Protective Role of Andrographolide in Bleomycin-Induced Pulmonary Fibrosis in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
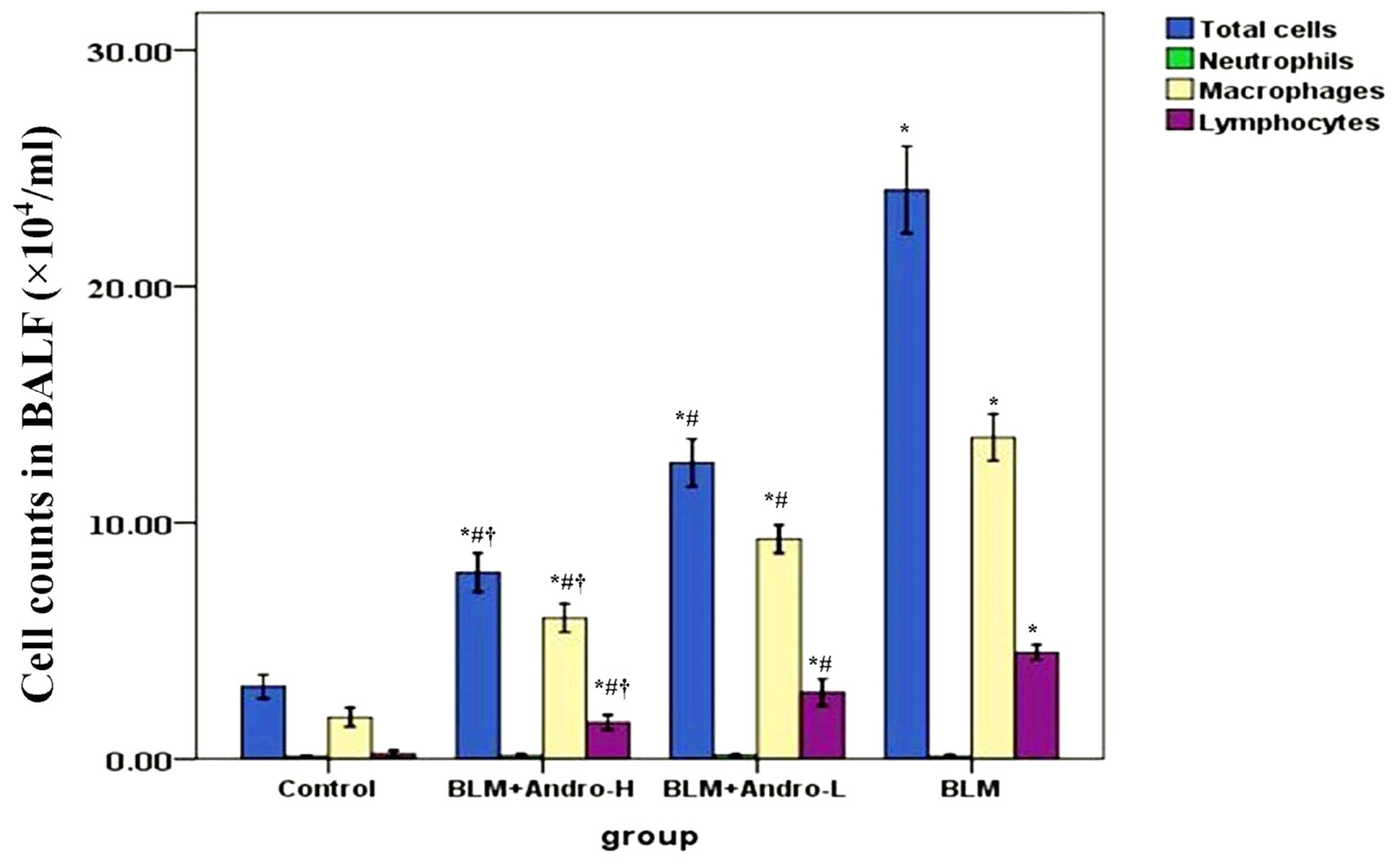
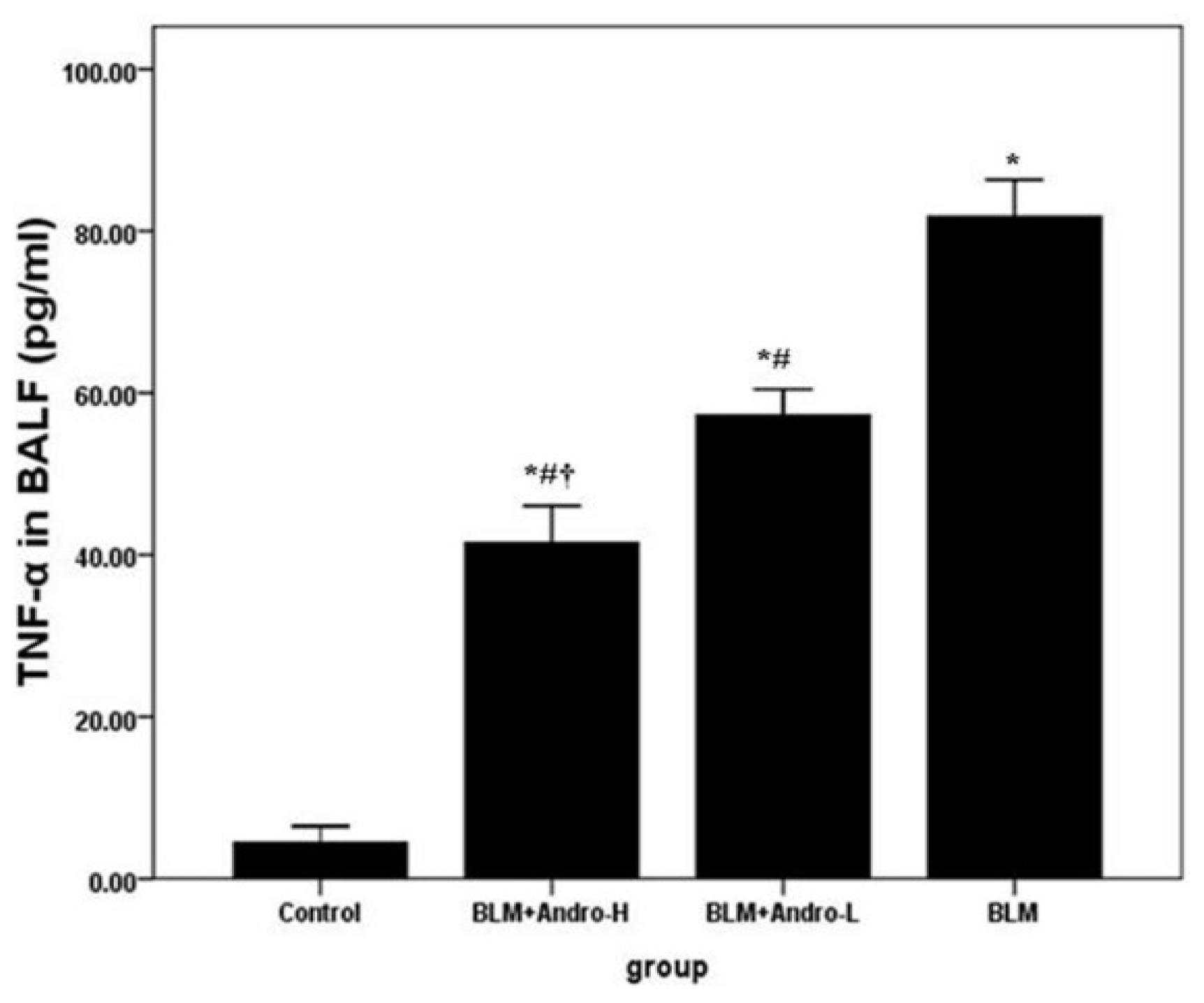
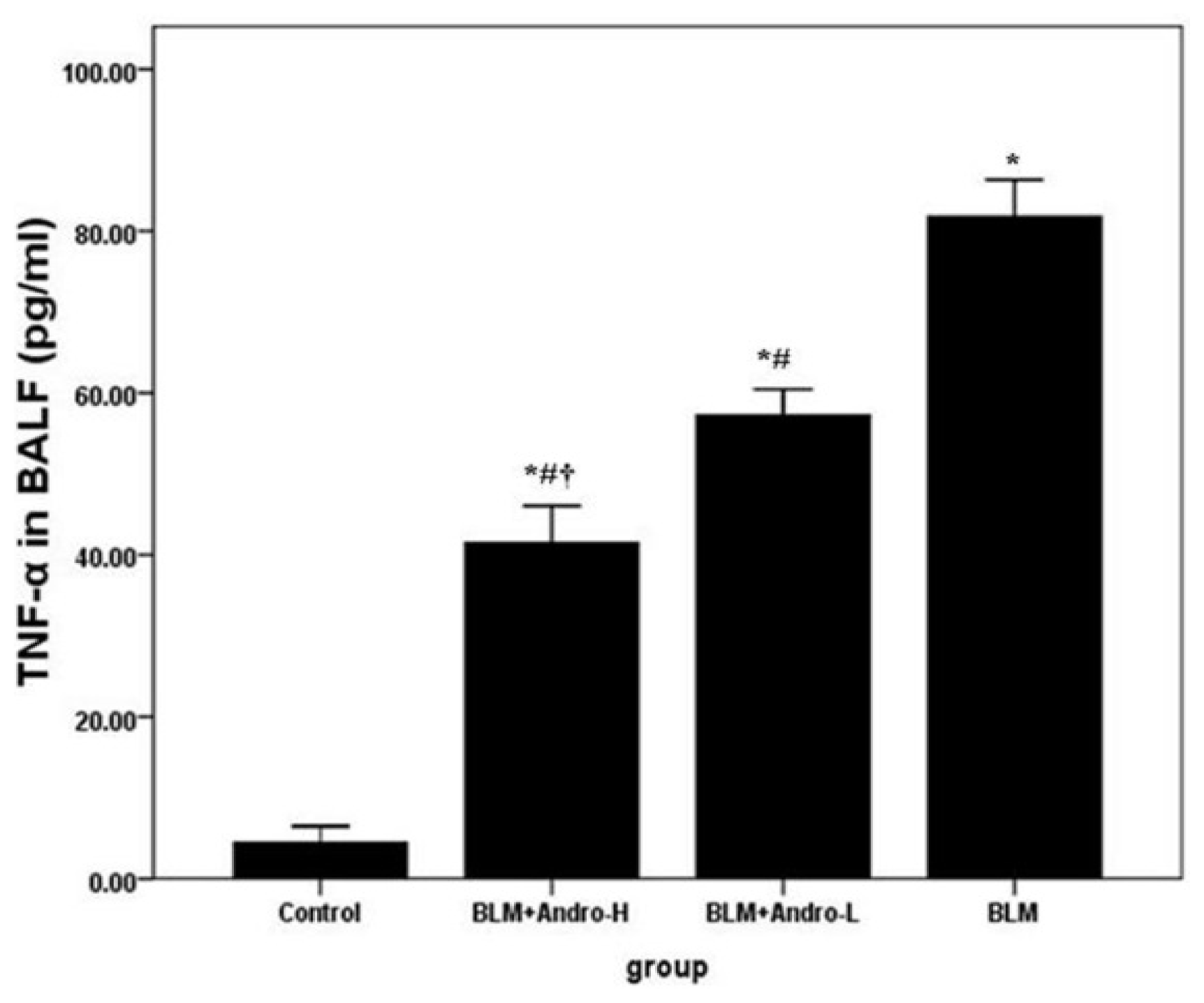
2.1. Andrographolide Reduces Cell Counts and TNF-α in BALF in BLM-Induced Pulmonary Fibrosis in Mice
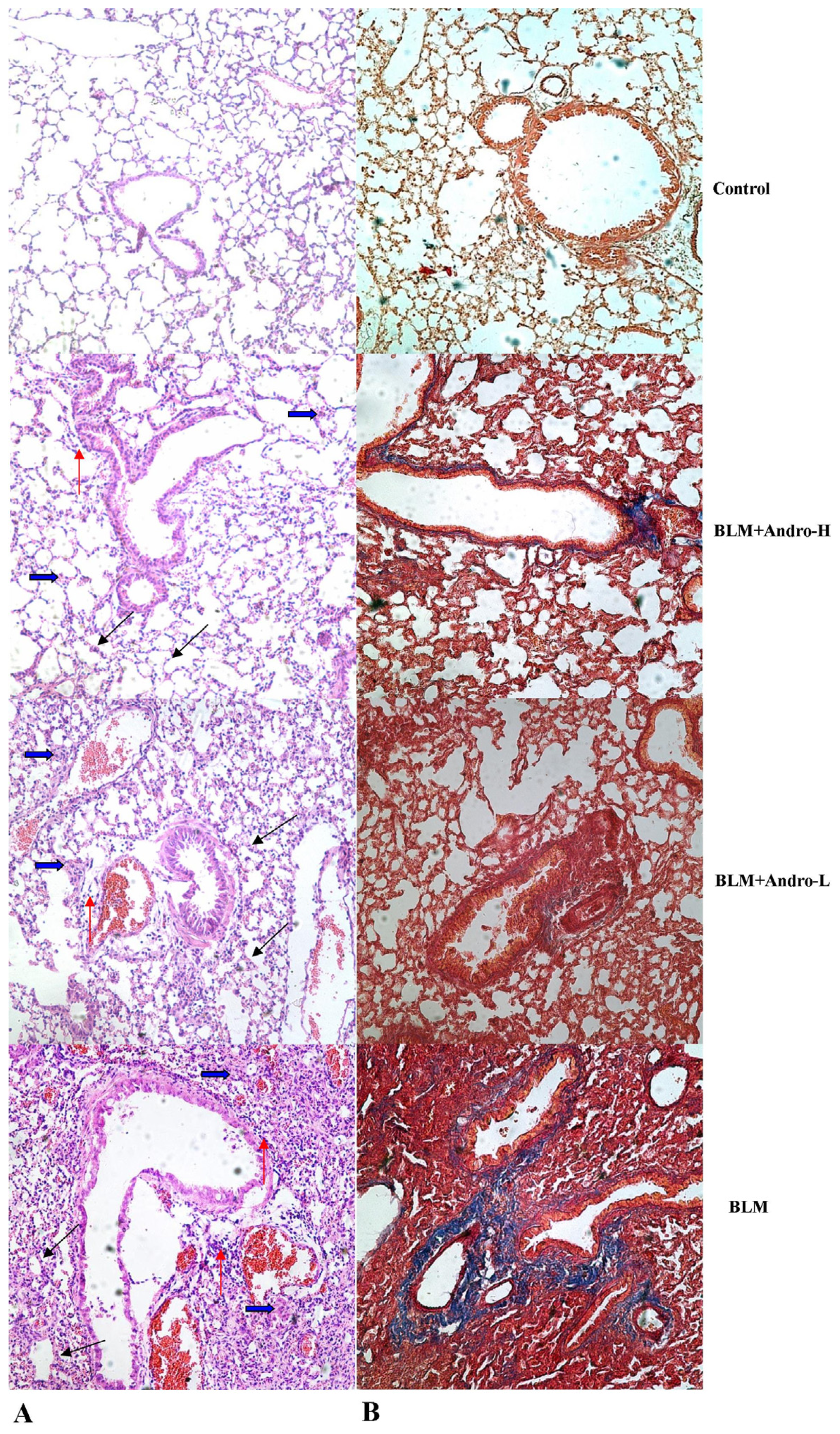
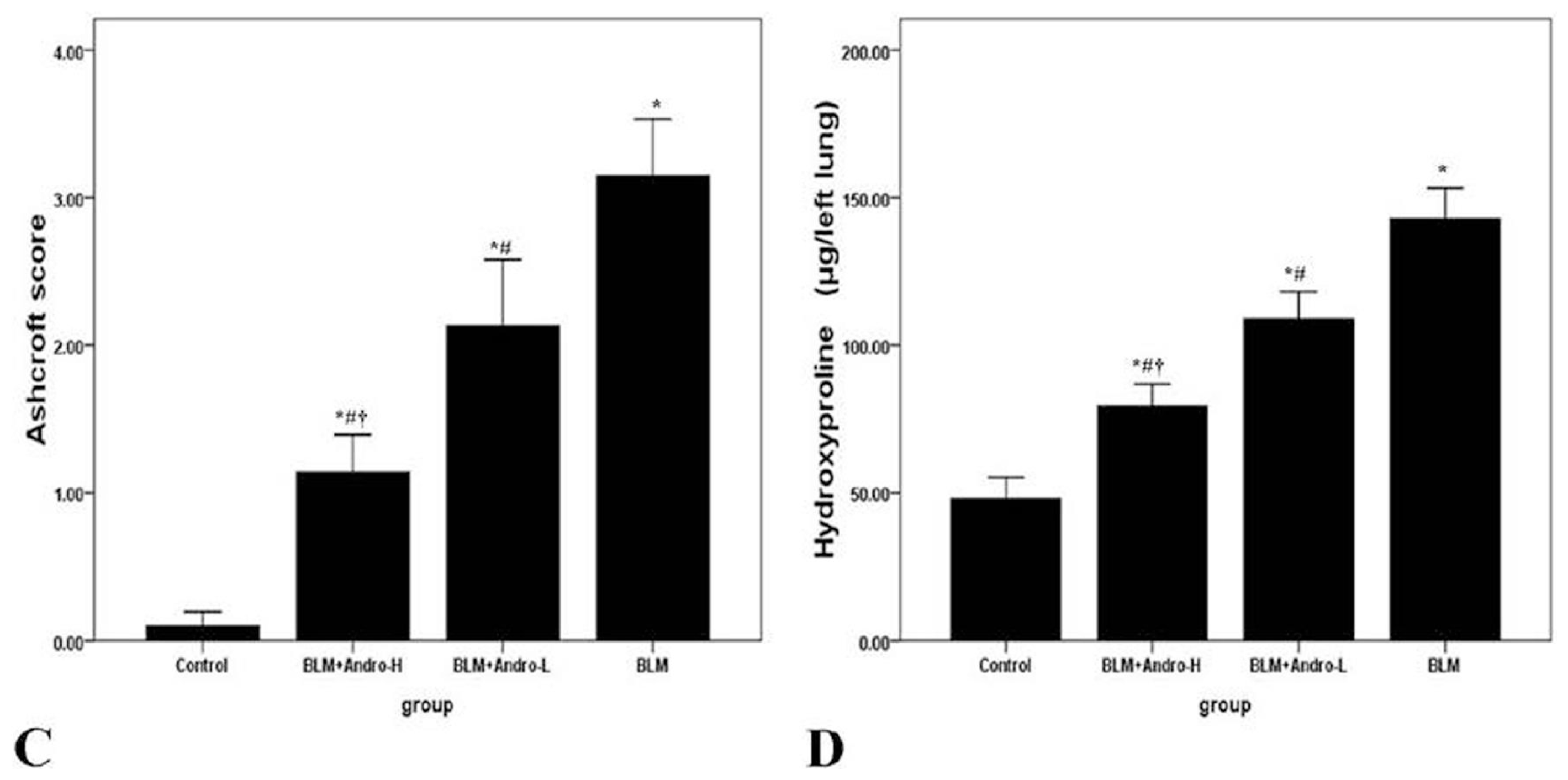
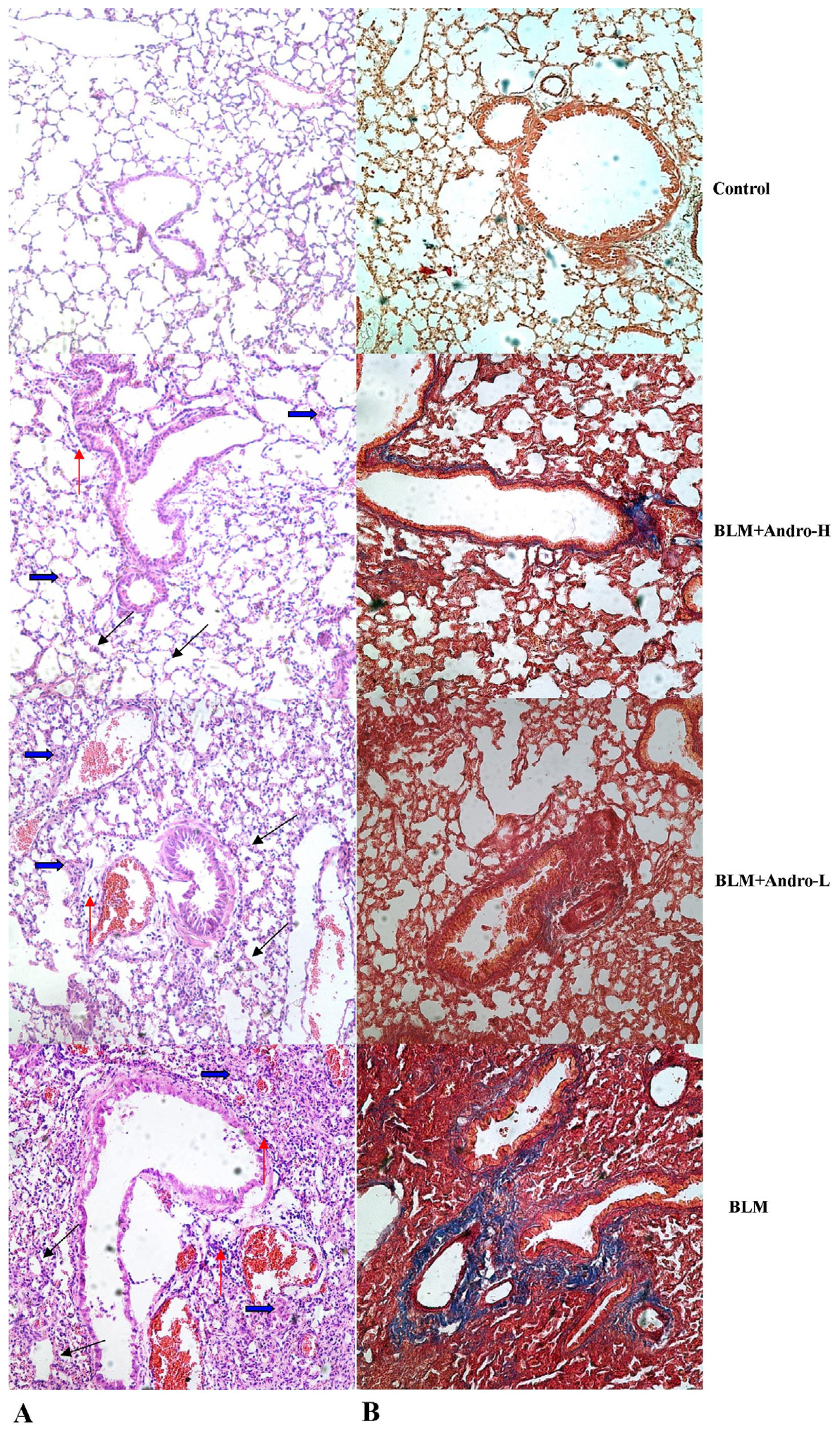
2.2. Andrographolide Attenuates BLM Induced Pulmonary Fibrosis in Mice
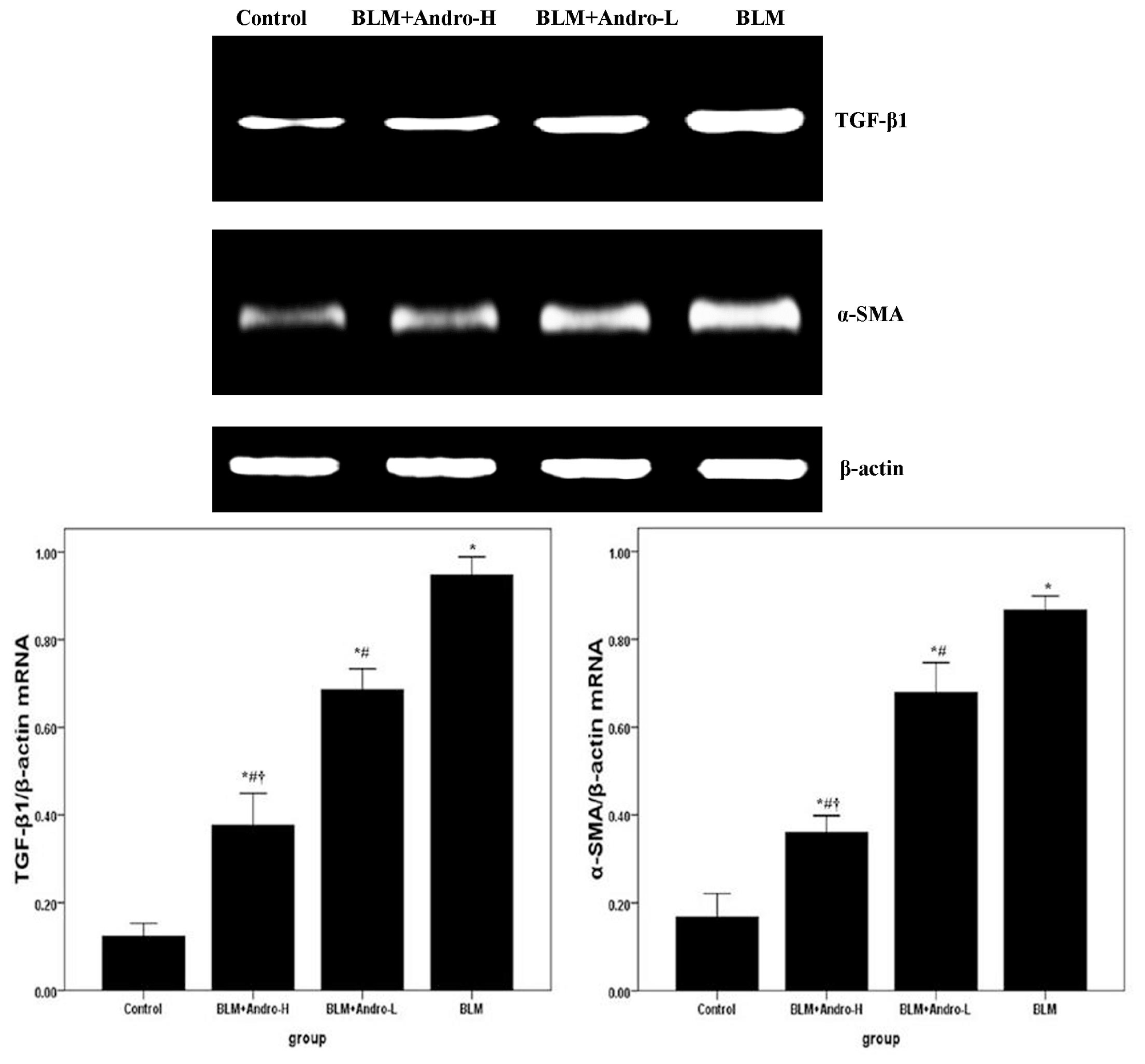
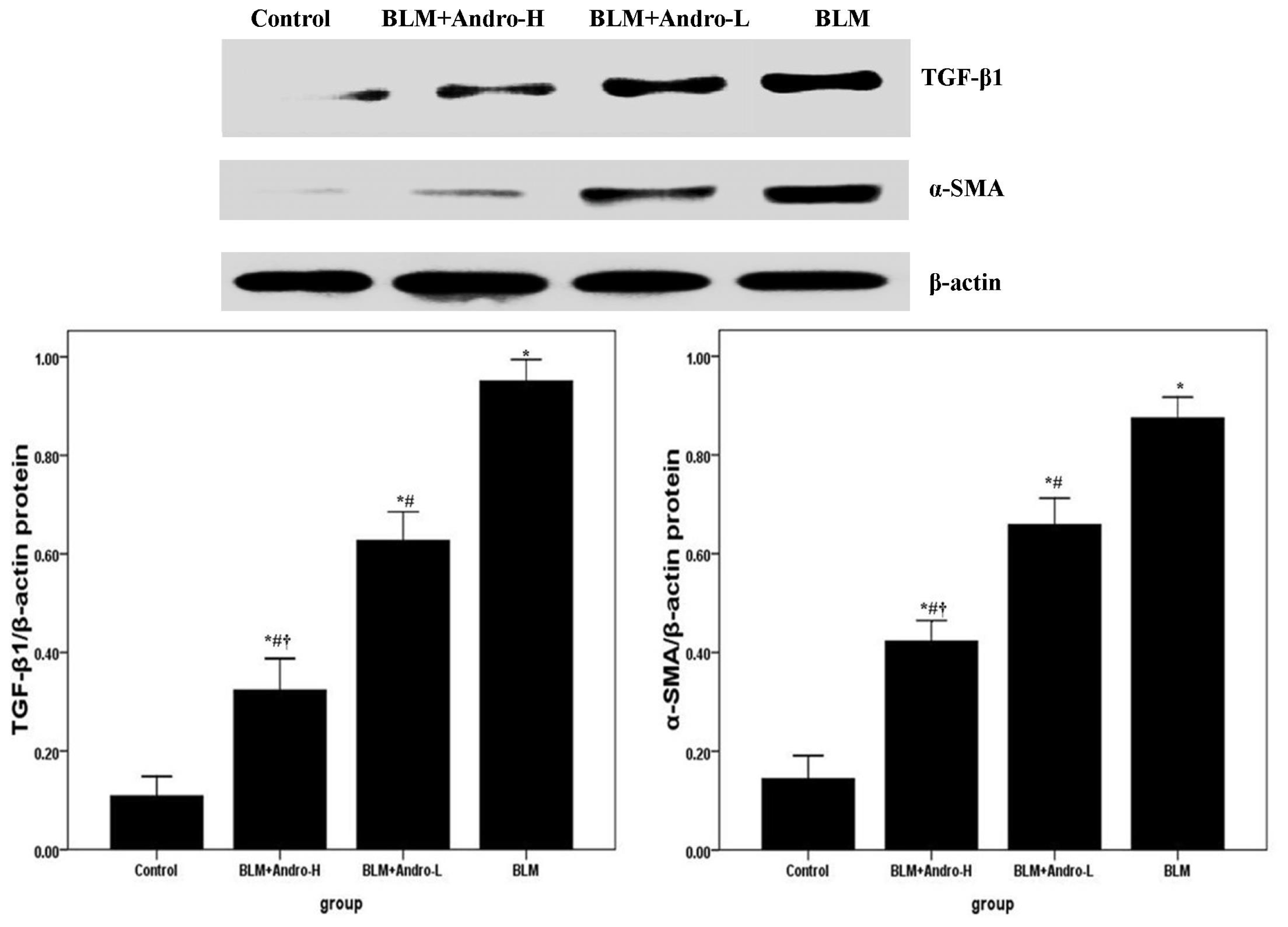
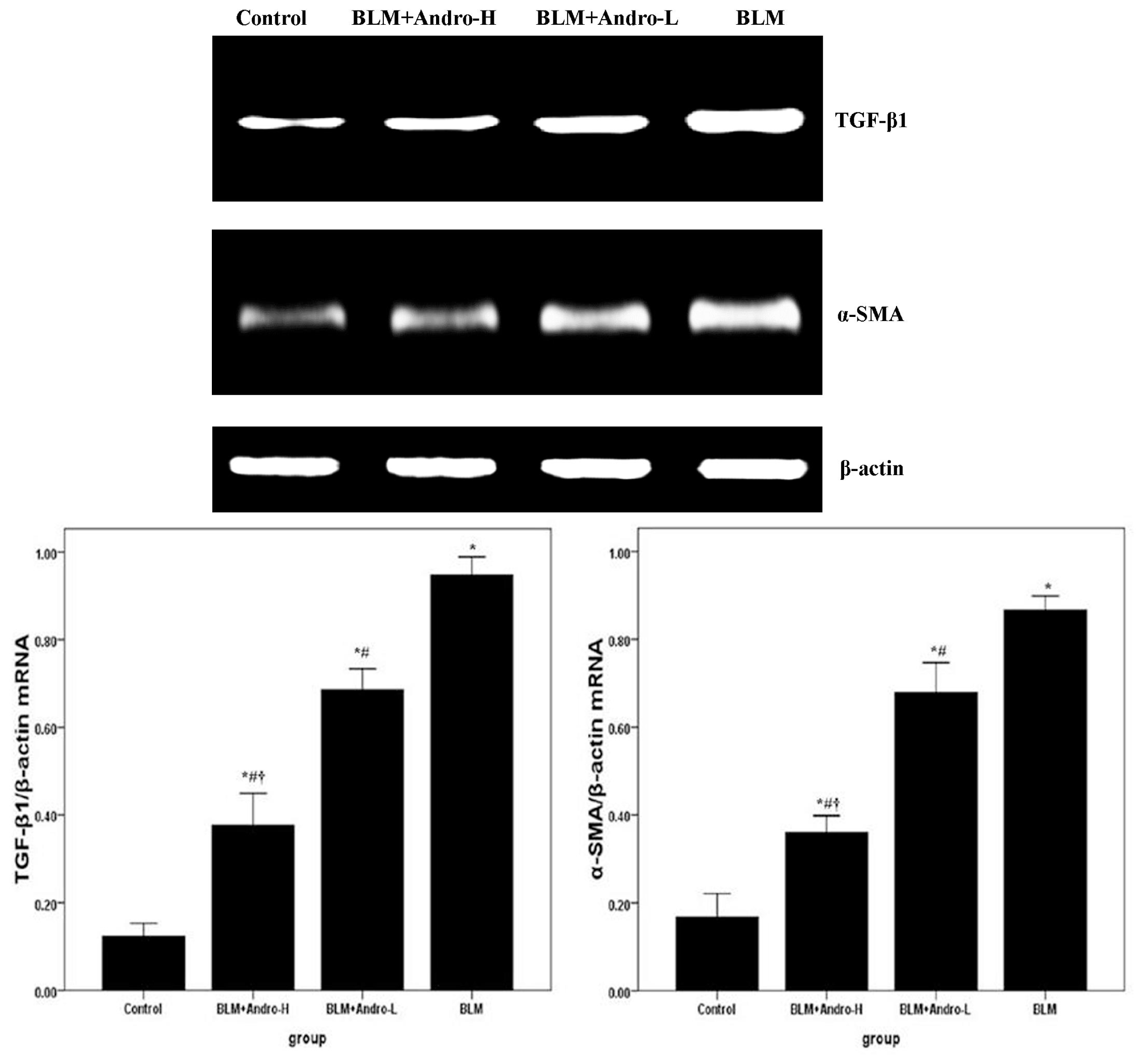
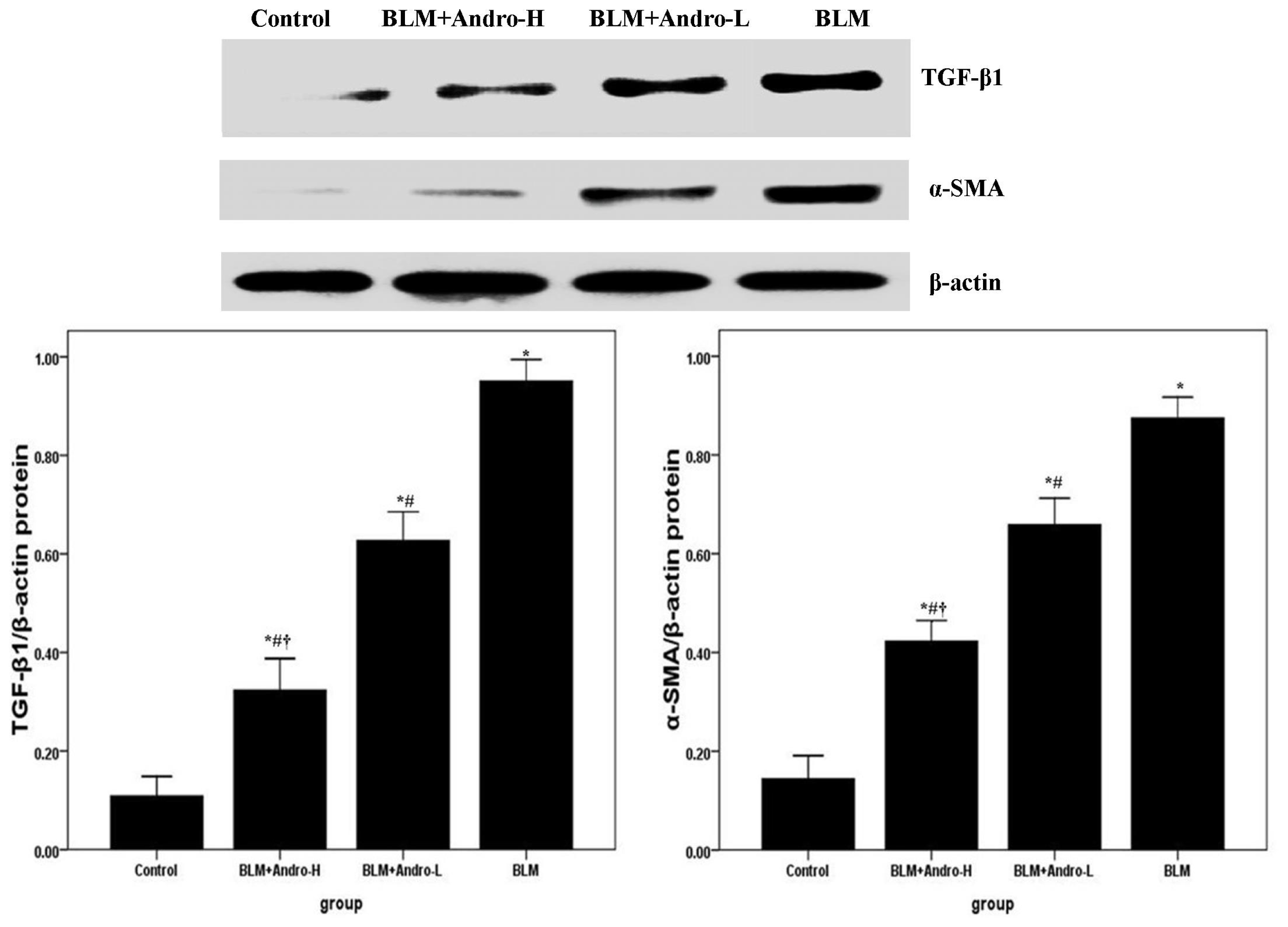
2.3. Andrographolide Inhibits TGF-β1 and α-SMA Expression in BLM-Induced Pulmonary Fibrosis in Mice
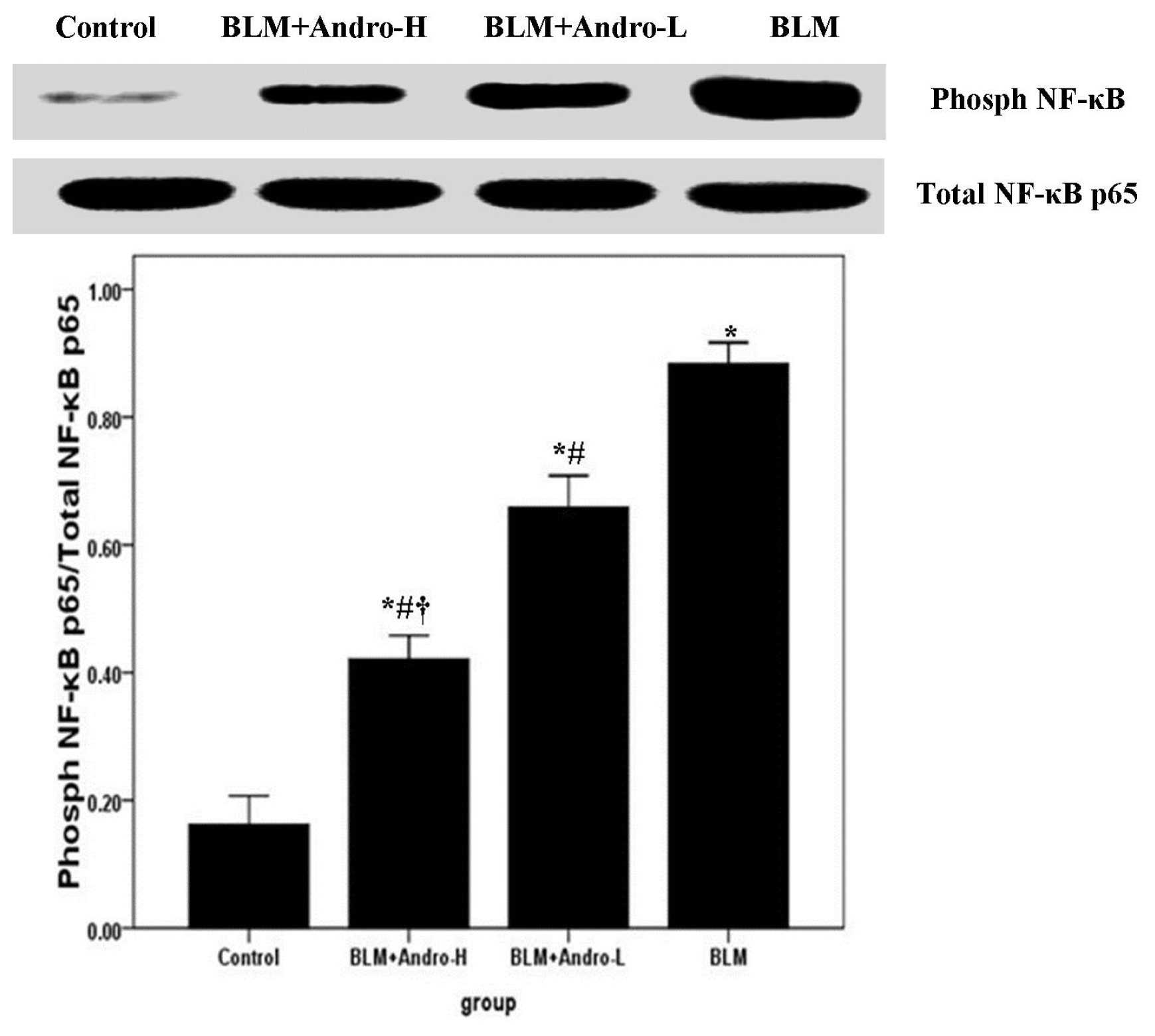
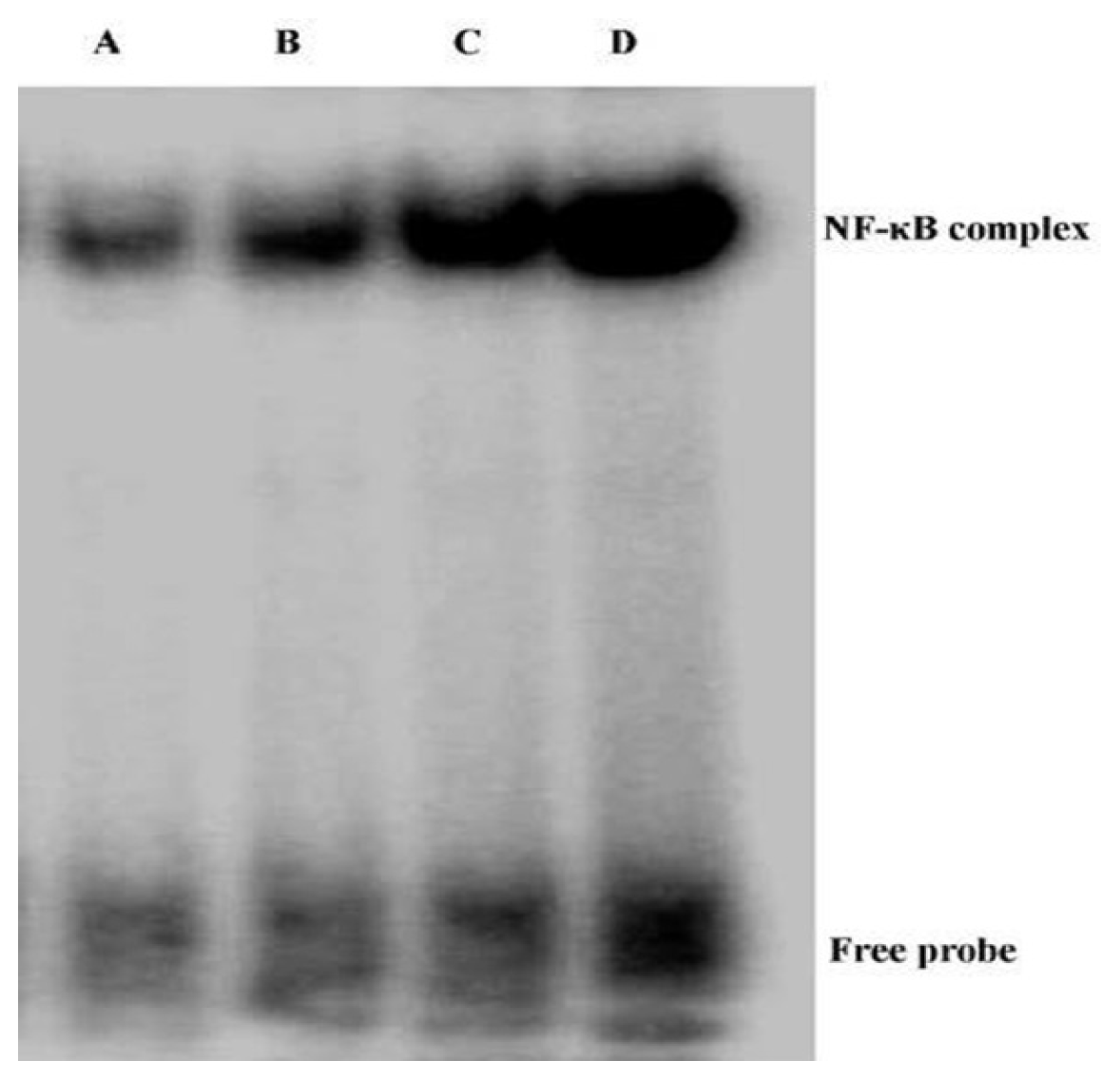
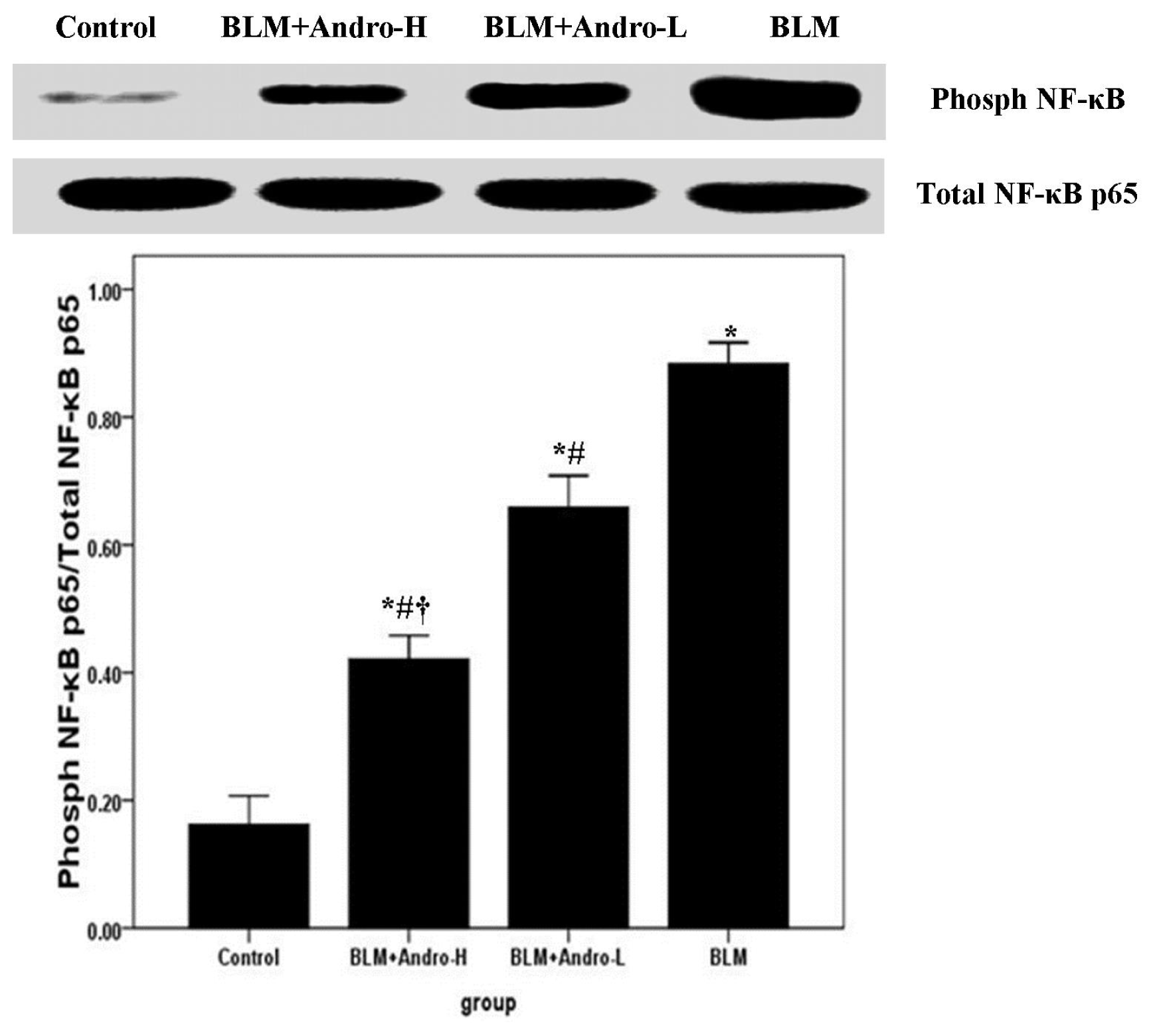
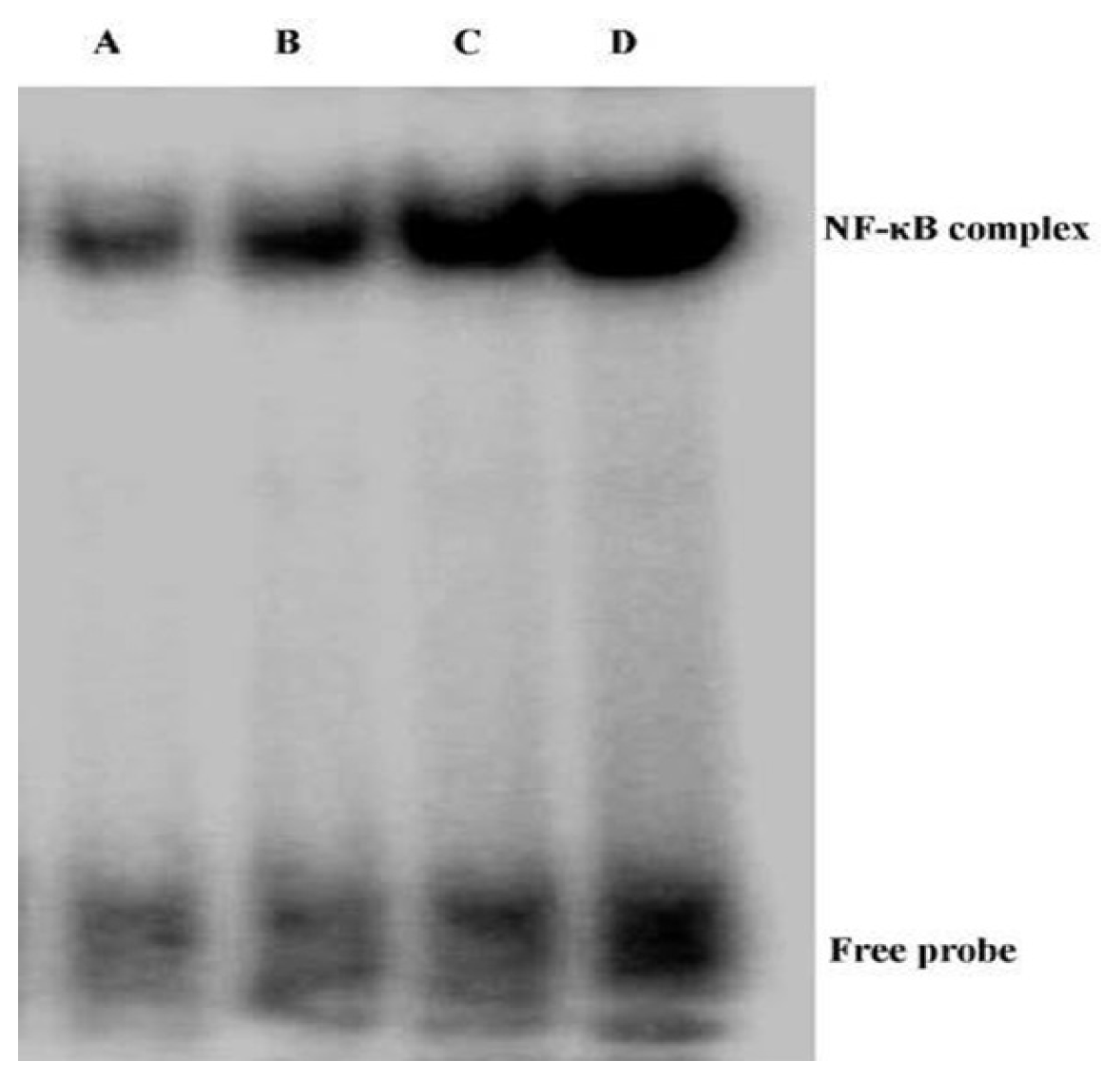
2.4. Andrographolide Inhibits NF-κB p65 Activation and DNA Binding Activity in BLM-Induced Pulmonary Fibrosis in Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Murine Model of BLM-Induced Pulmonary Fibrosis
4.3. Bronchoalveolar Lavage Fluid (BALF) and Cell Counting
4.4. TNF-α in BALF
4.5. Histopathological Examination
4.6. Hydroxyproline Assay
4.7. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.8. Western Blotting
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Statistical Analysis
5. Conclusions
Conflicts of Interest
References
- Tzouvelekis, A.; Paspaliaris, V.; Koliakos, G.; Ntolios, P.; Bouros, E.; Oikonomou, A.; Zissimopoulos, A.; Boussios, N.; Dardzinski, B.; Gritzalis, D.; et al. A prospective, non-randomized, no placebo-controlled, phase Ib clinical trial to study the safety of the adipose derived stromal cells-stromal vascular fraction in idiopathic pulmonary fibrosis. J. Transl. Med 2013, 11, 171. [Google Scholar]
- Collard, H.R.; Yow, E.; Richeldi, L.; Anstrom, K.J.; Glazer, C. IPFnet investigators. Suspected acute exacerbation of idiopathic pulmonary fibrosis as an outcome measure in clinical trials. Respir. Res 2013, 14, 73. [Google Scholar]
- Woodcock, H.V.; Molyneaux, P.L.; Maher, T.M. Reducing lung function decline in patients with idiopathic pulmonary fibrosis: Potential of nintedanib. Drug Des. Dev. Ther 2013, 7, 503–510. [Google Scholar]
- Collard, H.R.; Ryu, J.H.; Douglas, W.W.; Schwarz, M.I.; Curran-Everett, D.; King, T.E., Jr.; Brown, K.K. Combined corticosteroid and cyclophosphamide therapy does not alter survival in idiopathic pulmonary fibrosis. Chest 2004, 125, 2169–2174. [Google Scholar]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.F.; Flaherty, K.R.; Lasky, J.A.; et al. ATS/ERS/JRS/ALAT Committee on Idiopathic Pulmonary Fibrosis. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med 2011, 183, 788–824. [Google Scholar]
- Ou, X.M.; Feng, Y.L.; Wen, F.Q.; Huang, X.Y.; Xiao, J.; Wang, K.; Wang, T. Simvastatin attenuates bleomycin-induced pulmonary fibrosis in mice. Chin. Med. J. (Engl.) 2008, 121, 1821–1829. [Google Scholar]
- Yang, T.; Chen, M.; Sun, T. Simvastatin attenuates TGF-β1-induced epithelial-mesenchymal transition in human alveolar epithelial cells. Cell. Physiol. Biochem 2013, 31, 863–874. [Google Scholar]
- Chen, Y.L.; Zhang, X.; Bai, J.; Gai, L.; Ye, X.L.; Zhang, L.; Xu, Q.; Zhang, Y.X.; Xu, L.; Li, H.P.; et al. Sorafenib ameliorates bleomycin-induced pulmonary fibrosis: Potential roles in the inhibition of epithelial-mesenchymal transition and fibroblast activation. Cell Death Dis 2013, 4, e665. [Google Scholar]
- Aihara, K.; Handa, T.; Oga, T.; Watanabe, K.; Tanizawa, K.; Ikezoe, K.; Taguchi, Y.; Sato, H.; Chin, K.; Nagai, S.; et al. Clinical relevance of plasma prostaglandin f2α metabolite concentrations in patients with idiopathic pulmonary fibrosis. PLoS One 2013, 8, e66017. [Google Scholar]
- Gabasa, M.; Royo, D.; Molina-Molina, M.; Roca-Ferrer, J.; Pujols, L.; Picado, C.; Xaubet, A.; Pereda, J. Lung myofibroblasts are characterized by down-regulated cyclooxygenase-2 and its main metabolite, prostaglandin E2. PLoS One 2013, 8, e65445. [Google Scholar]
- Zhu, T.; Wang, D.X.; Zhang, W.; Liao, X.Q.; Guan, X.; Bo, H.; Sun, J.Y.; Huang, N.W.; He, J.; Zhang, Y.K.; et al. Andrographolide protects against LPS-induced acute lung injury by inactivation of NF-κB. PLoS One 2013, 8, e56407. [Google Scholar]
- Fraternale, A.; Crinelli, R.; Casabianca, A.; Paoletti, M.F.; Orlandi, C.; Carloni, E.; Smietana, M.; Palamara, A.T.; Magnani, M. Molecules altering the intracellular thiol content modulate NF-κB and STAT-1/IRF-1 signalling pathways and IL-12 p40 and IL-27 p28 production in murine macrophages. PLoS One 2013, 8, e57866. [Google Scholar]
- Chitra, P.; Saiprasad, G.; Manikandan, R.; Sudhandiran, G. Berberine attenuates bleomycin induced pulmonary toxicity and fibrosis via suppressing NF-κB dependant TGF-β activation: A biphasic experimental study. Toxicol. Lett 2013, 219, 178–193. [Google Scholar]
- Deng, X.; Xu, M.; Yuan, C.; Yin, L.; Chen, X.; Zhou, X.; Li, G.; Fu, Y.; Feghali-Bostwick, C.A.; Pang, L. Transcriptional regulation of increased CCL2 expression in pulmonary fibrosis involves nuclear factor-κB and activator protein-1. Int. J. Biochem. Cell Biol 2013, 45, 1366–1376. [Google Scholar]
- Jiang, C.; Huang, H.; Liu, J.; Wang, Y.; Lu, Z.; Xu, Z. Fasudil, a rho-kinase inhibitor, attenuates bleomycin-induced pulmonary fibrosis in mice. Int. J. Mol. Sci 2012, 13, 8293–8307. [Google Scholar]
- Chetty, A.; Cao, G.J.; Nielsen, H.C. Insulin-like growth factor-I signaling mechanisms, type I collagen and alpha smooth muscle actin in human fetal lung fibroblasts. Pediatr. Res 2006, 60, 389–394. [Google Scholar]
- Lin, X.; Sime, P.J.; Xu, H.; Williams, M.A.; LaRussa, L.; Georas, S.N.; Guo, J. Yin yang 1 is a novel regulator of pulmonary fibrosis. Am. J. Respir. Crit. Care Med 2011, 183, 1689–1697. [Google Scholar]
- Krug, L.T.; Torres-González, E.; Qin, Q.; Sorescu, D.; Rojas, M.; Stecenko, A.; Speck, S.H.; Mora, A.L. Inhibition of NF-κB signaling reduces virus load and gamma herpesvirus-induced pulmonary fibrosis. Am. J. Pathol 2010, 177, 608–621. [Google Scholar]
- Abu-Ghefreh, A.A.; Canatan, H.; Ezeamuzie, C.I. In vitro and in vivo anti-inflammatory effects of andrographolide. Int. Immunopharmacol 2009, 9, 313–318. [Google Scholar]
- Bao, Z.; Guan, S.; Cheng, C.; Wu, S.; Wong, S.H.; Kemeny, D.M.; Leung, B.P.; Wong, W.S. A novel antiinflammatory role for andrographolide in asthma via inhibition of the nuclear factor-kappaB pathway. Am. J. Respir. Crit. Care Med 2009, 179, 657–665. [Google Scholar]
- Li, B.; Chen, M.; Liu, X.; Guo, S.W. Constitutive and tumor necrosis factor-α-induced activation of nuclear factor-κB in adenomyosis and its inhibition by andrographolide. Fertil. Steril 2013, 100, 568–577. [Google Scholar]
- Burgos, R.A.; Hancke, J.L.; Bertoglio, J.C.; Aguirre, V.; Arriagada, S.; Calvo, M.; Cáceres, D.D. Efficacy of an Andrographis paniculata composition for the relief of rheumatoid arthritis symptoms: A prospective randomized placebo-controlled trial. Clin. Rheumatol 2009, 28, 931–946. [Google Scholar]
- Todd, N.W.; Scheraga, R.G.; Galvin, J.R.; Iacono, A.T.; Britt, E.J.; Luzina, I.G.; Burke, A.P.; Atamas, S.P. Lymphocyte aggregates persist and accumulate in the lungs of patients with idiopathic pulmonary fibrosis. J. Inflamm. Res 2013, 6, 63–70. [Google Scholar]
- Bargagli, E.; Prasse, A.; Olivieri, C.; Muller-Quernheim, J.; Rottoli, P. Macrophage-derived biomarkers of idiopathic pulmonary fibrosis. Pulm. Med 2011, 2011, 717130. [Google Scholar]
- Subbian, S.; Tsenova, L.; O’Brien, P.; Yang, G.; Koo, M.S.; Peixoto, B.; Fallows, D.; Zeldis, J.B.; Muller, G.; Kaplan, G. Phosphodiesterase-4 inhibition combined with isoniazid treatment of rabbits with pulmonary tuberculosis reduces macrophage activation and lung pathology. Am. J. Pathol 2011, 179, 289–301. [Google Scholar]
- Zhu, T.; Guan, X.; Zhang, W.; Wang, D. Dehydroandrographolide succinate inhibits oxidative stress in mice with lipopolysaccharide-induced acute lung injury by inactivating iNOS. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 1238–1241. [Google Scholar]
- Iruretagoyena, M.I.; Tobar, J.A.; González, P.A.; Sepúlveda, S.E.; Figueroa, C.A.; Burgos, R.A.; Hancke, J.L.; Kalergis, A.M. Andrographolide interferes with T cell activation and reduces experimental autoimmune encephalomyelitis in the mouse. J. Pharmacol. Exp. Ther 2005, 312, 366–372. [Google Scholar]
- Wan, Y.Y.; Tian, G.Y.; Guo, H.S.; Kang, Y.M.; Yao, Z.H.; Li, X.L.; Liu, Q.H.; Lin, D.J. Endostatin, an angiogenesis inhibitor, ameliorates bleomycin-induced pulmonary fibrosis in rats. Respir. Res 2013, 14, 56. [Google Scholar]
- Chen, Y.; Nie, Y.C.; Luo, Y.L.; Lin, F.; Zheng, Y.F.; Cheng, G.H.; Wu, H.; Zhang, K.J.; Su, W.W.; Shen, J.G.; et al. Protective effects of naringin against paraquat-induced acute lung injury and pulmonary fibrosis in mice. Food Chem. Toxicol 2013, 58, 133–140. [Google Scholar]
- Cu, A.; Ye, Q.; Sarria, R.; Nakamura, S.; Guzman, J.; Costabel, U. N-acetylcysteine inhibits TNF-alpha, sTNFR, and TGF-beta1 release by alveolar macrophages in idiopathic pulmonary fibrosis in vitro. Sarcoidosis Vasc. Diffus. Lung Dis. 2009, 26, 147–154. [Google Scholar]
- Oikonomou, N.; Harokopos, V.; Zalevsky, J.; Valavanis, C.; Kotanidou, A.; Szymkowski, D.E.; Kollias, G.; Aidinis, V. Soluble TNF mediates the transition from pulmonary inflammation to fibrosis. PLoS One 2006, 1, e108. [Google Scholar]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforming growth factor-beta1: Potential role in idiopathic pulmonary fibrosis. Am. J. Pathol 2005, 166, 1321–1332. [Google Scholar]
- Imanifooladi, A.A.; Yazdani, S.; Nourani, M.R. The role of nuclear factor-kappaB in inflammatory lung disease. Inflamm. Allergy Drug Targets 2010, 9, 197–205. [Google Scholar]
- Chen, X.H.; Sun, R.S.; Hu, J.M.; Mo, Z.Y.; Yang, Z.F.; Jin, G.Y.; Guan, W.D.; Zhong, N.S. Inhibitory effect of emodin on bleomycin-induced pulmonary fibrosis in mice. Clin. Exp. Pharmacol. Physiol 2009, 36, 146–153. [Google Scholar]
- Zhu, T.; Zhang, W.; Wang, D.X.; Huang, N.W.; Bo, H.; Deng, W.; Deng, J. Rosuvastatin attenuates mucus secretion in a murine model of chronic asthma by inhibiting the gamma-aminobutyric acid type A receptor. Chin. Med. J. (Engl.) 2012, 125, 1457–1464. [Google Scholar]
- Xiao, M.; Zhu, T.; Wang, T.; Wen, F.Q. Hydrogen-rich saline reduces airway remodeling via inactivation of NF-κB in a murine model of asthma. Eur. Rev. Med. Pharmacol. Sci 2013, 17, 1033–1043. [Google Scholar]
- Zhu, T.; Zhang, W.; Wang, D.X. Insulin up-regulates epithelial sodium channel in LPS-induced acute lung injury model in rats by SGK1 activation. Injury 2012, 43, 1277–1283. [Google Scholar]
- Song, Y.A.; Park, Y.L.; Kim, K.Y.; Chung, C.Y.; Lee, G.H.; Cho, D.H.; Ki, H.S.; Park, K.J.; Cho, S.B.; Lee, W.S.; et al. Black tea extract prevents lipopolysaccharide-induced NF-κB signaling and attenuates dextran sulfate sodium-induced experimental colitis. BMC Complement. Altern. Med 2011, 11, 91. [Google Scholar]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhu, T.; Zhang, W.; Xiao, M.; Chen, H.; Jin, H. Protective Role of Andrographolide in Bleomycin-Induced Pulmonary Fibrosis in Mice. Int. J. Mol. Sci. 2013, 14, 23581-23596. https://doi.org/10.3390/ijms141223581
Zhu T, Zhang W, Xiao M, Chen H, Jin H. Protective Role of Andrographolide in Bleomycin-Induced Pulmonary Fibrosis in Mice. International Journal of Molecular Sciences. 2013; 14(12):23581-23596. https://doi.org/10.3390/ijms141223581
Chicago/Turabian StyleZhu, Tao, Wei Zhang, Min Xiao, Hongying Chen, and Hong Jin. 2013. "Protective Role of Andrographolide in Bleomycin-Induced Pulmonary Fibrosis in Mice" International Journal of Molecular Sciences 14, no. 12: 23581-23596. https://doi.org/10.3390/ijms141223581