The Interaction of Adrenomedullin and Macrophages Induces Ovarian Cancer Cell Migration via Activation of RhoA Signaling Pathway

Abstract
:1. Introduction
2. Results and Discussion
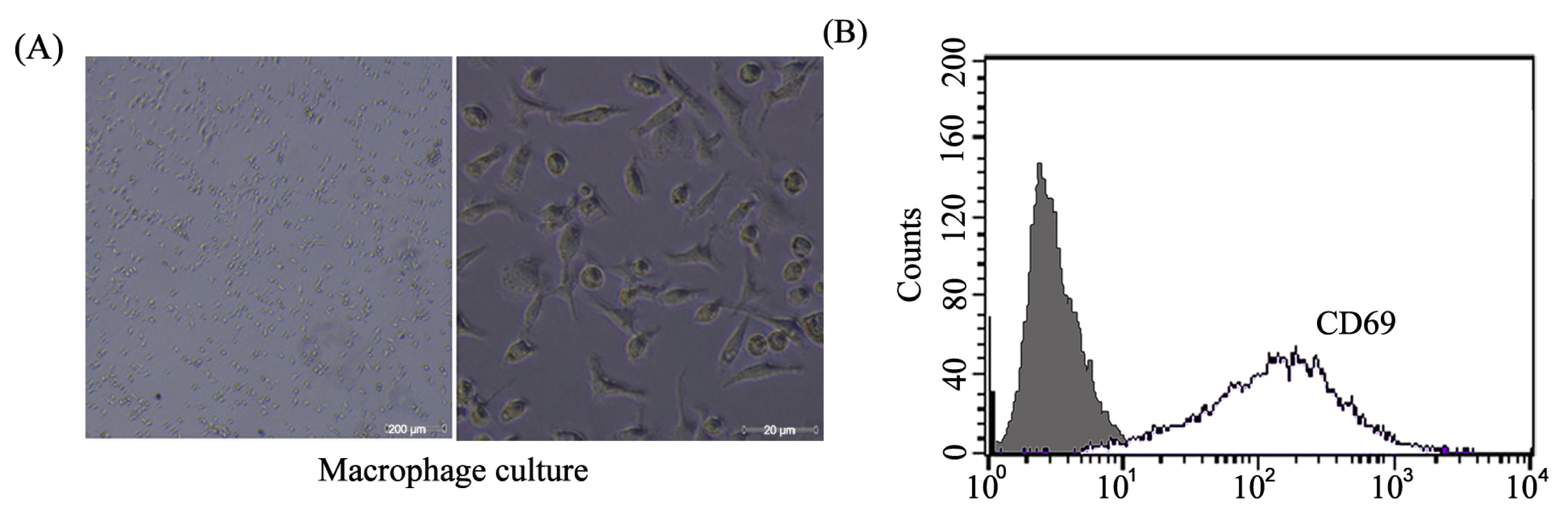
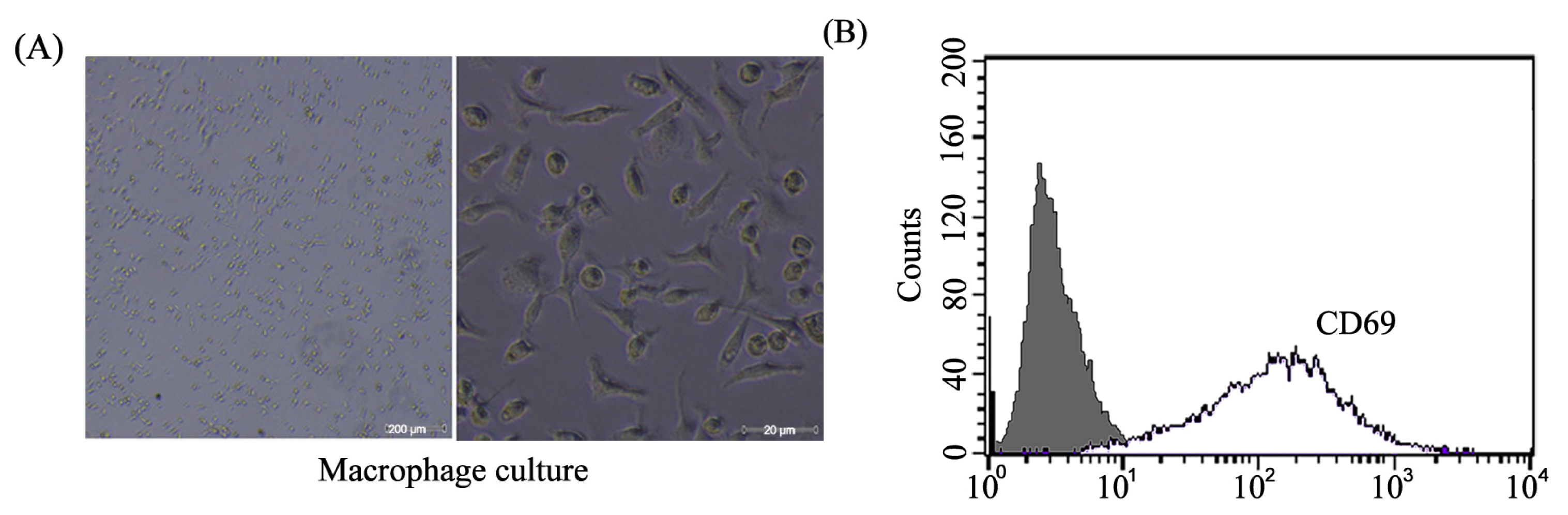
2.1. Mice Peritoneal Macrophages Isolated and Determined
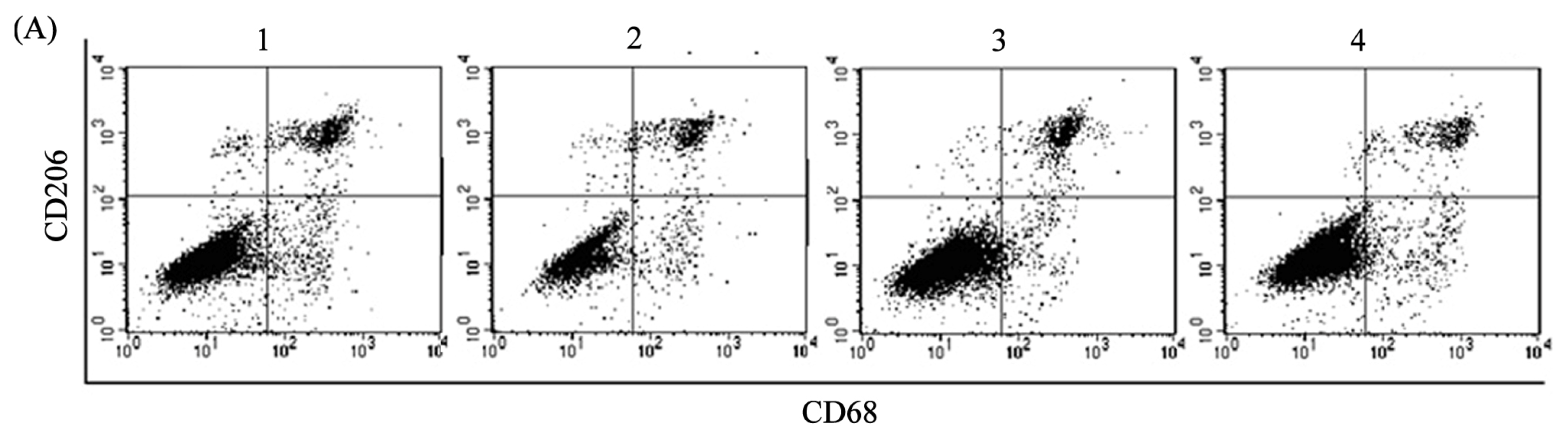
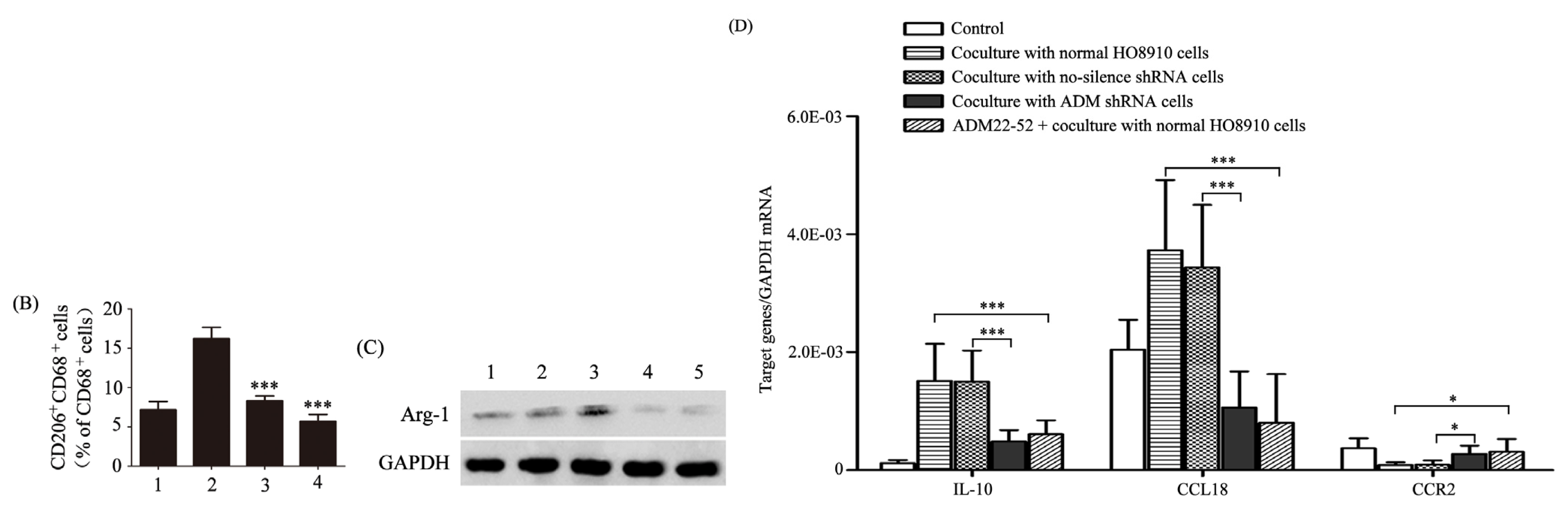
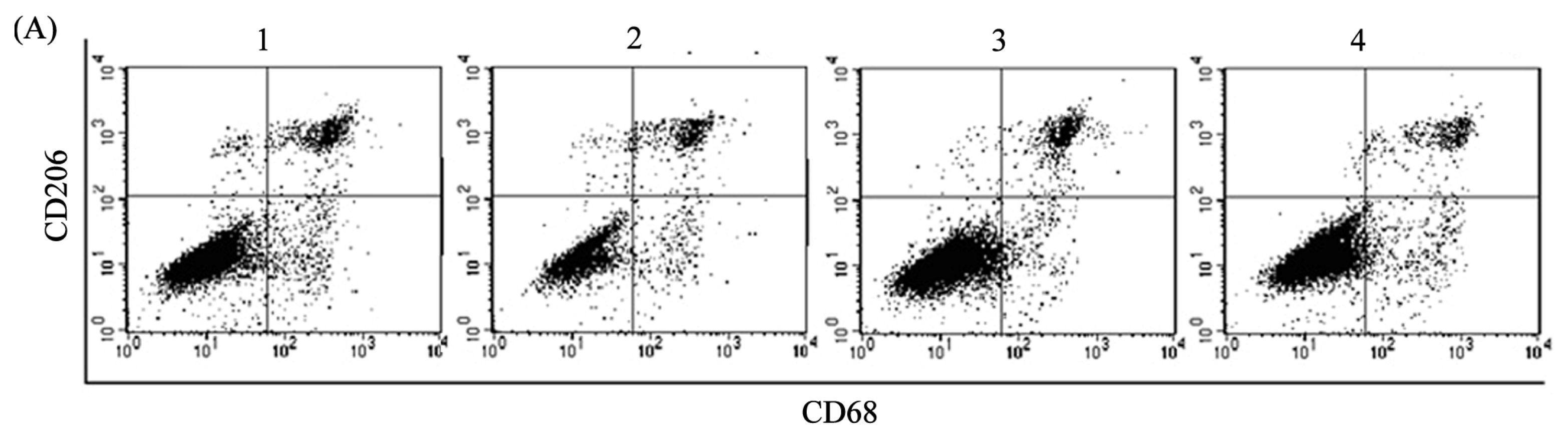
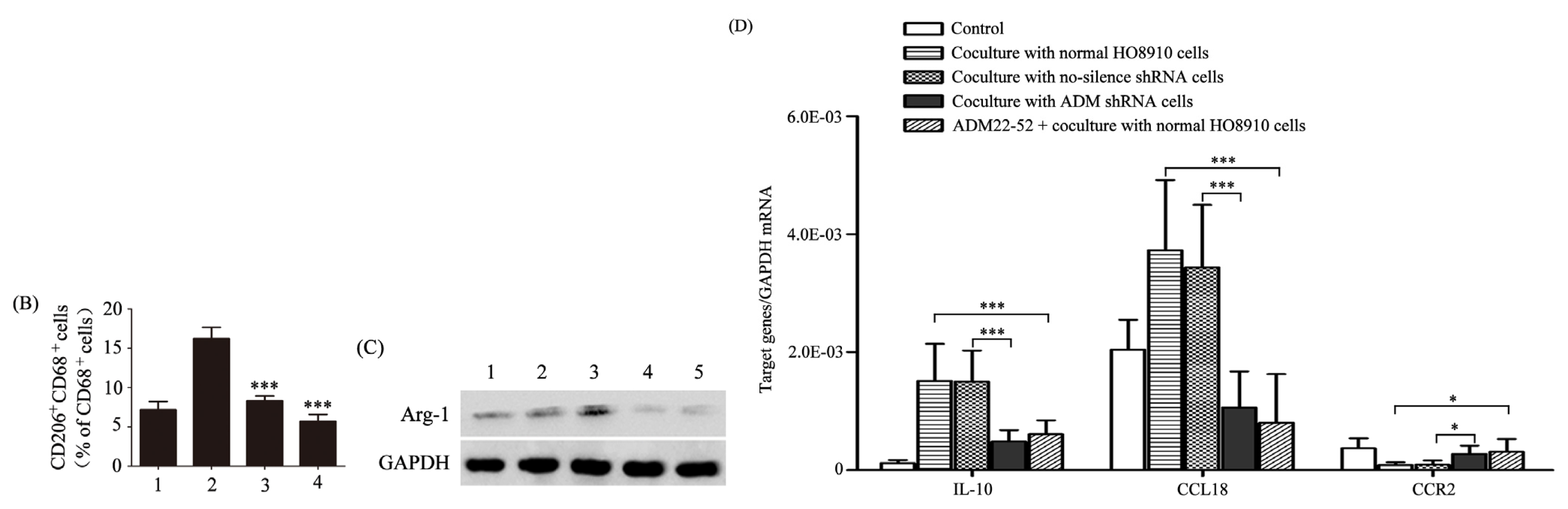
2.2. ADM Induced Macrophage Phenotype and Cytokine Production Similar to TAMs
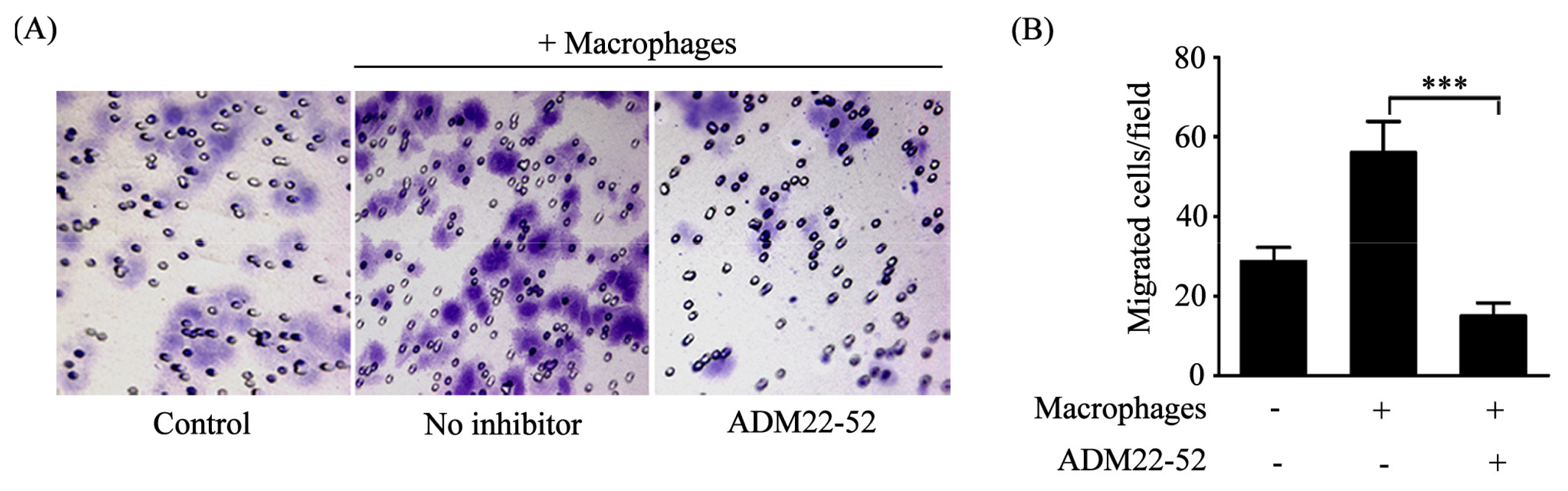
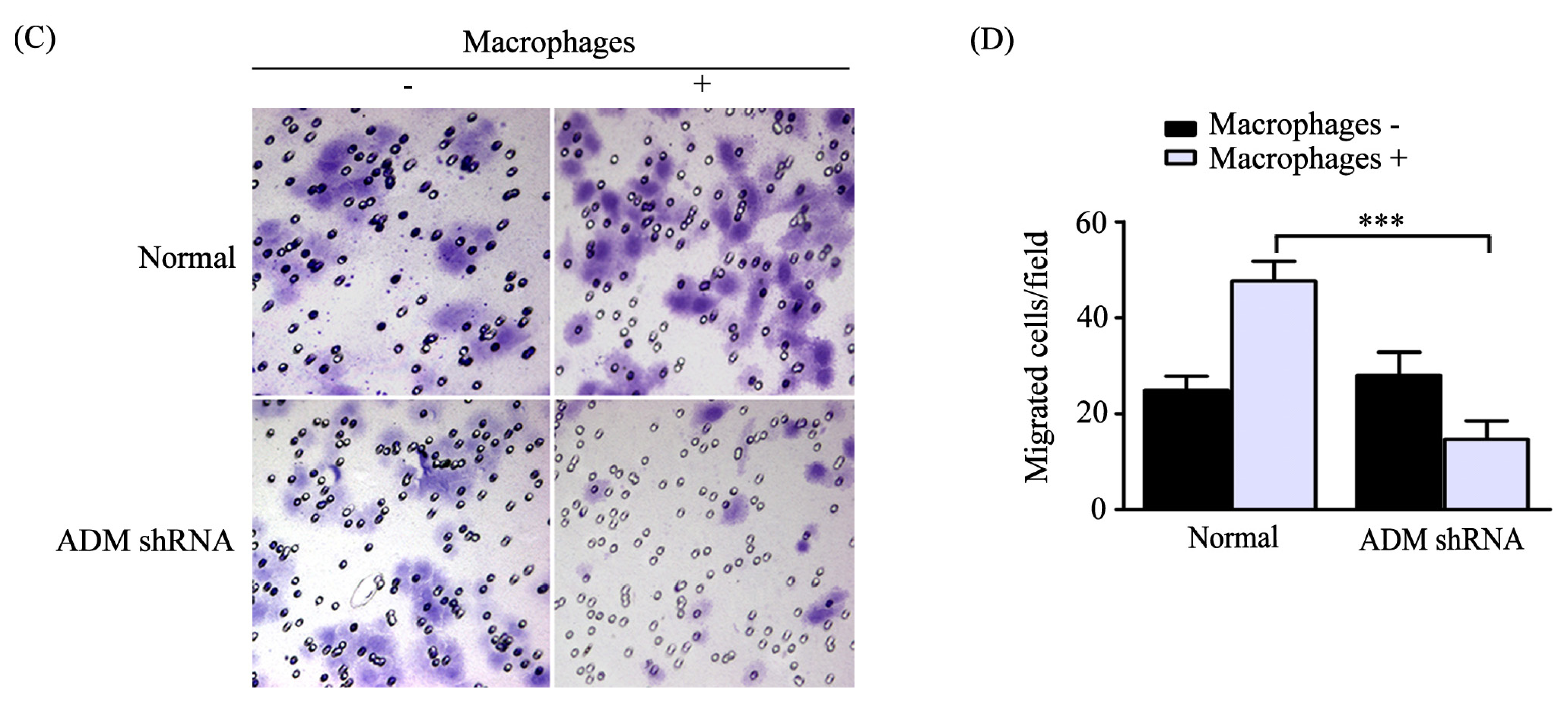
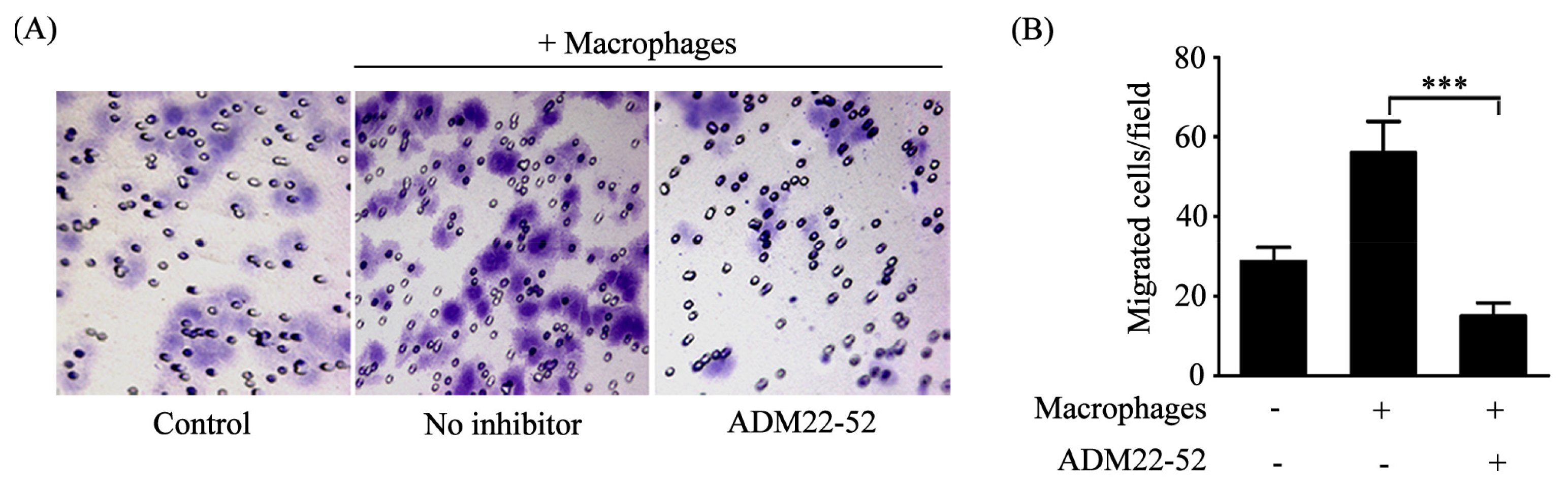
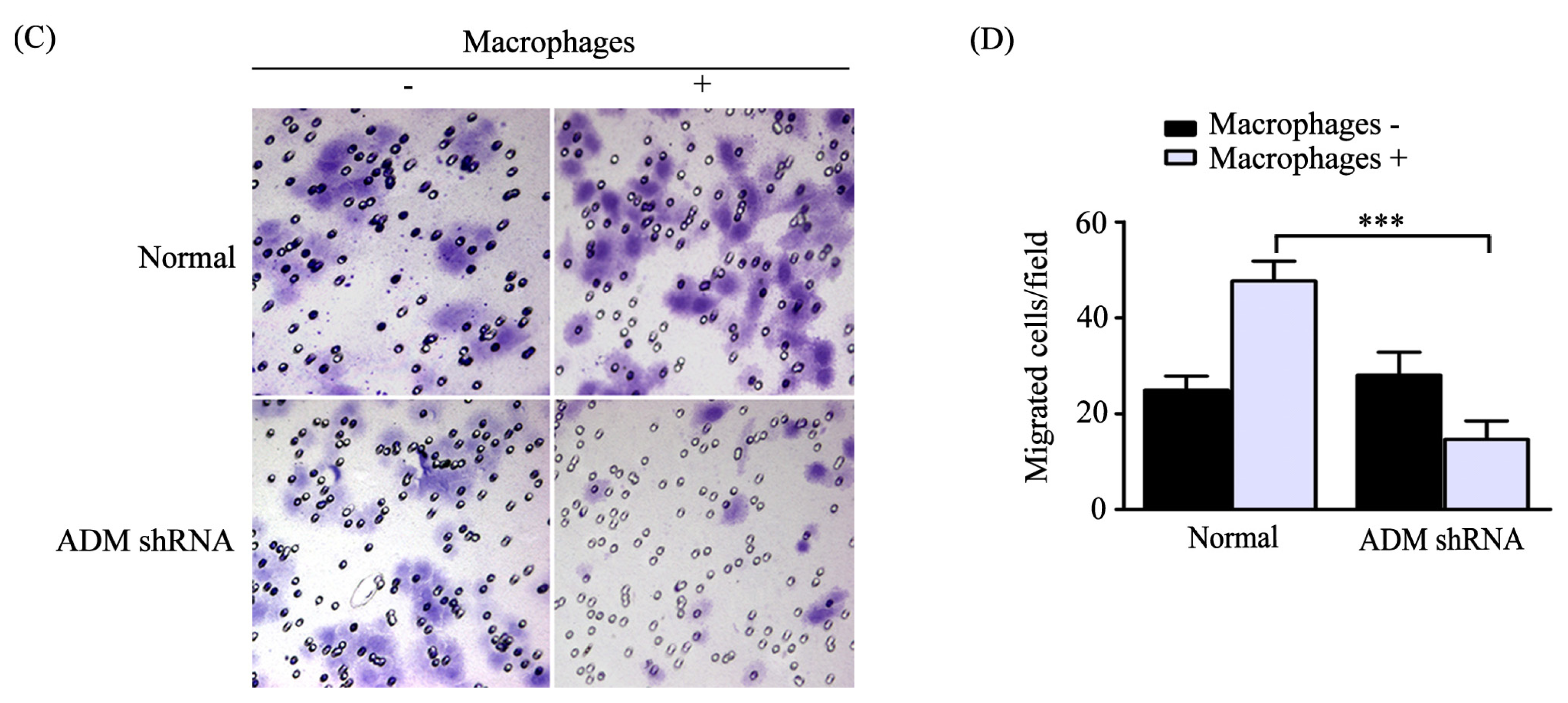
2.3. Macrophages Polarized by ADM Promoted the HO8910 Cell Migration
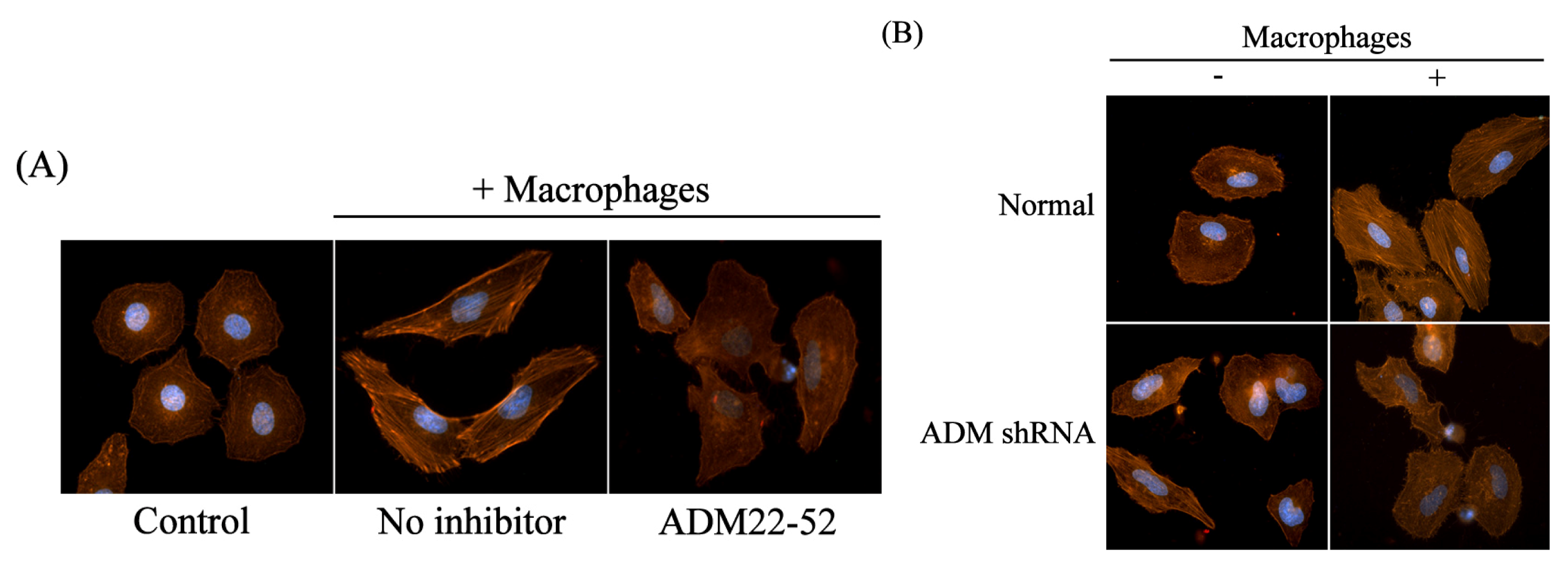
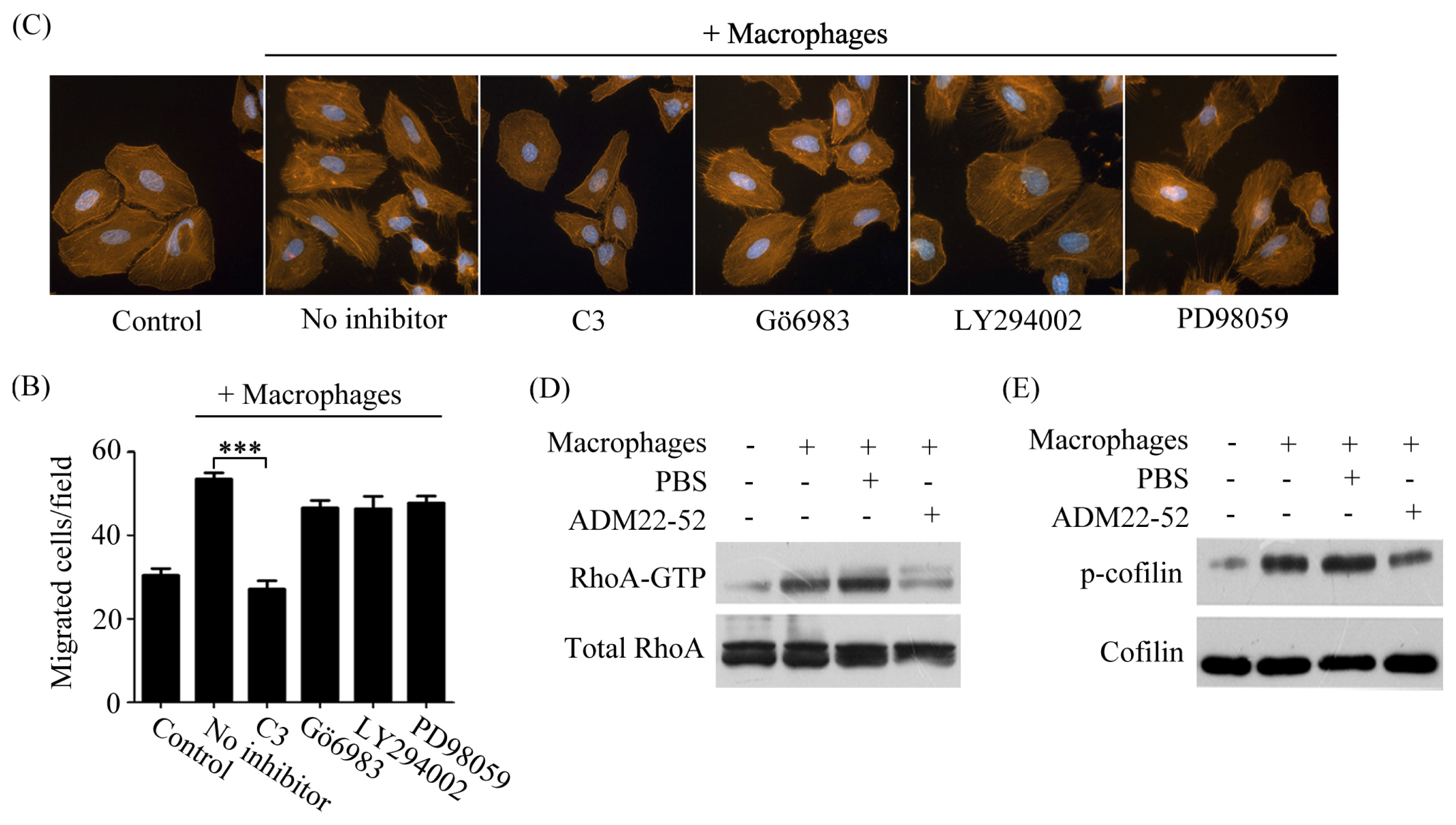
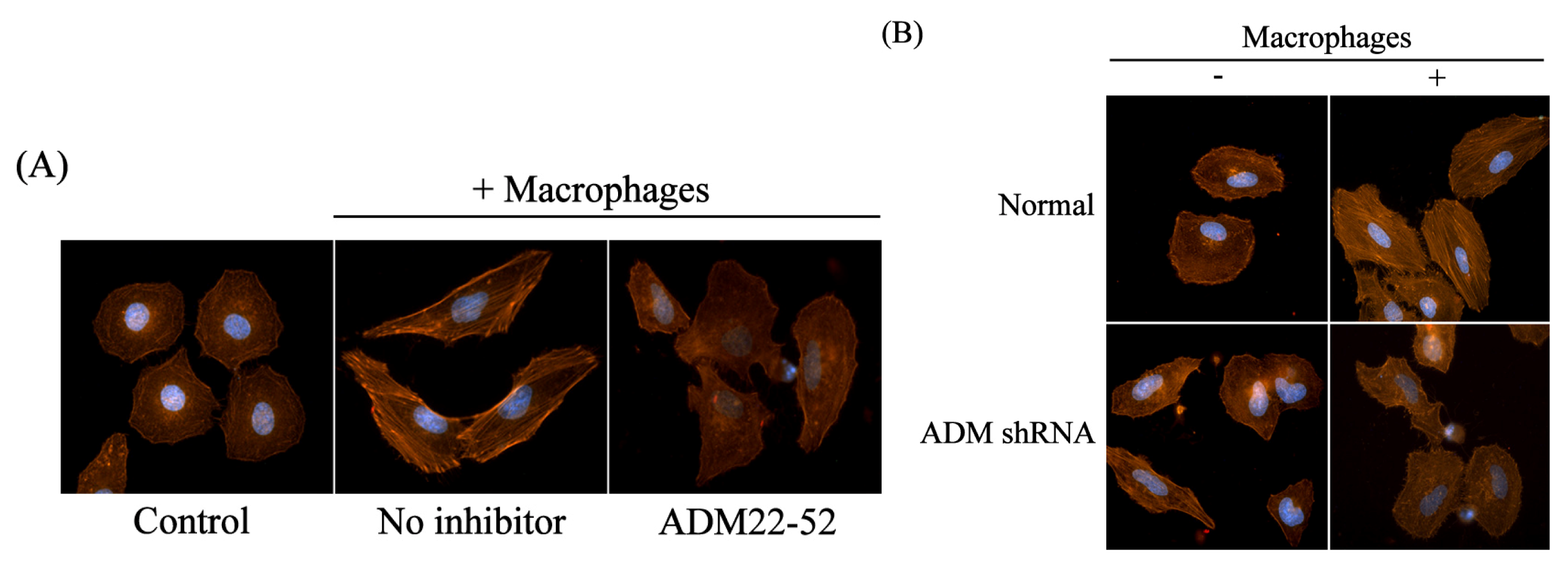
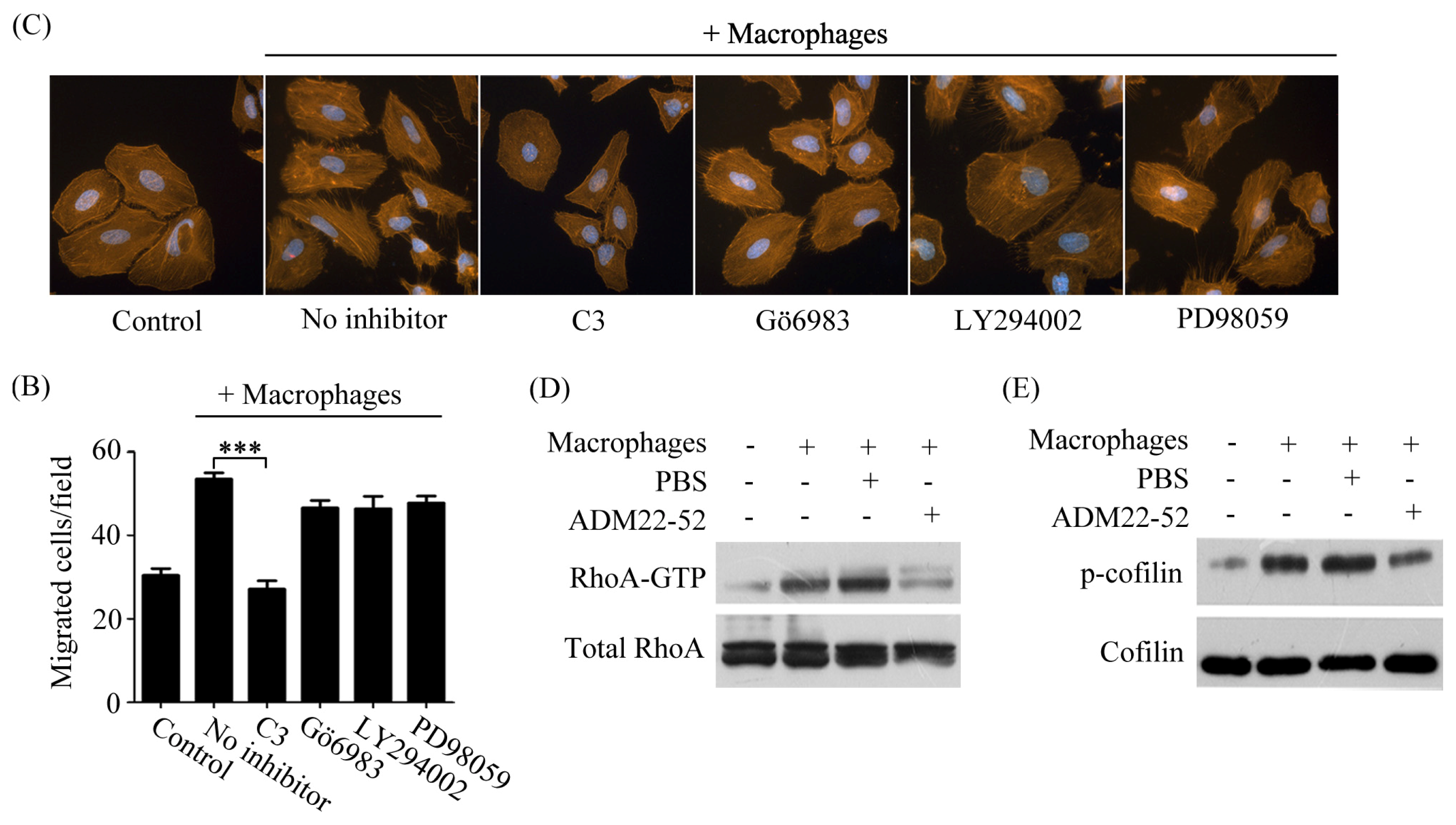
2.4. Macrophages Polarized by ADM Induced HO8910 Cell Cytoskeleton Rearrangement
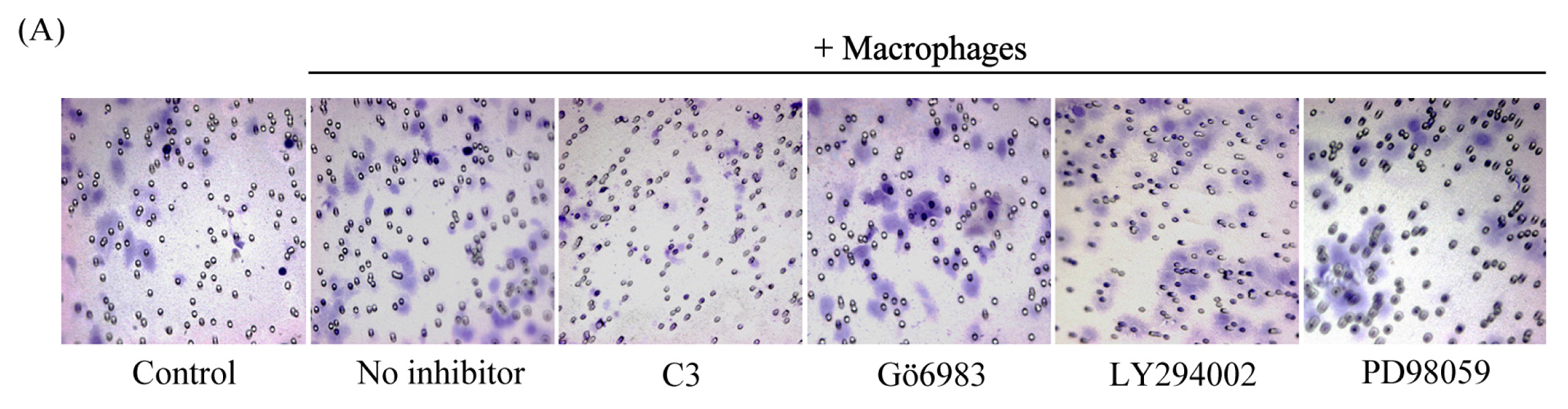
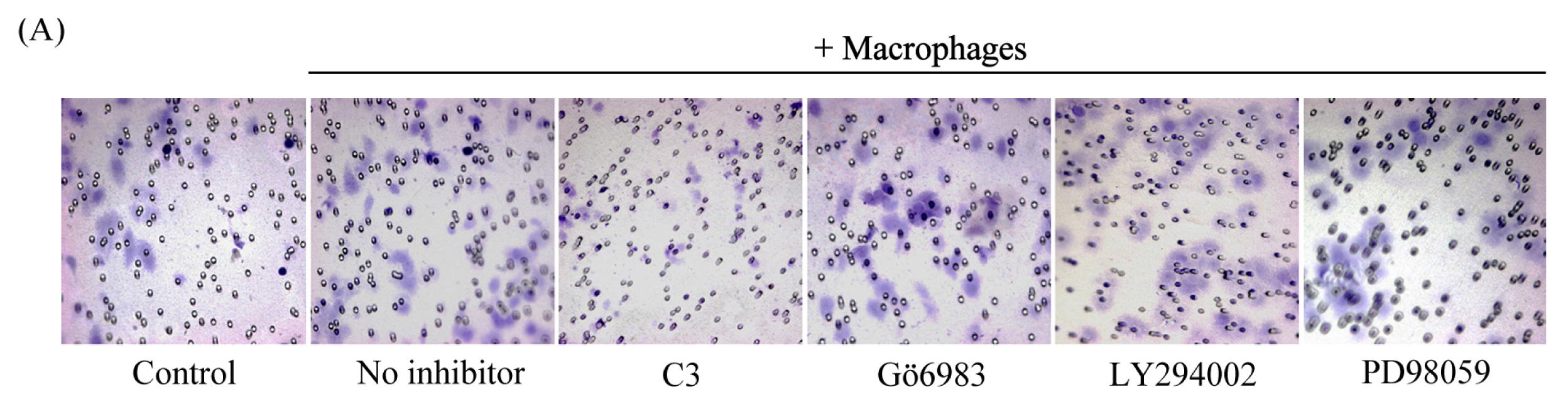
2.5. Macrophage-Induced Migration of HO8910 Cells via Activation of RhoA Signaling Pathway
2.6. Discussion
3. Materials and Methods
3.1. Cell Culture
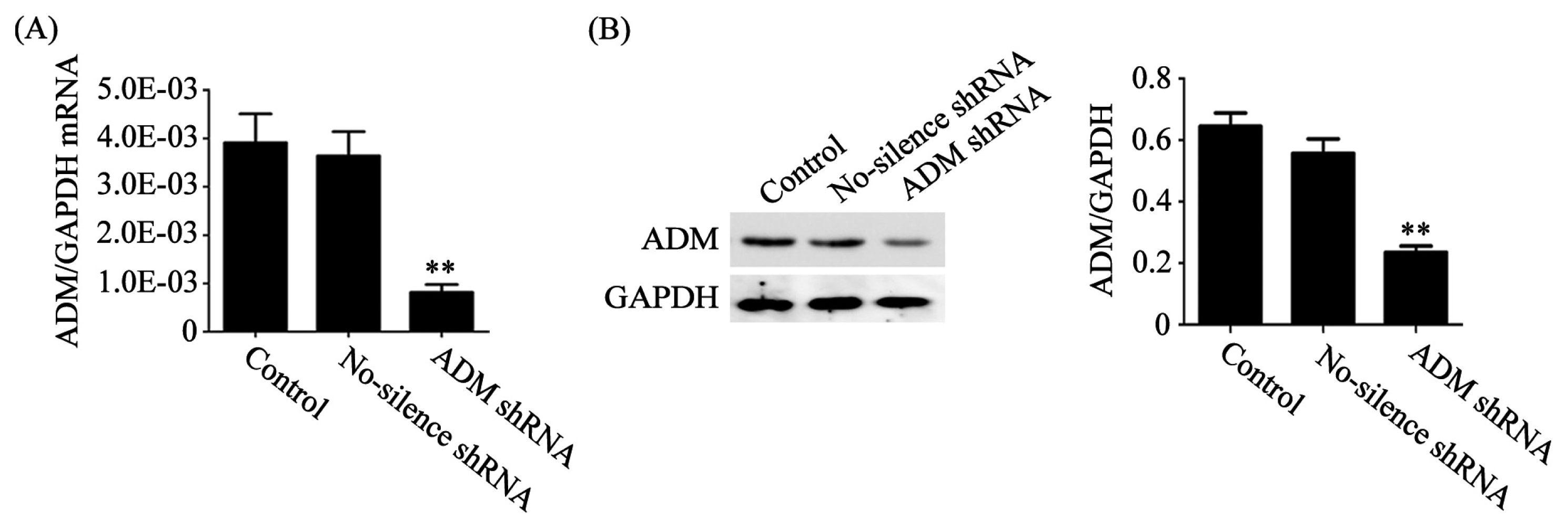
3.2. Knockdown of ADM Expression with Specific shRNA
3.3. Western Blot
3.4. Preparation of Peritoneal Macrophages and Culture
3.5. Macrophage and HO8910 Cell Coculture
3.6. Flow Cytometry
3.7. Real-Time PCR
3.8. Cell Migration Assay
3.9. Cell Cytoskeleton Staining
3.10. RhoA Activity Assay
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Chappell, N.P.; Teng, P.N.; Hood, B.L.; Wang, G.; Darcy, K.M.; Hamilton, C.A.; Maxwell, G.L.; Conrads, T.P. Mitochondrial proteomic analysis of cisplatin resistance in ovarian cancer. J. Proteome Res 2012, 11, 4605–4614. [Google Scholar]
- Stout, R.D.; Jiang, C.; Matta, B.; Tietzel, I.; Watkins, S.K.; Suttles, J. Macrophages sequentially change their functional phenotype in response to changes in microenvironmental influences. J. Immunol 2005, 175, 342–349. [Google Scholar]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 2002, 23, 549–555. [Google Scholar]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar]
- Rolny, C.; Mazzone, M.; Tugues, S.; Laoui, D.; Johansson, I.; Coulon, C.; Squadrito, M.L.; Segura, I.; Li, X.; Knevels, E.; et al. HRG inhibits tumor growth and metastasis by inducing macrophage polarization and vessel normalization through downregulation of PlGF. Cancer Cell 2011, 19, 31–44. [Google Scholar]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 2004, 25, 677–686. [Google Scholar]
- Kitamura, K.; Kangawa, K.; Kawamoto, M.; Ichiki, Y.; Nakamura, S.; Matsuo, H.; Eto, T. Adrenomedullin: A novel hypotensive peptide isolated from human pheochromocytoma. Biochem. Biophys. Res. Commun 1993, 192, 553–560. [Google Scholar]
- Miller, M.J.; Martínez, A.; Unsworth, E.J.; Thiele, C.J.; Moody, T.W.; Elsasser, T.; Cuttitta, F. Adrenomedullin expression in human tumor cell lines. Its potential role as an autocrine growth factor. J. Biol. Chem 1996, 271, 23345–23351. [Google Scholar]
- Zudaire, E.; Martínez, A.; Cuttitta, F. Adrenomedullin and cancer. Regul. Pept 2003, 112, 175–183. [Google Scholar]
- Deng, B.; Zhang, S.; Miao, Y.; Han, Z.; Zhang, X.; Wen, F.; Zhang, Y. Adrenomedullin expression in epithelial ovarian cancers and promotes HO8910 cell migration associated with upregulating integrin α5β1 and phosphorylating FAK and paxillin. J. Exp. Clin. Cancer Res 2012, 31, 19. [Google Scholar]
- Chen, P.; Huang, Y.; Bong, R.; Ding, Y.; Song, N.; Wang, X.; Song, X.; Luo, Y. Tumor-associated macrophages promote angiogenesis and melanoma growth via adrenomedullin in a paracrine and autocrine manner. Clin. Cancer Res 2011, 17, 7230–7239. [Google Scholar]
- Zhang, L.J.; Tao, B.B.; Wang, M.J.; Jin, H.M.; Zhu, Y.C. PI3K p110α isoform-dependent Rho GTPase Rac1 activation mediates H(2)S-promoted endothelial cell migration via actin cytoskeleton reorganization. PLoS One 2012, 7, e44590. [Google Scholar]
- Horiuchi, A.; Kikuchi, N.; Osada, R.; Wang, C.; Hayashi, A.; Nikaido, T.; Konishi, I. Overexpression of RhoA enhances peritoneal dissemination: RhoA suppression with Lovastatin may be useful for ovarian cancer. Cancer Sci 2008, 99, 2532–2539. [Google Scholar]
- Chiu, H.Y.; Sun, K.H.; Chen, S.Y.; Wang, H.H.; Lee, M.Y.; Tsou, Y.C.; Jwo, S.C.; Sun, G.H.; Tang, S.J. Autocrine CCL2 promotes cell migration and invasion via PKC activation and tyrosine phosphorylation of paxillin in bladder cancer cells. Cytokine 2012, 59, 423–432. [Google Scholar]
- Kong, L.; Schäfer, G.; Bu, H.; Zhang, Y.; Zhang, Y.; Klocker, H. Lamin A/C protein is overexpressed in tissue-invading prostate cancer and promotes prostate cancer cell growth, migration and invasion through the PI3K/AKT/PTEN pathway. Carcinogenesis 2012, 33, 751–759. [Google Scholar]
- Zhang, L.; Teng, Y.; Zhang, Y.; Liu, J.; Xu, L.; Qu, J.; Hou, K.; Yang, X.; Liu, Y.; Qu, X. C-Src-mediated RANKL-induced breast cancer cell migration by activation of the ERK and Akt pathway. Oncol. Lett 2012, 3, 395–400. [Google Scholar]
- Ohashi, K.; Nagata, K.; Maekawa, M.; Ishizaki, T.; Narumiya, S.; Mizuno, K. Rho-associated kinase ROCK activates LIM-kinase 1 by phosphorylation at threonine 508 within the activation loop. J. Biol. Chem 2000, 275, 3577–3582. [Google Scholar]
- Yang, N.; Higuchi, O.; Ohashi, K.; Nagata, K.; Wada, A.; Kangawa, K.; Nishida, E.; Mizuno, K. Cofilin phosphorylation by LIM-kinase 1 and its role in Rac-mediated actin reorganization. Nature 1998, 393, 809–812. [Google Scholar]
- Li, B.; Zhao, W.D.; Tan, Z.M.; Fang, W.G.; Zhu, L.; Chen, Y.H. Involvement of Rho/ROCK signalling in small cell lung cancer migration through human brain microvascular endothelial cells. FEBS Lett 2006, 580, 4252–4260. [Google Scholar]
- Bingle, L.; Brown, N.J.; Lewis, C.E. The role of tumour-associated macrophages in tumour progression: Implications for new anticancer therapies. J. Pathol 2002, 196, 254–265. [Google Scholar]
- Hata, K.; Takebayashi, Y.; Akiba, S.; Fujiwaki, R.; Iida, K.; Nakayama, K.; Nakayama, S.; Fukumoto, M.; Miyazaki, K. Expression of the adrenomedullin gene in epithelial ovarian cancer. Mol. Hum. Reprod 2000, 6, 867–872. [Google Scholar]
- Baranello, C.; Mariani, M.; Andreoli, M.; Fanelli, M.; Martinelli, E.; Ferrandina, G.; Scambia, G.; Shahabi, S.; Ferlini, C. Adrenomedullin in ovarian cancer: Foe in vitro and friend in vivo? PLoS One 2012, 7, e40678. [Google Scholar]
- Schutyser, E.; Struyf, S.; Proost, P.; Opdenakker, G.; Laureys, G.; Verhasselt, B.; Peperstraete, L.; van de Putte, I.; Saccani, A.; Allavena, P.; et al. Identification of biologically active chemokine isoforms from ascitic fluid and elevated levels of CCL18/pulmonary and activation-regulated chemokine in ovarian carcinoma. J. Biol. Chem 2002, 277, 24584–24593. [Google Scholar]
- Sica, A.; Saccani, A.; Bottazzi, B.; Bernasconi, S.; Allavena, P.; Gaetano, B.; Fei, F.; LaRosa, G.; Scotton, C.; Balkwill, F.; et al. Defective expression of the monocyte chemotactic protein-1 receptor CCR2 in macrophages associated with human ovarian carcinoma. J. Immunol 2000, 164, 733–738. [Google Scholar]
- Chen, P.; Pang, X.; Zhang, Y.; He, Y. Effect of inhibition of the adrenomedullin gene on the growth and chemosensitivity of ovarian cancer cells. Oncol. Rep 2012, 27, 1461–1466. [Google Scholar]
- Hagemann, T.; Robinson, S.C.; Schulz, M.; Trümper, L.; Balkwill, F.R.; Binder, C. Enhanced invasiveness of breast cancer cell lines upon co-cultivation with macrophages is due to TNF-α dependent up-regulation of matrix metalloproteases. Carcinogenesis 2004, 25, 1543–1549. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| Homo-ADM | Forward: 5′-TCCCCCTATTTTAAGACGTGAATG-3′ Reverse: 5′-CATGCACACAAA CACACTCACAT-3′ |
| Homo-GAPDH | Forward: 5′-GAAGGTGAAGGTCGGAGT-3′ Reverse: 5′-GAAAGATGGTGATGGGATTTC-3′ |
| Mus-IL-10 | Forward: 5′-TGAGGCGCTGTCGTCATCGATTTCTCCC-3′ Reverse: 5′-GGTTGCCAAGCCTTATCGGA-3′ |
| Mus-CCL18 | Forward: 5′-CCCTCCTTGTCCTCGTCTG-3′ Reverse: 5′-GCTTCAGGTCGCTGATGTATT-3′ |
| Mus-CCR2 | Forward: 5′-TTTGTTTTTGCAGATGATTCAA-3′ Reverse: 5′-TGCCATCATAAAGGAGCCAT-3′ |
| Mus-GAPDH | Forward: 5′-TGCATCCTGCACCACCAACTGCTTAG-3′ Reverse: 5′-TTCACCACCATGGAGAAGGC-3′ |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pang, X.; Shang, H.; Deng, B.; Wen, F.; Zhang, Y. The Interaction of Adrenomedullin and Macrophages Induces Ovarian Cancer Cell Migration via Activation of RhoA Signaling Pathway. Int. J. Mol. Sci. 2013, 14, 2774-2787. https://doi.org/10.3390/ijms14022774
Pang X, Shang H, Deng B, Wen F, Zhang Y. The Interaction of Adrenomedullin and Macrophages Induces Ovarian Cancer Cell Migration via Activation of RhoA Signaling Pathway. International Journal of Molecular Sciences. 2013; 14(2):2774-2787. https://doi.org/10.3390/ijms14022774
Chicago/Turabian StylePang, Xiaoyan, Hai Shang, Boya Deng, Fang Wen, and Yi Zhang. 2013. "The Interaction of Adrenomedullin and Macrophages Induces Ovarian Cancer Cell Migration via Activation of RhoA Signaling Pathway" International Journal of Molecular Sciences 14, no. 2: 2774-2787. https://doi.org/10.3390/ijms14022774
APA StylePang, X., Shang, H., Deng, B., Wen, F., & Zhang, Y. (2013). The Interaction of Adrenomedullin and Macrophages Induces Ovarian Cancer Cell Migration via Activation of RhoA Signaling Pathway. International Journal of Molecular Sciences, 14(2), 2774-2787. https://doi.org/10.3390/ijms14022774