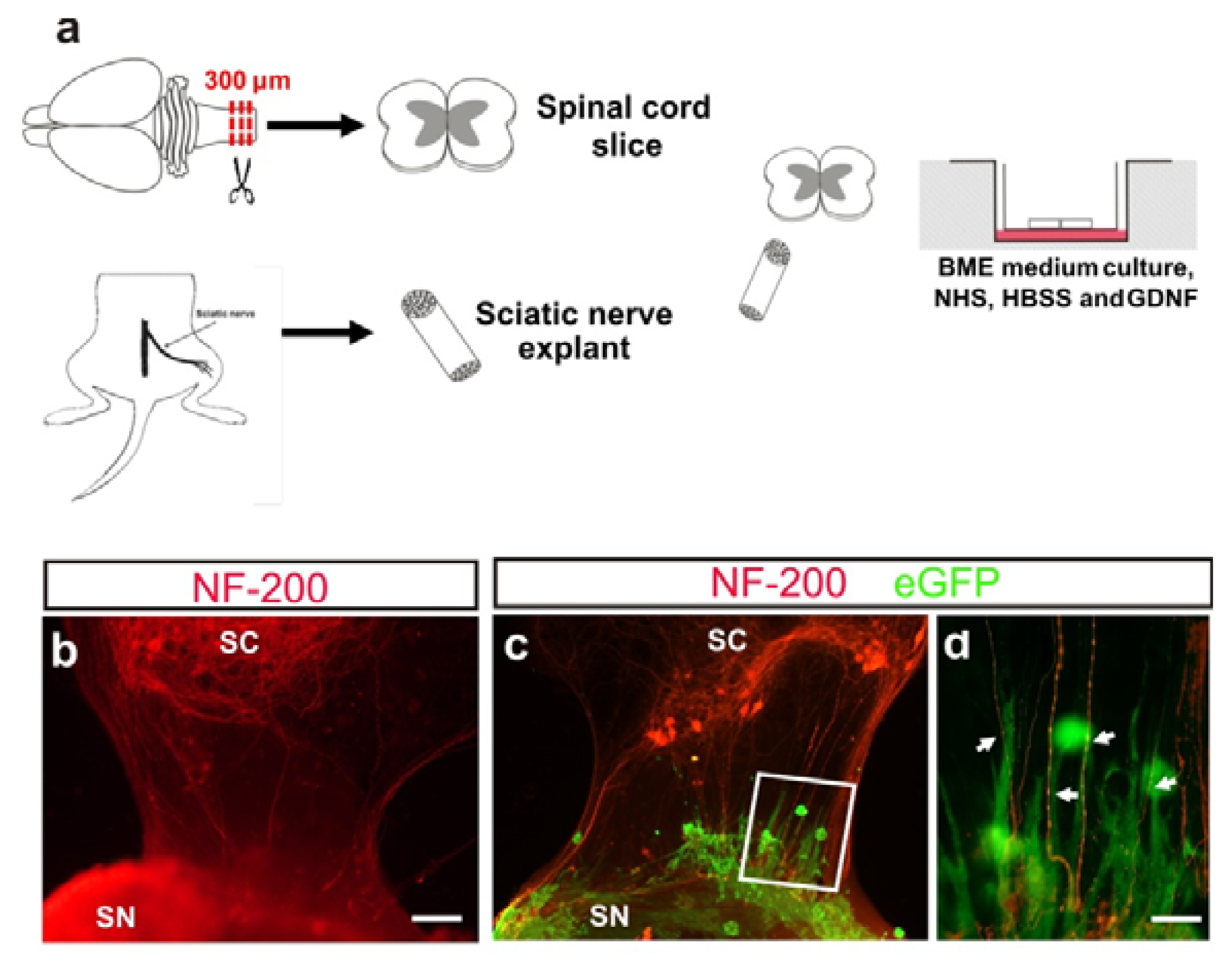
2.4. M-OECs Survive in an Organotypic Co-Culture of Peripheral Nerve and Spinal Cord
3. Discussion
4. Experimental Section
4.1. MNP Synthesis
4.2. OEC Cultures
4.3. Spinal Cord (SC) Isolation and Sciatic Nerve(SN) Isolation
4.4. PI Dye Exclusion Assay through Fluorescence Microscopy
4.5. PI Dye Exclusion Assay through Flow Cytometry
4.6. Immunoblotting Analysis
4.7. Phalloidin Staining
4.8. Immunocytochemistry of SC/SN Slice/OEC Co-Cultures
4.9. SEM/FIB Analysis
4.10. Cell Migration Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-14-10852-s001.pdfAcknowledgments
Conflict of Interest
References
- Doucette, R. Olfactory ensheathing cells-potential for glial-cell transplantation into areas of Cns injury. Histol. Histopathol 1995, 10, 503–507. [Google Scholar]
- Ramoncueto, A.; Valverde, F. Olfactory-bulb ensheathing glia: A unique cell-type with axonal growth-promoting properties. Glia 1995, 14, 163–173. [Google Scholar]
- Ramoncueto, A.; Nietosampedro, M. Regeneration into the spinal-cord of transected dorsal-root axons is promoted by ensheathing glia transplants. Exp. Neurol 1994, 127, 232–244. [Google Scholar]
- Li, Y.; Field, P.M.; Raisman, G. Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science 1997, 277, 2000–2002. [Google Scholar]
- Ramon-Cueto, A.; Cordero, M.I.; Santos-Benito, F.F.; Avila, J. Functional recovery of paraplegic rats and motor axon regeneration in their spinal cords by olfactory ensheathing glia. Neuron 2000, 25, 425–435. [Google Scholar]
- Garcia-Alias, G.; Lopez-Vales, R.; Fores, J.; Navarro, X.; Verdu, E. Acute transplantation of olfactory ensheathing cells or Schwann cells promotes recovery after spinal cord injury in the rat. J. Neurosci. Res 2004, 75, 632–641. [Google Scholar]
- Ruitenberg, M.J.; Plant, G.W.; Christensen, C.L.; Blits, B.; Niclou, S.P.; Harvey, A.R.; Boer, G.J.; Verhaagen, J. Viral vector-mediated gene expression in olfactory ensheathing glia implants in the lesioned rat spinal cord. Gene Ther 2002, 9, 135–146. [Google Scholar]
- Moreno-Flores, M.T.; Bradbury, E.J.; Martin-Bermejo, M.J.; Agudo, M.; Lim, F.; Pastrana, E.; Avila, J.; Diaz-Nido, J.; McMahon, S.B.; Wandosell, F. A clonal cell line from immortalized olfactory ensheathing glia promotes functional recovery in the injured spinal cord. Mol. Ther 2006, 13, 598–608. [Google Scholar]
- Raisman, G. Repair of spinal cord injury by transplantation of olfactory ensheathing cells. C. R. Biol 2007, 330, 557–560. [Google Scholar]
- Ramon-Cueto, A.; Munoz-Quiles, C. Clinical application of adult olfactory bulb ensheathing glia for nervous system repair. Exp. Neurol 2011, 229, 181–194. [Google Scholar]
- Ramoncueto, A.; Perez, J.; Nietosampedro, M. In vitro Enfolding of Olfactory Neurites by P75 Ngf Receptor-Positive Ensheathing Cells from Adult-Rat Olfactory-Bulb. Eur. J. Neurosci 1993, 5, 1172–1180. [Google Scholar]
- Franklin, R.J.M.; Gilson, J.M.; Franceschini, I.A.; Barnett, S.C. Schwann cell-like myelination following transplantation of an olfactory bulb-ensheathing cell line into areas of demyelination in the adult CNS. Glia 1996, 17, 217–224. [Google Scholar]
- Imaizumi, T.; Lankford, K.L.; Waxman, S.G.; Greer, C.A.; Kocsis, J.D. Transplanted olfactory ensheathing cells remyelinate and enhance axonal conduction in the demyelinated dorsal columns of the rat spinal cord. J. Neurosci 1998, 18, 6176–6185. [Google Scholar]
- Oudega, M. Schwann cell and olfactory ensheathing cell implantation for repair of the contused spinal cord. Acta Physiol 2007, 189, 181–189. [Google Scholar]
- Gudino-Cabrera, G.; Pastor, A.M.; de la Cruz, R.R.; Delgado-Garcia, J.M.; Nieto-Sampedro, M. Limits to the capacity of transplants of olfactory glia to promote axonal regrowth in the CNS. Neuroreport 2000, 11, 467–471. [Google Scholar]
- Nieto-Sampedro, M. Central nervous system lesions that can and those that cannot be repaired with the help of olfactory bulb ensheathing cell transplants. Neurochem. Res 2003, 28, 1659–1676. [Google Scholar]
- Lu, P.; Yang, H.; Culbertson, M.; Graham, L.; Roskams, A.J.; Tuszynski, M.H. Olfactory ensheathing cells do not exhibit unique migratory or axonal growth-promoting properties after spinal cord injury. J. Neurosci 2006, 26, 11120–11130. [Google Scholar]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci 2004, 5, 146–156. [Google Scholar]
- Fawcett, J.W. Overcoming inhibition in the damaged spinal cord. J. Neurotrauma 2006, 23, 371–383. [Google Scholar]
- Yan, H.L.; Lu, D.C.; Rivkees, S.A. Lysophosphatidic acid regulates the proliferation and migration of olfactory ensheathing cells in vitro. Glia 2003, 44, 26–36. [Google Scholar]
- Cao, L.; Su, Z.; Zhou, Q.; Lv, B.; Liu, X.; Jiao, L.; Li, Z.; Zhu, Y.; Huang, Z.; Huang, A.; et al. Glial cell line-derived neurotrophic factor promotes olfactory ensheathing cells migration. Glia 2006, 54, 536–544. [Google Scholar]
- Windus, L.C.E.; Chehrehasa, F.; Lineburg, K.E.; Claxton, C.; Mackay-Sim, A.; Key, B.; St. John, J.A. Stimulation of olfactory ensheathing cell motility enhances olfactory axon growth. Cell. Mol. Life Sci 2011, 68, 3233–3247. [Google Scholar]
- Su, Z.; Cao, L.; Zhu, Y.; Liu, X.; Huang, Z.; Huang, A.; He, C. Nogo enhances the adhesion of olfactory ensheathing cells and inhibits their migration. J. Cell Sci. 2007, 120, 1877–1887. [Google Scholar]
- Lee, I.H.; Bulte, J.W.M.; Schweinhardt, P.; Douglas, T.; Trifunovski, A.; Hofstetter, C.; Olson, L.; Spenger, C. In vivo magnetic resonance tracking of olfactory ensheathing glia grafted into the rat spinal cord. Exp. Neurol 2004, 187, 509–516. [Google Scholar]
- Jain, T.K.; Richey, J.; Strand, M.; Leslie-Pelecky, D.L.; Flask, C.A.; Labhasetwar, V. Magnetic nanoparticles with dual functional properties: Drug delivery and magnetic resonance imaging. Biomaterials 2008, 29, 4012–4021. [Google Scholar]
- Bulte, J.W.; Douglas, T.; Witwer, B.; Zhang, S.C.; Strable, E.; Lewis, B.K.; Zywicke, H.; Miller, B.; van Gelderen, P.; Moskowitz, B.W.; et al. Magnetodendrimers allow endosomal magnetic labeling and in vivo tracking of stem cells. Nat. Biotechnol 2001, 19, 1141–1147. [Google Scholar]
- Maier-Hauff, K.; Ulrich, F.; Nestler, D.; Niehoff, H.; Wust, P.; Thiesen, B.; Orawa, H.; Budach, V.; Jordan, A. Efficacy and safety of intratumoral thermotherapy using magnetic iron-oxide nanoparticles combined with external beam radiotherapy on patients with recurrent glioblastoma multiforme. J. Neuro-Oncol 2011, 103, 317–324. [Google Scholar]
- Muthana, M.; Scott, S.D.; Farrow, N.; Morrow, F.; Murdoch, C.; Grubb, S.; Brown, N.; Dobson, J.; Lewis, C.E. A novel magnetic approach to enhance the efficacy of cell-based gene therapies. Gene Ther 2008, 15, 902–910. [Google Scholar]
- Pankhurst, Q.A.; Connolly, J.; Jones, S.K.; Dobson, J. Applications of magnetic nanoparticles in biomedicine. J. Phys. D 2003, 36, R167–R181. [Google Scholar]
- Dobson, J. Remote control of cellular behaviour with magnetic nanoparticles. Nat. Nanotechnol 2008, 3, 139–143. [Google Scholar]
- Pensabene, V.; Vittorio, O.; Raffa, V.; Ziaei, A.; Menciassi, A.; Dario, P. Neuroblastoma cells displacement by magnetic carbon nanotubes. IEEE Trans. Nanobiosci 2008, 7, 105–110. [Google Scholar]
- Raffa, V.; Vittorio, O.; Ciofani, G.; Pensabene, V.; Cuschieri, A. Cell creeping and controlled migration by magnetic carbon nanotubes. Nanoscale Res Lett 2010, 5, 257–262. [Google Scholar]
- Vittorio, O.; Quaranta, P.; Raffa, V.; Funel, N.; Campani, D.; Pelliccioni, S.; Longoni, B.; Mosca, F.; Pietrabissa, A.; Cuschieri, A. Magnetic carbon nanotubes: A new tool for shepherding mesenchymal stem cells by magnetic fields. Nanomedicine 2011, 6, 43–54. [Google Scholar]
- Riggio, C.; Calatayud, M.P.; Hoskins, C.; Pinkernelle, J.; Sanz, B.; Torres, T.E.; Ibarra, M.R.; Wang, L.; Keilhoff, G.; Goya, G.F.; et al. Poly-l-lysine-coated magnetic nanoparticles as intracellular actuators for neural guidance. Int. J. Nanomed 2012, 7, 3155–3166. [Google Scholar]
- Nocentini, S.; Reginensi, D.; Garcia, S.; Carulla, P.; Moreno-Flores, M.T.; Wandosell, F.; Trepat, X.; Bribian, A.; del Río, J.A. Myelin-associated proteins block the migration of olfactory ensheathing cells: An in vitro study using single-cell tracking and traction force microscopy. Cell. Mol. Life Sci 2012, 69, 1689–1703. [Google Scholar]
- Vittorio, O.; Raffa, V.; Riggio, C.; Pietrabissa, A.; Cuschieri, A. PC12 Interaction with magnetic nanotubes: Effects on viability, cell differentiation and cell translocation induced by a magnetic field. Curr Nanosci 2011, 7, 337–344. [Google Scholar]
- Morales, R.; Riss, M.; Wang, L.; Gavin, R.; Del Rio, J.A.; Alcubilla, R.; Claverol-Tinture, E. Integrating multi-unit electrophysiology and plastic culture dishes for network neuroscience. Lab. Chip 2008, 8, 1896–1905. [Google Scholar]
- Vyas, A.; Li, Z.; Aspalter, M.; Feiner, J.; Hoke, A.; Zhou, C.; O’Daly, A.; Abdullah, M.; Rohde, C.; Brushart, T.M. An in vitro model of adult mammalian nerve repair. Exp. Neurol 2010, 223, 112–118. [Google Scholar]
- Allodi, I.; Guzman-Lenis, M.S.; Hernandez, J.; Navarro, X.; Udina, E. In vitro comparison of motor and sensory neuron outgrowth in a 3D collagen matrix. J. Neurosci. Methods 2011, 198, 53–61. [Google Scholar]
- Penna, V.; Stark, G.B.; Wewetzer, K.; Radtke, C.; Lang, E.M. Comparison of Schwann cells and olfactory ensheathing cells for peripheral nerve gap bridging. Cells Tissues Organs 2012, 196, 534–542. [Google Scholar]
- Wewetzer, K.; Verdu, E.; Angelov, D.N.; Navarro, X. Olfactory ensheathing glia and Schwann cells: Two of a kind? Cell Tissue Res 2002, 309, 337–345. [Google Scholar]
- Tetzlaff, W.; Okon, E.B.; Karimi-Abdolrezaee, S.; Hill, C.E.; Sparling, J.S.; Plemel, J.R.; Plunet, W.T.; Tsai, E.C.; Baptiste, D.; Smithson, L.J.; et al. A systematic review of cellular transplantation therapies for spinal cord injury. J. Neurotrauma 2011, 28, 1611–1682. [Google Scholar]
- Radtke, C.; Wewetzer, K.; Reimers, K.; Vogt, P.M. Transplantation of olfactory ensheathing cells as adjunct cell therapy for peripheral nerve injury. Cell Transplant 2011, 20, 145–152. [Google Scholar]
- Plant, G.W.; Harvey, A.R.; Leaver, S.G.; Lee, S.V. Olfactory ensheathing glia: Repairing injury to the mammalian visual system. Exp. Neurol 2011, 229, 99–108. [Google Scholar]
- Ekberg, J.A.; Amaya, D.; Mackay-Sim, A.; St. John, J.A. The migration of olfactory ensheathing cells during development and regeneration. Neurosignals 2012, 20, 147–158. [Google Scholar]
- Kocsis, J.D.; Lankford, K.L.; Sasaki, M.; Radtke, C. Unique in vivo properties of olfactory ensheathing cells that may contribute to neural repair and protection following spinal cord injury. Neurosci. Lett 2009, 456, 137–142. [Google Scholar]
- Radtke, C.; Aizer, A.A.; Agulian, S.K.; Lankford, K.L.; Vogt, P.M.; Kocsis, J.D. Transplantation of olfactory ensheathing cells enhances peripheral nerve regeneration after microsurgical nerve repair. Brain Res 2009, 1254, 10–17. [Google Scholar]
- Bartolomei, J.C.; Greer, C.A. Olfactory ensheathing cells: Bridging the gap in spinal cord injury. Neurosurgery 2000, 47, 1057–1069. [Google Scholar]
- Mackay-Sim, A.; St John, J.A. Olfactory ensheathing cells from the nose: Clinical application in human spinal cord injuries. Exp. Neurol 2011, 229, 174–180. [Google Scholar]
- Pellitteri, R.; Spatuzza, M.; Stanzani, S.; Zaccheo, D. Biomarkers expression in rat olfactory ensheathing cells. Front Biosci 2010, 2, 289–298. [Google Scholar]
- Sasaki, M.; Lankford, K.L.; Radtke, C.; Honmou, O.; Kocsis, J.D. Remyelination after olfactory ensheathing cell transplantation into diverse demyelinating environments. Exp. Neurol 2011, 229, 88–98. [Google Scholar]
- Huang, Z.H.; Wang, Y.; Cao, L.; Su, Z.D.; Zhu, Y.L.; Chen, Y.Z.; Yuan, X.B.; He, C. Migratory properties of cultured olfactory ensheathing cells by single-cell migration assay. Cell Res 2008, 18, 479–490. [Google Scholar]
- Dash, A.; Blackman, B.R.; Wamhoff, B.R. Organotypic systems in drug metabolism and toxicity: Challenges and opportunities. Expert Opin. Drug Metab. Toxicol 2012, 8, 999–1014. [Google Scholar]
- Pena, F. Organotypic cultures as tool to test long-term effects of chemicals on the nervous system. Curr. Med. Chem 2010, 17, 987–1001. [Google Scholar]
- Del Rio, J.A.; Soriano, E. Regenerating cortical connections in a dish: The entorhino-hippocampal organotypic slice co-culture as tool for pharmacological screening of molecules promoting axon regeneration. Nat. Protoc 2010, 5, 217–226. [Google Scholar]
- Calatayud, M.P.; Cristina, R.; Vittoria, R.; Beatriz, S.; Teobaldo, E.; Torres, M.; Ricardo, I.; Clare, H.; Alfred, C.; Wang, L.Y.; et al. Goya Neuronal cells loaded with PEI-coated Fe3O4 nanoparticles for magnetically-guided nerve regeneration. J. Mater. Chem. Submitted for publication. 2012. [Google Scholar]
- Rudolph, J.G.; Lemasters, J.J.; Crews, F.T. Use of a multiwell fluorescence scanner with propidium iodide to assess NMDA mediated excitotoxicity in rat cortical neuronal cultures. Neurosci. Lett 1997, 221, 149–152. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Riggio, C.; Nocentini, S.; Catalayud, M.P.; Goya, G.F.; Cuschieri, A.; Raffa, V.; Del Río, J.A. Generation of Magnetized Olfactory Ensheathing Cells for Regenerative Studies in the Central and Peripheral Nervous Tissue. Int. J. Mol. Sci. 2013, 14, 10852-10868. https://doi.org/10.3390/ijms140610852
Riggio C, Nocentini S, Catalayud MP, Goya GF, Cuschieri A, Raffa V, Del Río JA. Generation of Magnetized Olfactory Ensheathing Cells for Regenerative Studies in the Central and Peripheral Nervous Tissue. International Journal of Molecular Sciences. 2013; 14(6):10852-10868. https://doi.org/10.3390/ijms140610852
Chicago/Turabian StyleRiggio, Cristina, Sara Nocentini, Maria Pilar Catalayud, Gerardo Fabian Goya, Alfred Cuschieri, Vittoria Raffa, and José Antonio Del Río. 2013. "Generation of Magnetized Olfactory Ensheathing Cells for Regenerative Studies in the Central and Peripheral Nervous Tissue" International Journal of Molecular Sciences 14, no. 6: 10852-10868. https://doi.org/10.3390/ijms140610852