The Lipid Transfer Protein StarD7: Structure, Function, and Regulation

Abstract
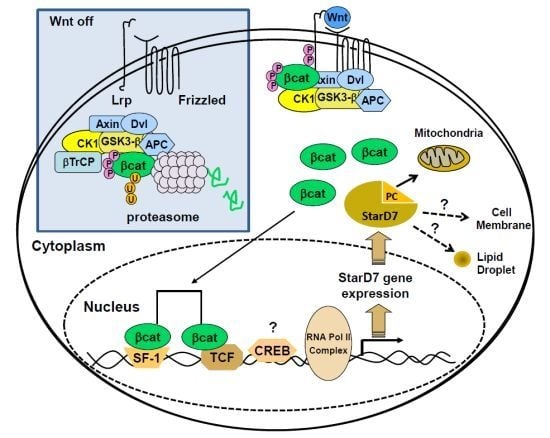
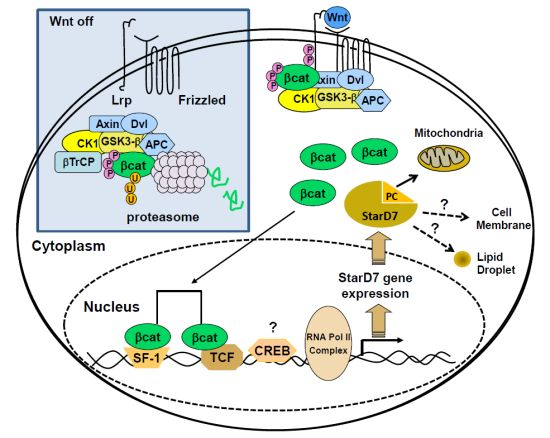
:
1. Introduction
The START Domain Protein Family
2. Structural and Biochemical Characteristics of StarD7
3. StarD7-Lipid Interaction
4. StarD7 Expression
5. Transcriptional Regulation of StarD7
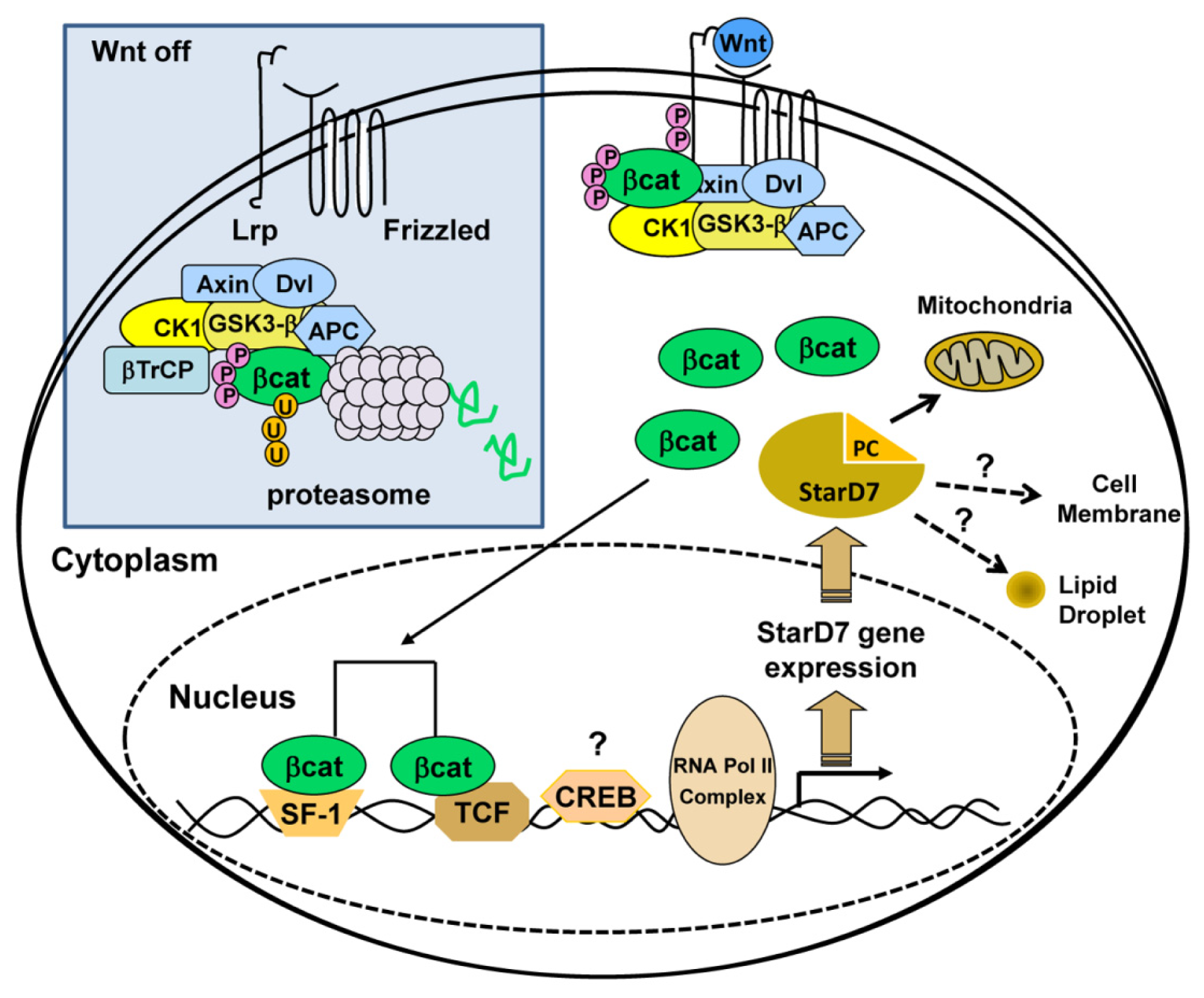
5.1. Regulation of StarD7 Expression by β-Catenin/T Cell-Specific Transcription Factor 4
5.2. Regulation of StarD7 Expression by cAMP and Steroidogenic Factor 1
5.3. Cross-Talk between β-Catenin/TCF4 and SF-1 Pathways
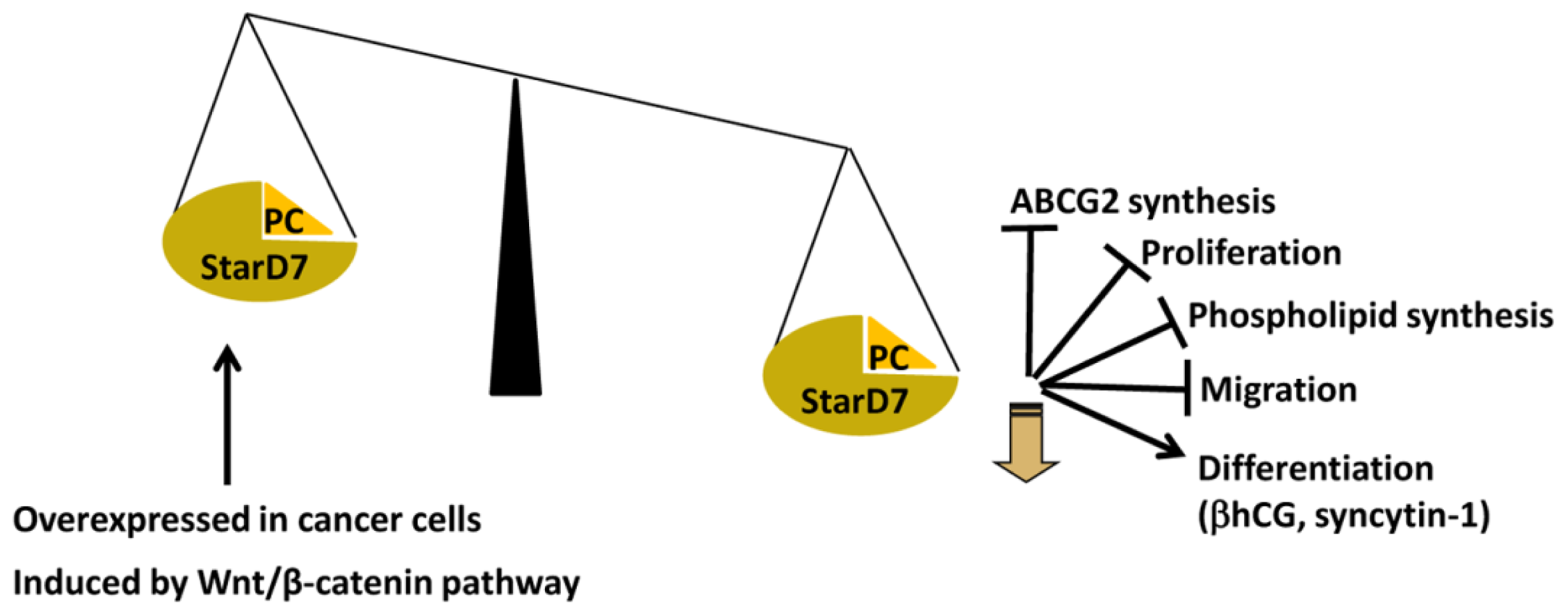
6. StarD7 Modulates Trophoblast Physiology
6.1. StarD7 Modulates ATP-Binding Cassette Subfamily G (WHITE) Member 2 Expression
6.2. StarD7 Modulates Cell Migration, Proliferation, and Differentiation
7. Conclusions
Acknowledgments
Conflict of Interest
References
- Prinz, W.A. Lipid trafficking sans vesicles: Where, why, how? Cell 2010, 143, 870–874. [Google Scholar]
- Holthuis, J.C.; Levine, T.P. Lipid traffic: Floppy drives and a superhighway. Nat. Rev. Mol. Cell Biol 2005, 6, 209–220. [Google Scholar]
- Soccio, R.E.; Breslow, J.L. StAR-Related lipid transfer (START) proteins: Mediators of intracellular lipid metabolism. J. Biol. Chem 2003, 278, 22183–22186. [Google Scholar]
- Thorsell, A.G.; Lee, W.H.; Persson, C.; Siponen, M.I.; Nilsson, M.; Busam, R.D.; Kotenyova, T.; Schuler, H.; Lehtio, L. Comparative structural analysis of lipid binding START domains. PLoS One 2011, 6, e19521. [Google Scholar]
- Lee, H.; Li, Z.; Silkov, A.; Fischer, M.; Petrey, D.; Honig, B.; Murray, D. High-Throughput computational structure-based characterization of protein families: START domains and implications for structural genomics. J. Struct. Funct. Genomics 2010, 11, 51–59. [Google Scholar]
- Ponting, C.P.; Aravind, L. START: A lipid-binding domain in StAR, HD-ZIP and signalling proteins. Trends Biochem. Sci 1999, 24, 130–132. [Google Scholar]
- Tsujishita, Y.; Hurley, J.H. Structure and lipid transport mechanism of a StAR-related domain. Nat. Struct. Biol 2000, 7, 408–414. [Google Scholar]
- Barros, M.H.; Johnson, A.; Gin, P.; Marbois, B.N.; Clarke, C.F.; Tzagoloff, A. The Saccharomyces cerevisiae COQ10 gene encodes a START domain protein required for function of coenzyme Q in respiration. J. Biol. Chem 2005, 280, 42627–42635. [Google Scholar]
- Mercier, K.A.; Mueller, G.A.; Acton, T.B.; Xiao, R.; Montelione, G.T.; Powers, R. (1)H, (13)C, and (15)N NMR assignments for the Bacillus subtilis yndB START domain. Biomol. NMR Assign. 2009, 3, 191–194. [Google Scholar]
- Schrick, K.; Nguyen, D.; Karlowski, W.M.; Mayer, K.F. START lipid/sterol-binding domains are amplified in plants and are predominantly associated with homeodomain transcription factors. Genome Biol 2004, 5, R41. [Google Scholar]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Adaptations of the helix-grip fold for ligand binding and catalysis in the START domain superfamily. Proteins 2001, 43, 134–144. [Google Scholar]
- Alpy, F.; Tomasetto, C. Give lipids a START: The StAR-related lipid transfer (START) domain in mammals. J. Cell Sci 2005, 118, 2791–2801. [Google Scholar]
- Stocco, D.M. StAR protein and the regulation of steroid hormone biosynthesis. Annu. Rev. Physiol 2001, 63, 193–213. [Google Scholar]
- Miller, W.L.; Bose, H.S. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res 2011, 52, 2111–2135. [Google Scholar]
- Romanowski, M.J.; Soccio, R.E.; Breslow, J.L.; Burley, S.K. Crystal structure of the Mus musculus cholesterol-regulated START protein 4 (StarD4) containing a StAR-related lipid transfer domain. Proc. Natl. Acad. Sci. USA 2002, 99, 6949–6954. [Google Scholar]
- Roderick, S.L.; Chan, W.W.; Agate, D.S.; Olsen, L.R.; Vetting, M.W.; Rajashankar, K.R.; Cohen, D.E. Structure of human phosphatidylcholine transfer protein in complex with its ligand. Nat. Struct. Biol 2002, 9, 507–511. [Google Scholar]
- Chan, W.W.; Roderick, S.L.; Cohen, D.E. Human phosphatidylcholine transfer protein: Purification, crystallization and preliminary X-ray diffraction data. Biochim. Biophys. Acta 2002, 1596, 1–5. [Google Scholar]
- Li, H.; Fung, K.L.; Jin, D.Y.; Chung, S.S.; Ching, Y.P.; Ng, I.O.; Sze, K.H.; Ko, B.C.; Sun, H. Solution structures, dynamics, and lipid-binding of the sterile alpha-motif domain of the deleted in liver cancer 2. Proteins 2007, 67, 1154–1166. [Google Scholar]
- Kudo, N.; Kumagai, K.; Matsubara, R.; Kobayashi, S.; Hanada, K.; Wakatsuki, S.; Kato, R. Crystal structures of the CERT START domain with inhibitors provide insights into the mechanism of ceramide transfer. J. Mol. Biol 2010, 396, 245–251. [Google Scholar]
- Lavigne, P.; Najmanivich, R.; Lehoux, J.G. Mammalian StAR-related lipid transfer (START) domains with specificity for cholesterol: Structural conservation and mechanism of reversible binding. Subcell. Biochem 2010, 51, 425–437. [Google Scholar]
- Chen, D.; Latham, J.; Zhao, H.; Bisoffi, M.; Farelli, J.; Dunaway-Mariano, D. Human brown fat inducible thioesterase variant 2 cellular localization and catalytic function. Biochemistry 2012, 51, 6990–6999. [Google Scholar]
- Durand, S.; Angeletti, S.; Genti-Raimondi, S. GTT1/StarD7, a novel phosphatidylcholine transfer protein-like highly expressed in gestational trophoblastic tumour: Cloning and characterization. Placenta 2004, 25, 37–44. [Google Scholar]
- Cohen, D.E.; Green, R.M.; Wu, M.K.; Beier, D.R. Cloning, tissue-specific expression, gene structure and chromosomal localization of human phosphatidylcholine transfer protein. Biochim. Biophys. Acta 1999, 1447, 265–270. [Google Scholar]
- Angeletti, S.; Rena, V.; Nores, R.; Fretes, R.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. Expression and localization of StarD7 in trophoblast cells. Placenta 2008, 29, 396–404. [Google Scholar]
- Leman, E.S.; Magheli, A.; Yong, K.M.; Netto, G.; Hinz, S.; Getzenberg, R.H. Identification of nuclear structural protein alterations associated with seminomas. J. Cell. Biochem 2009, 108, 1274–1279. [Google Scholar]
- Horibata, Y.; Sugimoto, H. StarD7 mediates the intracellular trafficking of phosphatidylcholine to mitochondria. J. Biol. Chem 2010, 285, 7358–7365. [Google Scholar]
- Kim, W.; Bennett, E.J.; Huttlin, E.L.; Guo, A.; Li, J.; Possemato, A.; Sowa, M.E.; Rad, R.; Rush, J.; Comb, M.J.; et al. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 2011, 44, 325–340. [Google Scholar]
- Hornbeck, P.V.; Kornhauser, J.M.; Tkachev, S.; Zhang, B.; Skrzypek, E.; Murray, B.; Latham, V.; Sullivan, M. PhosphoSitePlus: A comprehensive resource for investigating the structure and function of experimentally determined post-translational modifications in man and mouse. Nucleic Acids Res 2012, 40, D261–D270. [Google Scholar]
- Bose, H.S.; Whittal, R.M.; Ran, Y.; Bose, M.; Baker, B.Y.; Miller, W.L. StAR-Like activity and molten globule behavior of StARD6, a male germ-line protein. Biochemistry 2008, 47, 2277–2288. [Google Scholar]
- Rodriguez, M.; Li, S.S.; Harper, J.W.; Songyang, Z. An oriented peptide array library (OPAL) strategy to study protein-protein interactions. J. Biol. Chem 2004, 279, 8802–8807. [Google Scholar]
- Guo, L.; Ying, W.; Zhang, J.; Yuan, Y.; Qian, X.; Wang, J.; Yang, X.; He, F. Tandem affinity purification and identification of the human TSC1 protein complex. Acta. Biochim. Biophys. Sin. (Shanghai) 2010, 42, 266–273. [Google Scholar]
- Angeletti, S.; Maggio, B.; Genti-Raimondi, S. Surface activity and interaction of StarD7 with phospholipid monolayers. Biochem. Biophys. Res. Commun 2004, 314, 181–185. [Google Scholar]
- Angeletti, S.; Sanchez, J.M.; Chamley, L.W.; Genti-Raimondi, S.; Perillo, M.A. StarD7 behaves as a fusogenic protein in model and cell membrane bilayers. Biochim. Biophys. Acta 2011, 1818, 425–433. [Google Scholar]
- Penno, A.; Hackenbroich, G.; Thiele, C. Phospholipids and lipid droplets. Biochim. Biophys. Acta 2013, 1831, 589–594. [Google Scholar]
- Voelker, D.R. Genetic and biochemical analysis of non-vesicular lipid traffic. Annu. Rev. Biochem 2009, 78, 827–856. [Google Scholar]
- Zhu, X.M.; Han, T.; Sargent, I.L.; Yin, G.W.; Yao, Y.Q. Differential expression profile of microRNAs in human placentas from preeclamptic pregnancies vs normal pregnancies. Am. J. Obstet. Gynecol 2009, 200, 1–7. [Google Scholar]
- Polyak, K.; Hu, M. Do myoepithelial cells hold the key for breast tumor progression? J. Mammary Gland Biol. Neoplasia 2005, 10, 231–247. [Google Scholar]
- Jelinek, D.F.; Tschumper, R.C.; Stolovitzky, G.A.; Iturria, S.J.; Tu, Y.; Lepre, J.; Shah, N.; Kay, N.E. Identification of a global gene expression signature of B-chronic lymphocytic leukemia. Mol. Cancer Res 2003, 1, 346–361. [Google Scholar]
- Wiese, A.H.; Auer, J.; Lassmann, S.; Nahrig, J.; Rosenberg, R.; Hofler, H.; Ruger, R.; Werner, M. Identification of gene signatures for invasive colorectal tumor cells. Cancer Detect. Prev 2007, 31, 282–295. [Google Scholar]
- Ikeda, Y.; Tanji, E.; Makino, N.; Kawata, S.; Furukawa, T. MicroRNAs associated with mitogen-activated protein kinase in human pancreatic cancer. Mol. Cancer Res 2011, 10, 259–269. [Google Scholar]
- Xi, Y.; Nakajima, G.; Schmitz, J.C.; Chu, E.; Ju, J. Multi-Level gene expression profiles affected by thymidylate synthase and 5-fluorouracil in colon cancer. BMC Genomics 2006, 7, 68. [Google Scholar]
- Wright, H.J.; Matthews, J.B.; Chapple, I.L.; Ling-Mountford, N.; Cooper, P.R. Periodontitis associates with a type 1 IFN signature in peripheral blood neutrophils. J. Immunol 2008, 181, 5775–5784. [Google Scholar]
- Kadl, A.; Meher, A.K.; Sharma, P.R.; Lee, M.Y.; Doran, A.C.; Johnstone, S.R.; Elliott, M.R.; Gruber, F.; Han, J.; Chen, W.; et al. Identification of a novel macrophage phenotype that develops in response to atherogenic phospholipids via Nrf2. Circ. Res 2010, 107, 737–746. [Google Scholar]
- Selman, M.; Pardo, A.; Barrera, L.; Estrada, A.; Watson, S.R.; Wilson, K.; Aziz, N.; Kaminski, N.; Zlotnik, A. Gene expression profiles distinguish idiopathic pulmonary fibrosis from hypersensitivity pneumonitis. Am. J. Respir. Crit. Care Med 2006, 173, 188–198. [Google Scholar]
- Yung, C.K.; Halperin, V.L.; Tomaselli, G.F.; Winslow, R.L. Gene expression profiles in end-stage human idiopathic dilated cardiomyopathy: altered expression of apoptotic and cytoskeletal genes. Genomics 2004, 83, 281–297. [Google Scholar]
- Jerez-Timaure, N.C.; Kearney, F.; Simpson, E.B.; Eisen, E.J.; Pomp, D. Characterization of QTL with major effects on fatness and growth on mouse chromosome 2. Obes. Res 2004, 12, 1408–1420. [Google Scholar]
- De Wilde, J.; Mohren, R.; van den Berg, S.; Boekschoten, M.; Dijk, K.W.; de Groot, P.; Muller, M.; Mariman, E.; Smit, E. Short-Term high fat-feeding results in morphological and metabolic adaptations in the skeletal muscle of C57BL/6J mice. Physiol. Genomics 2008, 32, 360–369. [Google Scholar]
- MacLaren, R.; Cui, W.; Simard, S.; Cianflone, K. Influence of obesity and insulin sensitivity on insulin signaling genes in human omental and subcutaneous adipose tissue. J. Lipid Res 2008, 49, 308–323. [Google Scholar]
- Sokolovic, M.; Sokolovic, A.; van Roomen, C.P.; Gruber, A.; Ottenhoff, R.; Scheij, S.; Hakvoort, T.B.; Lamers, W.H.; Groen, A.K. Unexpected effects of fasting on murine lipid homeostasis—Transcriptomic and lipid profiling. J. Hepatol 2010, 52, 737–744. [Google Scholar]
- Rome, S.; Clement, K.; Rabasa-Lhoret, R.; Loizon, E.; Poitou, C.; Barsh, G.S.; Riou, J.P.; Laville, M.; Vidal, H. Microarray profiling of human skeletal muscle reveals that insulin regulates approximately 800 genes during a hyperinsulinemic clamp. J. Biol. Chem 2003, 278, 18063–18068. [Google Scholar]
- Kostek, M.C.; Chen, Y.W.; Cuthbertson, D.J.; Shi, R.; Fedele, M.J.; Esser, K.A.; Rennie, M.J. Gene expression responses over 24 h to lengthening and shortening contractions in human muscle: Major changes in CSRP3, MUSTN1, SIX1, and FBXO32. Physiol. Genomics 2007, 31, 42–52. [Google Scholar]
- Park, C.C.; Gale, G.D.; de Jong, S.; Ghazalpour, A.; Bennett, B.J.; Farber, C.R.; Langfelder, P.; Lin, A.; Khan, A.H.; Eskin, E.; et al. Gene networks associated with conditional fear in mice identified using a systems genetics approach. BMC Syst. Biol 2011, 5, 43. [Google Scholar]
- Wada, K.; Howard, J.T.; McConnell, P.; Whitney, O.; Lints, T.; Rivas, M.V.; Horita, H.; Patterson, M.A.; White, S.A.; Scharff, C.; et al. A molecular neuroethological approach for identifying and characterizing a cascade of behaviorally regulated genes. Proc. Natl. Acad. Sci. USA 2006, 103, 15212–15217. [Google Scholar]
- Lee, H.S.; Park, M.H.; Yang, S.J.; Park, K.C.; Kim, N.S.; Kim, Y.S.; Kim, D.I.; Yoo, H.S.; Choi, E.J.; Yeom, Y.I. Novel candidate targets of Wnt/beta-catenin signaling in hepatoma cells. Life Sci 2007, 80, 690–698. [Google Scholar]
- Fuerer, C.; Nusse, R.; Ten Berge, D. Wnt signalling in development and disease. Max delbruck center for molecular medicine meeting on wnt signaling in development and disease. EMBO Rep 2008, 9, 134–138. [Google Scholar]
- Sonderegger, S.; Husslein, H.; Leisser, C.; Knofler, M. Complex expression pattern of Wnt ligands and frizzled receptors in human placenta and its trophoblast subtypes. Placenta 2007, 28, S97–S102. [Google Scholar]
- Pollheimer, J.; Loregger, T.; Sonderegger, S.; Saleh, L.; Bauer, S.; Bilban, M.; Czerwenka, K.; Husslein, P.; Knofler, M. Activation of the canonical wingless/T-cell factor signaling pathway promotes invasive differentiation of human trophoblast. Am. J. Pathol 2006, 168, 1134–1147. [Google Scholar]
- Getsios, S.; Chen, G.T.; MacCalman, C.D. Regulation of beta-catenin mRNA and protein levels in human villous cytotrophoblasts undergoing aggregation and fusion in vitro: Correlation with E-cadherin expression. J. Reprod. Fertil 2000, 119, 59–68. [Google Scholar]
- Wong, N.C.; Novakovic, B.; Weinrich, B.; Dewi, C.; Andronikos, R.; Sibson, M.; Macrae, F.; Morley, R.; Pertile, M.D.; Craig, J.M.; et al. Methylation of the adenomatous polyposis coli (APC) gene in human placenta and hypermethylation in choriocarcinoma cells. Cancer Lett 2008, 268, 56–62. [Google Scholar]
- Hewitt, D.P.; Mark, P.J.; Dharmarajan, A.M.; Waddell, B.J. Placental expression of secreted frizzled related protein-4 in the rat and the impact of glucocorticoid-induced fetal and placental growth restriction. Biol. Reprod 2006, 75, 75–81. [Google Scholar]
- Sonderegger, S.; Haslinger, P.; Sabri, A.; Leisser, C.; Otten, J.V.; Fiala, C.; Knofler, M. Wingless (Wnt)-3A induces trophoblast migration and matrix metalloproteinase-2 secretion through canonical Wnt signaling and protein kinase B/AKT activation. Endocrinology 2010, 151, 211–220. [Google Scholar]
- Sonderegger, S.; Pollheimer, J.; Knofler, M. Wnt signalling in implantation, decidualisation and placental differentiation—Review. Placenta 2010, 31, 839–847. [Google Scholar]
- Fitzgerald, J.S.; Germeyer, A.; Huppertz, B.; Jeschke, U.; Knofler, M.; Moser, G.; Scholz, C.; Sonderegger, S.; Toth, B.; Markert, U.R. Governing the invasive trophoblast: Current aspects on intra- and extracellular regulation. Am. J. Reprod. Immunol 2010, 63, 492–505. [Google Scholar]
- Rena, V.; Angeletti, S.; Panzetta-Dutari, G.; Genti-Raimondi, S. Activation of beta-catenin signalling increases StarD7 gene expression in JEG-3 cells. Placenta 2009, 30, 876–883. [Google Scholar]
- Morohashi, K.; Hatano, O.; Nomura, M.; Takayama, K.; Hara, M.; Yoshii, H.; Takakusu, A.; Omura, T. Function and distribution of a steroidogenic cell-specific transcription factor, Ad4BP. J. Steroid Biochem. Mol. Biol 1995, 53, 81–88. [Google Scholar]
- Parker, K.L.; Rice, D.A.; Lala, D.S.; Ikeda, Y.; Luo, X.; Wong, M.; Bakke, M.; Zhao, L.; Frigeri, C.; Hanley, N.A.; et al. Steroidogenic factor 1: An essential mediator of endocrine development. Recent Prog. Horm. Res 2002, 57, 19–36. [Google Scholar]
- Lavoie, H.A.; King, S.R. Transcriptional regulation of steroidogenic genes: STARD1, CYP11A1 and HSD3B. Exp. Biol. Med. (Maywood) 2009, 234, 880–907. [Google Scholar]
- Mulholland, D.J.; Dedhar, S.; Coetzee, G.A.; Nelson, C.C. Interaction of nuclear receptors with the Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know? Endocr. Rev 2005, 26, 898–915. [Google Scholar]
- Rena, V.; Flores-Martín, J.; Angeletti, S.; Panzetta-Dutari, G.; Genti-Raimondi, S. StarD7 gene expression in trophoblast cells: Contribution of SF-1 and Wnt-b-catenin signalling. Mol. Endocrinol 2011, 8, 1364–1375. [Google Scholar]
- Mizusaki, H.; Kawabe, K.; Mukai, T.; Ariyoshi, E.; Kasahara, M.; Yoshioka, H.; Swain, A.; Morohashi, K. Dax-1 (dosage-sensitive sex reversal-adrenal hypoplasia congenita critical region on the X chromosome, gene 1) gene transcription is regulated by wnt4 in the female developing gonad. Mol. Endocrinol 2003, 17, 507–519. [Google Scholar]
- Schinner, S.; Willenberg, H.S.; Krause, D.; Schott, M.; Lamounier-Zepter, V.; Krug, A.W.; Ehrhart-Bornstein, M.; Bornstein, S.R.; Scherbaum, W.A. Adipocyte-Derived products induce the transcription of the StAR promoter and stimulate aldosterone and cortisol secretion from adrenocortical cells through the Wnt-signaling pathway. Int. J. Obes. (Lond. ) 2007, 31, 864–870. [Google Scholar]
- Parakh, T.N.; Hernandez, J.A.; Grammer, J.C.; Weck, J.; Hunzicker-Dunn, M.; Zeleznik, A.J.; Nilson, J.H. Follicle-Stimulating hormone/cAMP regulation of aromatase gene expression requires beta-catenin. Proc. Natl. Acad. Sci. USA 2006, 103, 12435–12440. [Google Scholar]
- Gardner, S.; Stavrou, E.; Rischitor, P.E.; Faccenda, E.; Pawson, A.J. Targeting mediators of Wnt signalling pathways by GnRH in gonadotropes. J. Mol. Endocrinol 2010, 44, 195–201. [Google Scholar]
- Hossain, A.; Saunders, G.F. Synergistic cooperation between the beta-catenin signaling pathway and steroidogenic factor 1 in the activation of the Mullerian inhibiting substance type II receptor. J. Biol. Chem 2003, 278, 26511–26516. [Google Scholar]
- Salisbury, T.B.; Binder, A.K.; Nilson, J.H. Welcoming beta-catenin to the gonadotropin-releasing hormone transcriptional network in gonadotropes. Mol. Endocrinol 2008, 22, 1295–1303. [Google Scholar]
- Gummow, B.M.; Winnay, J.N.; Hammer, G.D. Convergence of Wnt signaling and steroidogenic factor-1 (SF-1) on transcription of the rat inhibin alpha gene. J. Biol. Chem 2003, 278, 26572–26579. [Google Scholar]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar]
- Shishova, E.Y.; Stoll, J.M.; Ersoy, B.A.; Shrestha, S.; Scapa, E.F.; Li, Y.; Niepel, M.W.; Su, Y.; Jelicks, L.A.; Stahl, G.L.; et al. Genetic ablation or chemical inhibition of phosphatidylcholine transfer protein attenuates diet-induced hepatic glucose production. Hepatology 2011, 54, 664–674. [Google Scholar]
- Ullmannova, V.; Popescu, N.C. Expression profile of the tumor suppressor genes DLC-1 and DLC-2 in solid tumors. Int. J. Oncol 2006, 29, 1127–1132. [Google Scholar]
- Durkin, M.E.; Ullmannova, V.; Guan, M.; Popescu, N.C. Deleted in liver cancer 3 (DLC-3), a novel Rho GTPase-activating protein, is downregulated in cancer and inhibits tumor cell growth. Oncogene 2007, 26, 4580–4589. [Google Scholar]
- Olayioye, M.A.; Hoffmann, P.; Pomorski, T.; Armes, J.; Simpson, R.J.; Kemp, B.E.; Lindeman, G.J.; Visvader, J.E. The phosphoprotein StarD10 is overexpressed in breast cancer and cooperates with ErbB receptors in cellular transformation. Cancer Res 2004, 64, 3538–3544. [Google Scholar]
- Al-Khan, A.; Aye, I.L.; Barsoum, I.; Borbely, A.; Cebral, E.; Cerchi, G.; Clifton, V.L.; Collins, S.; Cotechini, T.; Davey, A.; et al. IFPA Meeting 2010 Workshops Report II: Placental pathology; trophoblast invasion; fetal sex; parasites and the placenta; decidua and embryonic or fetal loss; trophoblast differentiation and syncytialisation. Placenta 2011, 32, S90–S99. [Google Scholar]
- Flores-Martin, J.; Rena, V.; Marquez, S.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. StarD7 knockdown modulates ABCG2 expression, cell migration, proliferation, and differentiation of human choriocarcinoma JEG-3 cells. PLoS One 2012, 7, e44152. [Google Scholar]
- Robey, R.W.; To, K.K.; Polgar, O.; Dohse, M.; Fetsch, P.; Dean, M.; Bates, S.E. ABCG2: A perspective. Adv. Drug Deliv. Rev 2009, 61, 3–13. [Google Scholar]
- Ni, Z.; Mao, Q. ATP-Binding cassette efflux transporters in human placenta. Curr. Pharm. Biotechnol. 2011, 12, 674–685. [Google Scholar]
- Vahakangas, K.; Myllynen, P. Drug transporters in the human blood-placental barrier. Br. J. Pharmacol 2009, 158, 665–678. [Google Scholar]
- Quazi, F.; Molday, R.S. Lipid transport by mammalian ABC proteins. Essays Biochem 2011, 50, 265–290. [Google Scholar]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. The xenobiotic transporter ABCG2 plays a novel role in differentiation of trophoblast-like BeWo cells. Placenta 2007, 28, S116–S120. [Google Scholar]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. ABC drug transporter expression and functional activity in trophoblast-like cell lines and differentiating primary trophoblast. Am. J. Physiol. Regul. Integr. Comp. Physiol 2006, 290, R1357–R1365. [Google Scholar]
- Evseenko, D.A.; Murthi, P.; Paxton, J.W.; Reid, G.; Emerald, B.S.; Mohankumar, K.M.; Lobie, P.E.; Brennecke, S.P.; Kalionis, B.; Keelan, J.A. The ABC transporter BCRP/ABCG2 is a placental survival factor, and its expression is reduced in idiopathic human fetal growth restriction. FASEB J 2007, 21, 3592–3605. [Google Scholar]
- Flores-Martín, J.; Rena, V.; Márquez, S.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. StarD7 siRNA modulates ABCG2 expression, cell migration and proliferation in epithelial cell lines. Biocell 2012, 66, 36. [Google Scholar]
- Vaitukaitis, J.L.; Ebersole, E.R. Evidence for altered synthesis of human chorionic gonadotropin in gestational trophoblastic tumors. J. Clin. Endocrinol. Metab 1976, 42, 1048–1055. [Google Scholar]
- Kharfi, A.; Giguere, Y.; de Grandpre, P.; Moutquin, J.M.; Forest, J.C. Human chorionic gonadotropin (hCG) may be a marker of systemic oxidative stress in normotensive and preeclamptic term pregnancies. Clin. Biochem 2005, 38, 717–721. [Google Scholar]
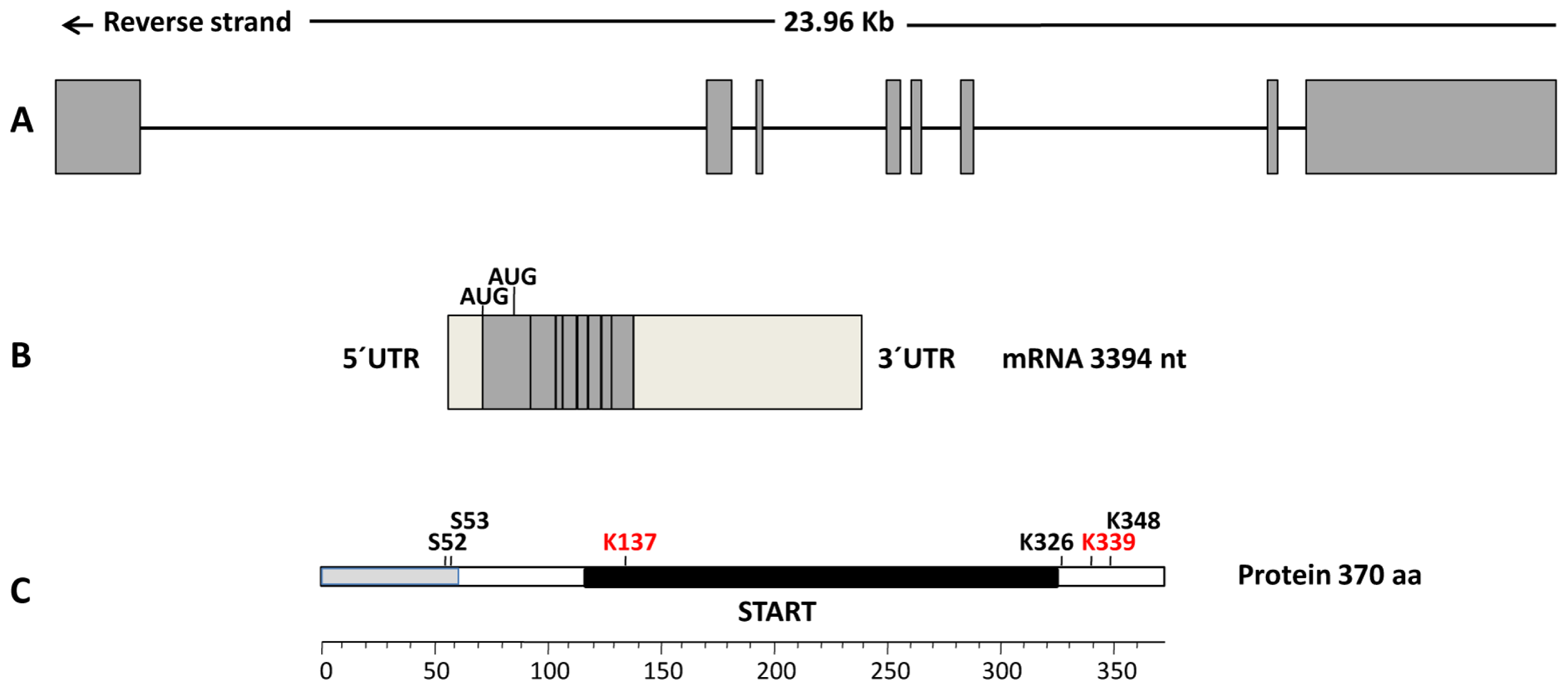



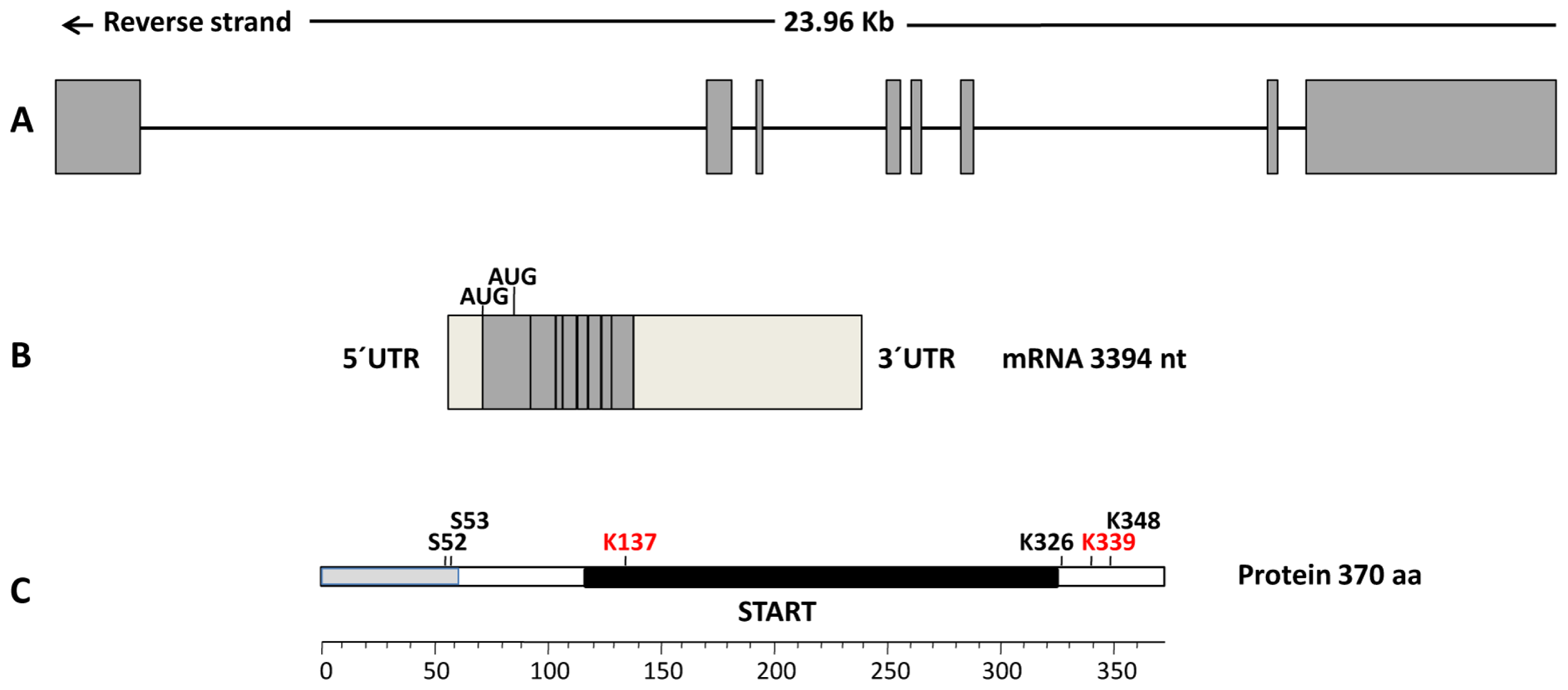
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological/Pathological Major Situations | Study | Cells/Tissue | Fold Change | Expression | Reference |
|---|---|---|---|---|---|
| Cancer | Ductal carcinoma in situ vs. normal breast tissue | Myoepithelial cells | 3 | Upregulated | [37] |
| B-chronic lymphocytic leukemia vs. normal B cells | Peripheral blood mononuclear cells | >97.7 | Upregulated | [38] | |
| Microdissected tumor cells | Colorectal tissue | 2.59 | Upregulated | [39] | |
| MIA PaCa-2 cells transfected with miR-193b | Pancreatic cell lines | 2 | Downregulated | [40] | |
| HCT-C18 cells treated with 5-fluorouracil | Human colon cancer cell lines | 2 | Upregulated | [41] | |
| Inflammation | Chronic periodontitis vs. normal | Peripheral blood neutrophils | 2.19 | Upregulated | [42] |
| Conventional M1 macrophages treated with oxidized phospholipid | Macrophages | 3.02 | Upregulated | [43] | |
| Hypersensitivity pneumonitis vs. idiopathic pulmonary fibrosis | Lung | ND ** | Upregulated | [44] | |
| Human idiopathic dilated cardiomyopathy vs. nonfailing | Heart | 0.62 | Downregulated | [45] | |
| Metabolic state | Quantitative trait loci on chromosome 2 associated with growth and fatness | Liver | 1.17 | Upregulated | [46] |
| Short-term high-fat-diet-fed mice | Skeletal muscle | 1.3 | Upregulated | [47] | |
| Insulin resistance vs. insulin sensitive | Omental adipose tissue | 0.28 | Downregulated | [48] | |
| Fasted mice | Small intestine | 1.4 | Upregulated | [49] | |
| Hyperinsulinemic clamp | Skeletal muscle | 3.31 | Upregulated | [50] | |
| Lengthening vs. shortening contraction | Leg muscle biopsies | 2 | Downregulated | [51] | |
| Behavior | Conditional fear | Brain | ND ** | Upregulated | [52] |
| Singing vs. nonsinging songbird behavior | Forebrain vocal nuclei of brain | DIH ** | Upregulated | [53] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Flores-Martin, J.; Rena, V.; Angeletti, S.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. The Lipid Transfer Protein StarD7: Structure, Function, and Regulation. Int. J. Mol. Sci. 2013, 14, 6170-6186. https://doi.org/10.3390/ijms14036170
Flores-Martin J, Rena V, Angeletti S, Panzetta-Dutari GM, Genti-Raimondi S. The Lipid Transfer Protein StarD7: Structure, Function, and Regulation. International Journal of Molecular Sciences. 2013; 14(3):6170-6186. https://doi.org/10.3390/ijms14036170
Chicago/Turabian StyleFlores-Martin, Jésica, Viviana Rena, Sofía Angeletti, Graciela M. Panzetta-Dutari, and Susana Genti-Raimondi. 2013. "The Lipid Transfer Protein StarD7: Structure, Function, and Regulation" International Journal of Molecular Sciences 14, no. 3: 6170-6186. https://doi.org/10.3390/ijms14036170