Does the Neuroprotective Role of Anandamide Display Diurnal Variations?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
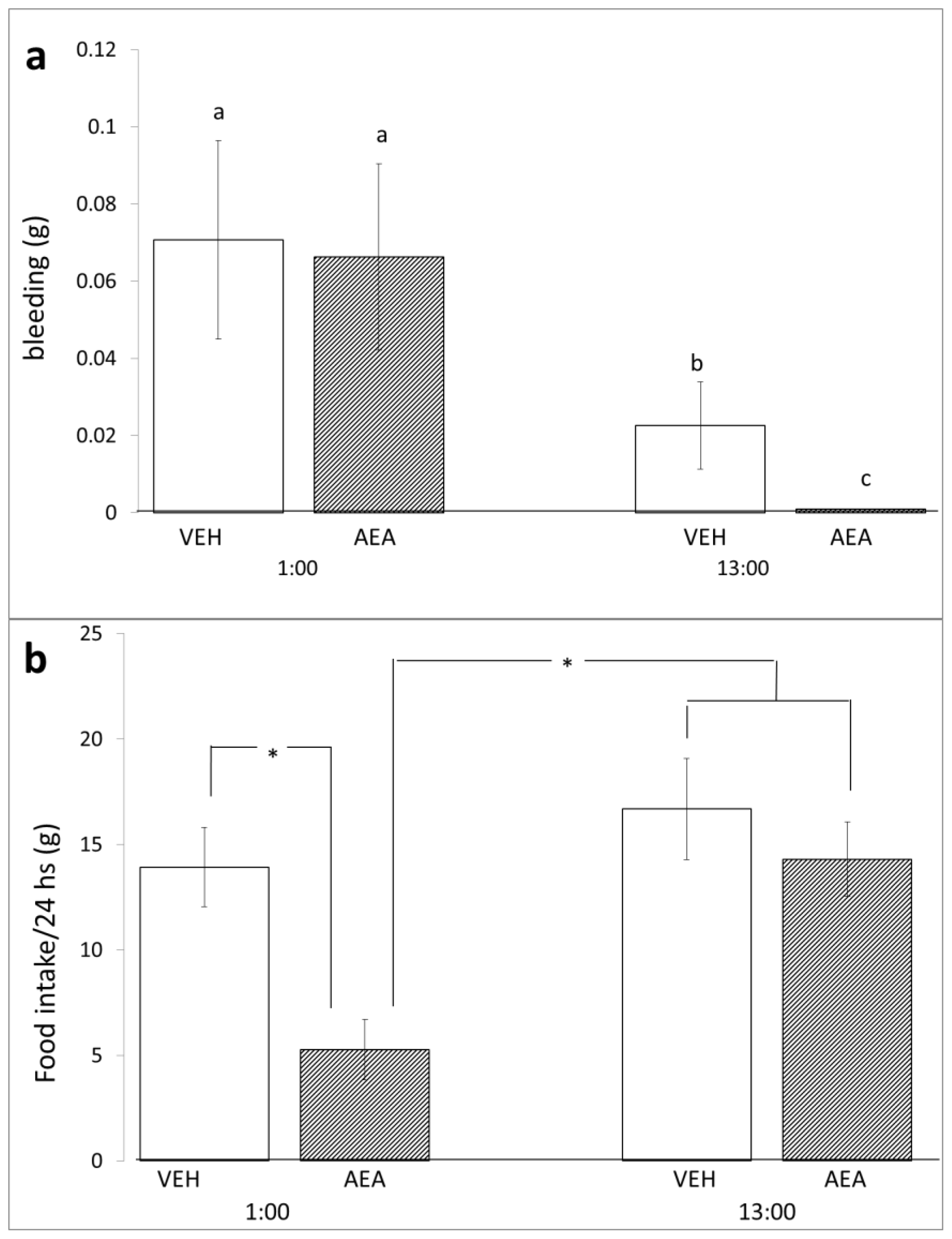
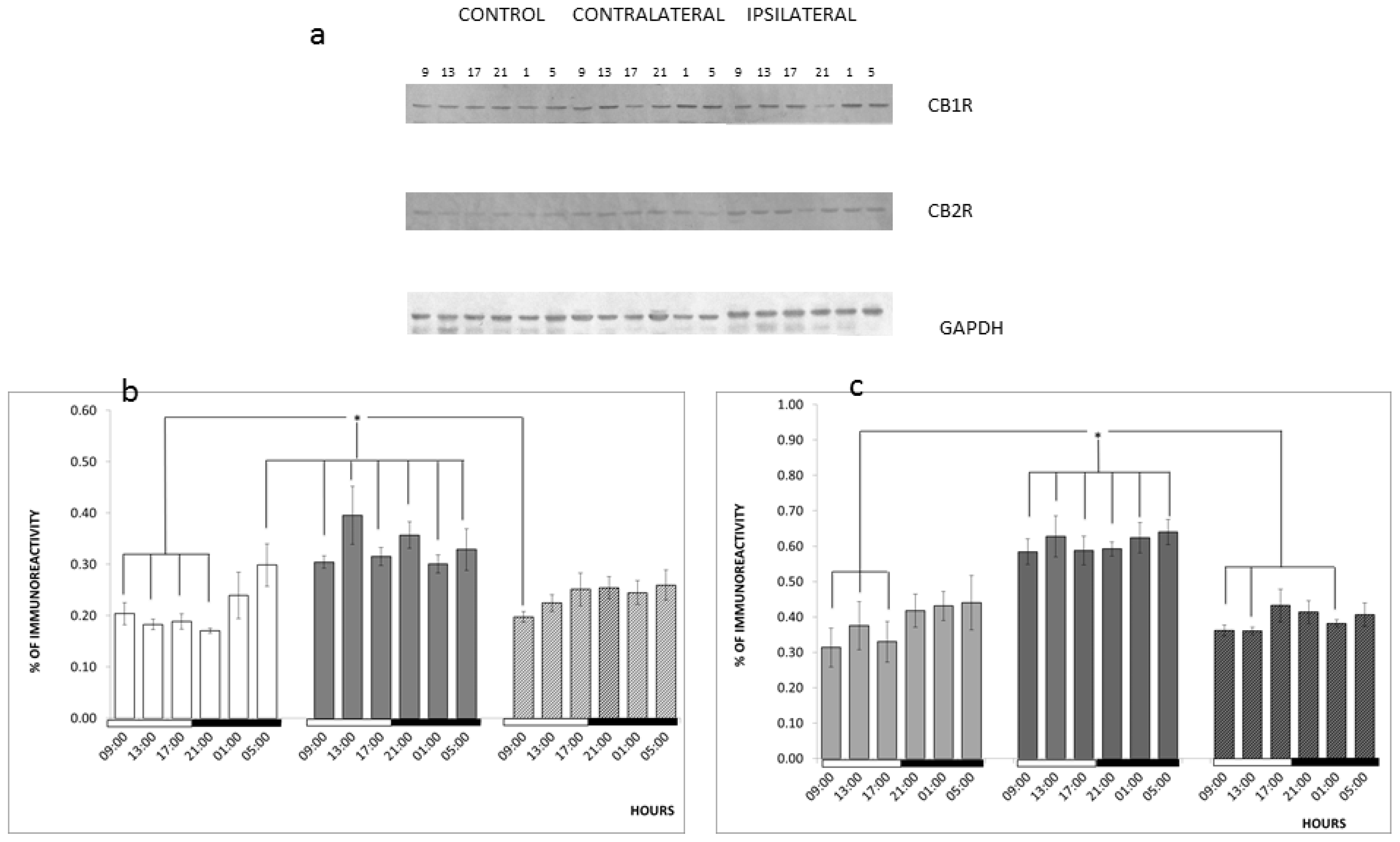
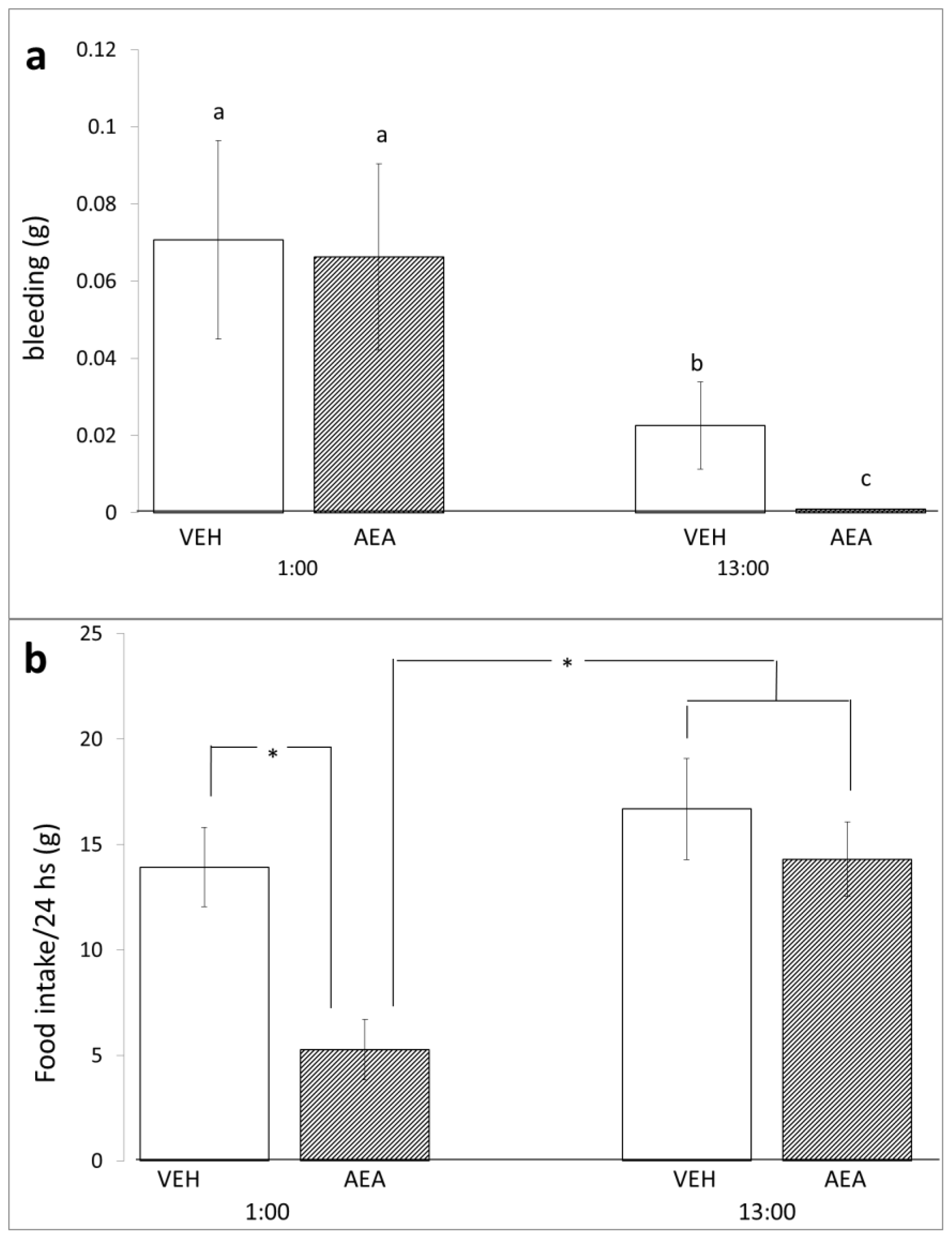
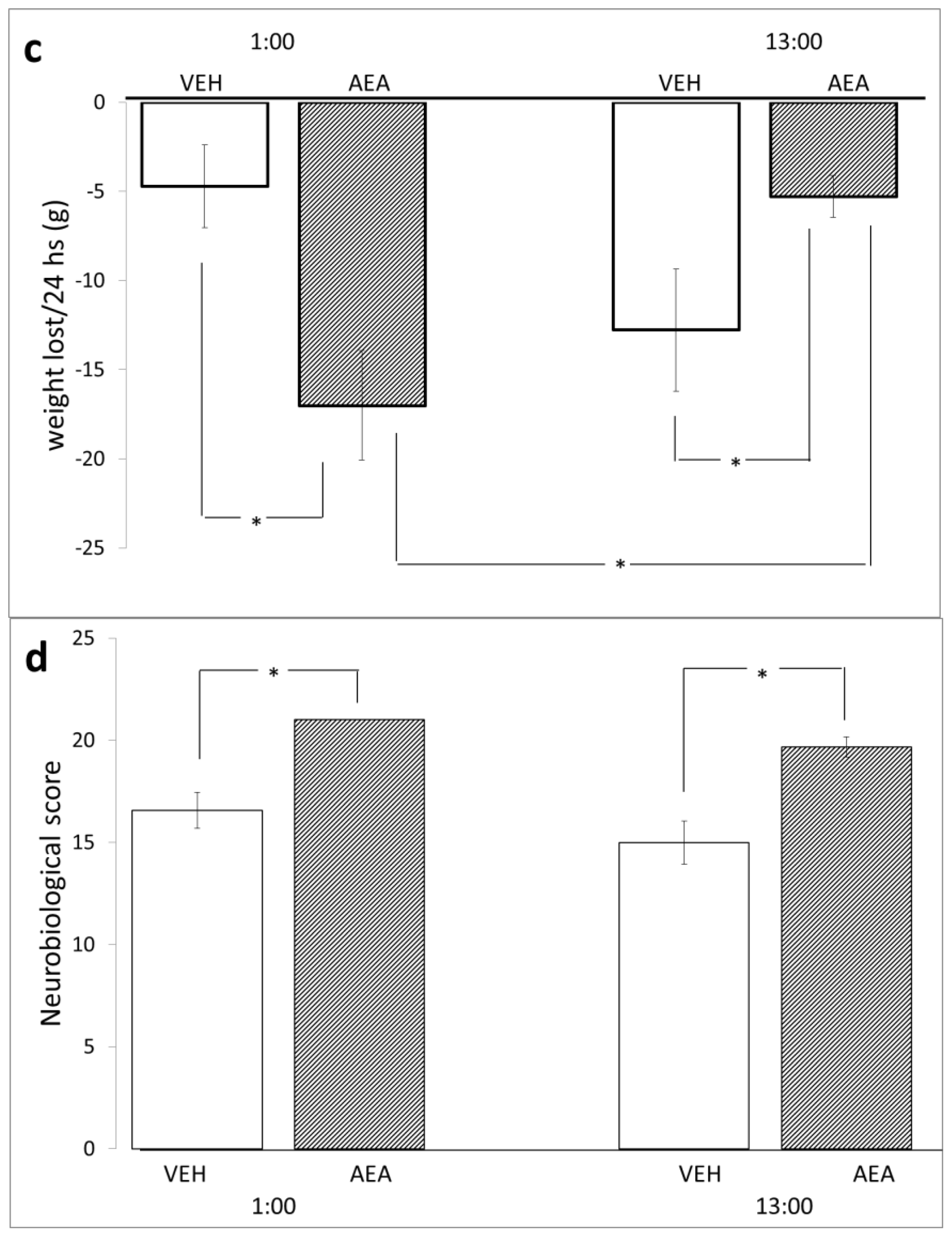
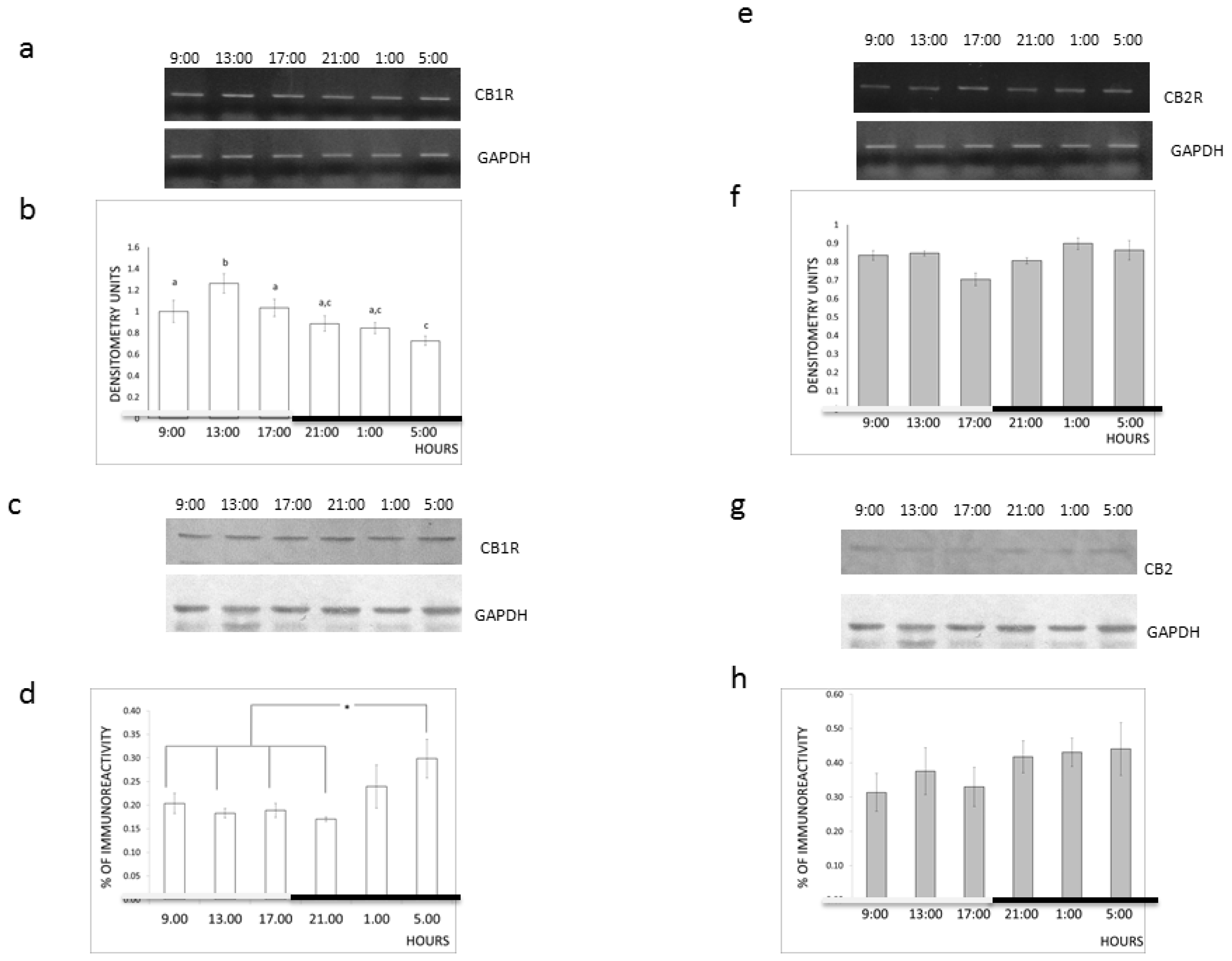
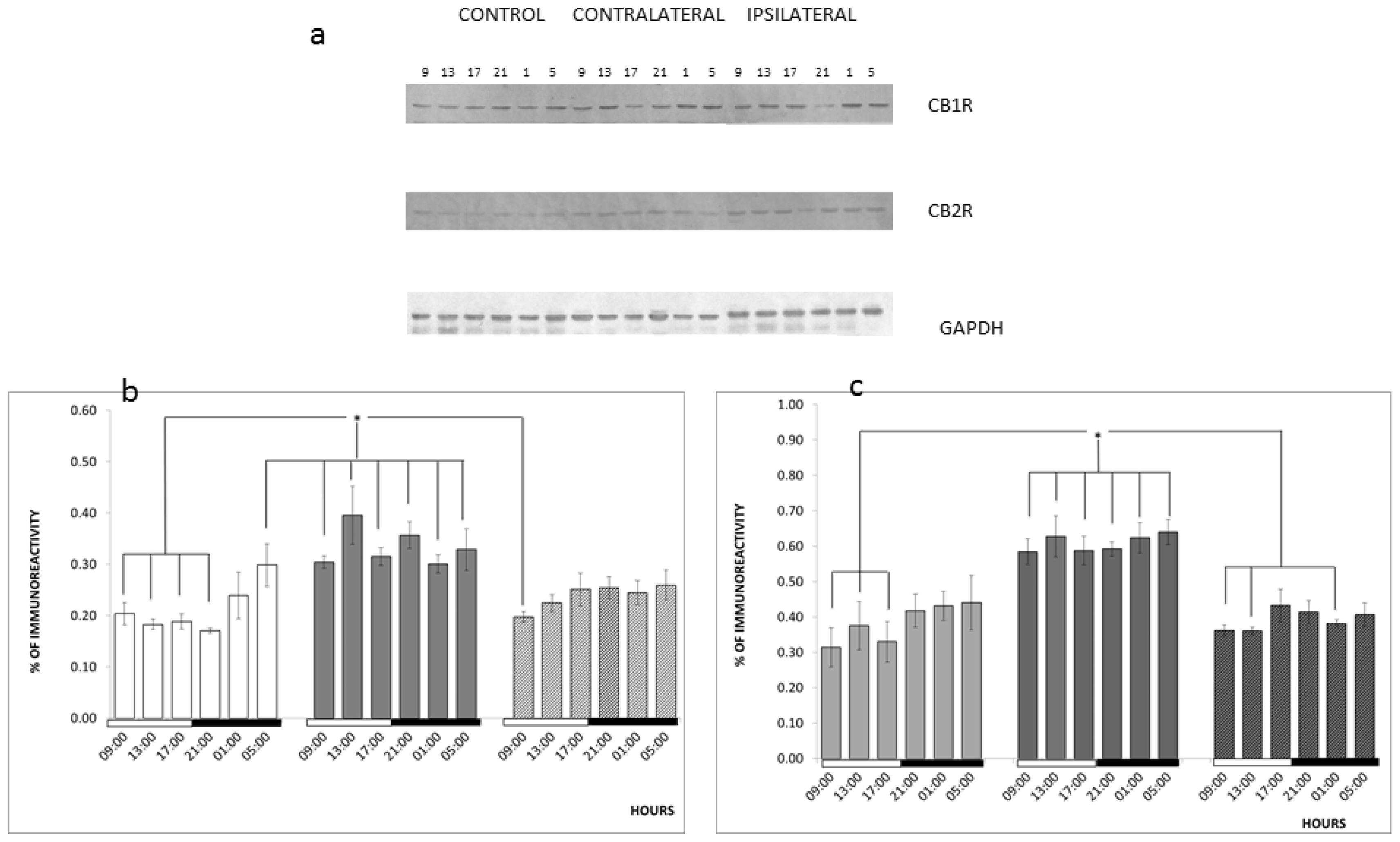
2. Results and Discussion
3. Experimental Section
3.1. Subjects
3.2. Intracerebroventricular Administration of Drugs and TBI
3.3. Bleeding
3.4. Neurological Damage
3.5. Diurnal Variations
3.6. Western Blotting
3.7. RNA Extraction
3.8. Reverse Transcriptase (RT)-PCR
3.9. Statistical Analyses
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mechoulam, R.; Panikashvili, D.; Shohami, E. Cannabinoids and brain injury: Therapeutic implications. Trends Mol. Med 2002, 8, 58–61. [Google Scholar]
- Panikashvili, D.; Mechoulam, R.; Beni, S.M.; Alexandrovich, A.; Shohami, E. CB1 cannabinoid receptors are involved in neuroprotection via NF-kappa B inhibition. J. Cereb. Blood Flow Metab 2005, 25, 477–484. [Google Scholar]
- Panikashvili, D.; Shein, N.A.; Mechoulam, R.; Trembovler, V.; Kohen, R.; Alexandrovich, A.; Shohami, E. The endocannabinoid 2-AG protects the blood-brain barrier after closed head injury and inhibits mRNA expression of proinflammatory cytokines. Neurobiol. Dis 2006, 22, 257–264. [Google Scholar]
- Mechoulam, R.; Shohami, E. Endocannabinoids and traumatic brain injury. Mol. Neurobiol 2007, 36, 68–74. [Google Scholar]
- Leker, R.R.; Shohami, E. Cerebral ischemia and trauma different etiologies yet similar mechanisms: Neuroprotective opportunities. Brain Res. Rev 2002, 39, 55–73. [Google Scholar]
- Turner, W.M. Epidemiology of nocturnal asthma. Am. J. Med 1988, 85, 6–8. [Google Scholar]
- Solomon, G.D. Circadian rhythms and migraine. Cleve Clin. J. Med 1992, 59, 326–329. [Google Scholar]
- Muller, J.E.; Stone, P.H.; Turi, Z.G.; Rutherford, J.D.; Czeisler, C.A.; Parker, C.; Poole, W.K.; Passamani, E.; Roberts, R.; Robertson, T.; et al. Circadian variation in the frequency of onset of acute myocardial infarction. N. Engl. J. Med 1985, 313, 1315–1322. [Google Scholar]
- Marshall, J. Diurnal variation in occurrence of strokes. Stroke 1977, 8, 230–231. [Google Scholar]
- Toni, D.; Argentino, C.; Gentile, M.; Sacchetti, M.L.; Girmenia, F.; Millefiorini, E.; Fieschi, C. Circadian variation in the onset of acute cerebral ischemia: Ethiopathogenetic correlates in 80 patients given angiography. Chronobiol. Int 1991, 8, 321–326. [Google Scholar]
- Vinall, P.E.; Kramer, M.S.; Heinel, L.A.; Rosenwasser, R.H. Temporal changes in sensitivity of rats to cerebral ischemic insult. J. Neurosurg 2000, 93, 82–89. [Google Scholar]
- Martinez-Vargas, M.; Gonzalez-Rivera, R.; Soto-Nuñez, M.; Cisneros-Martinez, M.; Huerta-Saquero, A.; Morales-Gpmez, J.; Molina-Guarneros, J.; Navarro, L. Recovery after a traumatic brain injury depends on diurnal variations. Effect of cystatin C. Neurosci. Lett 2006, 400, 21–24. [Google Scholar]
- Martínez-Vargas, M.; Murillo-Rodriguez, E.; González-Rivera, R.; Landa, A.; Méndez-Diaz, M.; Prospero-Garcia, O.; Navarro, L. Sleep modulates cannabinoid receptor 1 expression in the pons of rats. Neuroscience 2003, 117, 197–201. [Google Scholar]
- Rueda-Orozco, P.E.; Soria-Gomez, E.; Montes-Rodriguez, C.J.; Martínez-Vargas, M.; Galicia, O.; Navarro, L.; Prospero-Garcia, O. A potential function of endocannabinoids in the selection of a navigation strategy by rats. Psychopharmacology 2008, 198, 565–576. [Google Scholar]
- Valenti, M.; Vigano, D.; Cascico, M.G.; Rubino, T.; Steardo, L.; Parolaro, D.; di Marzo, V. Differential diurnal variations of anandamide and 2-arachidonoyl-glycerol levels in rat brain. Cell Mol. Life Sci 2004, 61, 945–950. [Google Scholar]
- Murillo-Rodriguez, E.; Désarnaud, F.; Prospéro-Garcia, O. Diurnal variation of arachidonoylethanolamine, palmitoylethanolamide and oleoylethanolamide in the brain of the rat. Life Sci 2006, 79, 30–37. [Google Scholar]
- Glaser, S.T.; Kaczocha, M. Temporal changes in mouse brain fatty acid amide hydrolase activity. Neuroscience 2009, 163, 594–600. [Google Scholar]
- Wittmann, G.; Deli, L.; Kallo, I.; Hrabovszky, E.; Watanabe, M.; Lipozits, Z.; Fekete, C. Distribution of type I cannabinoid receptor (CB1)-immunoreactive axons in the mouse hypothalamous. J. Comp. Neurol 2007, 503, 270–279. [Google Scholar]
- Acuna, G.C.; Obrietan, K.V. Cannabinoids excite circadian clock neurons. J. Neurosci 2010, 30, 10061–10066. [Google Scholar]
- Sanford, A.E.; Castillo, E.; Gannon, R.L. Cannabinoids and hamster circadian activity rhythms. Brain Res 2008, 1222, 141–148. [Google Scholar]
- Montagnana, M.; Salvagno, G.L.; Lippi, G. Circadian variation within hemostasis: An underrecognized link between biology and disease? Semin. Thromb. Hemost 2009, 35, 23–33. [Google Scholar]
- Soulban, G.; Labrecque, G. Circadian rhythms of blood clotting time and coagulation factors II, VII, IX and X in rats. Life Sci 1989, 45, 2485–2489. [Google Scholar]
- Maccarrone, M.; Bari, M.; Menichelli, A.; del Principe, D.; Agro, A.F. Anandamide activates human platelets through a pathway independent of the arachidonate cascade. FEBS Lett 1999, 447, 277–282. [Google Scholar]
- Braud, S.; Bon, C.; Touqui, L.; Mounier, C. Activation of rabbit blood platelets by anandamide through its cleavage into arachidonic acid. FEBS Lett 2000, 471, 12–16. [Google Scholar]
- Sritharan, K.; Thompson, H. Understanding the metabolic response to trauma. Br. J. Hosp. Med 2009, 70, M156–M518. [Google Scholar]
- Kazanis, I.; Bozas, E.; Philippidis, H.; Stylianopoulou, F. Neuroprotective effects of insulin-like growth factor-I (IGF-I) following a penetrating brain injury in rats. Brain Res 2003, 991, 34–45. [Google Scholar]
- Vickers, S.P.; Kennett, G.A. Cannabinoids and the regulation of ingestive behavior. Curr. Drug Targets 2005, 6, 215–223. [Google Scholar]
- Schmid, P.C.; Krebsbach, R.J.; Perry, S.R.; Dettmer, T.M.; Maasson, J.L.; Schmid, H.H. Occurrence and postmortem generation of anandamide and other long-chain N-acylethanolamines in mammalian brain. FEBS Lett 1995, 375, 143–147. [Google Scholar]
- Shen, M.; Thayer, S.A. Cannabinoid receptor agonists protect cultured rat hippocampal neurons from excitotoxicity. Mol. Pharmacol 1998, 54, 459–462. [Google Scholar]
- Nagayama, T.; Sinor, A.D.; Simon, R.P.; Chen, J.; Graham, S.H.; Jin, K.; Greenberg, D.A. Cannabinoids and neuroprotection in global and focal cerebral ischemia and in neuronal cultures. J. Neuroci 1999, 19, 2987–2995. [Google Scholar]
- Schomacher, M.; Müller, H.D.; Sommer, C.; Schwab, S.; Schabitz, W.R. Endocannabinoids mediate neuroprotection after transient focal cerebral ischemia. Brain Res 2008, 1240, 213–220. [Google Scholar]
- Tominaga, K.; Shinohara, K.; Otori, Y.; Fukuhara, C.; Inouye, S.T. Circadian rhythms of vasopressin content in the suprachiasmatic nucleus of the rat. Neuroreport 1992, 3, 809–812. [Google Scholar]
- Burbach, J.P.; Liu, B.; Voorhuis, T.A.; van Tol, H.H. Diurnal variation in vasopressin and oxytocin messenger RNAs in hypothalamic nuclei of the rat. Brain Res 1988, 464, 157–160. [Google Scholar]
- Bruinink, A.; Lichtensteiger, W.; Schlumpf, M. Ontogeny of diurnal rhythms of central dopamine, serotonin and spirodecanone bindings sites and of motor activity in the rat. Life Sci 1983, 33, 31–38. [Google Scholar]
- Naber, D.; Wirz, J.A.; Kafka, M.S.; Tobler, I.; Borbely, A.A. Seasonal variations in the endogenous rhythm of dopamine receptor binding in rat striatum. Biol. Psychiatry 1981, 16, 831–835. [Google Scholar]
- Torner, C.; Sanchez, H.S.; Aguilar-Roblñero, R. Diurnal variations of striatal D2 dopaminergic receptors and its relation with haloperidol-induced catalepsy. Biol. Rhythm Res 1998, 29, 538–545. [Google Scholar]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 388, 773–778. [Google Scholar]
- Romero, J.; Berrendero, F.; Manzanares, J.; Perez, A.; Cochero, J.; Fuentes, J.A.; Fernandez-Ruiz, J.J.; Ramos, J.A. Time-course of the cannabinoid receptor down-regulation in the adult rat brain caused by repeated exposure to delta-9-tethahydrocannabinol. Synapse 1998, 30, 298–308. [Google Scholar]
- Zhuang, S.Y.; Kittler, J.; Grigorensko, E.V.; Kirby, M.T.; Sim, L.J.; Hapson, R.E.; Childers, S.R.; Deadwyler, S.A. Effects of long-term exposure to delta-9-THC on expression of cannabinoid receptor (CB1) mRNA in different brain regions. Mol. Brain Res 1998, 62, 141–149. [Google Scholar]
- Borner, C.; Hollt, V.; Sebald, W.; Kraus, J. Transcriptional regulation of the cannabinoid receptor type 1 gene in T cells by cannabinoids. J. Leukoc. Biol 2007, 81, 336–343. [Google Scholar]
- White, T.E.; Ford, G.D.; Surles-Zeigler, M.C.; Gates, A.S.; LaPlaca, M.C.; Ford, B.D. Gene expression patterns following unilateral traumatic brain injury reveals a local proinflammatory and remote anti-inflammatory response. Genomics 2013, 14, 282. [Google Scholar]
- Garcia-Ovejero, D.; Arevalo-Martin, A.; Petrosino, S.; Docagne, F.; Hagen, C.; Bisogno, T.; Watanabe, M.; Guaza, C.; di Marzo, V.; Molina-Holgado, E. The endocannabinoid system is modulated in response to spinal cord injury in rats. Neurobiol. Dis 2009, 33, 57–71. [Google Scholar]
- Jin, K.L.; Mao, X.O.; Godsmith, P.C.; Greenberg, D.A. CB1 cannabinoid receptor induction in experimental stroke. Ann. Neurol 2000, 48, 257–261. [Google Scholar]
- Schomacher, M.; Müller, H.D.; Sommer, C. Short-term ischemia usually used for ischemic preconditioning down-regulates central cannabinoid receptors in the gerbil hippocampus. Acta Neuropathol 2006, 111, 8–14. [Google Scholar]
- Zhang, M.; Martin, B.R.; Adler, M.W.; Razdan, R.K.; Ganea, D.; Tuma, R.F. Modulation of the balance between cannabinoid CB(1) and CB(2) receptor activation during cerebral ischemic/reperfusion injury. Neuroscience 2008, 152, 753–760. [Google Scholar]
- Paul, T.; Lemmer, B. Disturbance of circadian rhythms in analgosedated intensive care unit patients with and without craniocerebral injury. Chronobiol. Int 2007, 24, 45–61. [Google Scholar]
- Llompart-Pou, J.; Perez, G.; Raurich, J.M.; Riesco, M.; Brell, M.; Ibañez, J.; Perez-Barcena, J.; Abadal, J.M.; Homar, J.; Burguera, B. Loss of cortisol circadian rhythm in patients with traumatic brain injury: A microdialysis evaluation. Neurocrit. Care 2010, 13, 211–216. [Google Scholar]
- Ayalon, L.; Borodkin, K.; Dishon, L.; Kanety, H.; Dagan, Y. Circadian rhythm sleep disorders following mild traumatic brain injury. Neurology 2007, 68, 1136–1140. [Google Scholar]
- Castriotta, R.J.; Murthy, J.N. Sleep disorders in patients with traumatic brain injury: A review. CNS Drugs 2010, 25, 175–185. [Google Scholar]
- Boone, D.R.; Sell, S.L.; Micci, M.A.; Crookshanks, J.M.; Parsley, M.; Uchida, T.; Prough, D.S.; DeWitt, D.S.; Hellmich, H.L. Traumatic brain injury-induced dysregulation of the circadian clock. PLoS One 2012, 7, e46204. [Google Scholar]
- Perron, R.R.; Tyson, R.L.; Sutherland, G.R. Delta-9-tetrahydrocannabinol increases brain temperature and inverts circadian. Neuroreport 2001, 12, 3791–3794. [Google Scholar]
- Oviedo, A.; Glowa, J.; Herkenham, M. Chronic cannabinoid administration alters cannabinoid receptor binding in rat brain: A quantitative autoradiographic study. Brain Res 1993, 616, 293–302. [Google Scholar]
- Rodríguez de Fonseca, F.; Gorriti, M.A.; Fernández-Ruiz, J.J.; Palomo, T.; Ramos, J.A. Down-regulation of rat brain cannabinoid binding sites after chronic delta 9-tetrahydrocannabinol. Pharmacol. Biochem. Behav 1994, 47, 33–34. [Google Scholar]
- Marco, E.M.; Rubino, T.; Adriani, W.; Viveros, M.P.; Parolaro, D.; Laviola, G. Long-term consequences of URB597 administration during adolescence on cannabinoid CB1 receptor binding in brain areas. Brain Res 2009, 1257, 25–31. [Google Scholar]
- Coutts, A.A.; Anavi-Goffer, S.; Ross, R.A.; MacEwan, D.J.; Mackie, K.; Pertwee, R.G.; Irving, A.J. Agonist-induced internalization and trafficking of cannabinoid CB1 receptors in hippocampal neurons. J. Neurosci 2001, 21, 2425–2433. [Google Scholar]
- Romero, J.; García, L.; Fernández-Ruiz, J.J.; Cebeira, M.; Ramos, J.A. Changes in rat brain cannabinoid binding sites after acute or chronic exposure to their endogenous agonist, anandamide, or to delta 9-tetrahydrocannabinol. Pharmacol. Biochem. Behav 1995, 51, 731–737. [Google Scholar]
- Heller, J.E.; Baty, D.E.; Zhang, M.; Li, H.; Adler, M.; Ganea, D.; Gaughan, J.; Loftus, C.M.; Jallo, J.I.; Tuma, R.F. The combination of selective inhibition of the cannabinoid CB1 receptor and activation of the cannabinoid CB2 receptor yields improved attenuation of motor and autonomic deficits in a mouse model of spinal cord injury. Clin. Neurosurg 2009, 56, 84–92. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Murillo-Rodríguez, E.; Sánchez-Alavez, M.; Navarro, L.; Martínez-Gonzalez, D.; Drucker-Colin, R.; Prospero-Garcia, O. Anandamida modulates sleep and memory in rats. Brain Res 1998, 812, 270–274. [Google Scholar]
- Hunter, A.J.; Hatcher, J.; Virley, D.; Nelson, P.; Irving, E.; Hadingham, S.J.; Parsons, A.A. Functional assessments in mice and rats after focal stroke. Neuropharmacology 2000, 39, 806–816. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Martinez-Vargas, M.; Morales-Gomez, J.; Gonzalez-Rivera, R.; Hernandez-Enriquez, C.; Perez-Arredondo, A.; Estrada-Rojo, F.; Navarro, L. Does the Neuroprotective Role of Anandamide Display Diurnal Variations? Int. J. Mol. Sci. 2013, 14, 23341-23355. https://doi.org/10.3390/ijms141223341
Martinez-Vargas M, Morales-Gomez J, Gonzalez-Rivera R, Hernandez-Enriquez C, Perez-Arredondo A, Estrada-Rojo F, Navarro L. Does the Neuroprotective Role of Anandamide Display Diurnal Variations? International Journal of Molecular Sciences. 2013; 14(12):23341-23355. https://doi.org/10.3390/ijms141223341
Chicago/Turabian StyleMartinez-Vargas, Marina, Julio Morales-Gomez, Ruben Gonzalez-Rivera, Carla Hernandez-Enriquez, Adan Perez-Arredondo, Francisco Estrada-Rojo, and Luz Navarro. 2013. "Does the Neuroprotective Role of Anandamide Display Diurnal Variations?" International Journal of Molecular Sciences 14, no. 12: 23341-23355. https://doi.org/10.3390/ijms141223341
APA StyleMartinez-Vargas, M., Morales-Gomez, J., Gonzalez-Rivera, R., Hernandez-Enriquez, C., Perez-Arredondo, A., Estrada-Rojo, F., & Navarro, L. (2013). Does the Neuroprotective Role of Anandamide Display Diurnal Variations? International Journal of Molecular Sciences, 14(12), 23341-23355. https://doi.org/10.3390/ijms141223341