Abnormal Skeletal Growth in Adolescent Idiopathic Scoliosis Is Associated with Abnormal Quantitative Expression of Melatonin Receptor, MT2

Abstract
:1. Introduction
2. Results and Discussion
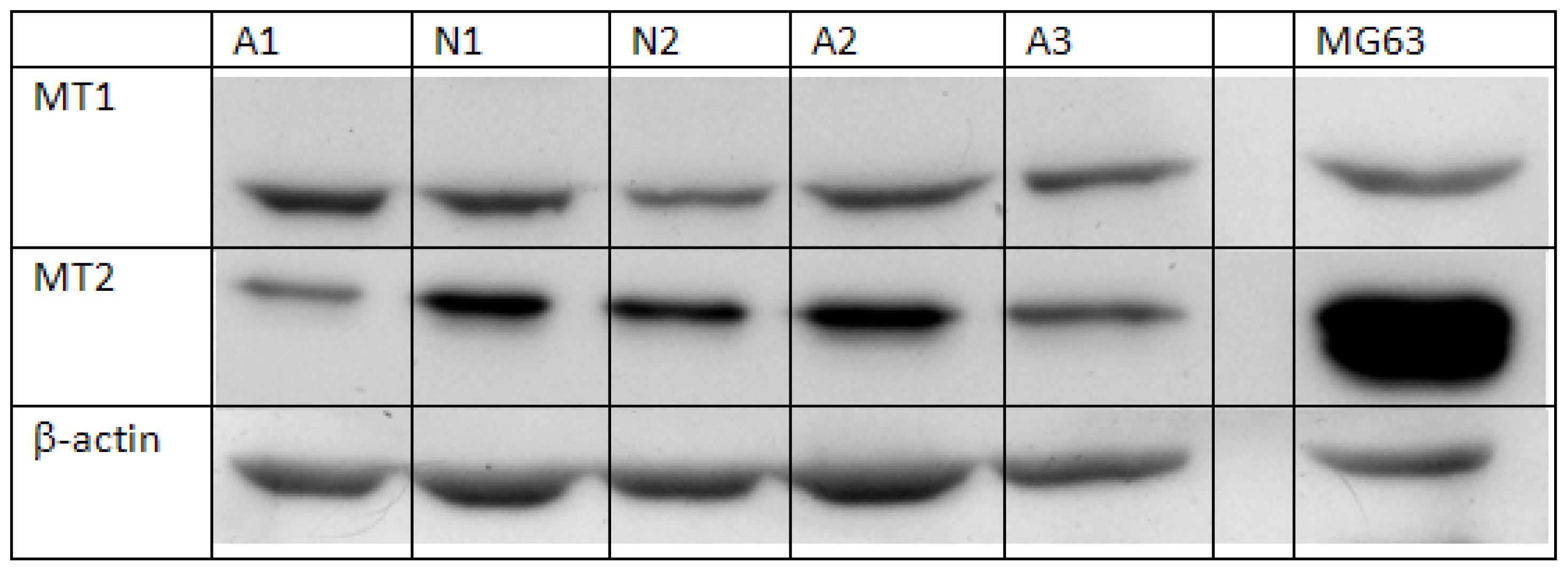
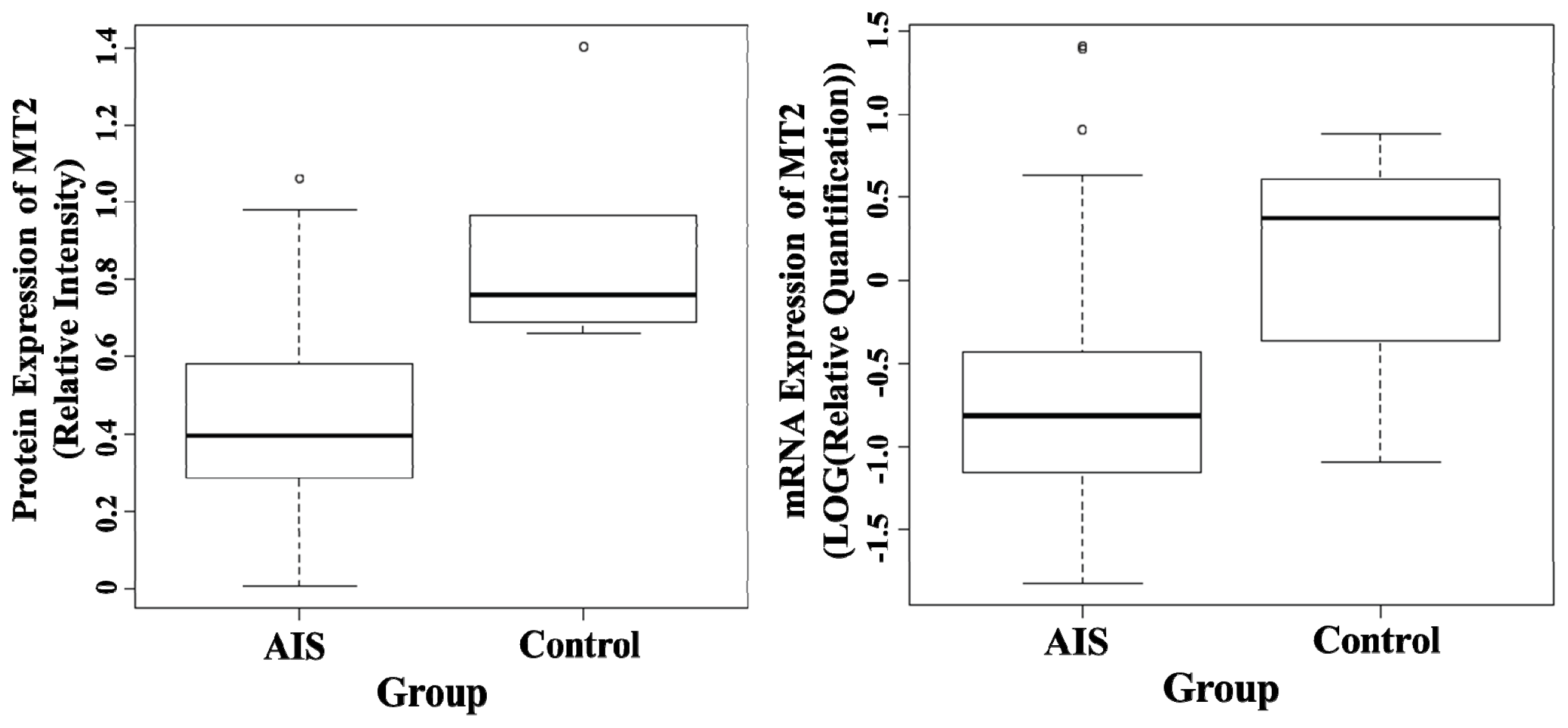
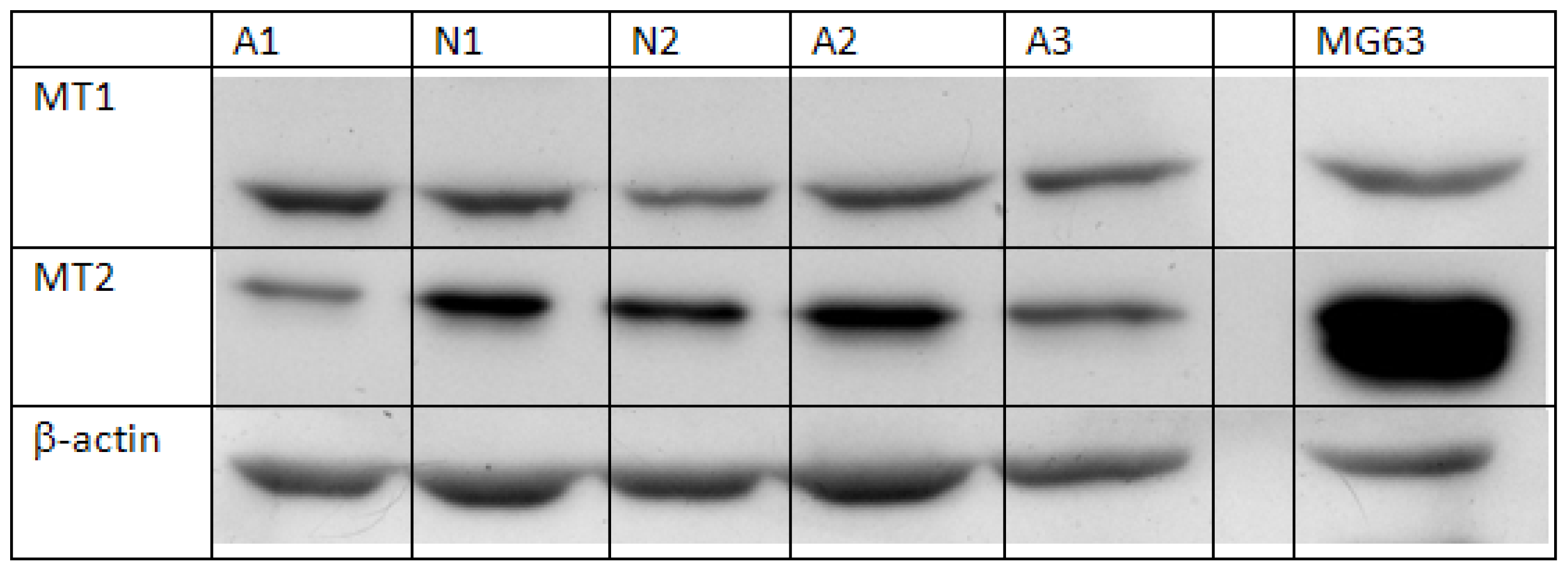
2.1. Semi-Quantification of Protein Expression Levels of MT1 and MT2 in Osteoblast
2.2. Quantification of mRNA Expression Levels of MT1 and MT2 in Osteoblast
2.3. Evaluation and Correlation of Anthropometric Parameters with Melatonin Receptors Expression Level
3. Experimental Section
3.1. Subjects Recruitments
3.2. Cell Isolation and Osteoblast Culture
3.3. Semi-Quantification of Protein Expression of MT1 and MT2 in Osteoblasts
3.4. Quantification of mRNA Expression of MT1 and MT2 in Osteoblast
3.5. Evaluation and Correlation of Anthropometric Parameters with Melatonin Receptors Expression
3.6. Data Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Wang, W.J.; Yeung, H.Y.; Chu, W.C.; Tang, N.L.; Lee, K.M.; Qiu, Y.; Burwell, R.G.; Cheng, J.C. Top theories for the etiopathogenesis of adolescent idiopathic scoliosis. J. Pediatr. Orthop 2011, 31, S14–S27. [Google Scholar]
- Weinstein, S.L.; Dolan, L.A.; Cheng, J.C.; Danielsson, A.; Morcuende, J.A. Adolescent idiopathic scoliosis. Lancet 2008, 371, 1527–1537. [Google Scholar]
- Hardeland, R. Neurobiology, pathophysiology, and treatment of melatonin deficiency and dysfunction. Sci. World J 2012, 2012, 640389. [Google Scholar]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J 2006, 273, 2813–2838. [Google Scholar]
- Armstrong, S.M. Melatonin and circadian control in mammals. Experientia 1989, 45, 932–938. [Google Scholar]
- Scheer, F.A.; van Montfrans, G.A.; van Someren, E.J.; Mairuhu, G.; Buijs, R.M. Daily nighttime melatonin reduces blood pressure in male patients with essential hypertension. Hypertension 2004, 43, 192–197. [Google Scholar]
- Guerrero, J.M.; Reiter, R.J. Melatonin-immune system relationships. Curr. Top. Med. Chem 2002, 2, 167–179. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Maldonado, M.D. Melatonin as an antioxidant: Physiology vs. pharmacology. J. Pineal Res 2005, 39, 215–216. [Google Scholar]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res 2011, 51, 1–16. [Google Scholar]
- Blask, D.E.; Dauchy, R.T.; Sauer, L.A. Putting cancer to sleep at night: The neuroendocrine/circadian melatonin signal. Endocrine 2005, 27, 179–188. [Google Scholar]
- Ladizesky, M.G.; Boggio, V.; Albornoz, L.E.; Castrillon, P.O.; Mautalen, C.; Cardinali, D.P. Melatonin increases oestradiol-induced bone formation in ovariectomized rats. J. Pineal Res 2003, 34, 143–151. [Google Scholar]
- Cardinali, D.P.; Ladizesky, M.G.; Boggio, V.; Cutrera, R.A.; Mautalen, C. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res 2003, 34, 81–87. [Google Scholar]
- Garcia-Maurino, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. Involvement of nuclear binding sites for melatonin in the regulation of IL-2 and IL-6 production by human blood mononuclear cells. J. Neuroimmunol 1998, 92, 76–84. [Google Scholar]
- Hardeland, R. Melatonin: Signaling mechanisms of a pleiotropic agent. Biofactors 2009, 35, 183–192. [Google Scholar]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron 1994, 13, 1177–1185. [Google Scholar]
- Reppert, S.M.; Godson, C.; Mahle, C.D.; Weaver, D.R.; Slaugenhaupt, S.A.; Gusella, J.F. Molecular characterization of a second melatonin receptor expressed in human retina and brain: The Mel1b melatonin receptor. Proc. Natl. Acad. Sci. USA 1995, 92, 8734–8738. [Google Scholar]
- Fujieda, H.; Hamadanizadeh, S.A.; Wankiewicz, E.; Pang, S.F.; Brown, G.M. Expression of mt1 melatonin receptor in rat retina: Evidence for multiple cell targets for melatonin. Neuroscience 1999, 93, 793–799. [Google Scholar]
- Meyer, P.; Pache, M.; Loeffler, K.U.; Brydon, L.; Jockers, R.; Flammer, J.; Wirz-Justice, A.; Savaskan, E. Melatonin MT-1-receptor immunoreactivity in the human eye. Br. J. Ophthalmol 2002, 86, 1053–1057. [Google Scholar]
- Savaskan, E.; Wirz-Justice, A.; Olivieri, G.; Pache, M.; Krauchi, K.; Brydon, L.; Jockers, R.; Muller-Spahn, F.; Meyer, P. Distribution of melatonin MT1 receptor immunoreactivity in human retina. J. Histochem. Cytochem 2002, 50, 519–526. [Google Scholar]
- Kong, X.; Li, X.; Cai, Z.; Yang, N.; Liu, Y.; Shu, J.; Pan, L.; Zuo, P. Melatonin regulates the viability and differentiation of rat midbrain neural stem cells. Cell Mol. Neurobiol 2008, 28, 569–579. [Google Scholar]
- Uz, T.; Arslan, A.D.; Kurtuncu, M.; Imbesi, M.; Akhisaroglu, M.; Dwivedi, Y.; Pandey, G.N.; Manev, H. The regional and cellular expression profile of the melatonin receptor MT1 in the central dopaminergic system. Brain Res. Mol. Brain Res 2005, 136, 45–53. [Google Scholar]
- Achong, R.; Nishimura, I.; Ramachandran, H.; Howell, T.H.; Fiorellini, J.P.; Karimbux, N.Y. Membrane type (MT) 1-matrix metalloproteinase (MMP) and MMP-2 expression in ligature-induced periodontitis in the rat. J. Periodontol 2003, 74, 494–500. [Google Scholar]
- Dillon, D.C.; Easley, S.E.; Asch, B.B.; Cheney, R.T.; Brydon, L.; Jockers, R.; Winston, J.S.; Brooks, J.S.; Hurd, T.; Asch, H.L. Differential expression of high-affinity melatonin receptors (MT1) in normal and malignant human breast tissue. Am. J. Clin. Pathol 2002, 118, 451–458. [Google Scholar]
- Lanoix, D.; Ouellette, R.; Vaillancourt, C. Expression of melatoninergic receptors in human placental choriocarcinoma cell lines. Hum. Reprod 2006, 21, 1981–1989. [Google Scholar]
- Naji, L.; Carrillo-Vico, A.; Guerrero, J.M.; Calvo, J.R. Expression of membrane and nuclear melatonin receptors in mouse peripheral organs. Life Sci 2004, 74, 2227–2236. [Google Scholar]
- Von Gall, C.; Stehle, J.H.; Weaver, D.R. Mammalian melatonin receptors: Molecular biology and signal transduction. Cell Tissue Res 2002, 309, 151–162. [Google Scholar]
- Vanecek, J. Cellular mechanisms of melatonin action. Physiol. Rev 1998, 78, 687–721. [Google Scholar]
- Park, K.H.; Kang, J.W.; Lee, E.M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin promotes osteoblastic differentiation through the BMP/ERK/Wnt signaling pathways. J. Pineal Res 2011, 51, 187–194. [Google Scholar]
- Sethi, S.; Radio, N.M.; Kotlarczyk, M.P.; Chen, C.T.; Wei, Y.H.; Jockers, R.; Witt-Enderby, P.A. Determination of the minimal melatonin exposure required to induce osteoblast differentiation from human mesenchymal stem cells and these effects on downstream signaling pathways. J. Pineal Res 2010, 49, 222–238. [Google Scholar]
- Roth, J.A.; Kim, B.G.; Lin, W.L.; Cho, M.I. Melatonin promotes osteoblast differentiation and bone formation. J. Biol. Chem 1999, 274, 22041–22047. [Google Scholar]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Tan, D.X.; Reiter, R.J. Scientific basis for the potential use of melatonin in bone diseases: osteoporosis and adolescent idiopathic scoliosis. J. Osteoporos 2010, 2010, 830231. [Google Scholar]
- Cheng, J.C.; Guo, X.; Sher, A.H. Persistent osteopenia in adolescent idiopathic scoliosis. A longitudinal follow up study. Spine 1999, 24, 1218–1222. [Google Scholar]
- Cheng, J.C.; Guo, X. Osteopenia in adolescent idiopathic scoliosis. A primary problem or secondary to the spinal deformity? Spine 1997, 22, 1716–1721. [Google Scholar]
- Cheung, K.M.; Wang, T.; Poon, A.M.; Carl, A.; Tranmer, B.; Hu, Y.; Luk, K.D.; Leong, J.C. The effect of pinealectomy on scoliosis development in young nonhuman primates. Spine 2005, 30, 2009–2013. [Google Scholar]
- Fjelldal, P.G.; Grotmol, S.; Kryvi, H.; Gjerdet, N.R.; Taranger, G.L.; Hansen, T.; Porter, M.J.; Totland, G.K. Pinealectomy induces malformation of the spine and reduces the mechanical strength of the vertebrae in Atlantic salmon, Salmo salar. J. Pineal Res 2004, 36, 132–139. [Google Scholar]
- Machida, M.; Dubousset, J.; Yamada, T.; Kimura, J.; Saito, M.; Shiraishi, T.; Yamagishi, M. Experimental scoliosis in melatonin-deficient C57BL/6J mice without pinealectomy. J. Pineal Res 2006, 41, 1–7. [Google Scholar]
- O’Kelly, C.; Wang, X.; Raso, J.; Moreau, M.; Mahood, J.; Zhao, J.; Bagnall, K. The production of scoliosis after pinealectomy in young chickens, rats, and hamsters. Spine 1999, 24, 35–43. [Google Scholar]
- Turgut, M.; Yenisey, C.; Uysal, A.; Bozkurt, M.; Yurtseven, M.E. The effects of pineal gland transplantation on the production of spinal deformity and serum melatonin level following pinealectomy in the chicken. Eur. Spine J 2003, 12, 487–494. [Google Scholar]
- Bagnall, K.; Raso, J.; Moreau, M.; Mahood, J.; Wang, X.; Beuerlein, M. The development of scoliosis following pinealectomy in young chickens is not the result of an artifact of the surgical procedure. Stud. Health Technol. Inform 2002, 88, 3–9. [Google Scholar]
- Thillard, M.J. Vertebral column deformities following epiphysectomy in the chick. C. R. Hebd. Seances Acad. Sci 1959, 248, 1238–1240. [Google Scholar]
- Machida, M.; Murai, I.; Miyashita, Y.; Dubousset, J.; Yamada, T.; Kimura, J. Pathogenesis of idiopathic scoliosis. Experimental study in rats. Spine 1999, 24, 1985–1989. [Google Scholar]
- Machida, M.; Dubousset, J.; Imamura, Y.; Miyashita, Y.; Yamada, T.; Kimura, J. Melatonin. A possible role in pathogenesis of adolescent idiopathic scoliosis. Spine 1996, 21, 1147–1152. [Google Scholar]
- Bagnall, K.M.; Raso, V.J.; Hill, D.L.; Moreau, M.; Mahood, J.K.; Jiang, H.; Russell, G.; Bering, M.; Buzzell, G.R. Melatonin levels in idiopathic scoliosis. Diurnal and nocturnal serum melatonin levels in girls with adolescent idiopathic scoliosis. Spine 1996, 21, 1974–1978. [Google Scholar]
- Brodner, W.; Krepler, P.; Nicolakis, M.; Langer, M.; Kaider, A.; Lack, W.; Waldhauser, F. Melatonin and adolescent idiopathic scoliosis. J. Bone Joint Surg. Br 2000, 82, 399–403. [Google Scholar]
- Moreau, A.; Wang, D.S.; Forget, S.; Azeddine, B.; Angeloni, D.; Fraschini, F.; Labelle, H.; Poitras, B.; Rivard, C.H.; Grimard, G. Melatonin signaling dysfunction in adolescent idiopathic scoliosis. Spine 2004, 29, 1772–1781. [Google Scholar]
- Azeddine, B.; Letellier, K.; Wang da, S.; Moldovan, F.; Moreau, A. Molecular determinants of melatonin signaling dysfunction in adolescent idiopathic scoliosis. Clin. Orthop. Relat. Res 2007, 462, 45–52. [Google Scholar]
- Qiu, X.S.; Tang, N.L.; Yeung, H.Y.; Lee, K.M.; Hung, V.W.; Ng, B.K.; Ma, S.L.; Kwok, R.H.; Qin, L.; Qiu, Y.; et al. Melatonin receptor 1B (MTNR1B) gene polymorphism is associated with the occurrence of adolescent idiopathic scoliosis. Spine 2007, 32, 1748–1753. [Google Scholar]
- Man, G.C.; Wang, W.W.; Yeung, B.H.; Lee, S.K.; Ng, B.K.; Hung, W.Y.; Wong, J.H.; Ng, T.B.; Qiu, Y.; Cheng, J.C. Abnormal proliferation and differentiation of osteoblasts from girls with adolescent idiopathic scoliosis to melatonin. J. Pineal Res 2010, 49, 69–77. [Google Scholar]
- Man, G.C.; Wong, J.H.; Wang, W.W.; Sun, G.Q.; Yeung, B.H.; Ng, T.B.; Lee, S.K.; Ng, B.K.; Qiu, Y.; Cheng, J.C. Abnormal melatonin receptor 1B expression in osteoblasts from girls with adolescent idiopathic scoliosis. J. Pineal Res 2011, 50, 395–402. [Google Scholar]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol 2011, 93, 350–384. [Google Scholar]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol 2012, 351, 152–166. [Google Scholar]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J. Pineal Res 2006, 40, 332–342. [Google Scholar]
- Shyy, W.; Wang, K.; Gurnett, C.A.; Dobbs, M.B.; Miller, N.H.; Wise, C.; Sheffield, V.C.; Morcuende, J.A. Evaluation of GPR50, hMel-1B, and ROR-alpha melatonin-related receptors and the etiology of adolescent idiopathic scoliosis. J. Pediatr. Orthop 2010, 30, 539–543. [Google Scholar]
- Nelson, L.M.; Ward, K.; Ogilvie, J.W. Genetic variants in melatonin synthesis and signaling pathway are not associated with adolescent idiopathic scoliosis. Spine 2011, 36, 37–40. [Google Scholar]
- Morocz, M.; Czibula, A.; Grozer, Z.B.; Szecsenyi, A.; Almos, P.Z.; Rasko, I.; Illes, T. Association study of BMP4, IL6, Leptin, MMP3, and MTNR1B gene promoter polymorphisms and adolescent idiopathic scoliosis. Spine 2011, 36, E123–130. [Google Scholar]
- Takahashi, Y.; Matsumoto, M.; Karasugi, T.; Watanabe, K.; Chiba, K.; Kawakami, N.; Tsuji, T.; Uno, K.; Suzuki, T.; Ito, M.; et al. Lack of association between adolescent idiopathic scoliosis and previously reported single nucleotide polymorphisms in MATN1, MTNR1B, TPH1, and IGF1 in a Japanese population. J. Orthop. Res. Off. Publ. Orthop. Res. Soc 2011, 29, 1055–1058. [Google Scholar]
- Slominski, A.; Pisarchik, A.; Zbytek, B.; Tobin, D.J.; Kauser, S.; Wortsman, J. Functional activity of serotoninergic and melatoninergic systems expressed in the skin. J. Cell Physiol 2003, 196, 144–153. [Google Scholar]
- Cheng, J.C.; Leung, S.S.; Lau, J. Anthropometric measurements and body proportions among Chinese children. Clin. Orthop. Relat. Res. 1996, 22–30. [Google Scholar]
- Johnson, B.E.; Westgate, H.D. Methods of predicting vital capacity in patients with thoracic scoliosis. J. Bone Joint Surg. Am 1970, 52, 1433–1439. [Google Scholar]
- Brown, J.K.; Whittemore, K.T.; Knapp, T.R. Is arm span an accurate measure of height in young and middle-age adults? Clin. Nurs. Res 2000, 9, 84–94. [Google Scholar]
- Siu King Cheung, C.; Tak Keung Lee, W.; Kit Tse, Y.; Ping Tang, S.; Man Lee, K.; Guo, X.; Qin, L.; Chun Yiu Cheng, J. Abnormal peri-pubertal anthropometric measurements and growth pattern in adolescent idiopathic scoliosis: A study of 598 patients. Spine 2003, 28, 2152–2157. [Google Scholar]
- Starcevic-Klasan, G.; Cvijanovic, O.; Peharec, S.; Zulle, M.; Arbanas, J.; Ivancic Jokic, N.; Bakarcic, D.; Malnar-Dragojevic, D.; Bobinac, D. Anthropometric parameters as predictors for iliopsoas muscle strength in healthy girls and in girls with adolescent idiopathic scoliosis. Coll. Antropol 2008, 32, 461–466. [Google Scholar]
- Yim, A.P.Y.; Yeung, H.Y.; Hung, V.W.; Lee, K.M.; Lam, T.P.; Ng, B.K.; Qiu, Y.; Cheng, J.C. Abnormal skeletal growth patterns in adolescent idiopathic scoliosis-a longitudinal study until skeletal maturity. Spine 2012, 37, E1148–E1154. [Google Scholar]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type I collagen synthesis in human bone cells in vitro. J. Pineal Res 1999, 27, 106–110. [Google Scholar]
- Toma, C.D.; Svoboda, M.; Arrich, F.; Ekmekcioglu, C.; Assadian, O.; Thalhammer, T. Expression of the melatonin receptor (MT) 1 in benign and malignant human bone tumors. J. Pineal Res 2007, 43, 206–213. [Google Scholar]

{kind=link}
{kind=link}
| Expression of | AIS (n = 41) (mean ± SD) | Control (n = 9) (mean ± SD) | p-value | |
|---|---|---|---|---|
| Protein (Relative intensity) | MT1 | 0.797 ± 0.199 | 0.793 ± 0.322 | 0.978 |
| MT2 | 0.429 ± 0.276 | 0.881 ± 0.317 | <0.01 * | |
| mRNA [Log (Relative quantification)] | MT1 | −0.378 ± 0.258 | −0.343 ± 0.225 | 0.707 |
| MT2 | −0.627 ± 0.742 | 0.129 ± 0.70 | 0.019 * |
| AIS patients (n = 38) mean ± SD † | z-score mean ± SD | p-value | |
|---|---|---|---|
| BW (kg) | 46.46 ± 6.36 | −0.15 ± 0.91 | 0.29 |
| BMI | 18.37 ± 2.16 | −0.45 ± 0.15 | <0.01 * |
| Arm Span (cm) | 158.96 ± 6.13 | 0.41 ± 0.83 | <0.01 * |
| Melatonin receptor MT1 (n = 38) | |||
|---|---|---|---|
| Group | Normal expression (n = 32) | Low expression (n = 6) | p-value |
| z-score of | |||
| BW | −0.136 ± 0.98 | −0.070 ± 0.62 | 0.875 |
| BMI | −0.429 ± 0.98 | −0.384 ± 0.67 | 0.915 |
| Arm span | 0.412 ± 0.92 | 0.471 ± 0.28 | 0.768 |
| Expression of mRNA (Log (relative quantification)) | −0.311 ± 0.18 | −0.751 ± 0.078 | <0.01 |
| Age | 15.3 ± 2.1 | 15.2 ± 1.5 | 0.940 |
| Melatonin receptor MT2 (n= 38) | |||
| Group | Normal expression (n= 27) | Low expression (n= 11) | p-value |
| z-score of | |||
| BW | −0.223 ± 0.83 | 0.112 ± 1.13 | 0.318 |
| BMI | −0.405 ± 0.83 | −0.464 ± 1.18 | 0.861 |
| Arm Span | 0.238 ± 0.79 | 0.870 ± 0.86 | 0.036 * |
| Expression of mRNA (Log (relative quantification)) | −0.367 ± 0.72 | 1.311 ± 0.21 | <0.01 |
| Age | 15.1 ± 1.9 | 15.7 ± 2.0 | 0.414 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yim, A.P.-y.; Yeung, H.-y.; Sun, G.; Lee, K.-m.; Ng, T.-b.; Lam, T.-p.; Ng, B.K.-w.; Qiu, Y.; Moreau, A.; Cheng, J.C.-y. Abnormal Skeletal Growth in Adolescent Idiopathic Scoliosis Is Associated with Abnormal Quantitative Expression of Melatonin Receptor, MT2. Int. J. Mol. Sci. 2013, 14, 6345-6358. https://doi.org/10.3390/ijms14036345
Yim AP-y, Yeung H-y, Sun G, Lee K-m, Ng T-b, Lam T-p, Ng BK-w, Qiu Y, Moreau A, Cheng JC-y. Abnormal Skeletal Growth in Adolescent Idiopathic Scoliosis Is Associated with Abnormal Quantitative Expression of Melatonin Receptor, MT2. International Journal of Molecular Sciences. 2013; 14(3):6345-6358. https://doi.org/10.3390/ijms14036345
Chicago/Turabian StyleYim, Annie Po-yee, Hiu-yan Yeung, Guangquan Sun, Kwong-man Lee, Tzi-bun Ng, Tsz-ping Lam, Bobby Kin-wah Ng, Yong Qiu, Alain Moreau, and Jack Chun-yiu Cheng. 2013. "Abnormal Skeletal Growth in Adolescent Idiopathic Scoliosis Is Associated with Abnormal Quantitative Expression of Melatonin Receptor, MT2" International Journal of Molecular Sciences 14, no. 3: 6345-6358. https://doi.org/10.3390/ijms14036345