Two Common Bean Genotypes with Contrasting Response to Phosphorus Deficiency Show Variations in the microRNA 399-Mediated PvPHO2 Regulation within the PvPHR1 Signaling Pathway

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
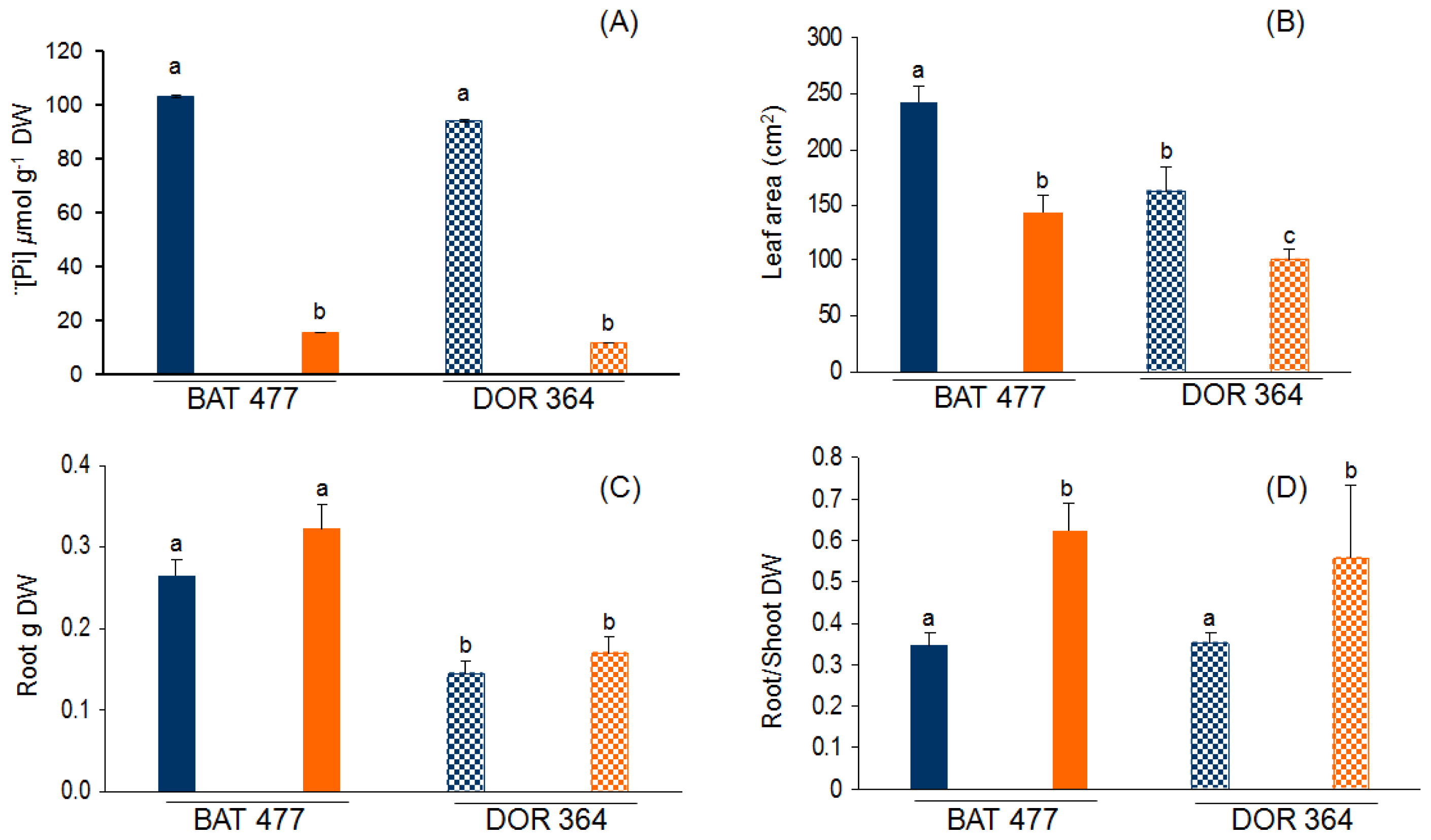
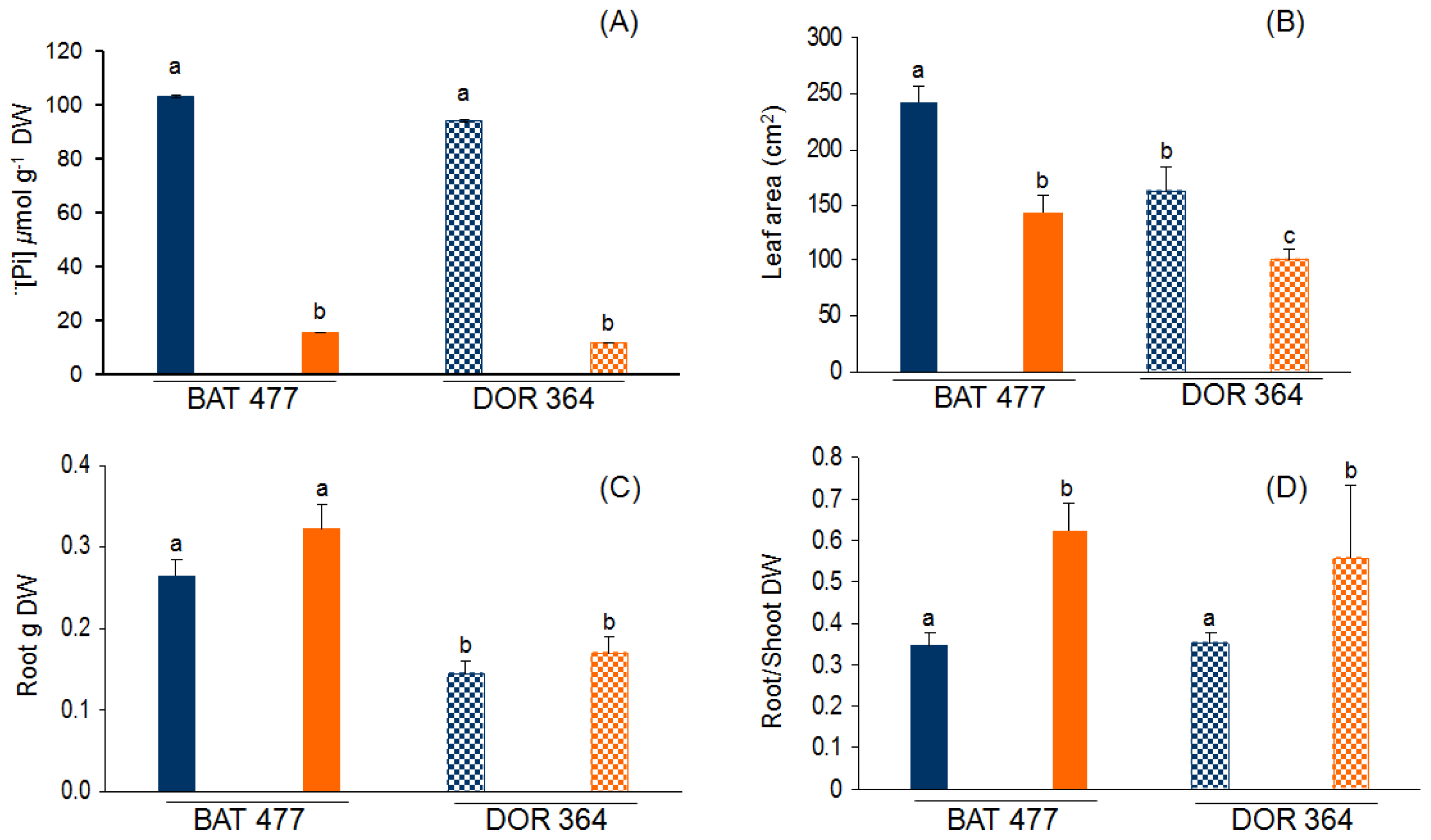
2.1. Comparative Phenotypic Analysis of BAT477 vs. DOR364 Plants under P Deficiency
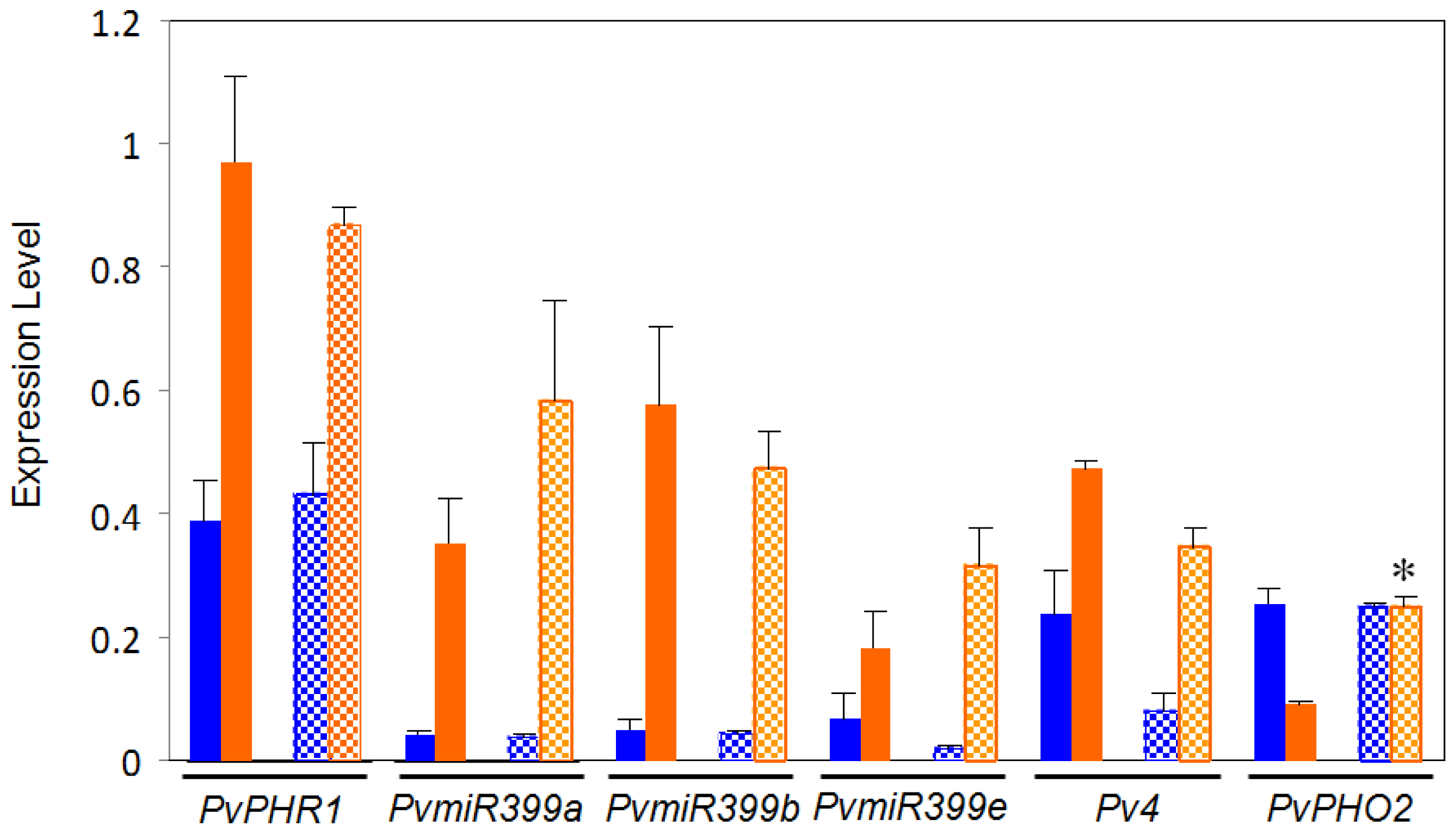
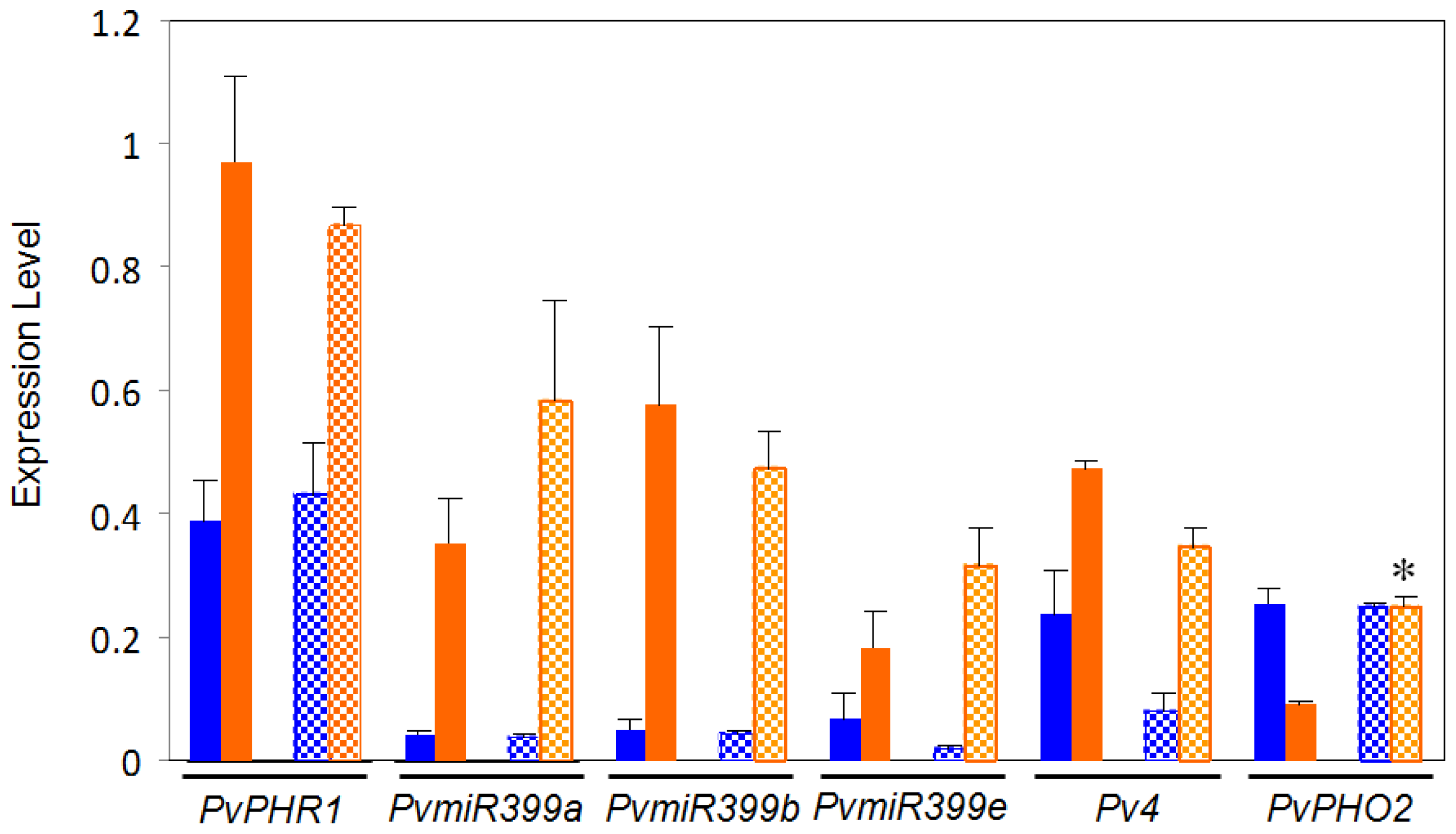
2.2. Comparative Expression Analysis of Regulatory Genes from the PvPHR1 Signaling Pathway, BAT477 vs. DOR364 P-Deficient Roots
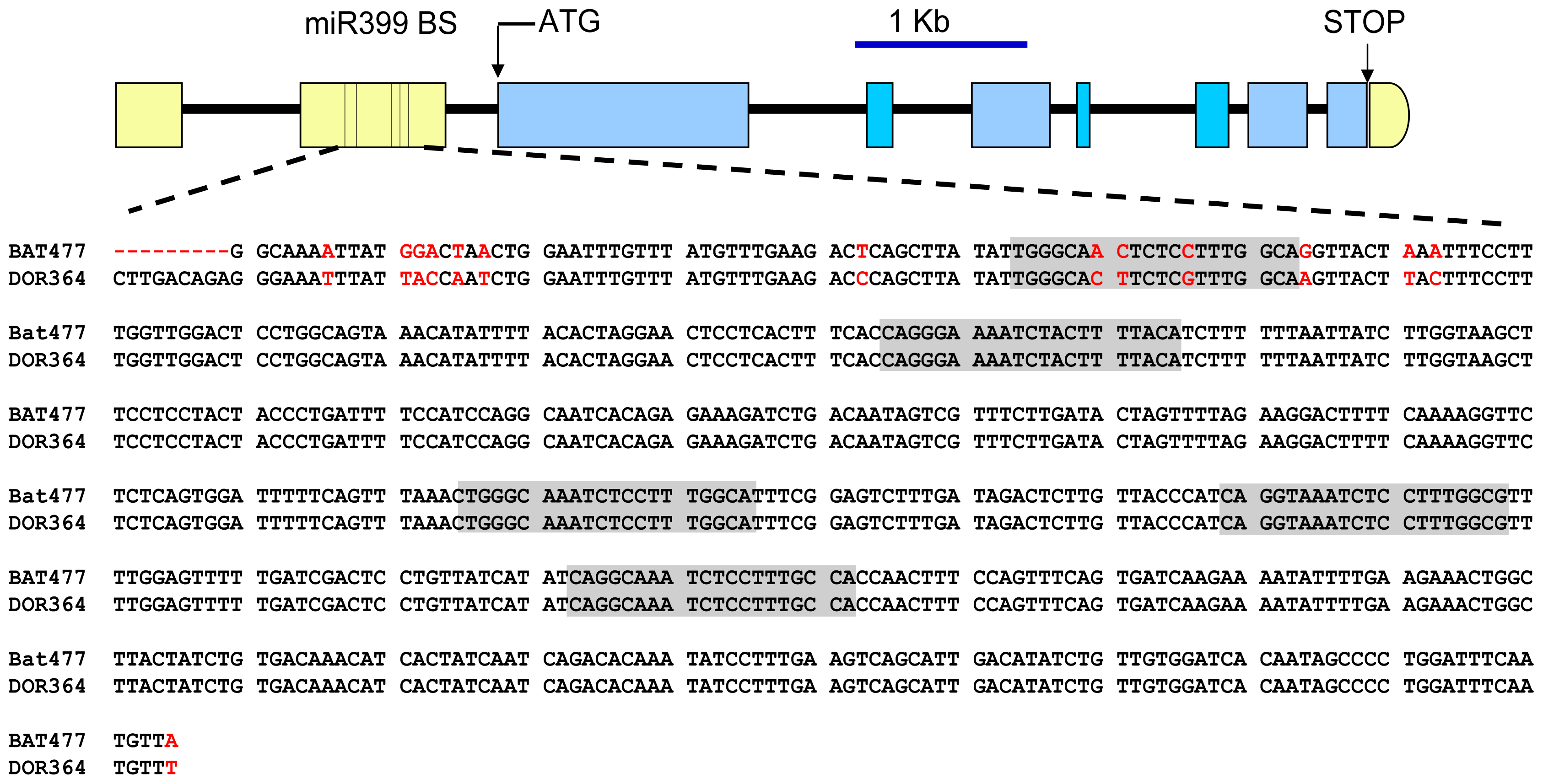
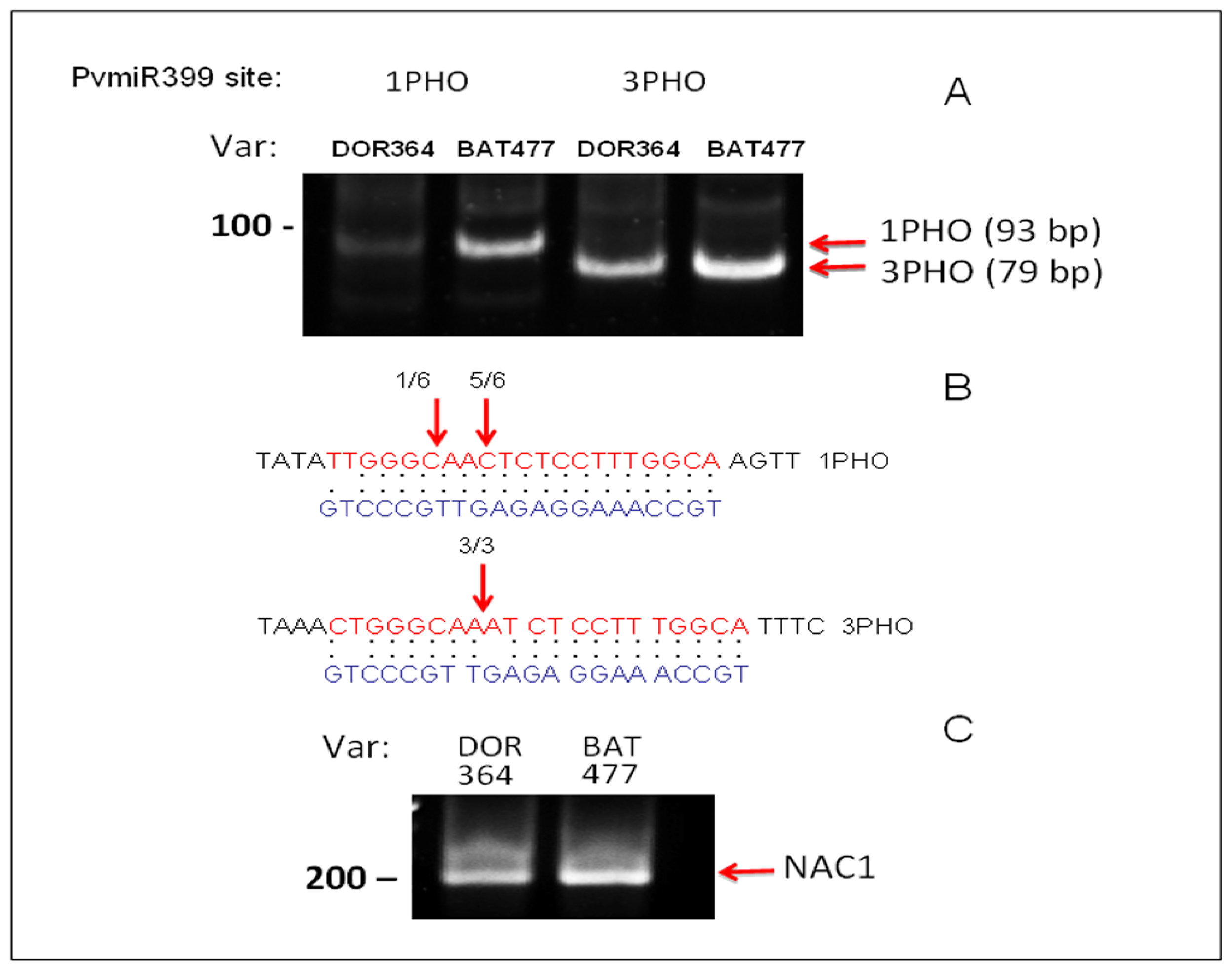
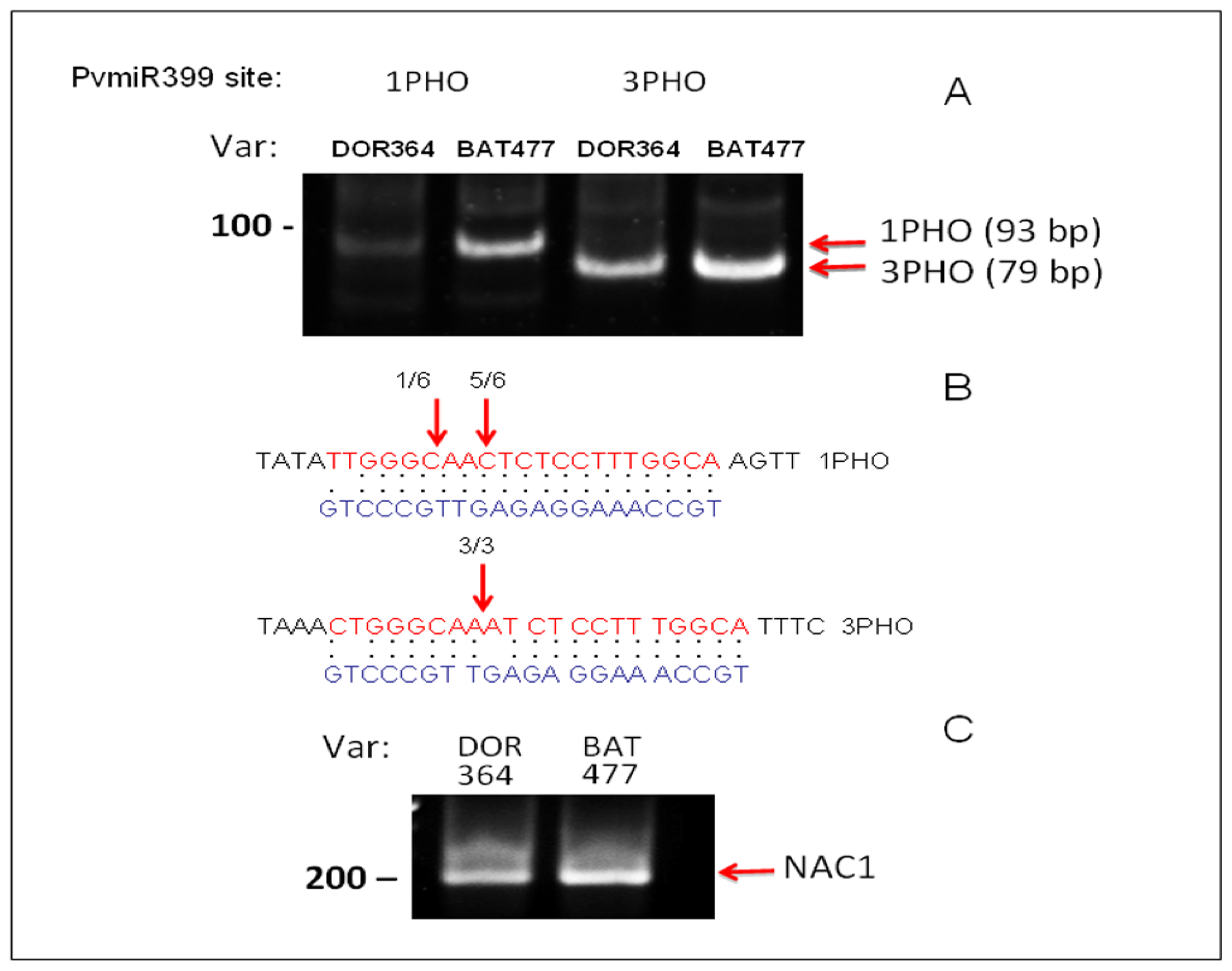
2.3. Differential Regulation of PvPHO2 by PvmiR399 in BAT477 vs. DOR364 P-Deficient Roots
3. Experimental Section
3.1. Plant Material and Growth Conditions
3.2. Phenotypic Characterization
3.3. Real-Time Quantitative RT-PCR (qRT-PCR)
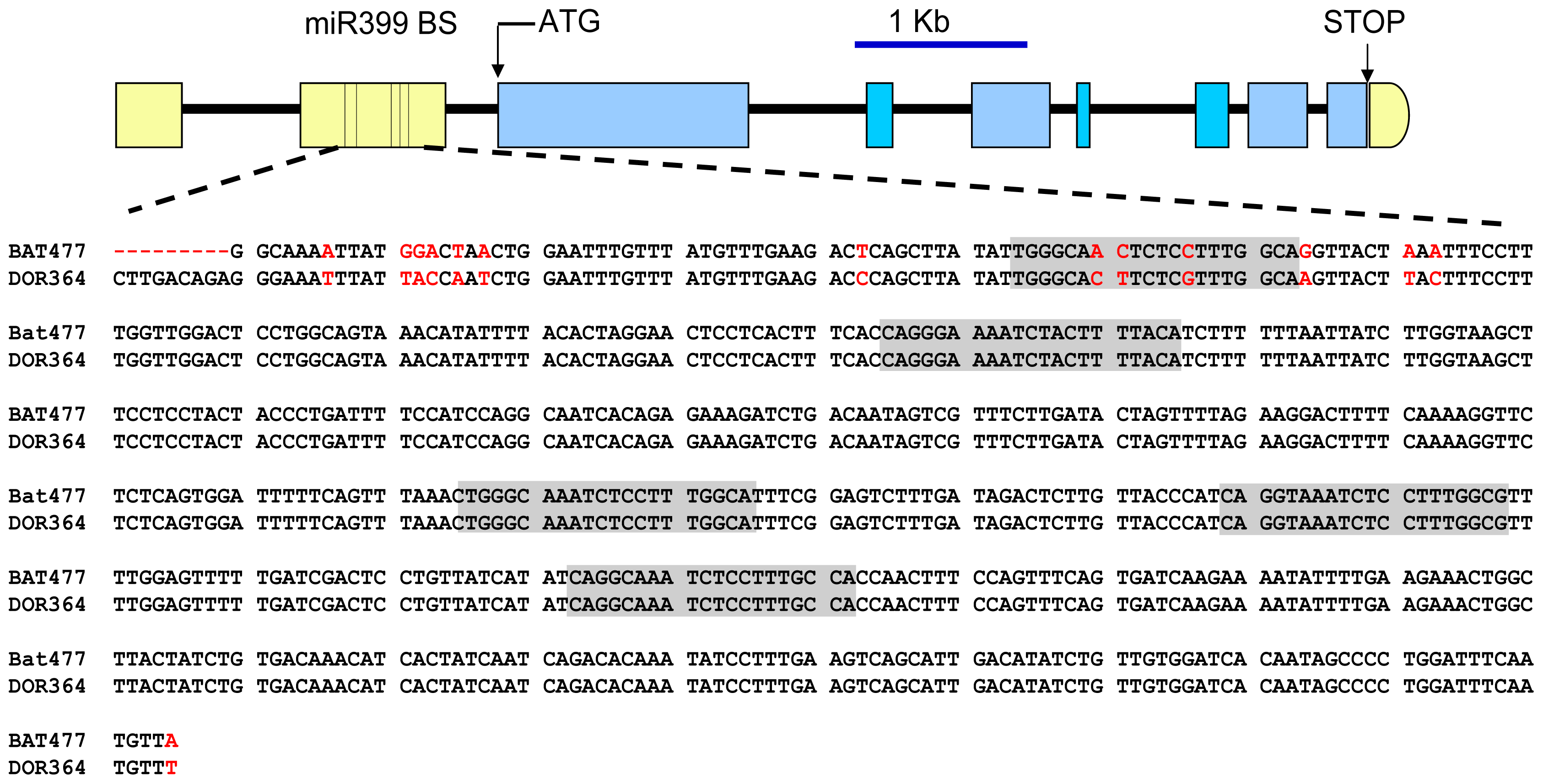
3.4. Cloning and DNA Sequence Analysis of Pv4 Gene and of the 5′ UTR from PvPHO2
3.5. Target Validation by 5′RACE of PvPHO2 mRNA
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Bieleski, R.L.; Fergunson, I.B. Physiology and Metabolism of Phosphate and Its Compounds. In Encyclopedia of Plant Physiol; Lauchli, A., Bieleski, R.L., Eds.; Springer-Verlag: Berlin, Germany, 1983; Volume 15A, pp. 422–449. [Google Scholar]
- Theodorou, M.E.; Plaxton, W.C. Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant. Physiol 1993, 101, 339–344. [Google Scholar]
- Chiou, T.J.; Aung, K.; Lin, S.I.; Wu, C.C.; Chiang, S.F.; Su, C.L. Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 2006, 18, 412–421. [Google Scholar]
- Vance, C.P.; Udhe-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptation by plants for securing a non renewable resource. New Phytol 2003, 157, 423–447. [Google Scholar]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang 2009, 19, 292–305. [Google Scholar]
- Franco-Zorrilla, J.M.; González, E.; Linhares, F.; Leyva, A.; Paz-Ares, J. The transcriptional control of plant responser to phosphate limitation. J. Exp. Bot 2004, 55, 285–293. [Google Scholar]
- Raghothama, K.G. Phosphate acquisition. Annu. Rev. Plant. Phys 1999, 50, 665–693. [Google Scholar]
- Valdés-López, O.; Hernández, G. Transcriptional regulation and signaling in phosphorus starvation: What about legumes? J. Integr. Plant. Biol 2008, 50, 1213–1222. [Google Scholar]
- Hammond, J.P.; Bennett, M.J.; Bowen, H.C.; Broadley, M.R.; Eastwood, D.C.; May, S.T.; Rahn, C.; Swarup, R.; Woolaway, K.E.; White, P.J. Changes in gene expression in Arabidopsis shoots during phosphate starvation and the potential for developing smart plants. Plant Physiol 2003, 132, 578–596. [Google Scholar]
- Wu, P.; Ma, L.; Hou, X.; Wang, M.; Wu, Y.; Liu, F.; Deng, X.W. Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves. Plant. Physiol 2003, 132, 1260–1271. [Google Scholar]
- Wasaki, J.; Yonetani, R.; Kuroda, S.; Shinano, T.; Yazaki, J.; Fujii, F.; Shimbo, K.; Yamamoto, K.; Sakata, K.; Sasaki, T. Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots. Plant. Cell Environ 2003, 26, 1515–1523. [Google Scholar]
- Uhde-Stone, C.; Zinn, K.E.; Ramirez-Yañez, M.; Li, A.; Vance, C.P.; Allan, D.L. Nylon filter arrays reveal differential gene expression in proteoid roots of white lupin in response to phosphorous deficiency. Plant Physiol 2003, 131, 1064–1079. [Google Scholar]
- Misson, J.; Raghothama, K.G.; Jain, A. A genome-wide transcriptional analysis using Arabidopsis thaliana Affimetrix gene chips determined plant responses to phosphate deprivation. Proc. Natl. Acad. Sci. USA 2005, 102, 11934–11939. [Google Scholar]
- Hernández, G.; Ramírez, M.; Valdés-López, O.; Graham, M.A.; Czechowki, T.; Schlereth, A.; Wandrey, M.; Erban, A.; Cheung, F.; Wu, H.C.; et al. Phosphorus stress in common bean: Root transcript and metabolic responses. Plant Physiol. 2007, 144, 752–767. [Google Scholar]
- Hernández, G.; Valdés-López, O.; Ramírez, M.; Goffard, N.; Weiller, G.; Aparicio-Fabre, R.; Fuentes, S.I.; Erban, A.; Kopka, J.; Udvardi, M.K.; et al. Global changes in the transcript and metabolic profiles during symbiotic nitrogen fixation in phosphorus-stressed common bean plants. Plant Physiol 2009, 151, 1221–1238. [Google Scholar]
- Devaiah, B.; Karthikeyan, A.S.; Raghothama, K.G. WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis. Plant Physiol 2007, 143, 1789–1801. [Google Scholar]
- Devaiah, B.; Nagarajna, V.K.; Raghothama, K.G. Phosphate homeostasis and root development in Arabidopsis are synchronized by the zing finger transcription factor ZAT6. Plant Physiol 2007, 145, 147–159. [Google Scholar]
- Chen, Z.H.; Nimmo, G.A.; Jeenkins, G.I.; Nimmo, H.G. BHLH32 modulates several biochemical and morphological process that respond to Pi starvation in Arabidopsis. Biochem. J 2007, 405, 191–198. [Google Scholar]
- Devaiah, B.; Madhuvanthi, R.; Karthikeyan, A.S.; Raghothama, K.G. Phosphate starvation responses and giberellic acid biosynthesis are regulated by the MYB62 transcription factor in Arabidopsis. Mol. Plant 2009, 2, 43–58. [Google Scholar]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plant and unicellular algae. Genes Dev 2001, 15, 2122–2133. [Google Scholar]
- Wykoff, D.; Grossman, A.; Weeks, D.P.; Usuda, H.; Shimogawara, K. Psr1, a nuclear localized protein that regulates phosphorus metabolism in Chlamydomonas. Proc. Natl. Acad. Sci. USA 1999, 96, 15336–15341. [Google Scholar]
- Nilsson, L.; Muller, R.; Nielsen, T.H. Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana. Plant Cell Environ 2007, 30, 1499–1512. [Google Scholar]
- Stefanovic, A.; Ribot, C.; Rouached, H.; Wang, Y.; Chong, J.; Belbahri, L.; Delessert, S.; Poirier, Y. Member of the PHO1 gene family show limited functional redundancy in phosphate transfer to the shoot, and are regulated by phosphate deficiency via distinct pathways. Plant J 2007, 50, 982, –994.. [Google Scholar]
- Ribot, C.; Wang, Y.; Poirier, Y. Expression analyses of three members of the AtPHO1 family reveal differential interactions between signaling pathways involved in phosphate deficiency and the responses to auxin, cytokinin, and abscisic acid. Planta 2008, 227, 1025, –1036.. [Google Scholar]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Ann. Rev. Plant Biol 2006, 57, 9–53. [Google Scholar]
- Lin, S.I.; Chiang, S.F.; Lin, W.Y.; Chen, J.W.; Tseng, C.Y. Regulatory network of microRNA399 and PHO2 by systemic signaling. Plant Physiol 2008, 147, 732–746. [Google Scholar]
- Pant, B.D.; Buhtz, A.; Kehr, J.; Scheible, W.R. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. Plant J 2008, 53, 731–738. [Google Scholar]
- Buhtz, A.; Pieritz, J.; Springer, F.; Kehr, J. Phloem small RNAs, nutrient stress responses, and systemic mobility. BMC Plant Biol 2010, 10, 64, :1–64:13.. [Google Scholar]
- Fujii, H.; Chiou, T.Z.; Lin, S.I.; Aung, K.; Zhu, J.K. A miRNA involved in phosphate starvation response in Arabidopsis. Curr. Biol 2005, 15, 2038–2043. [Google Scholar]
- Aung, K.; Lin, S.I.; Wu, C.C.; Huang, Y.T.; Su, C.L.; Chiou, T.J. pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a microRNA399 target gene. Plant Physiol. 2006, 141, 1000–1011. [Google Scholar]
- Bari, R.; Datt, P.; Stitt, M.; Scheible, W.R. PHO2, MicroRNA 399 and PHR1 define a phosphate signaling pathway in plants. Plant Physiol 2006, 141, 988–999. [Google Scholar]
- Ciereszko, I.; Johansson, H.; Hurry, V.; Kleczkowski, L.A. Phosphate status affects the gene expression, protein content and enzymatic activity of UDP-glucose pyrophosphorylase in wild-type and pho mutants of Arabidopsis. Planta 2001, 212, 598–605. [Google Scholar]
- Franco-Zorrilla, J.M.; Valli, A.; Tudesco, M. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet 2007, 39, 1033–1037. [Google Scholar]
- Liu, F.; Wang, Z.; Ren, H.; Shen, C.; Li, Y.; Ling, H.Q.; Wu, C.; Lian, X.; Wu, P. OsSPX1 suppresses the function of OsPHR2 in the regulation of expression of OsPT2 and phosphate homeostasis in shoots of rice. Plant J 2010, 62, 508–517. [Google Scholar]
- Branscheid, A.; Sieh, D.; Pant, B.D.; May, P.; Devers, A. Expression pattern suggests a role of miR399 in the regulation of the cellular response to local Pi increase during arbuscular mycorrhizal symbiosis. Mol. Plant Microbe Interact. 2010, 23, 915–926. [Google Scholar]
- Valdés-López, O.; Arenas-Huertero, C.; Girard, L.; Vance, C.P.; Reyes, J.L.; Hernández, G. Essential role of MYB transcription factor: PvPHR1 miRNA: PvmiR399 in the phosphorus deficiency signaling in common bean roots. Plant Cell Environ 2008, 31, 1834–1843. [Google Scholar]
- Liu, J.Q.; Allan, D.L.; Vance, C.P. Systemic signaling and local sensing of phosphate in common bean: Cross-talk between photosynthate and microRNA399. Mol. Plant 2010, 3, 428–437. [Google Scholar]
- Broughton, W.J.; Hernández, G.; Blair, M.; Beeb, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar]
- Yan, X.; Beebe, S.E.; Lynch, J.P. Phosphorus efficiency in common bean genotypes in contrasting soil types: II. Yield response. Crop Sci 1995, 35, 1094–1099. [Google Scholar]
- Vadez, V.; Lasso, J.H.; Becck, D.P.; Drevon, J.J. Variability of N2-fixation in common bean (Phaseolus vulgaris) under P deficiency is related to P use efficiency. Euphytica 1999, 109, 231–242. [Google Scholar]
- Tang, C.; Hinsinger, P.; Jaillard, B.; Rengelz, Z.; Drevon, J.J. Effect of phosphorus deficiency on the growth, symbiotic N2 fixation and proton release by two bean (Phaseolus vulgaris) genotypes. Agronomie 2001, 21, 683–689. [Google Scholar]
- Tang, C.; Drevon, J.J.; Jaillard, B.; Souche, G.; Hinsinger, P. Proton release of two genotypes of bean (Phaseolus vulgaris L.) as affected by N nutrition and P deficiency. Plant Soil 2004, 260, 59–68. [Google Scholar]
- Drevon, J.J.; Alkama, N.; Araujo, A.; Beebe, B.; Aslan Attar, H.; Benoit, J.; Lopez, A.; Martinez-Romero, E.; Rodino, P.; Tajini, F.; et al. Nodular diagnosis for ecological engineering of the symbiotic nitrogen fixation with legumes. Proc. Environ. Sci 2011, 9, 40–46. [Google Scholar]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar]
- Gaume, A.; Mächler, F.; Frossard, E. Aluminum resistance in two cultivars of Zea may L.: Root exudation of organic acids and influence of phosphorous nutrition. Plant Soil 2001, 234, 73–81. [Google Scholar]
- Gilbert, G.A.; Knight, J.D.; Vance, C.P.; Allan, D.L. Proteoid root development of phosphorus deficient lupin is mimicked by auxin and phosphonate. Ann. Bot 2000, 85, 921–928. [Google Scholar]
- Ma, Z.; Baskin, T.I.; Brown, K.M.; Lynch, J.P. Regulation of root elongation under phosphorus stress involves changes in ethylene responsiveness. Plant Physiol 2003, 131, 1381–1390. [Google Scholar]
- Peláez, P.; Trejo, S.T.; Iñiguez, L.P.; Estrada-Navarrete, G.; Covarrubias, A.A.; Reyes, J.L.; Sánchez, F. Identification and characterization of microRNAs in Phaseolus vulgaris by high-throughput sequencing. BMC Genomics 2012, 13, 1–18. [Google Scholar]
- phytozome. Available online: http://www.phytozome.net (accessed on 20 October 2012).
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res 2012, 40, 1178–1186. [Google Scholar]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar]
- Summerfield, R.J.; Huxley, P.A.; Minchin, F.R. Plant husbandry and management techniques for growing grain legumes under simulated tropical conditions in controlled environments. Exp. Agric 1977, 13, 113–121. [Google Scholar]
- The Computational Biology and Functional Genomics Laboratory. The Cene Index Project. Available online: http://compbio.dfci.harvard.edu/tgi/V4.0 (accessed on 15 January 2012).
- psRNATarget: A Plant Small RNA Target Analysis Server. Available online: http://plantgrn.noble.org/psRNATarget/ (accessed on 10 October 2012).
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res 2011, 39, 1–5. [Google Scholar]
- Arenas-Huertero, C.; Perez, B.; Rabanal, F.; Blanco-Melo, D.; de la Rosa, C.; Estrada-Navarrete, G.; Sánchez, F.; Covarrubias, A.A.; Reyes, J.L. Conserved and novel miRNAs in the legume Phaseolus vulgaris in response to stress. Plant Mol. Biol 2009, 70, 385–401. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | PvmiR399 binding site PHO2 5′ UTR region | BAT477 miRNA:mRNA pairing (score) | DOR364 miRNA:mRNA pairing (score) |
|---|---|---|---|
| PvmiR399a | 1 | I-IIIIII-IIIIIIIIIIII (2.0) | I-IIIII--IIII-IIIIIII (4.0) |
| PvmiR399b | 1 | I-IIIIIIIIIIIIIIIIIII (1.0) | I-IIIII-:IIII-IIIIIII (3.5) |
| PvmiR399e | 1 | IIIIIIII-IIIIIIIIIIII (1.0) | IIIIIII--IIII-IIIIIII (3.0) |
| PvmiR399a | 2 | IIIII-IIIIII-IIII--II (4.0) | IIIII-IIIIII-IIII--II (4.0) |
| PvmiR399b | 2 | IIIIII---III-IIII--II (6.0) | IIIIII---III-IIII--II (6.0) |
| PvmiR399e | 2 | I-III-IIIIII-IIII--II (5.0) | I-III-IIIIII-IIII--II (5.0) |
| PvmiR399a | 3 | I-IIIIIIIIIIIIIIIIIII (1.0) | I-IIIIIIIIIIIIIIIIIII (1.0) |
| PvmiR399b | 3 | I-IIIIIIIII-IIIIIIIII (2.0) | I-IIIIIIIII-IIIIIIIII (2.0) |
| P-miR399e | 3 | IIIIIIIIIIIIIIIIIIIII (0.0) | IIIIIIIIIIIIIIIIIIIII (0.0) |
| PvmiR399a | 4 | I--II:IIIIIIIIIIIIII: (3.0) | I--II:IIIIIIIIIIIIII: (3.0) |
| PvmiR399b | 4 | I--II:II-IIIIIIIIIII: (4.0) | I--II:II-IIIIIIIIIII: (4.0) |
| PvmiR399e | 4 | I--II:IIIIIIIIIIIIII: (3.0) | I--II:IIIIIIIIIIIIII: (3.0) |
| PvmiR399a | 5 | I--IIIIIIIIIIIIIII-II (3.0) | I--IIIIIIIIIIIIIII-II (3.0) |
| PvmiR399b | 5 | I--IIIII-IIIIIIIII-II (4.0) | I--IIIII-IIIIIIIII-II (4.0) |
| PvmiR399e | 5 | I--IIIIIIIIIIIIIII-II (3.0) | I--IIIIIIIIIIIIIII-II (3.0) |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ramírez, M.; Flores-Pacheco, G.; Reyes, J.L.; Luz Alvarez, A.; Drevon, J.J.; Girard, L.; Hernández, G. Two Common Bean Genotypes with Contrasting Response to Phosphorus Deficiency Show Variations in the microRNA 399-Mediated PvPHO2 Regulation within the PvPHR1 Signaling Pathway. Int. J. Mol. Sci. 2013, 14, 8328-8344. https://doi.org/10.3390/ijms14048328
Ramírez M, Flores-Pacheco G, Reyes JL, Luz Alvarez A, Drevon JJ, Girard L, Hernández G. Two Common Bean Genotypes with Contrasting Response to Phosphorus Deficiency Show Variations in the microRNA 399-Mediated PvPHO2 Regulation within the PvPHR1 Signaling Pathway. International Journal of Molecular Sciences. 2013; 14(4):8328-8344. https://doi.org/10.3390/ijms14048328
Chicago/Turabian StyleRamírez, Mario, Gerardo Flores-Pacheco, José Luis Reyes, Ana Luz Alvarez, Jean Jacques Drevon, Lourdes Girard, and Georgina Hernández. 2013. "Two Common Bean Genotypes with Contrasting Response to Phosphorus Deficiency Show Variations in the microRNA 399-Mediated PvPHO2 Regulation within the PvPHR1 Signaling Pathway" International Journal of Molecular Sciences 14, no. 4: 8328-8344. https://doi.org/10.3390/ijms14048328