Genetic Diversity and Conservation of the Prespa Trout in the Balkans

Abstract
:1. Introduction
2. Results
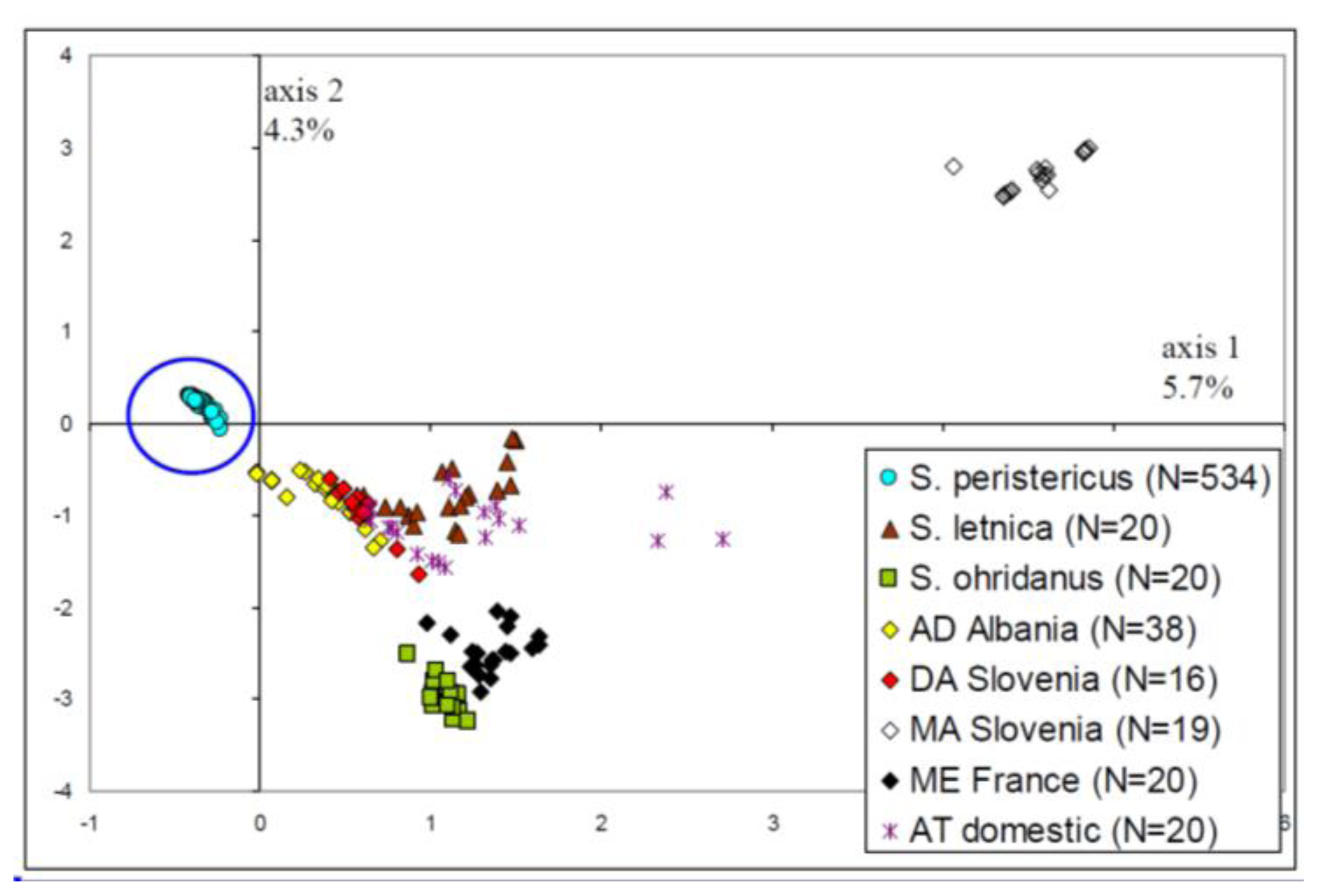
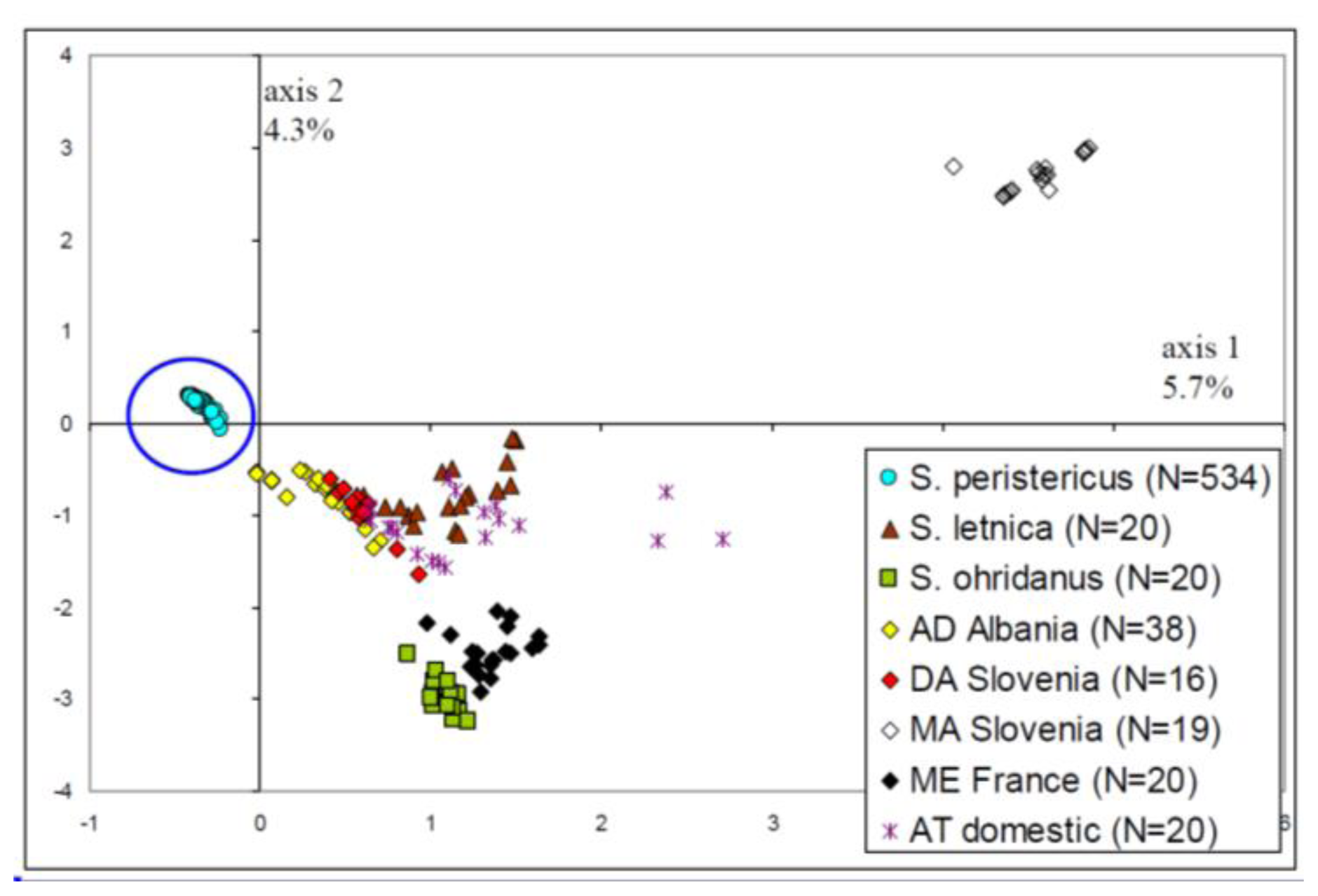
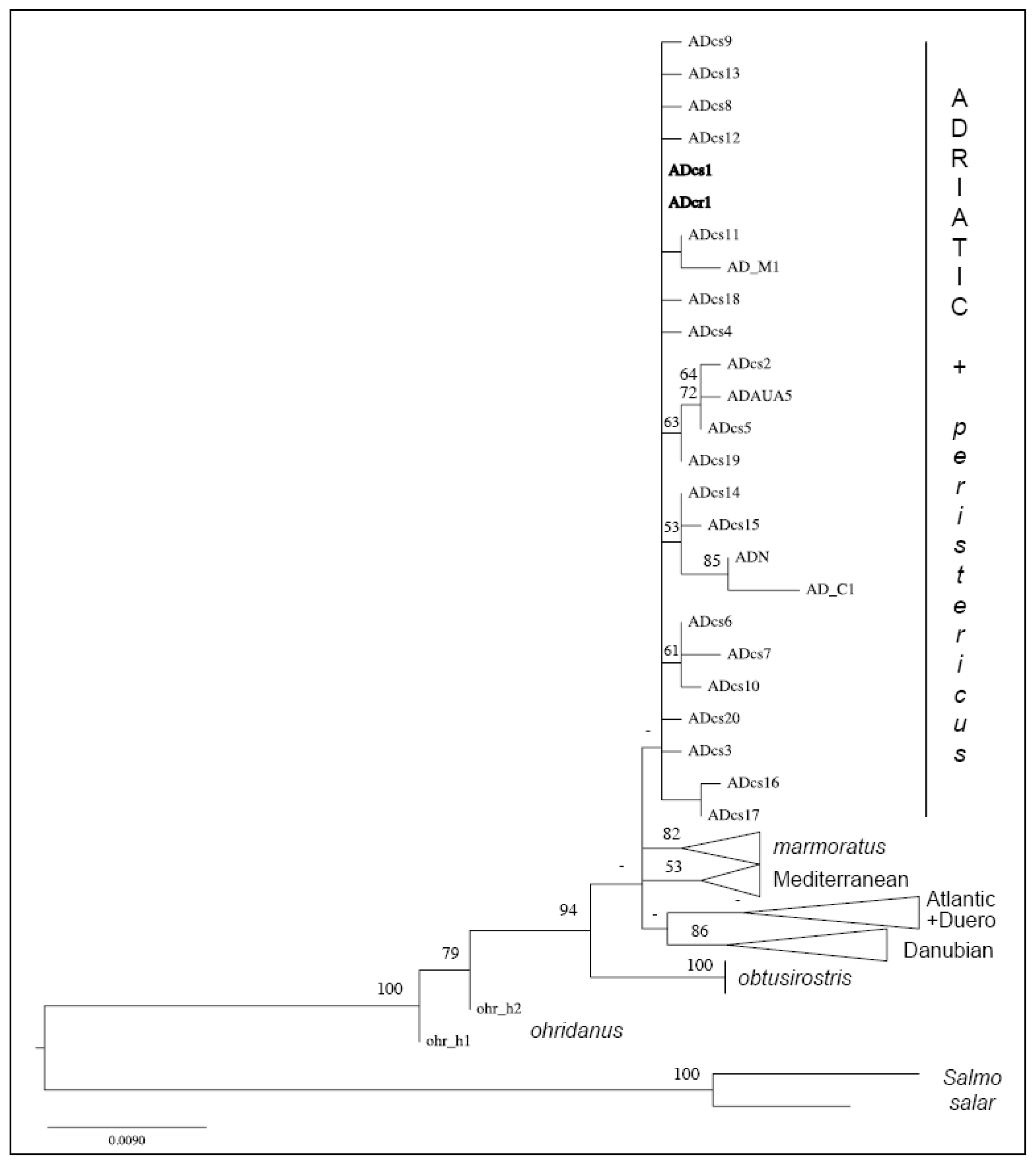
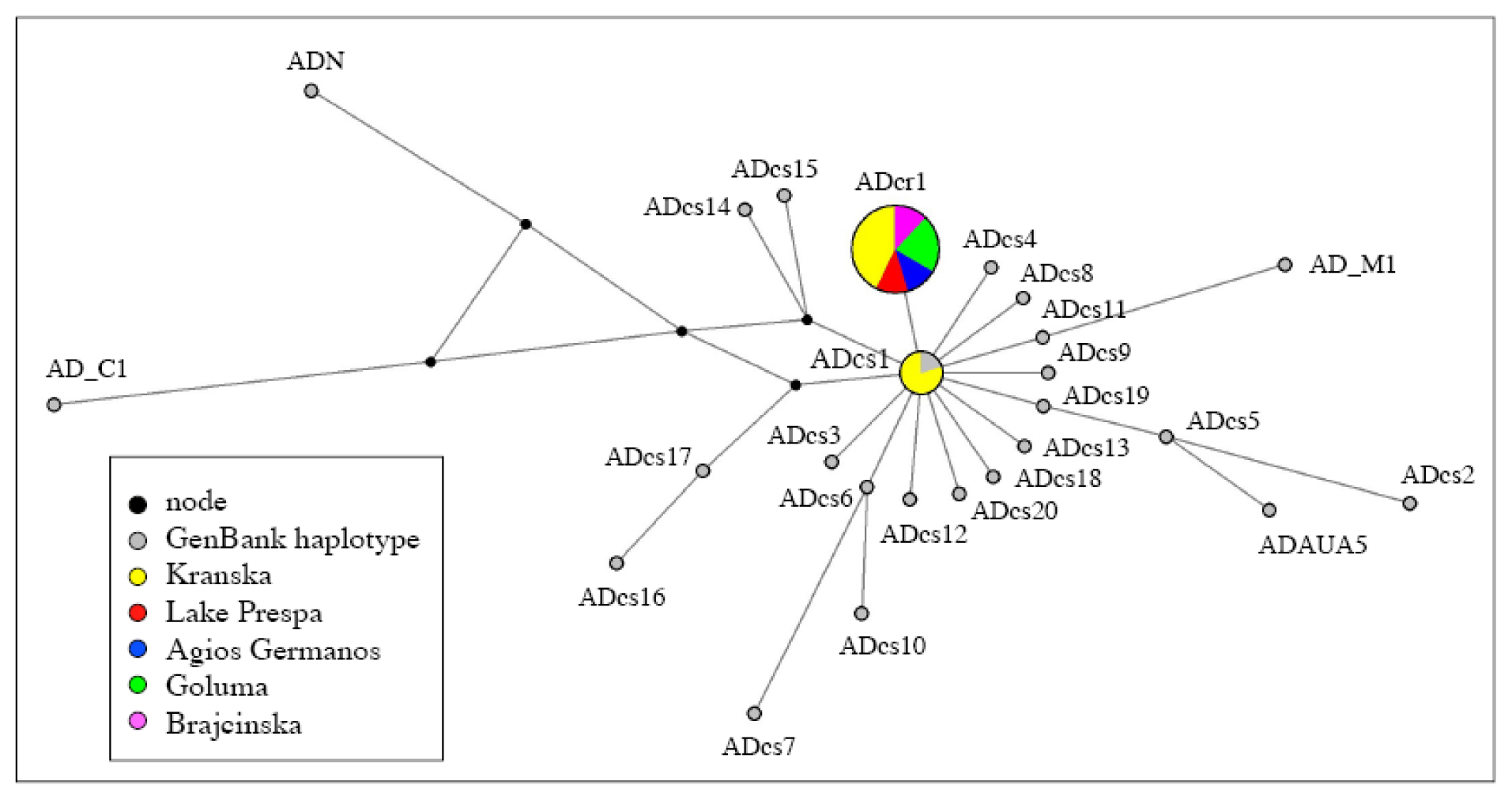
2.1. Distinction of the Prespa Trout Taxon
2.2. Biology of the Populations
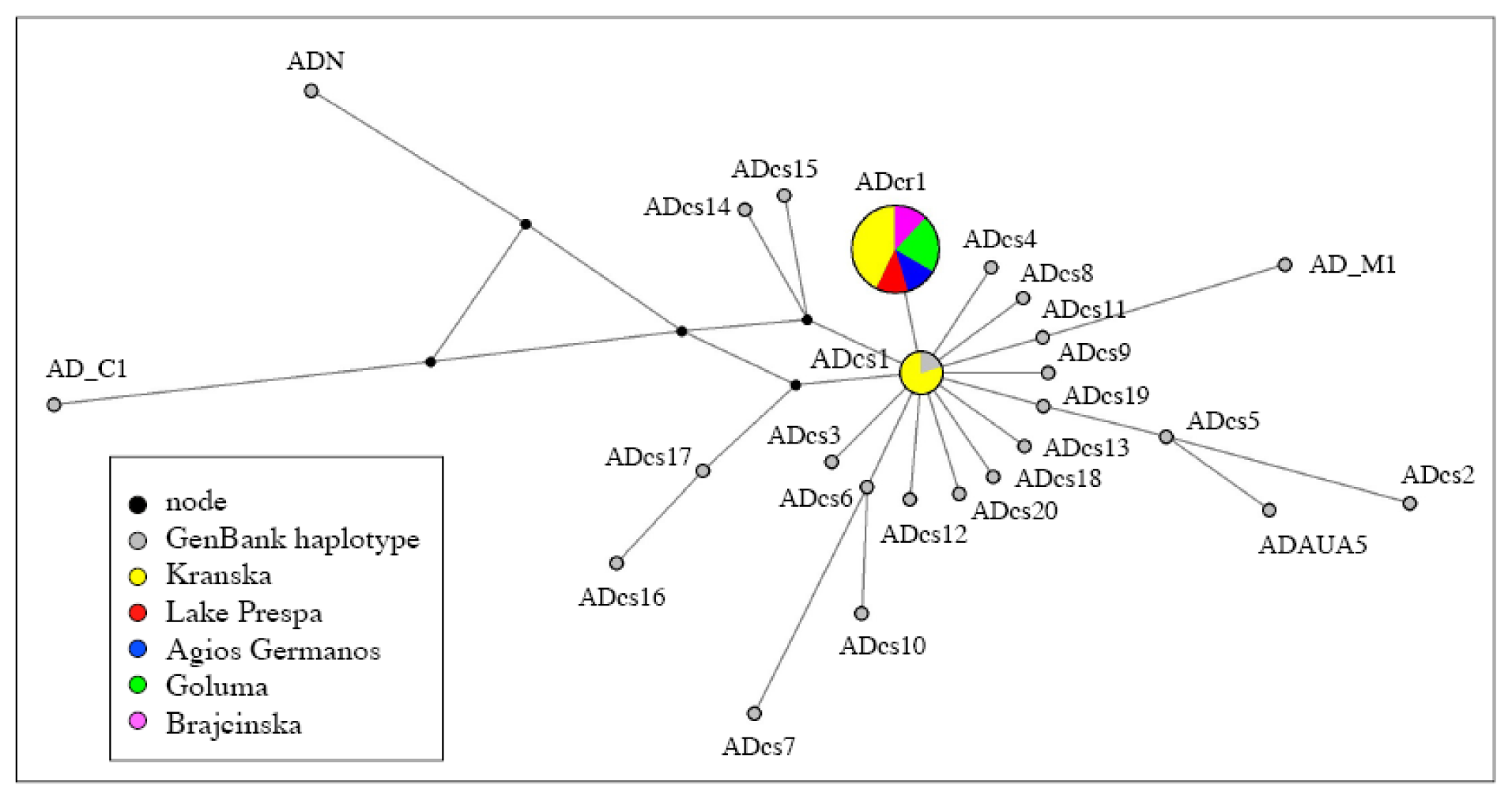
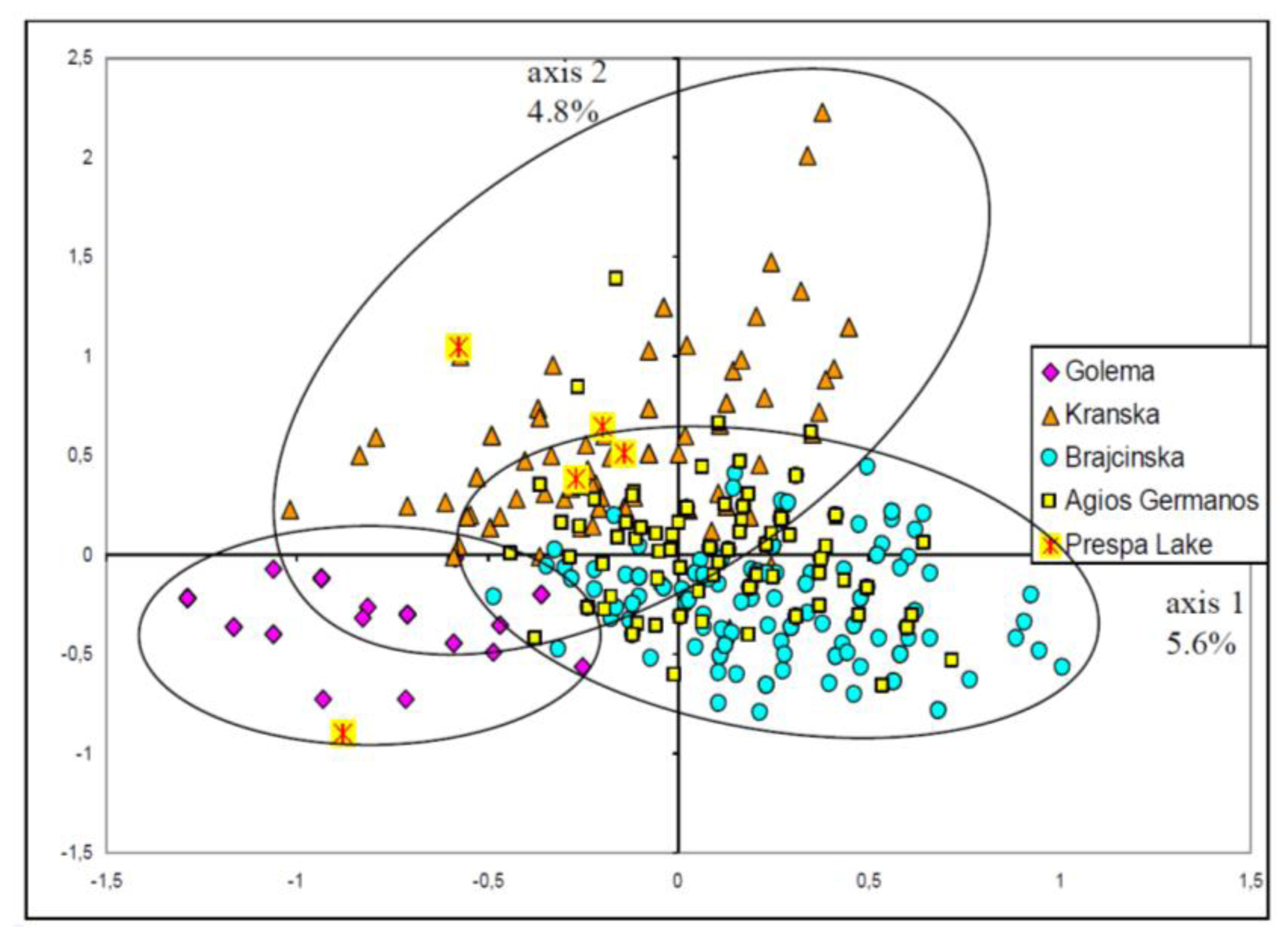
2.3. Population Structure
3. Discussion
3.1. A Structured Endemic Taxon
3.2. A Threatened Species
3.3. Management Considerations
4. Materials and Methods
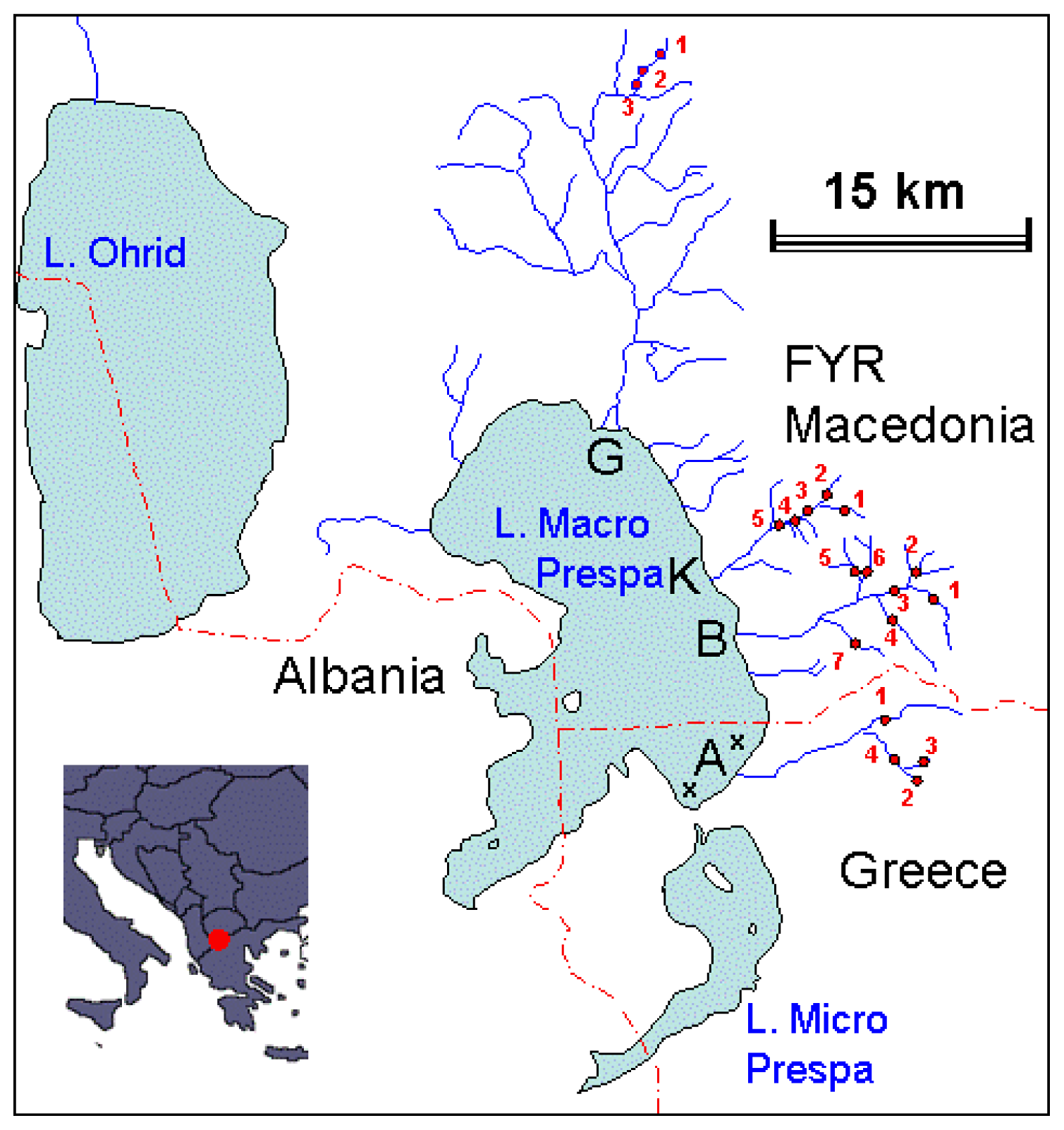
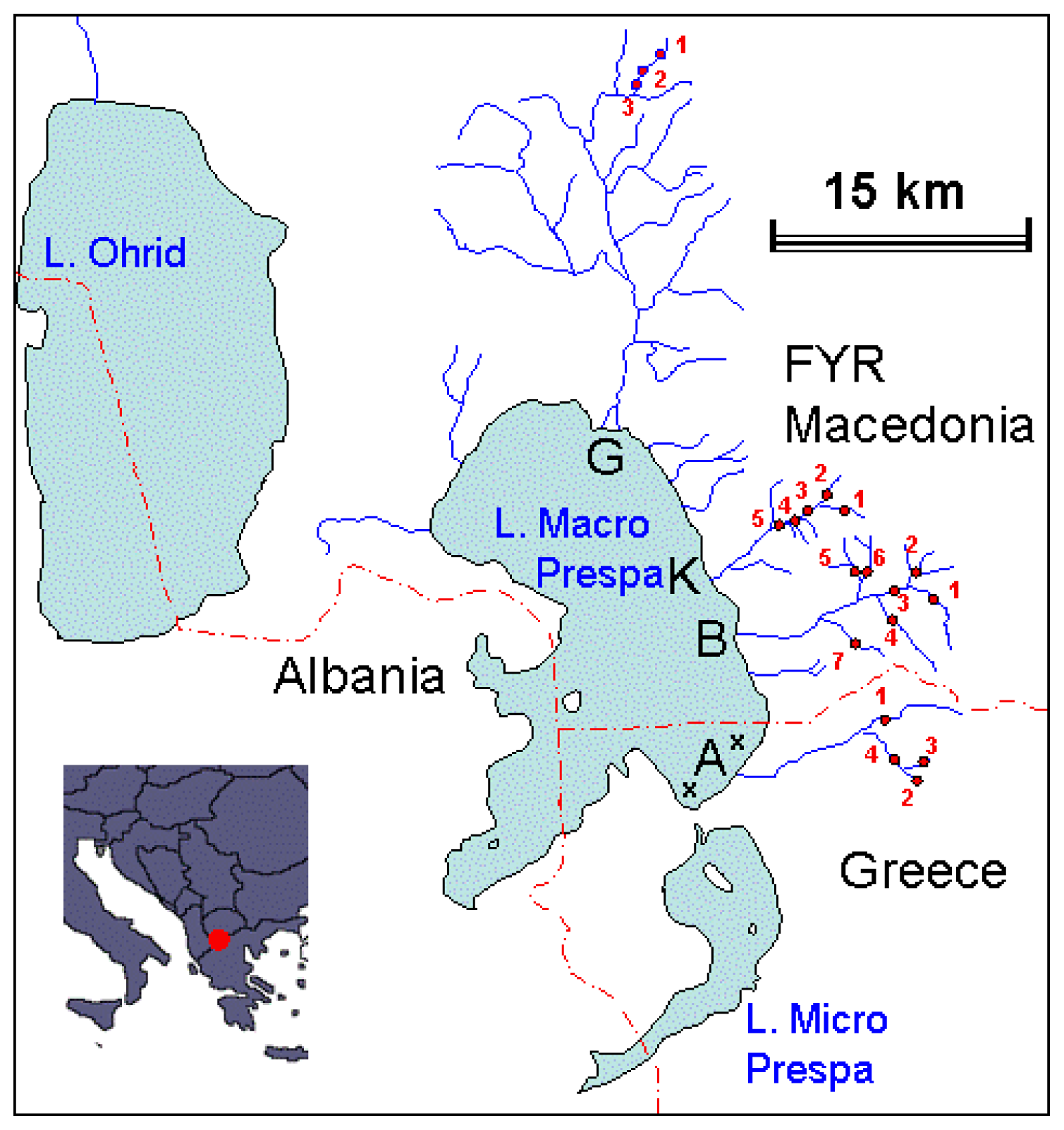
4.1. Sampling Strategy
4.2. DNA Extraction and Genotyping
4.3. Mitochondrial DNA Amplification, Sequencing and Data Analysis
4.4. Microsatellite Descriptive Statistics
- A general picture of the trout diversity was obtained through multidimensional analyses (here Factorial Correspondences Analysis [FCA]). This method was based on the microsatellite data only, without any origin information. It was used to confirm that all samples belonged to the Prespa trout taxon, before analyzing intra-taxon genetic differentiation among rivers.
- A genetic polymorphism estimation (unbiased estimate of average heterozygosity Hnb [42]) was obtained for each sample and for each river or river system.
- The inter-sample or inter-river system differentiations (Fst) and the intra-river panmixia (Fis) were estimated [43]. The significance of the Fis and Fst values was obtained by random permutation procedures (5000 allele permutations within samples for Fis and 5000 individual permutations between samples for Fst).
- Linkage disequilibrium screening was performed using the Black and Krafsur method [44] and the significance was determined by permutation tests (5000 permutations).
- The sequential Bonferroni correction was applied for multiple tests [45].
- Since the same site may have been sampled during the summer of 2005, 2006 and 2007, samples from the same locality collected two years in a row were compared using Fst statistics. The two annual samples were only mixed when the inter-sample Fst could be considered as statistically null.
- Finally, a research of errors due to stuttering, long allele dropout and null allele was performed using Micro-Checker[46].
4.5. Bottleneck Detection
4.6. Microsatellite-Based Assignment Tests
5. Conclusions
Supplementary Information
ijms-14-23454-s001.pdfAcknowledgments
Conflicts of Interest
References
- Crivelli, A.J.; Maitland, P.S. Endemic freshwater of the northern Mediterranean region. Biol. Conserv 1995, 72, 121–337. [Google Scholar]
- Kryštufek, B.; Reed, J.M. Pattern and Process in Balkan Biodiversity—An Overview. In Balkan Biodiversity, Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 203–217. [Google Scholar]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cosson, J.-F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol 1998, 7, 453–464. [Google Scholar]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc 1999, 68, 87–112. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland, 2007. [Google Scholar]
- Karaman, S. Contribution to the knowledge of Yugoslavian freshwater fish. Bull. Soc. Sci. Skopje 1938, 18, 131–139. [Google Scholar]
- Lo Brutto, S.; Hristovski, N.; Arculeo, M. Genetic divergence between morphological forms of brown trout Salmo trutta L. in the Balkan region of Macedonia. J. Fish Biol 2010, 76, 1220–1227. [Google Scholar]
- Snoj, A.; Maric, S.; Berrebi, P.; Crivelli, A.J.; Shumka, S.; Sušnik, S. Genetic architecture of trout from Albania as revealed by mtDNA control region. Genet. Sel. Evol 2009, 41, 22. [Google Scholar]
- Sušnik, S.; Snoj, A.; Wilson, I.; Mrdak, D.; Weiss, S. Historical demography of brown trout (Salmo trutta) in the Adriatic drainage including the putative S. letnica endemic to Lake Ohrid. Mol. Phylogenet. Evol 2007, 44, 63–76. [Google Scholar]
- Sušnik, S.; Knizhin, I.; Snoj, A.; Weiss, S. Genetic and morphological characterization of a Lake Ohrid endemic, Salmo (Acantholingua) ohridanus with a comparison to sympatric. Salmo trutta. J. Fish Biol 2006, 68, 2–23. [Google Scholar]
- Bernatchez, L.; Guyomard, R.; Bonhomme, F. DNA sequence variation of the mitochondrial control region among geographically and morphologically remote European brown trout Salmo trutta populations. Mol. Ecol 1992, 1, 161–173. [Google Scholar]
- Bernatchez, L. The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation. Evolution 2001, 55, 351–379. [Google Scholar]
- Cortey, M.; Pla, C.; Garcia-Marin, J.-L. Historical biogeography of Mediterranean trout. The role of allopatry and dispersal events. Mol. Phylogenet. Evol 2004, 33, 831–844. [Google Scholar]
- Delling, B. Diversity of western and southern Balkan trouts, with the description of a new species from the Louros River, Greece (Teleostei, Salmonidae). Ichthyol. Explor. Freshw 2010, 21, 331–344. [Google Scholar]
- Koutseri, I.; Crivelli, A.J.; Petkovski, S.; Kazoglou, Y. Species Action Plan for the Endemic Prespa Trout, Salmo peristericus: A Conservation Tool. Proceedings of the BALWOIS Water Observation and Information System for Balkan Countries, 25–29 May 2010; BALWOIS: Ohrid, FYR of Macedonia, 2010; pp. 1–18. [Google Scholar]
- Economidis, P.S. Endangered freshwater fishes of Greece. Biol. Conserv 1995, 72, 201–211. [Google Scholar]
- Smith, K.G.; Darwall, W.R.T. The Status and Distribution of Freshwater Fish Endemic to the Mediterranean Basin; IUCN: Gland, Switzerland, Cambridge, UK, 2006; pp. 1–34. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol 2005, 14, 2611–2620. [Google Scholar]
- Presa, P.; Guyomard, R. Conservation of microsatellite in three species of salmonids. J. Fish. Biol 1996, 49, 1326–1329. [Google Scholar]
- Angers, B.; Bernatchez, L.; Angers, A.; Desgroseillers, L. Specific microsatellite loci for brook charr (Salvelinus fontinalis Mitchill) reveal strong population subdivision on a microgeographic scale. J. Fish Biol 1995, 48, 177–185. [Google Scholar]
- McConnell, S.; Hamilton, L.; Morris, D.; Cook, D.; Paquet, D.; Bentzen, P.; Wright, J. Isolation of salmonid microsatellite loci and their application to the population genetics of Canadian east coast stocks of Atlantic salmon. Aquaculture 1995, 137, 19–30. [Google Scholar]
- Scribner, K.T.; Gust, J.R.; Fields, R.L. Isolation and characterization of novel salmon microsatellite loci: Cross-species amplification and population genetic applications. Can. J. Fish. Aquat. Sci 1996, 53, 833–841. [Google Scholar]
- O’Reilly, P.T.; Hamilton, L.C.; McConnell, S.K.; Wright, J.M. Rapid analysis of genetic variation in Atlantic salmon (Salmo salar) by PCR multiplexing of dinucleotide and tetranucleotide microsatellites. Can. J. Fish. Aquat. Sci 1996, 53, 2292–2298. [Google Scholar]
- Rexroad, C.E., III; Coleman, R.L.; Hershberger, W.K.; Killefer, J. Thirty-eight polymorphic microsatellite markers for mapping in rainbow trout. J. Anim. Sci 2002, 80, 541–542. [Google Scholar]
- Bohling, J.H.; Adams, J.R.; Waits, L.P. Evaluating the ability of Bayesian clustering methods to detect hybridization and introgression using an empirical red wolf data set. Mol. Ecol 2013, 22, 74–86. [Google Scholar]
- Karakousis, Y.; Triantaphyllidis, C. Genetic structure and differentiation among Greek brown trout (Salmo trutta L.) populations. Heredity 1990, 64, 297–304. [Google Scholar]
- Apostolidis, A.P.; Stoumboudi, M.T.; Kalogianni, E.; Côté, G.; Bernatchez, L. Genetic divergence among native trout Salmo trutta populations from southern Balkans based on mitochondrial DNA and microsatellite variation. J. Fish Biol 2011, 79, 1950–1960. [Google Scholar]
- Serbezov, D.; Bernatchez, L.; Olsen, E.M.; Vollestad, L.A. Quantitative genetic parameters for wild stream-living brown trout: Heritability and parental effects. J. Evol. Biol 2010, 23, 1631–1641. [Google Scholar]
- Waples, R.S. Pacific Salmon, Oncorhynchus spp., and the definition of “species” under the endangered species act. Mar. Fish. Rev 1991, 53, 11–22. [Google Scholar]
- Palsboll, P.J.; Bérubé, M.; Allendorf, F.W. Identification of management units using population genetic data. Trends Ecol. Evol 2006, 22, 11–16. [Google Scholar]
- Crivelli, A. Tour du Valat; Unpublished Observations; 2013. [Google Scholar]
- Walsh, P.S.; Metzger, D.A.; Higushi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar]
- Estoup, A.; Largiader, C.R.; Perrot, E.; Chourrout, D. Rapid one-tube DNA extraction for reliable PCR detection of fish polymorphic markers and transgenes. Mol. Mar. Biol. Biotechnol 1996, 5, 295–298. [Google Scholar]
- Cortey, M.; García-Marín, J.-L. Evidence for phylogeographically informative sequence variation in the mitochondrial control region of Atlantic brown trout. J. Fish Biol 2002, 60, 1058–1063. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Ne, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum-likelihood. Syst. Biol 2003, 52, 696–704. [Google Scholar]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol 1985, 22, 160–174. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar]
- Mardulyn, P. Trees and/or networks to display intraspecific DNA sequence variation? Mol. Ecol 2012, 21, 3385–3390. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining network for inferring intraspecific phylogenies. Mol. Biol. Evol 1999, 16, 37–48. [Google Scholar]
- Belkhir, K.; Borsa, P.; Goudet, J.; Bonhomme, F. GENETIX 4.05: Logiciel sous Windows pour la génétique des populations; Laboratoire Génome et Population, CNRS-UPR, Université de Montpellier II: Montpellier, France, 2004. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Black, W.C.; Krafsur, E.S. A FORTRAN program for the calculation and analysis of two-locus linkage disequilibrium coefficients. Theor. Appl. Genet 1985, 70, 491–496. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar]
- Cornuet, J.-M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar]
- Piry, S.G.; Luikart, G.; Cornuet, J.M. Bottleneck, a computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered 1999, 90, 502–503. [Google Scholar]
- Luikart, G.; Sherwin, W.B.; Steele, B.M.; Allendorf, F.W. Usefulness of molecular markers for detecting population bottlenecks via monitoring genetic change. Mol. Ecol 1998, 7, 963–974. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resour 2012, 4, 359–361. [Google Scholar]
- Pritchard, J.K.; Wen, W.; Falush, D. Documentation for Structure Software: Version 2.2, 2007. Available online: http://pritch.bsd.uchicago.edu/software (accessed on 24 October 2012).
- Gilbert, K.J.; Andrew, R.L.; Bock, D.G.; Franklin, M.T.; Kane, N.C.; Moore, J.-S.; Moyers, B.T.; Renaut, S.; Rennison, D.J.; Veen, T.; et al. Recommendations for utilizing and reporting population genetic analyses: The reproducibility of genetic clustering using the program Structure. Mol. Ecol 2012, 21, 4925–4930. [Google Scholar]
- Corander, J.; Marttinen, P.; Sirén, J.; Tang, J. Enhanced Bayesian modeling in BAPS software for learning genetic structures of populations. BMC Bioinforma 2008, 9, 539. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station (map) | River system | Tributary | Date of capture | N nDNA | N mtDNA |
|---|---|---|---|---|---|
| G1b | Golema | Leva Reka | 21 August 2006 | 10 | - |
| G1c | Golema | Leva Reka | 26 July 2007 | 15 | 5 |
| G2b | Golema | Leva Reka | 21 August 2006 | 9 | (1) |
| G2c | Golema | Leva Reka | 26 July 2007 | 4 | - |
| G3b | Golema | Leva Reka | 21 August 2006 | 11 | (4) |
| G3c | Golema | Leva Reka | 26 July 2007 | 10 | - |
| K1c | Kranska | main | 25 July 2007 | 5 | - |
| K2c | Kranska | Srbino | 25 July 2007 | 16 | - |
| K3b | Kranska | main | 21 August 2006 | 10 | (1) |
| K3c | Kranska | main | 25 July 2007 | 16 | 4 |
| K4b | Kranska | main | 21 August 2006 | 10 | - |
| K4c | Kranska | main | 25 July 2007 | 21 | - |
| K5b | Kranska | main | 21 August 2006 | 10 | (1) |
| K5c | Kranska | main | 25 July 2007 | 15 | - |
| B1b | Brajcinska | main | 19 August 2006 | 10 | (1) |
| B1c | Brajcinska | main | 23 July 2007 | 25 | - |
| B2c | Brajcinska | main | 27 July 2007 | 31 | 5 |
| B3c | Brajcinska | Kriva Kobila | 27 July 2007 | 37 | - |
| B4b | Brajcinska | Rzanska | 19 August 2006 | 10 | (1) |
| B4c | Brajcinska | Rzanska | 25 July 2007 | 51 | - |
| B5b | Brajcinska | Drmisar | 19 August 2006 | 10 | (1) |
| B5c | Brajcinska | Drmisar | 30 July 2007 | 28 | - |
| B6c | Brajcinska | Marusica | 31 July 2007 | 16 | - |
| B7b | Brajcinska | Baltanska | 12 August 2006 | 3 | (2) |
| B7c | Brajcinska | Baltanska | 30 July 2007 | 29 | - |
| A1a | Agios Germanos | Siroka | 17 August 2005 | 39 | (5) |
| A2a | Agios Germanos | Gaidouritsa | 19 August 2005 | 10 | (3) |
| A2c | Agios Germanos | Gaidouritsa | 3 August 2007 | 20 | 5 |
| A3c | Agios Germanos | Gaidouritsa | 3 August 2007 | 28 | - |
| A4c | Agios Germanos | Gaidouritsa | 8 August 2007 | 22 | - |
| L2006 | Lake Macro Prespa | lake | 30 October 2006 | 2 | 2 |
| L2009 | Lake Macro Prespa | lake | 21 November 2009 | 1 | 1 |
| L2011 | Lake Macro Prespa | lake | 1 August 2011 | 2 | 2 |
| River systems | N | Hn.b. | Hobs. | P (0.99) | Â | Fis |
|---|---|---|---|---|---|---|
| Golema | 59 | 0.16 | 0.16 | 0.50 | 2.00 | 0.05 NS |
| SD | 0.21 | 0.21 | ||||
| Kranska | 103 | 0.37 | 0.35 | 0.83 | 3.17 | 0.06 NS |
| SD | 0.23 | 0.22 | ||||
| Brajcinska | 250 | 0.32 | 0.23 | 0.83 | 3.17 | 0.29 *** |
| SD | 0.25 | 0.18 | ||||
| Agios Germanos | 119 | 0.34 | 0.32 | 0.67 | 2.50 | 0.01 NS |
| SD | 0.27 | 0.25 | ||||
| Lake trout | 5 | 0.64 | 0.70 | 1.00 | 4.00 | −0.11 NS |
| SD | 0.26 | 0.30 |
| Sample names | G | K | BU | BR | BD |
|---|---|---|---|---|---|
| Golema (G) | 0.92 | - | - | - | - |
| Kranska (K) | 0.05 | 0.78 | 0.13 | - | 0.10 |
| Brajcinska upstream (BU) | - | 0.07 | 0.57 | 0.19 | 0.14 |
| Brajcinska Rzanska (BR) | - | - | 0.09 | 0.79 | 0.06 |
| Brajcinska downstream (BD) | - | - | 0.17 | 0.10 | 0.67 |
| Agios Germanos R. | 0.05 | 0.23 | 0.32 | 0.22 | 0.18 |
| Lake Prespa | 0.11 | 0.75 | - | 0.08 | - |
| Prespa 2006 trout | - | 0.91 | - | - | - |
| Prespa 2006 trout | - | 0.89 | 0.05 | - | - |
| Prespa 2009 trout | 0.48 | 0.12 | 0.06 | 0.29 | 0.06 |
| Prespa 2011 trout | - | 0.90 | - | 0.06 | - |
| Prespa 2011 trout | - | 0.95 | - | - | - |
| Locus | MgCl2 (mM) | Annealing temprature (°C) | Allele size (bp) in Prespa trout | Allele size (bp) in S. trutta | Reference |
|---|---|---|---|---|---|
| Sfo1 | 2 | 59 | 124–138 | 110–170 | [20] |
| SsoSL311 | 2 | 57 | 122–144 | 110–196 | [21] |
| Omm1105 | 2 | 53 | 114–218 | 106–310 | [24] |
| Oneμ9 | 1.5 | 60 | 195–205 | 185–205 | [22] |
| Ssa197 | 2 | 53 | 131–151 | 123–179 | [23] |
| Mst85 | 2 | 52 | 149 | 147–193 | [19] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Berrebi, P.; Tougard, C.; Dubois, S.; Shao, Z.; Koutseri, I.; Petkovski, S.; Crivelli, A.J. Genetic Diversity and Conservation of the Prespa Trout in the Balkans. Int. J. Mol. Sci. 2013, 14, 23454-23470. https://doi.org/10.3390/ijms141223454
Berrebi P, Tougard C, Dubois S, Shao Z, Koutseri I, Petkovski S, Crivelli AJ. Genetic Diversity and Conservation of the Prespa Trout in the Balkans. International Journal of Molecular Sciences. 2013; 14(12):23454-23470. https://doi.org/10.3390/ijms141223454
Chicago/Turabian StyleBerrebi, Patrick, Christelle Tougard, Sophie Dubois, Zhaojun Shao, Irene Koutseri, Svetozar Petkovski, and Alain J. Crivelli. 2013. "Genetic Diversity and Conservation of the Prespa Trout in the Balkans" International Journal of Molecular Sciences 14, no. 12: 23454-23470. https://doi.org/10.3390/ijms141223454