Citrange Fruit Extracts Alleviate Obesity-Associated Metabolic Disorder in High-Fat Diet-Induced Obese C57BL/6 Mouse

Abstract
:1. Introduction
2. Results
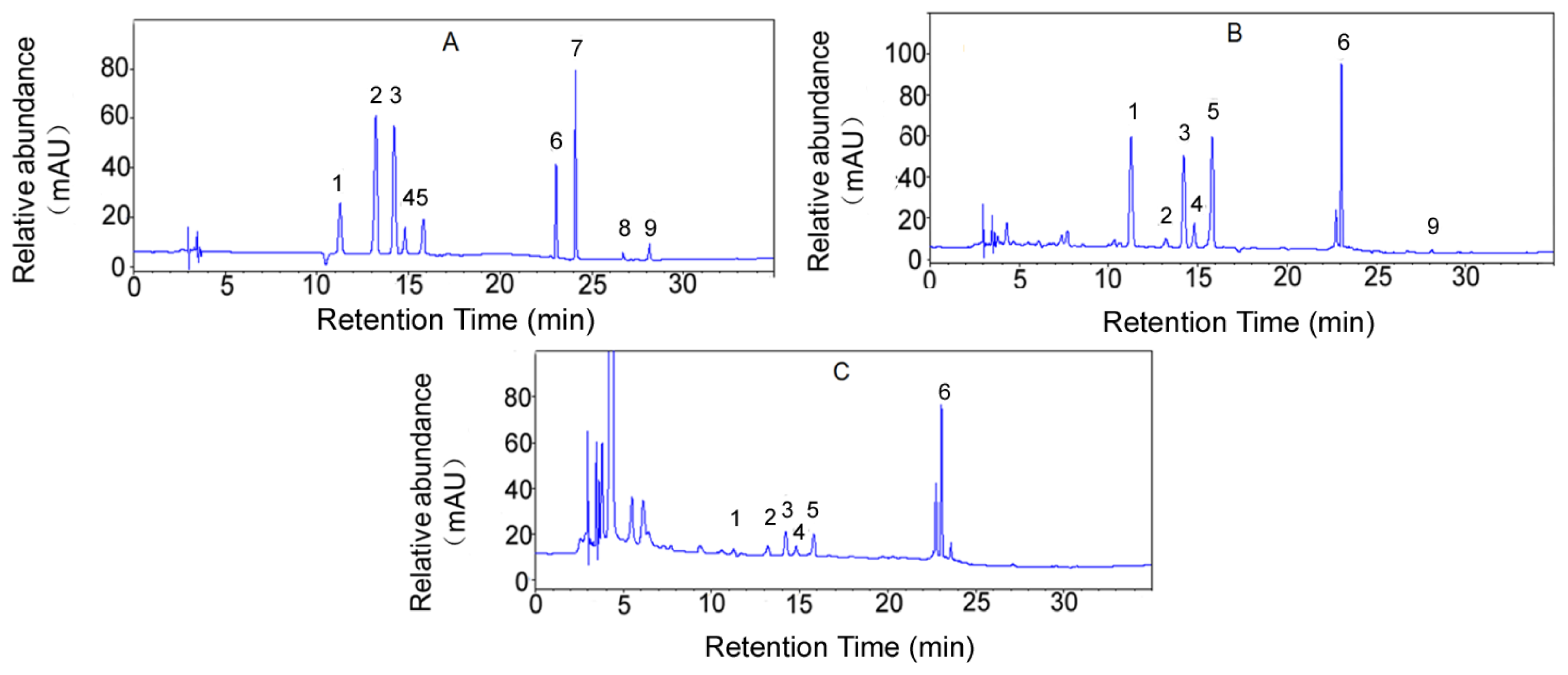
2.1. Flavonoid Contents in CPE and CFSE
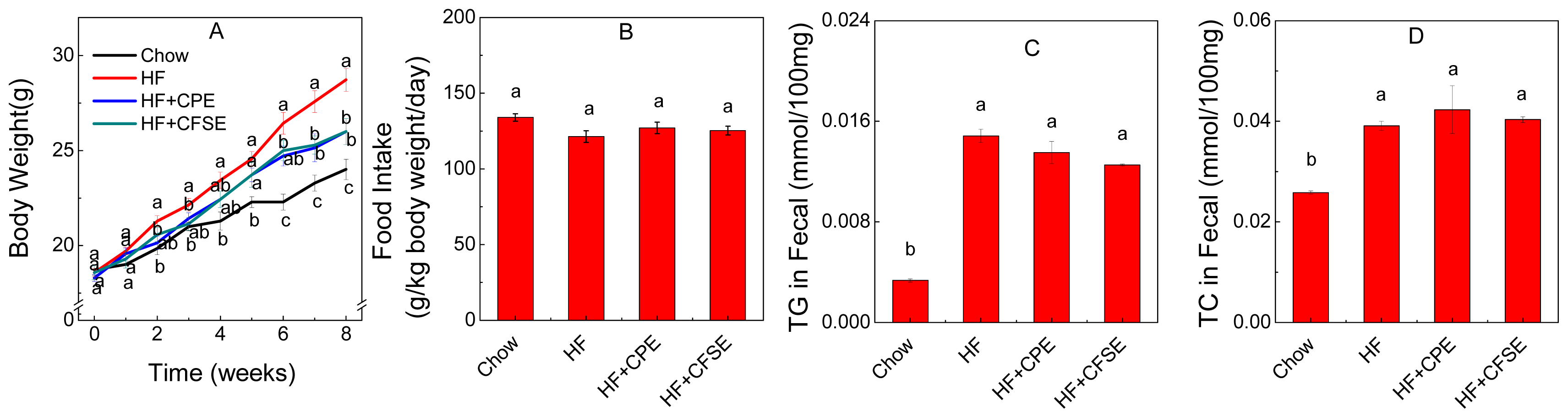
2.2. Body Weight Gain
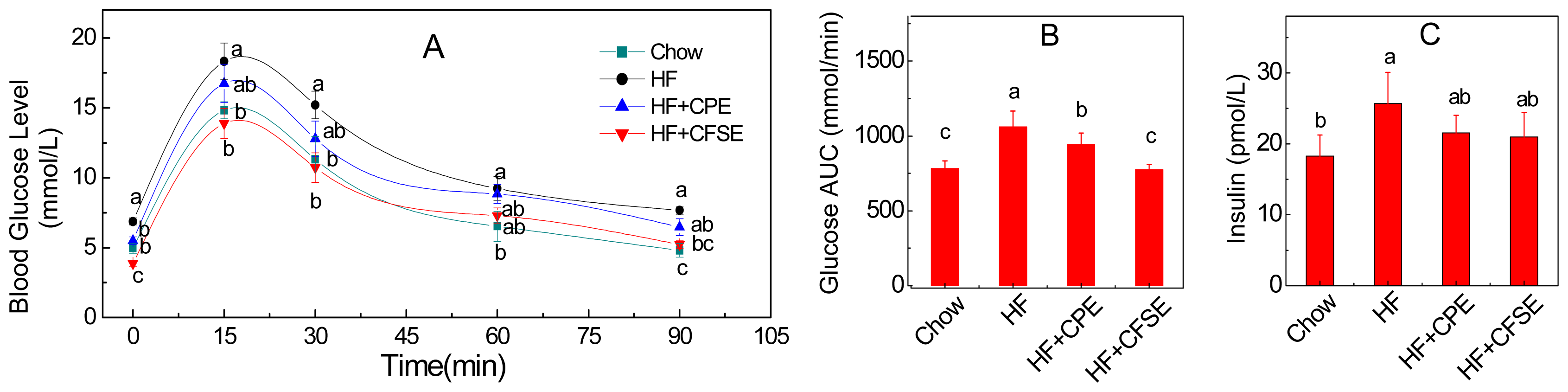
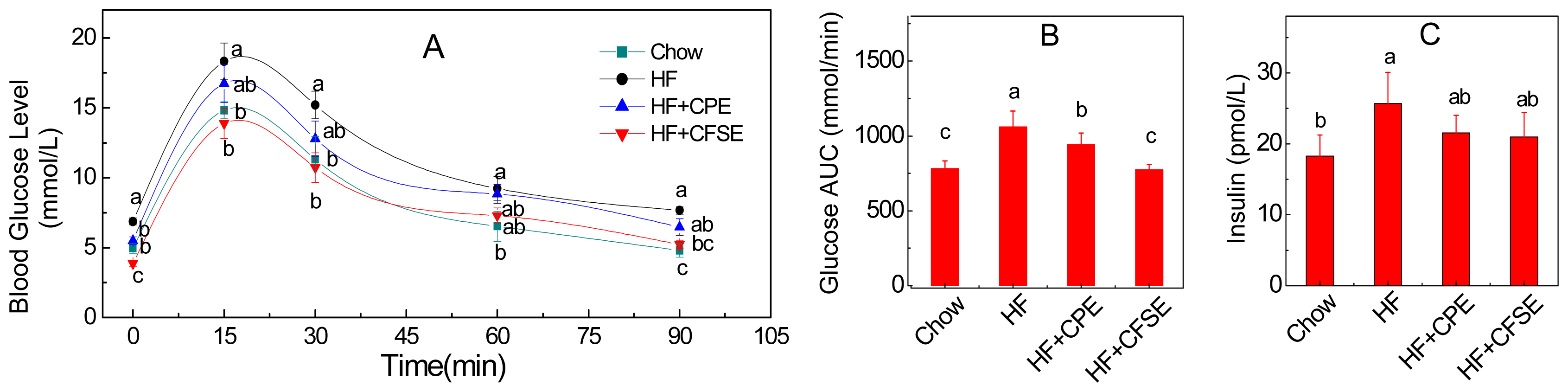
2.3. CPE and CFSE Improves Glucose Tolerance
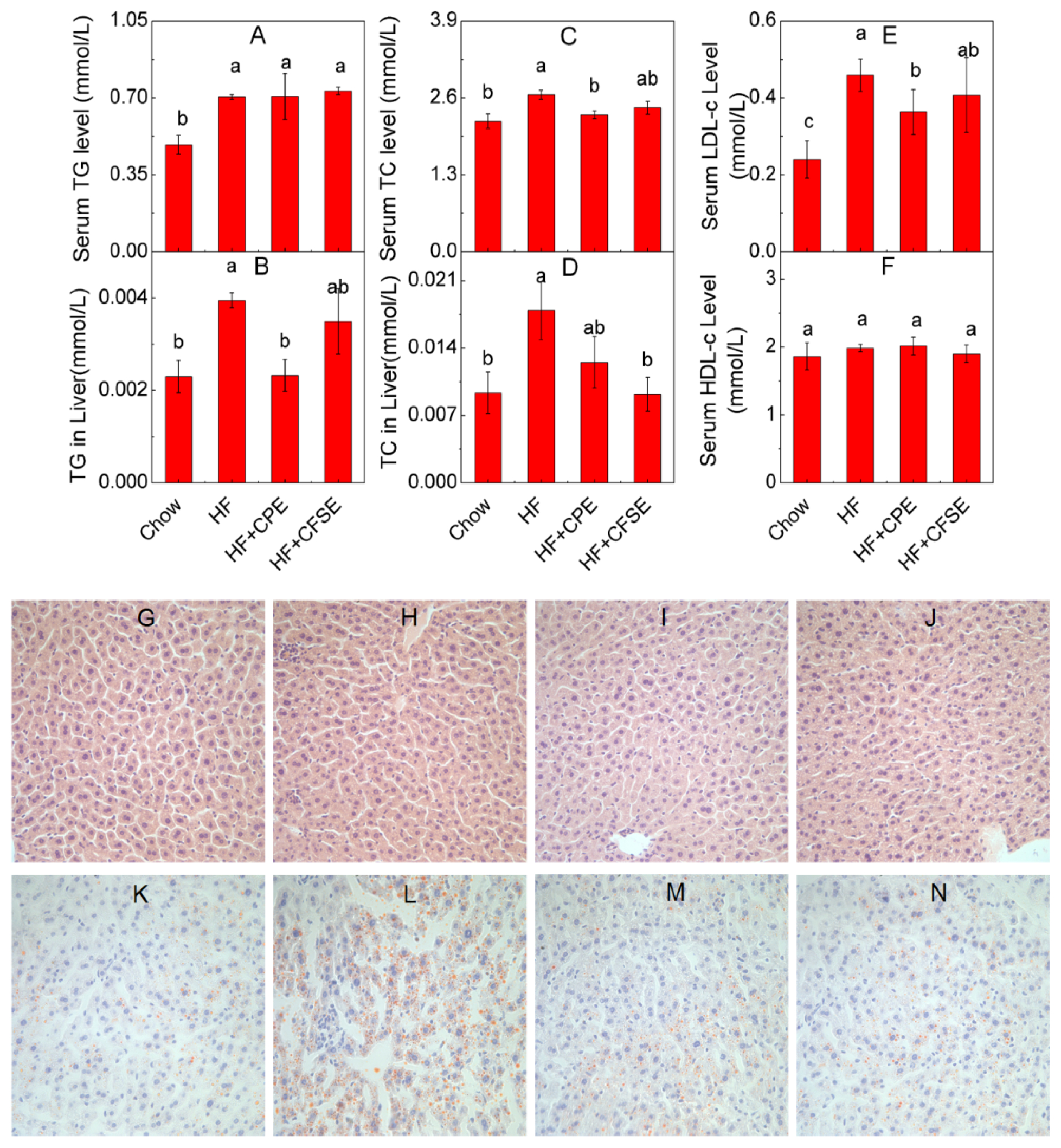
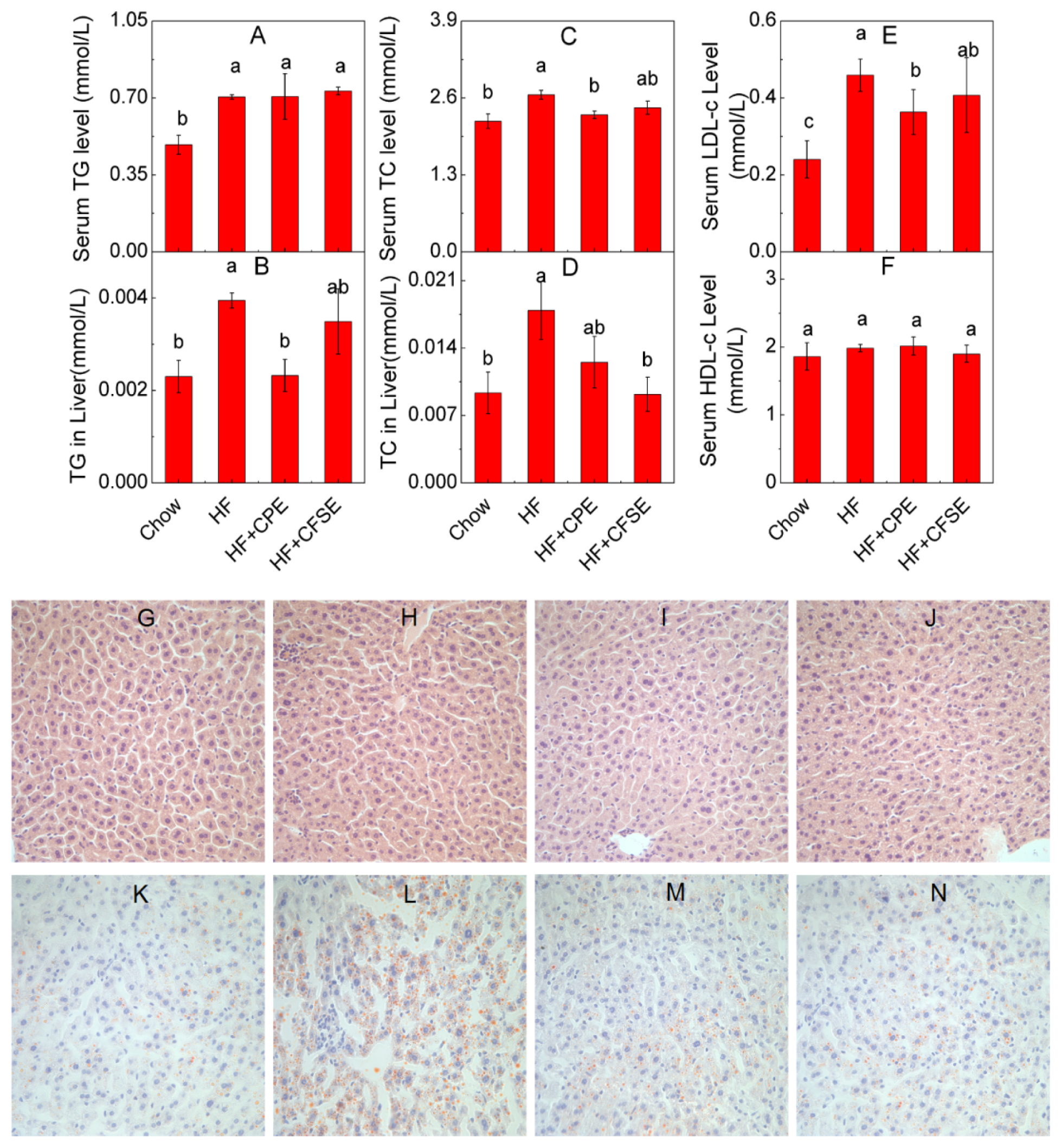
2.4. CPE and CFSE Prevents Lipid Accumulation
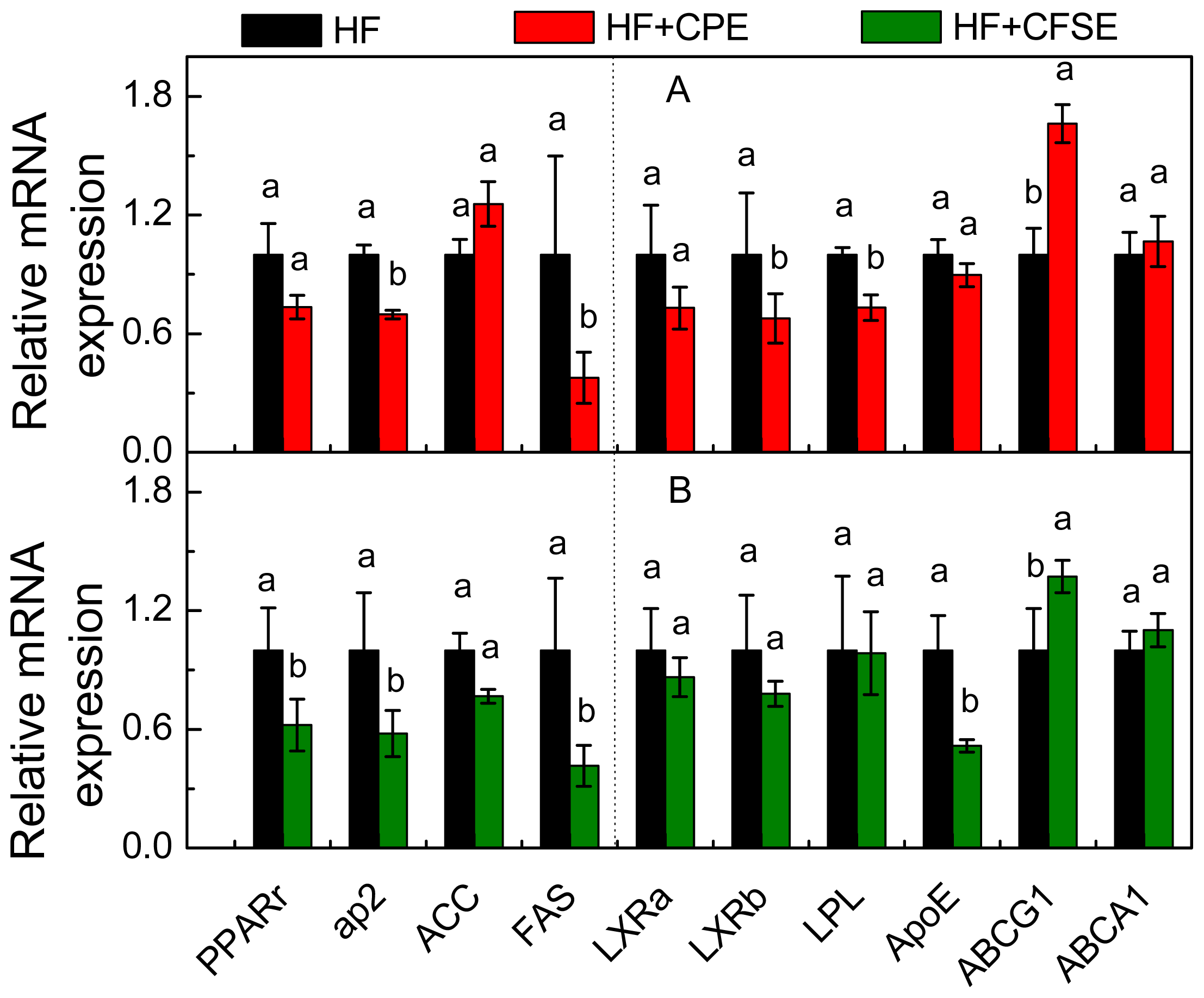
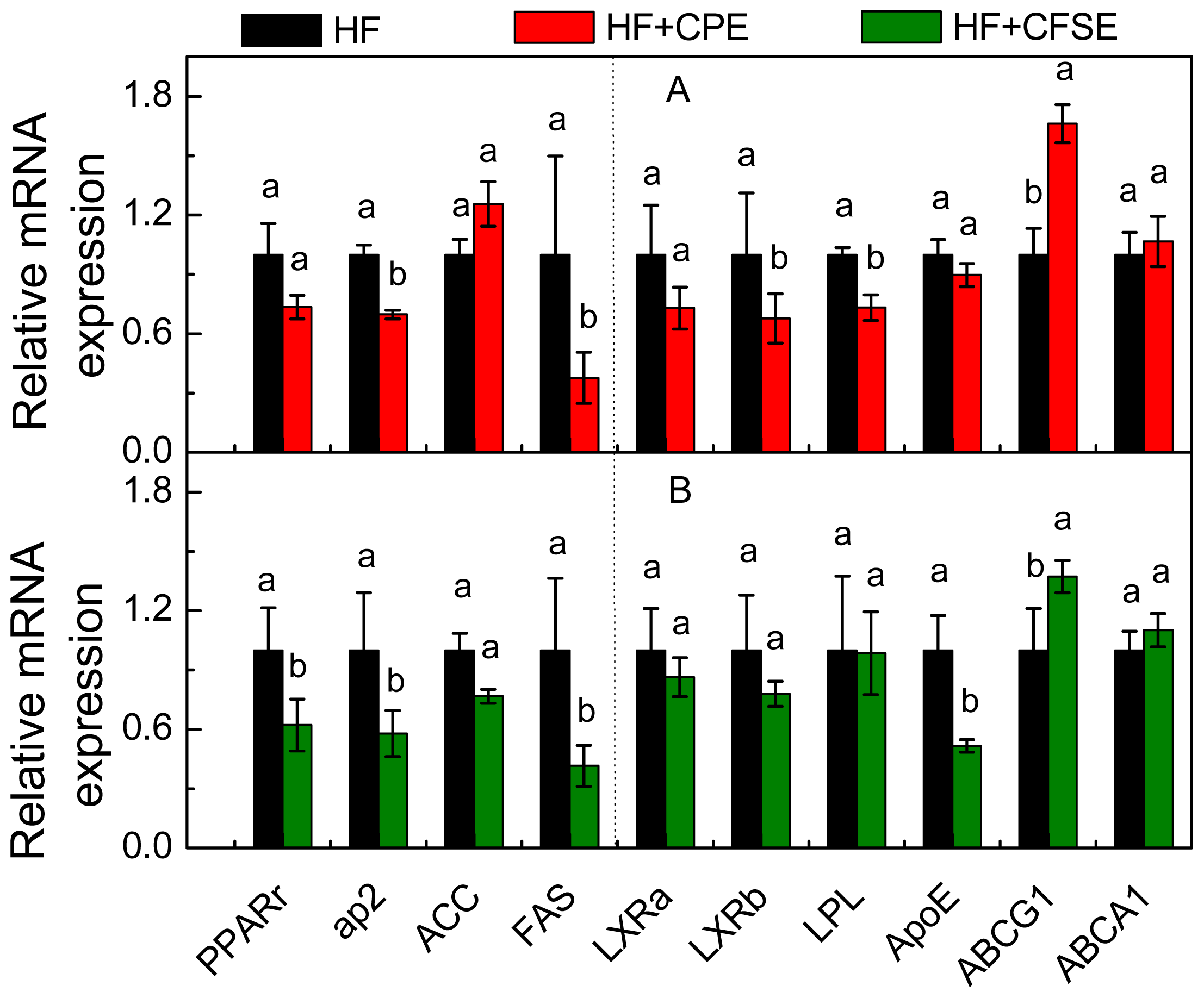
2.5. Gene Expression Analysis by Real-Time PCR
3. Discussion
4. Experimental Section
4.1. Preparation of Citrange Fruit Extracts
4.2. HPLC Analysis
4.3. Animals and Diets
4.4. Intraperitoneal Glucose Tolerance Test (ipGTT)
4.5. Serum Chemistry Analysis
4.6. Liver and Fecal Lipid Content Analysis
4.7. Histology of Liver
4.8. RNA Isolation and Gene Expression
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- WHO. Obesity and Overweight. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 23 June 2013).
- Fernstrom, J.D.; Choi, S. The development of tolerance to drugs that suppress food intake. Pharmacol. Ther 2008, 117, 105–122. [Google Scholar]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox, and impact of weight loss. J. Am. Coll. Cardiol 2009, 53, 1925–1932. [Google Scholar]
- Li, Z.; Maglione, M.; Tu, W.; Mojica, W.; Arterburn, D.; Shugarman, L.R.; Hilton, L.; Suttorp, M.; Solomon, V.; Shekelle, P.G.; et al. Meta-analysis: Pharmacologic treatment of obesity. Ann. Intern. Med 2005, 142, 532–546. [Google Scholar]
- Alappat, L.; Awad, A.B. Curcumin and obesity: Evidence and mechanisms. Nutr. Rev 2010, 68, 729–738. [Google Scholar]
- Alam, M.A.; Kauter, K.; Brown, L. Naringin improves diet-induced cardiovascular dysfunction and obesity in high carbohydrate, high fat diet-fed rats. Nutrients 2013, 5, 637–650. [Google Scholar]
- Jing, L.; Zhang, Y.; Fan, S.; Gu, M.; Guan, Y.; Lu, X.; Huang, C.; Zhou, Z. Preventive and ameliorating effects of citrus d-limonene on dyslipidemia and hyperglycemia in mice with high-fat diet-induced obesity. Eur. J. Pharmacol 2013, 715, 46–55. [Google Scholar]
- Choudhary, M.; Grover, K. Development of functional food products in relation to obesity. Funct. Foods Health Dis 2012, 2, 188–197. [Google Scholar]
- Vasudeva, N.; Yadav, N.; Sharma, S.K. Natural products: A safest approach for obesity. Chin. J. Integr. Med 2012, 18, 473–480. [Google Scholar]
- Gorinstein, S.; Caspi, A.; Libman, I.; Lerner, H.T.; Huang, D.; Leontowicz, H.; Leontowicz, M.; Tashma, Z.; Katrich, E.; Feng, S.; et al. Red grapefruit positively influences serum triglyceride level in patients suffering from coronary atherosclerosis: Studies in vitro and in humans. J. Agric. Food Chem 2006, 54, 1887–1892. [Google Scholar]
- Jung, U.J.; Kim, H.J.; Lee, J.S.; Lee, M.K.; Kim, H.O.; Park, E.J.; Kim, H.K.; Jeong, T.S.; Choi, M.S. Naringin supplementation lowers plasma lipids and enhances erythrocyte antioxidant enzyme activities in hypercholesterolemic subjects. Clin. Nutr 2003, 22, 561–568. [Google Scholar]
- Preuss, H.G.; DiFerdinando, D.; Bagchi, M.; Bagchi, D. Citrus aurantium as a thermogenic, weight-reduction replacement for ephedra: An overview. J. Med 2002, 33, 247–264. [Google Scholar]
- Yamada, T.; Hayasaka, S.; Shibata, Y.; Ojima, T.; Saegusa, T.; Gotoh, T.; Ishikawa, S.; Nakamura, Y.; Kayaba, K. Frequency of citrus fruit intake is associated with the incidence of cardiovascular disease: The Jichi Medical School cohort study. J. Epidemiol. Jpn. Epidemiol. Assoc 2011, 21, 169–175. [Google Scholar]
- Shin, H.S.; Kang, S.; Ko, H.C.; Kim, H.M.; Hong, Y.S.; Yoon, S.A.; Kim, S.J. Anti-inflammatory effect of the immature peel extract of Jinkyool (Citrus sunki Hort. ex Tanaka). Food Sci. Biotechnol 2011, 20, 1235–1241. [Google Scholar]
- Tanaka, T.; Tanaka, M.; Kuno, T. Cancer chemoprevention by citrus pulp and juices containing high amounts of beta-cryptoxanthin and hesperidin. J. Biomed. Biotechnol 2012, 2012. [Google Scholar] [CrossRef]
- Manthey, J.A.; Grohmann, K. Phenols in citrus peel byproducts. Concentrations of hydroxycinnamates and polymethoxylated flavones in citrus peel molasses. J. Agric. Food Chem 2001, 49, 3268–3273. [Google Scholar]
- Neville, F.; Pchelintsev, N.A.; Broderick, M.J.; Gibson, T.; Millner, P.A. Novel one-pot synthesis and characterization of bioactive thiol-silicate nanoparticles for biocatalytic and biosensor applications. Nanotechnology 2009, 20. [Google Scholar] [CrossRef]
- People’s Republic of China Ministry of Health, Chinese Pharmacopoeia, 8th ed.; Chemical Industry Press: Beijing, China, 2005.
- Jung, H.; Jeong, Y.; Park, C.D.; Park, C.H.; Hong, J.H. Inhibitory effect of citrus peel extract on lipid accumulation of 3T3-L1 adipocytes. J. Korean Soc. Appl. Biol. Chem 2011, 54, 169–176. [Google Scholar]
- Tsujita, T.; Takaku, T. Lipolysis induced by segment wall extract from Satsuma mandarin orange (Citrus unshu Mark). J. Nutr. Sci. Vitaminol 2007, 53, 547–551. [Google Scholar]
- Bok, S.H.; Lee, S.H.; Park, Y.B.; Bae, K.H.; Son, K.H.; Jeong, T.S.; Choi, M.S. Plasma and hepatic cholesterol and hepatic activities of 3-hydroxy-3-methyl-glutaryl-CoA reductase and acyl CoA: Cholesterol transferase are lower in rats fed citrus peel extract or a mixture of citrus bioflavonoids. J. Nutr 1999, 129, 1182–1185. [Google Scholar]
- Kang, S.I.; Shin, H.S.; Kim, H.M.; Hong, Y.S.; Yoon, S.A.; Kang, S.W.; Kim, J.H.; Kim, M.H.; Ko, H.C.; Kim, S.J. Immature Citrus sunki peel extract exhibits antiobesity effects by beta-oxidation and lipolysis in high-fat diet-induced obese mice. Biol. Pharm. Bull 2012, 35, 223–230. [Google Scholar]
- Ding, X.; Fan, S.; Lu, Y.; Zhang, Y.; Gu, M.; Zhang, L.; Liu, G.; Guo, L.; Jiang, D.; Lu, X.; et al. Citrus ichangensis peel extract exhibits anti-metabolic disorder effects by the inhibition of PPAR and LXR signaling in high-fat diet-induced C57BL/6 mouse. Evid. Based Complement. Altern. Med 2012, 2012. [Google Scholar] [CrossRef]
- Goldwasser, J.; Cohen, P.Y.; Yang, E.; Balaguer, P.; Yarmush, M.L.; Nahmias, Y. Transcriptional regulation of human and rat hepatic lipid metabolism by the grapefruit flavonoid naringenin: Role of PPARalpha, PPARgamma and LXRalpha. PLoS One 2010, 5, e12399. [Google Scholar]
- Lee, C.H.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med 2004, 10, 355–361. [Google Scholar]
- Pakala, R.; Rha, S.W.; Kuchulakanti, P.K.; Cheneau, E.; Baffour, R.; Waksman, R. Peroxisome proliferator-activated receptor gamma; Its role in atherosclerosis and restenosis. Cardiovasc. Radiat. Med 2004, 5, 44–48. [Google Scholar]
- Cho, K.W.; Kim, Y.O.; Andrade, J.E.; Burgess, J.R.; Kim, Y.C. Dietary naringenin increases hepatic peroxisome proliferators-activated receptor alpha protein expression and decreases plasma triglyceride and adiposity in rats. Eur. J. Nutr 2011, 50, 81–88. [Google Scholar]
- Kim, G.S.; Park, H.J.; Woo, J.H.; Kim, M.K.; Koh, P.O.; Min, W.; Ko, Y.G.; Kim, C.H.; Won, C.K.; Cho, J.H. Citrus aurantium flavonoids inhibit adipogenesis through the Akt signaling pathway in 3T3-L1 cells. BMC Complement. Altern. Med 2012, 12, 31. [Google Scholar]
- Kurowska, E.M.; Manthey, J.A.; Casaschi, A.; Theriault, A.G. Modulation of HepG2 cell net apolipoprotein B secretion by the citrus polymethoxy flavone, tangeretin. Lipids 2004, 39, 143–151. [Google Scholar]
- Li, R.W.; Theriault, A.G.; Au, K.; Douglas, T.D.; Casaschi, A.; Kurowska, E.M.; Mukherjee, R. Citrus polymethoxylated flavones improve lipid and glucose homeostasis and modulate adipocytokines in fructose-induced insulin resistant hamsters. Life Sci 2006, 79, 365–373. [Google Scholar]
- Haaz, S.; Fontaine, K.R.; Cutter, G.; Limdi, N.; Perumean-Chaney, S.; Allison, D.B. Citrus aurantium and synephrine alkaloids in the treatment of overweight and obesity: An update. Obes. Rev 2006, 7, 79–88. [Google Scholar]
- Sharma, A.K.; Bharti, S.; Ojha, S.; Bhatia, J.; Kumar, N.; Ray, R.; Kumari, S.; Arya, D.S. Up-regulation of PPARgamma, heat shock protein-27 and -72 by naringin attenuates insulin resistance, beta-cell dysfunction, hepatic steatosis and kidney damage in a rat model of type 2 diabetes. Br. J. Nutr 2011, 106, 1713–1723. [Google Scholar]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid composition of fruit tissues of citrus species. Biosci. Biotechnol. Biochem 2006, 70, 178–192. [Google Scholar]
- Avula, B.; Joshi, V.C.; Weerasooriya, A.; Khan, I.A. Liquid hromatography for separation and quantitative determination of adrenergic amines and flavonoids from Poncirus trifoliatus raf. fruits at different stages of growth. Chromatographia 2005, 62, 379–383. [Google Scholar]
- Zhu, X.; Luo, F.; Zheng, Y.; Zhang, J.; Huang, J.; Sun, C.; Li, X.; Chen, K. Characterization, purification of poncirin from edible citrus ougan (Citrus reticulate cv. suavissima) and its growth inhibitory effect on human gastric cancer cells SGC-7901. Int. J. Mol. Sci 2013, 14, 8684–8697. [Google Scholar]
- Yoon, H.Y.; Yun, S.I.; Kim, B.Y.; Jin, Q.; Woo, E.R.; Jeong, S.Y.; Chung, Y.S. Poncirin promotes osteoblast differentiation but inhibits adipocyte differentiation in mesenchymal stem cells. Eur. J. Pharmacol 2011, 664, 54–59. [Google Scholar]
- Pu, P.; Gao, D.-M.; Mohamed, S.; Chen, J.; Zhang, J.; Zhou, X.-Y.; Zhou, N.-J.; Xie, J.; Jiang, H. Naringin ameliorates metabolic syndrome by activating AMP-activated protein kinase in mice fed a high-fat diet. Arch. Biochem. Biophys 2012, 518, 61–70. [Google Scholar]
- Chandra Jagetia, G.; Koti Reddy, T.; Venkatesha, V.A.; Kedlaya, R. Influence of naringin on ferric iron induced oxidative damage. in vitro. Clin. Chim. Acta 2004, 347, 189–197. [Google Scholar]
- Camargo, C.A.; Gomes-Marcondes, M.C.C.; Wutzki, N.C.; Aoyama, H. Naringin inhibits tumor growth and reduces interleukin-6 and tumor necrosis factor α levels in rats with Walker 256 carcinosarcoma. Anticancer Res 2012, 32, 129–133. [Google Scholar]
- Inês Amaro, M.; Rocha, J.; Vila-Real, H.; Eduardo-Figueira, M.; Mota-Filipe, H.; Sepodes, B.; Ribeiro, M.H. Anti-inflammatory activity of naringin and the biosynthesised naringenin by naringinase immobilized in microstructured materials in a model of DSS-induced colitis in mice. Food Res. Int 2009, 42, 1010–1017. [Google Scholar]
- Lee, S.; Muniyappa, R.; Yan, X.; Chen, H.; Yue, L.Q.; Hong, E.G.; Kim, J.K.; Quon, M.J. Comparison between surrogate indexes of insulin sensitivity and resistance and hyperinsulinemic euglycemic clamp estimates in mice. Am. J. Physiol. Endocrinol. Metab 2008, 294, E261–E270. [Google Scholar]
- Kim, J.K.; Gavrilova, O.; Chen, Y.; Reitman, M.L.; Shulman, G.I. Mechanism of insulin resistance in A-ZIP/F-1 fatless mice. J. Biol. Chem 2000, 275, 8456–8460. [Google Scholar]
- McGarry, J.D.; Foster, D.W. Regulation of hepatic fatty acid oxidation and ketone body production. Annu. Rev. Biochem 1980, 49, 395–420. [Google Scholar]
- Tripoli, E.; Guardia, M.A.; Giammanco, S.; Majo, D.D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem 2007, 104, 466–479. [Google Scholar]
- Choi, M.S.; Do, K.M.; Park, Y.S.; Jeon, S.M.; Jeong, T.S.; Lee, Y.K.; Lee, M.K.; Bok, S.H. Effect of naringin supplementation on cholesterol metabolism and antioxidant status in rats fed high cholesterol with different levels of vitamin E. Ann. Nutr. Metab 2001, 45, 193–201. [Google Scholar]
- Jung, U.J.; Lee, M.K.; Jeong, K.S.; Choi, M.S. The hypoglycemic effects of hesperidin and naringin are partly mediated by hepatic glucose-regulating enzymes in C57BL/KsJ-db/db mice. J. Nutr 2004, 134, 2499–2503. [Google Scholar]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Kang, M.A.; Choi, M.S. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mRNA levels in type-2 diabetic mice. Int. J. Biochem. Cell Biol 2006, 38, 1134–1145. [Google Scholar]
- Flier, J.S. Obesity wars: Molecular progress confronts an expanding epidemic. Cell 2004, 116, 337–350. [Google Scholar]
- Zhang, Y.; Fan, S.J.; Hu, N.; Gu, M.; Chu, C.; Li, Y.; Lu, X.; Huang, C. Rhein reduces fat weight in db/db mouse and prevents diet-induced obesity in C57BL/6 mouse through the inhibition of PPARgamma signaling. PPAR Res 2012, 2012. [Google Scholar] [CrossRef]
- Gong, Z.; Huang, C.; Sheng, X.; Zhang, Y.; Li, Q.; Wang, M.W.; Peng, L.; Zang, Y.Q. The role of tanshinone IIA in the treatment of obesity through peroxisome proliferator-activated receptor gamma antagonism. Endocrinology 2009, 150, 104–113. [Google Scholar]
- Mitro, N.; Mak, P.A.; Vargas, L.; Godio, C.; Hampton, E.; Molteni, V.; Kreusch, A.; Saez, E. The nuclear receptor LXR is a glucose sensor. Nature 2007, 445, 219–223. [Google Scholar]
- Pruessner, J.C.; Kirschbaum, C.; Meinlschmid, G.; Hellhammer, D.H. Two formulas for computation of the area under the curve represent measures of total hormone concentration versus time-dependent change. Psychoneuroendocrinology 2003, 28, 916–931. [Google Scholar]
- Chun, T.H.; Inoue, M.; Morisaki, H.; Yamanaka, I.; Miyamoto, Y.; Okamura, T.; Sato-Kusubata, K.; Weiss, S.J. Genetic link between obesity and MMP14-dependent adipogenic collagen turnover. Diabetes 2010, 59, 2484–2494. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward primer | Reverse primer |
|---|---|---|
| β-Actin | TGTCCACCTTCCAGCAGATGT | AGCTCAGTAACAGTCCGCCTAGA |
| PPARγ | CGCTGATGCACTGCCTATGA | AGAGGTCCACAGAGCTGATTCC |
| aP2 | CATGGCCAAGCCCAACAT | CGCCCAGTTTGAAGGAAATC |
| ACC | GAATCTCCTGGTGACAATGCTTATT | GGTCTTGCTGAGTTGGGTTAGCT |
| FAS | CTGAGATCCCAGCACTTCTTGA | GCCTCCGAAGCCAAATGAG |
| LXRα | GAGTGTCGACTTCGCAAATGC | AGCTCAGTAACAGTCCGCCTAGA |
| LXRβ | CAGGCTTGCAGGTGGAATTC | ATGGCGATAAGCAAGGCATACT |
| ApoE | GAACCGCTTCTGGGATTACCT | TCAGTGCCGTCAGTTCTTGTG |
| LPL | ATCGGAGAACTGCTCATGATGA | CGGATCCTCTCGATGACGAA |
| ABCG1 | TCCCCACCTGTAAGTAATTGCA | TCGGACCCTTATCATTCTCTACAGA |
| ABCA1 | GGCAATGAGTGTGCCAGAGTTA | TAGTCACATGTGGCACCGTTTT |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lu, Y.; Xi, W.; Ding, X.; Fan, S.; Zhang, Y.; Jiang, D.; Li, Y.; Huang, C.; Zhou, Z. Citrange Fruit Extracts Alleviate Obesity-Associated Metabolic Disorder in High-Fat Diet-Induced Obese C57BL/6 Mouse. Int. J. Mol. Sci. 2013, 14, 23736-23750. https://doi.org/10.3390/ijms141223736
Lu Y, Xi W, Ding X, Fan S, Zhang Y, Jiang D, Li Y, Huang C, Zhou Z. Citrange Fruit Extracts Alleviate Obesity-Associated Metabolic Disorder in High-Fat Diet-Induced Obese C57BL/6 Mouse. International Journal of Molecular Sciences. 2013; 14(12):23736-23750. https://doi.org/10.3390/ijms141223736
Chicago/Turabian StyleLu, Yan, Wanpeng Xi, Xiaobo Ding, Shengjie Fan, Yu Zhang, Dong Jiang, Yiming Li, Cheng Huang, and Zhiqin Zhou. 2013. "Citrange Fruit Extracts Alleviate Obesity-Associated Metabolic Disorder in High-Fat Diet-Induced Obese C57BL/6 Mouse" International Journal of Molecular Sciences 14, no. 12: 23736-23750. https://doi.org/10.3390/ijms141223736