Hyperoside Downregulates the Receptor for Advanced Glycation End Products (RAGE) and Promotes Proliferation in ECV304 Cells via the c-Jun N-Terminal Kinases (JNK) Pathway Following Stimulation by Advanced Glycation End-Products In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
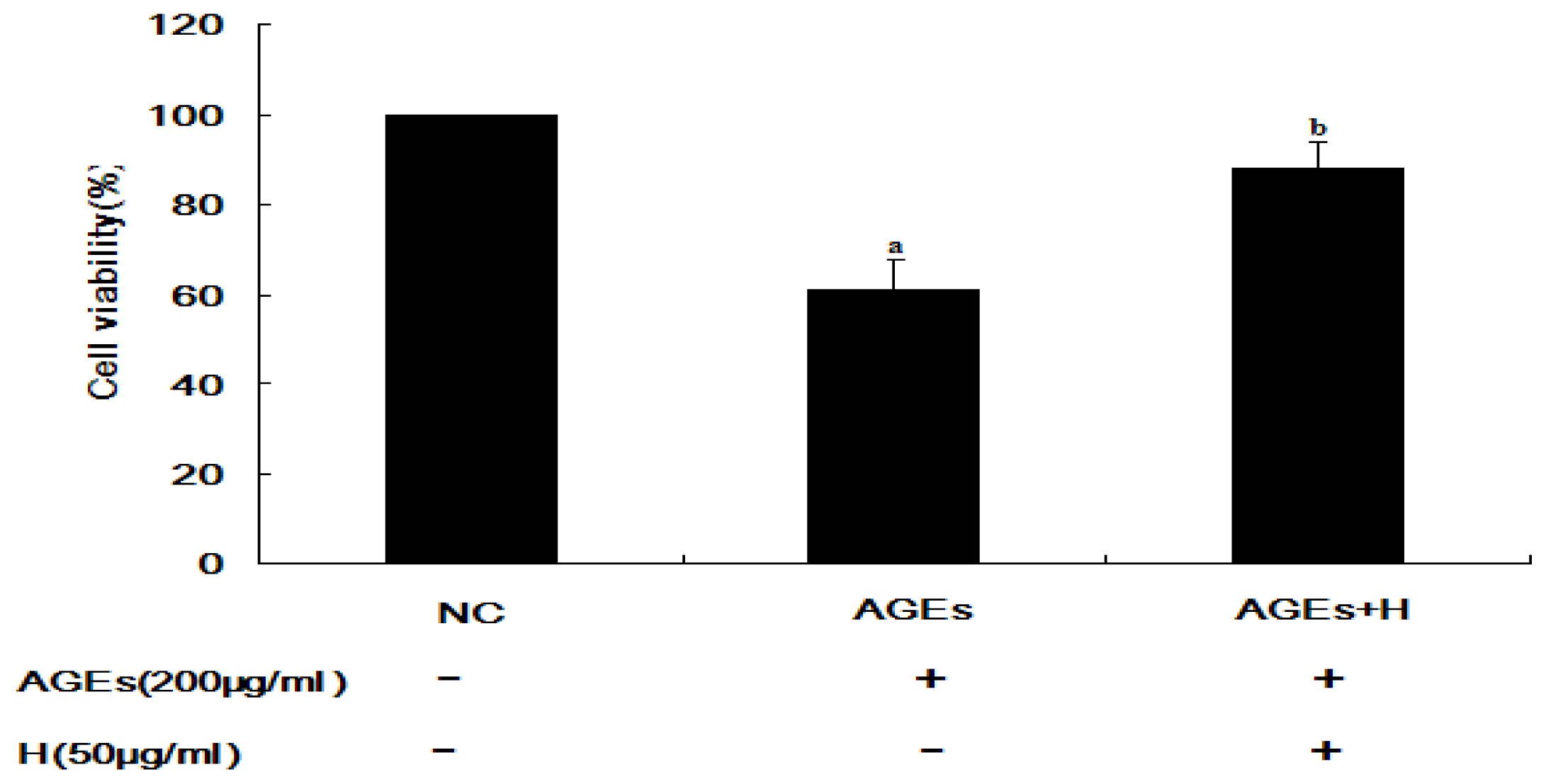
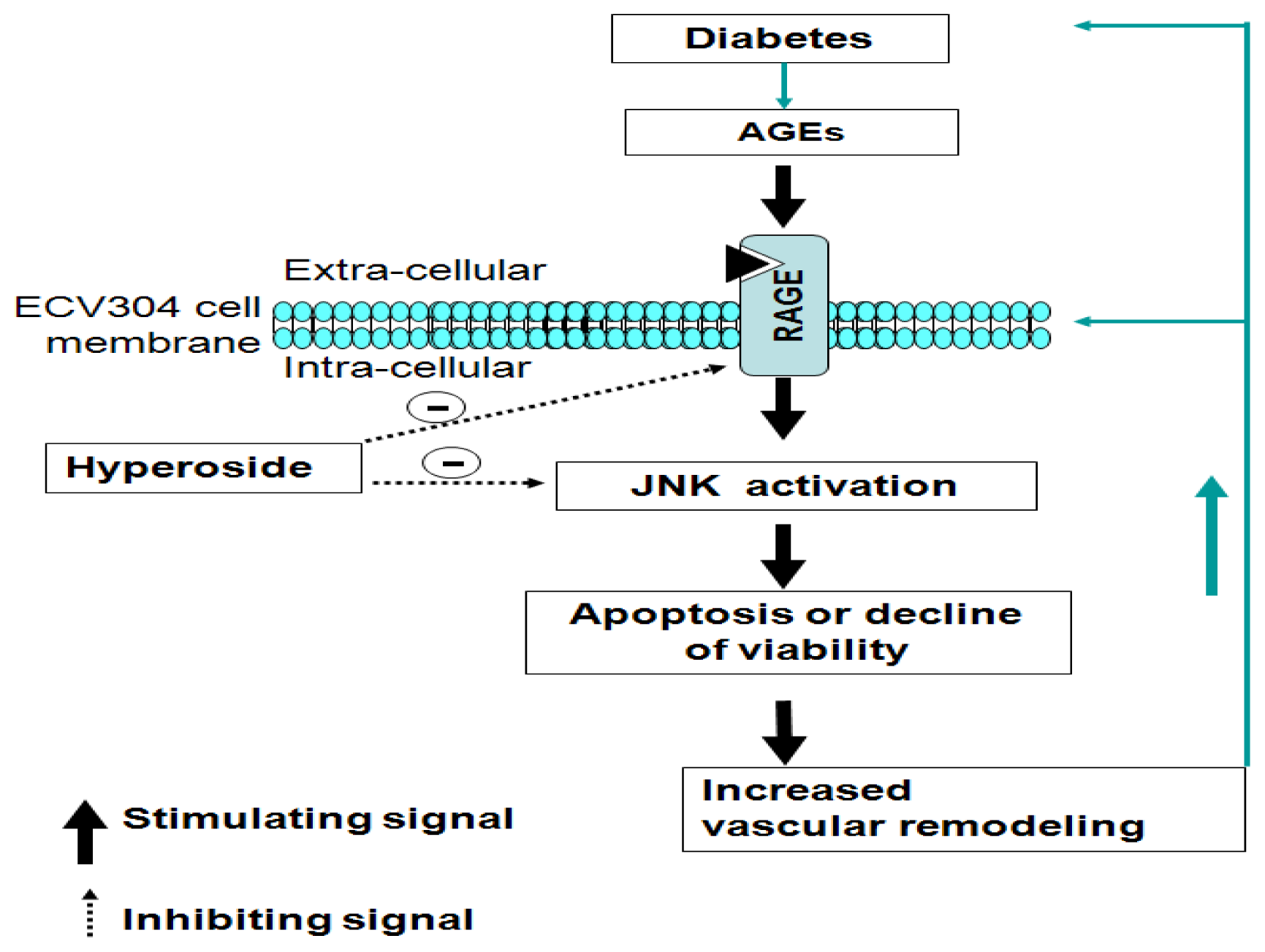
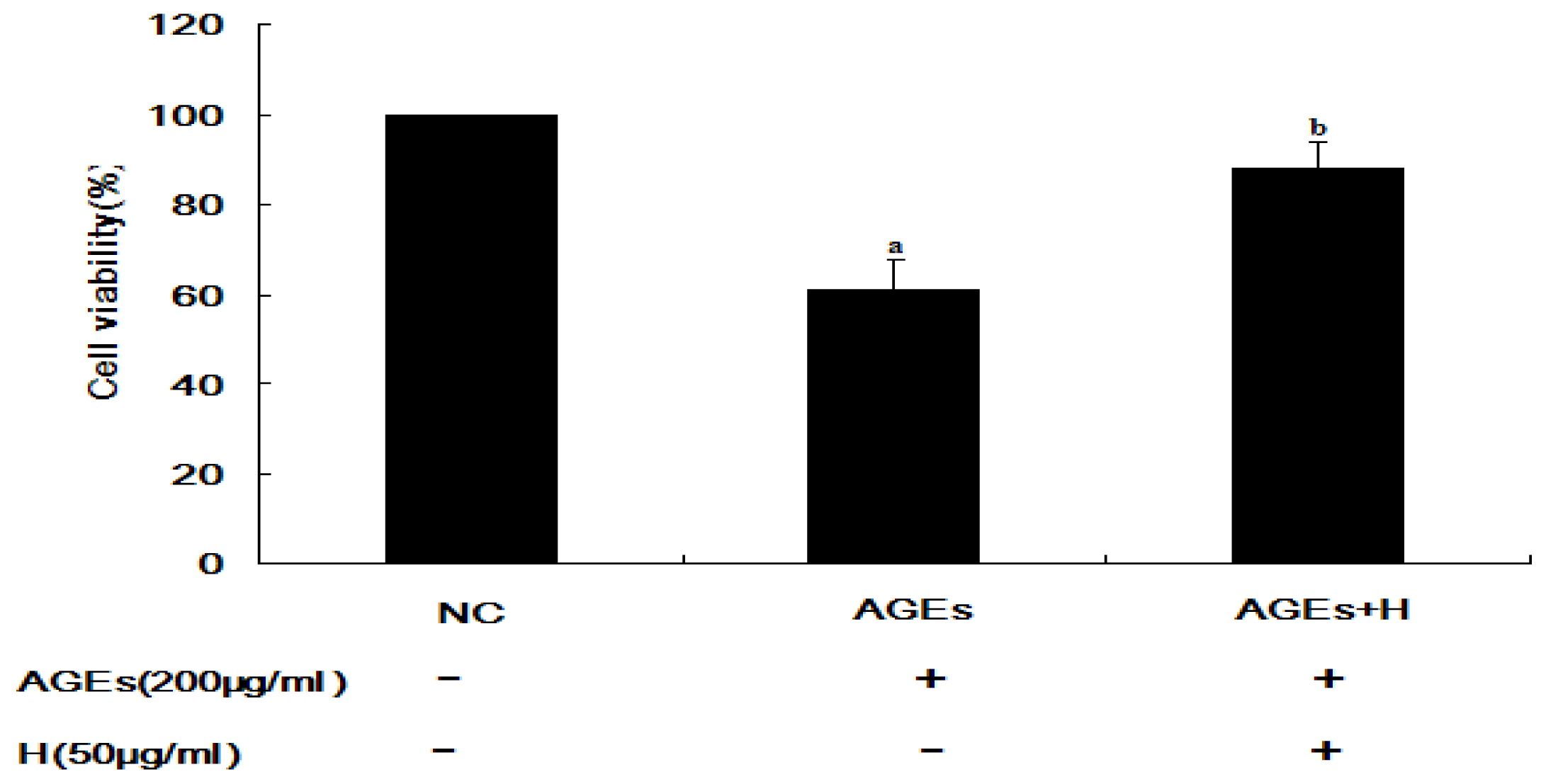
2.1. Hyperoside Promotes Cell Viability and Attenuates the Effects of AGEs (MTT)
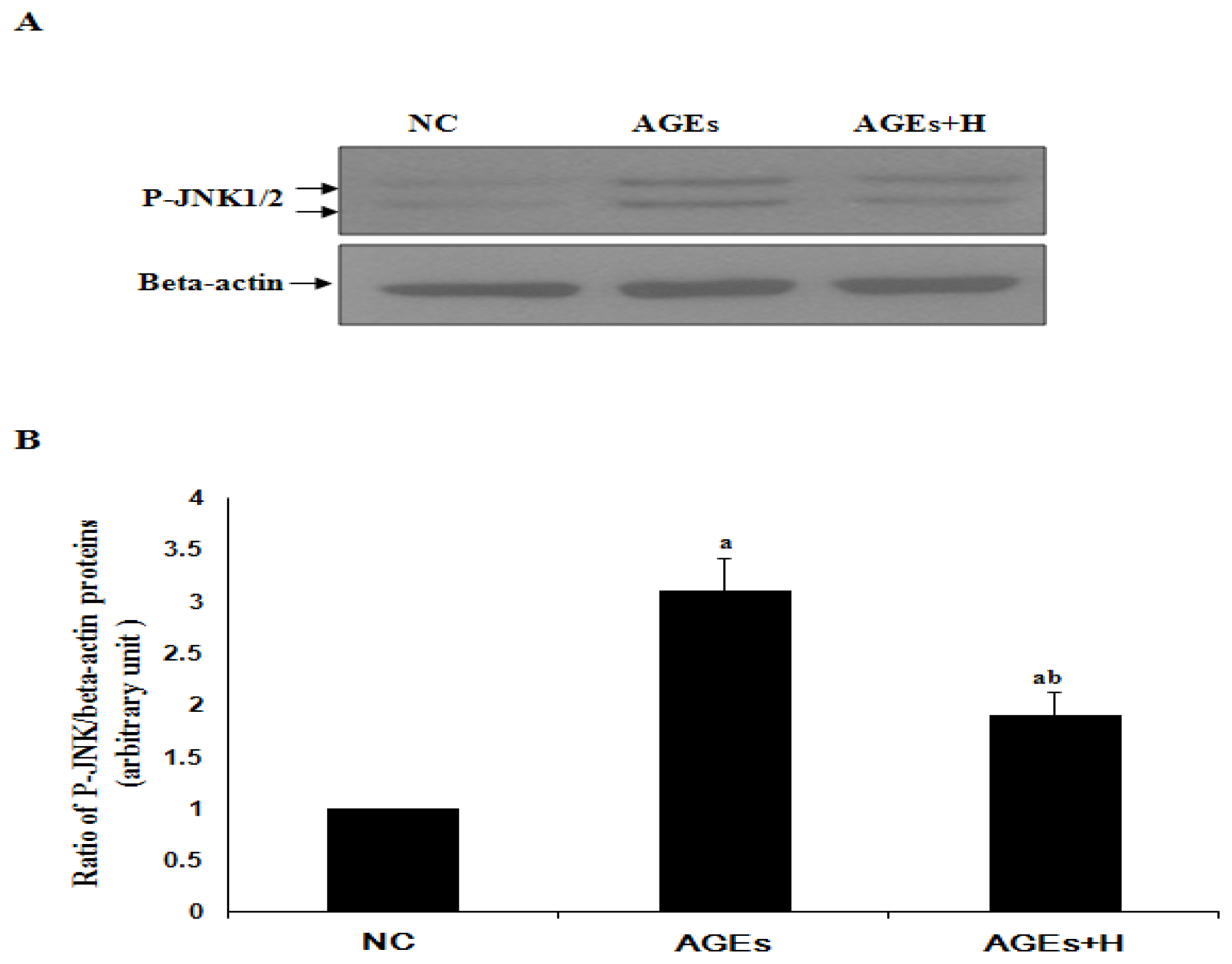
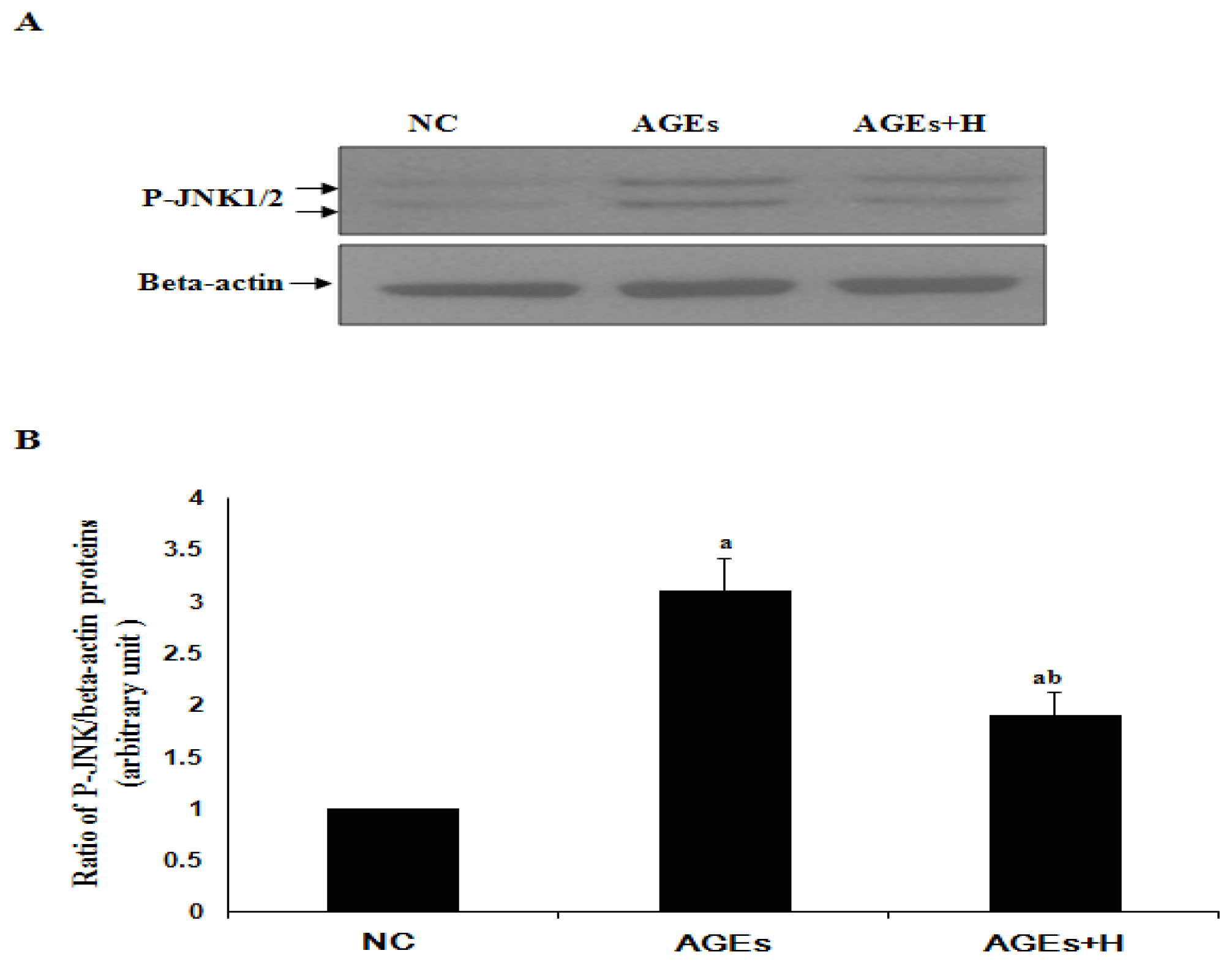
2.2. Hyperoside Inhibits AGE-Induced Activation of JNK in ECV304 Cells
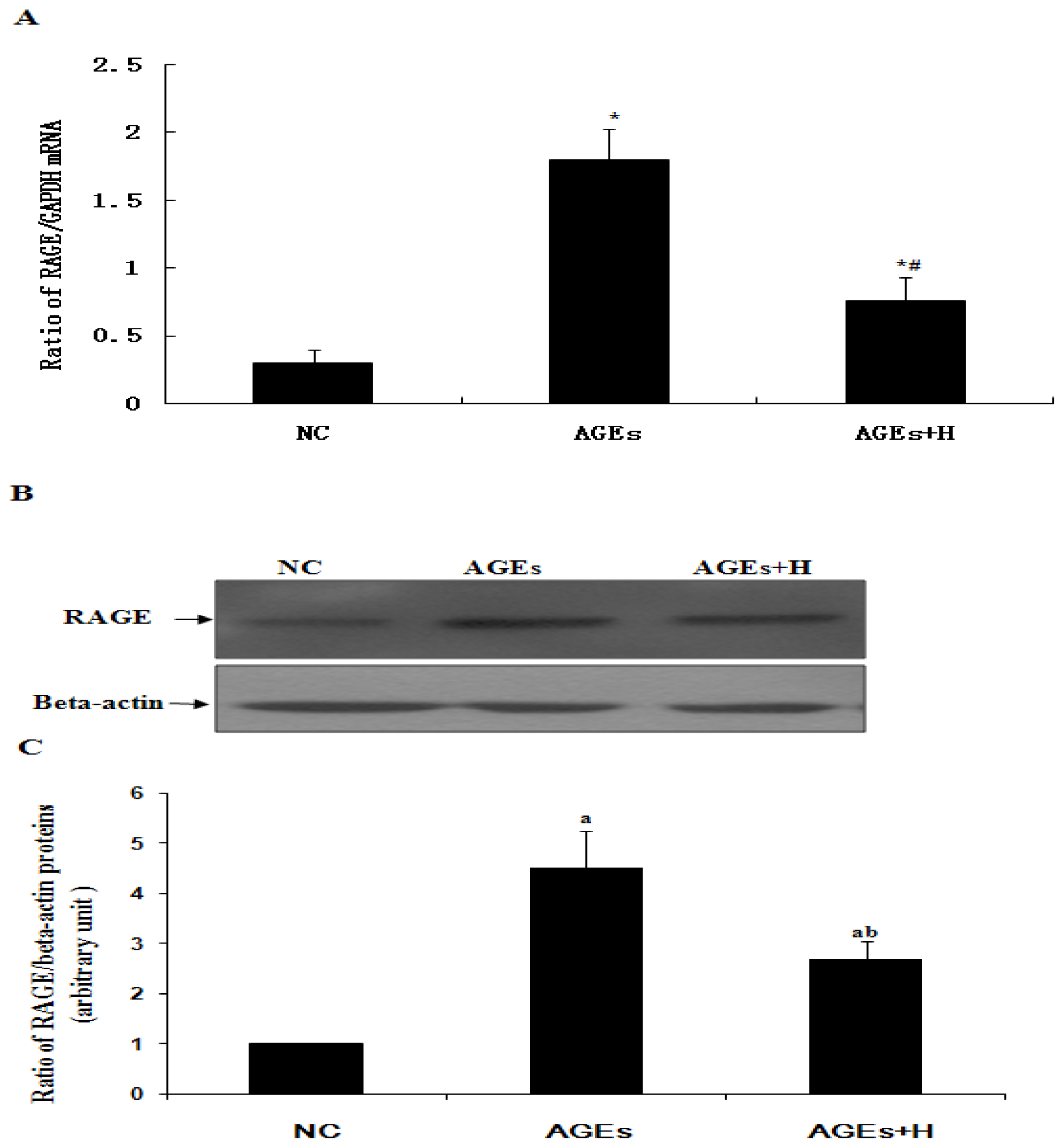
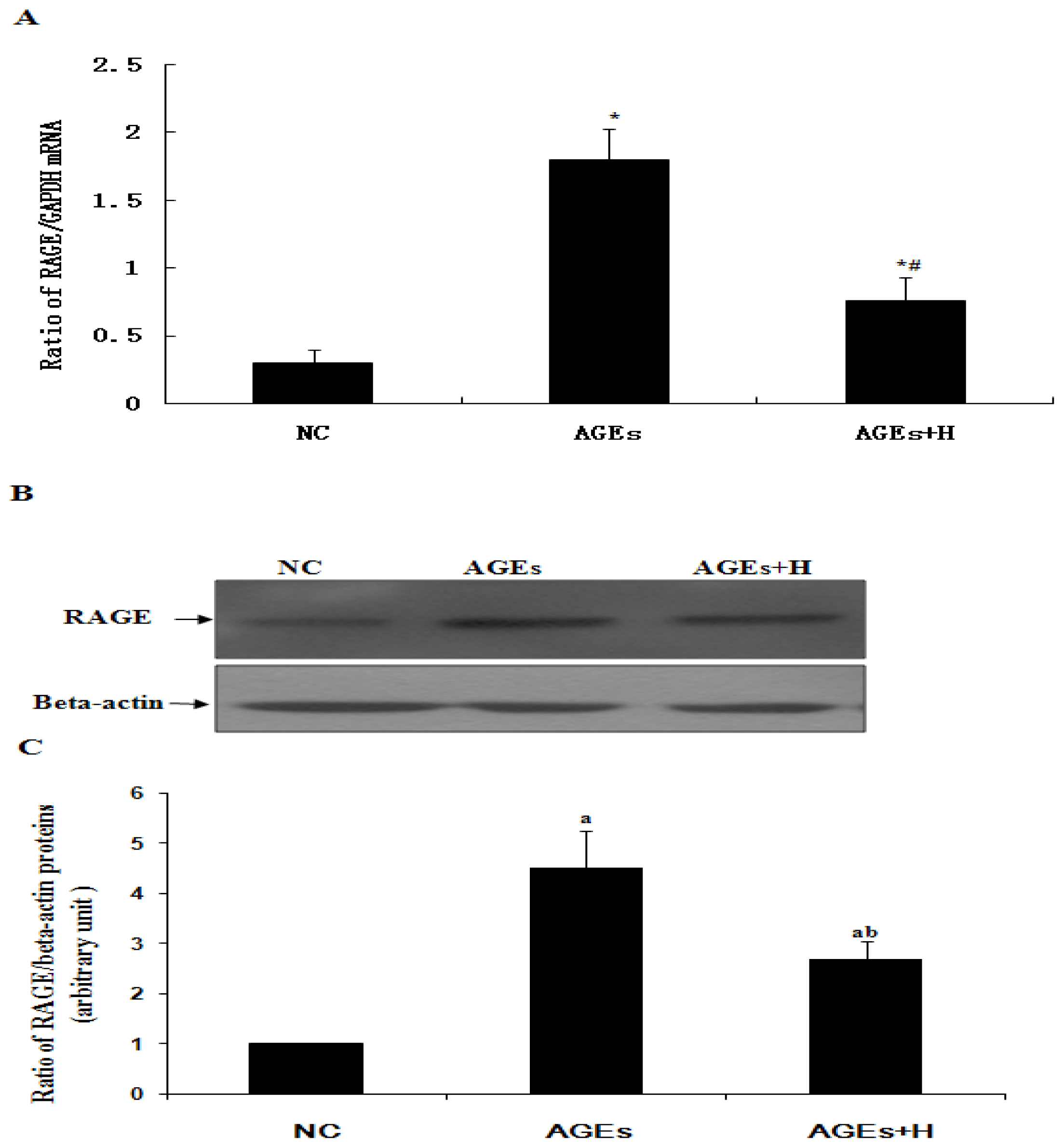
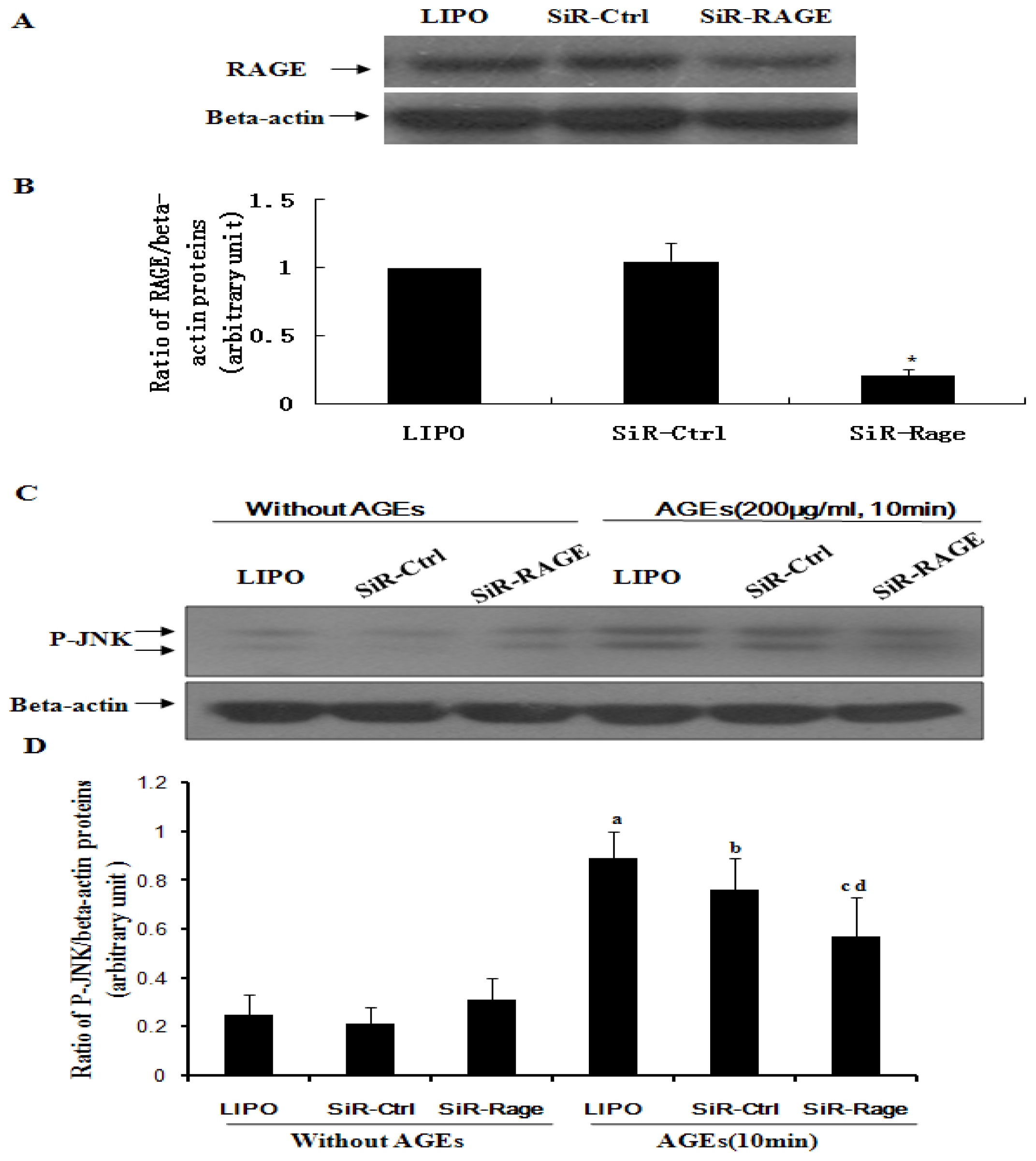
2.3. Hyperoside Downregulates AGE-Induced Expression of RAGE in ECV304 Cells
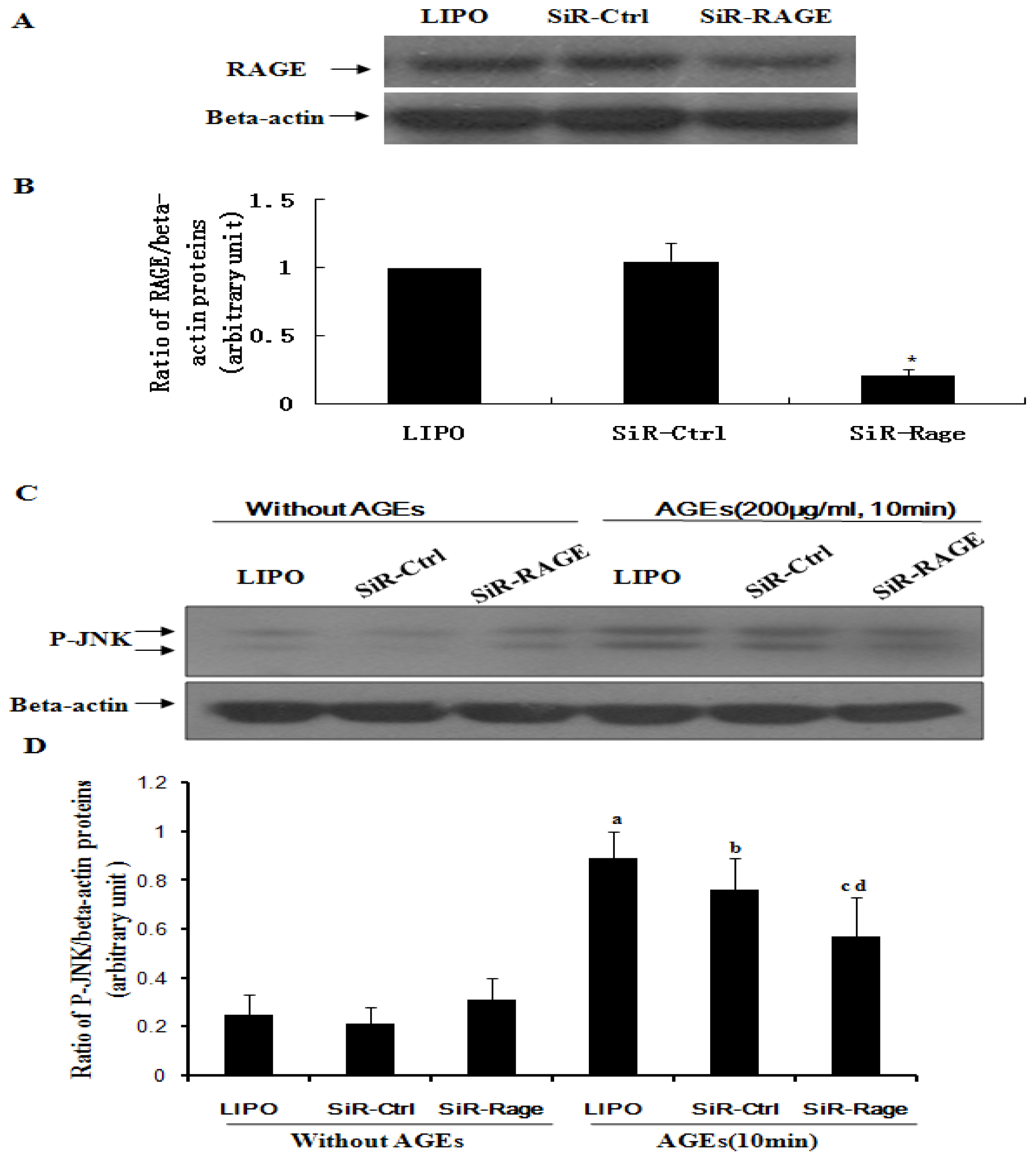
2.4. Knockdown of RAGE Inhibits AGE-Induced Activation of JNK in ECV304 Cells
2.5. Discussion
3. Materials and Methods
3.1. ECV304 Cell Culture
3.2. AGE Preparation
3.3. RNA Interference
3.4. Cell Viability Assay (MTT)
3.5. Knockdown of RAGE in ECV304 Cells
3.6. Western Blot Analysis
3.7. Real-Time qPCR
3.8. Statistical Analysis
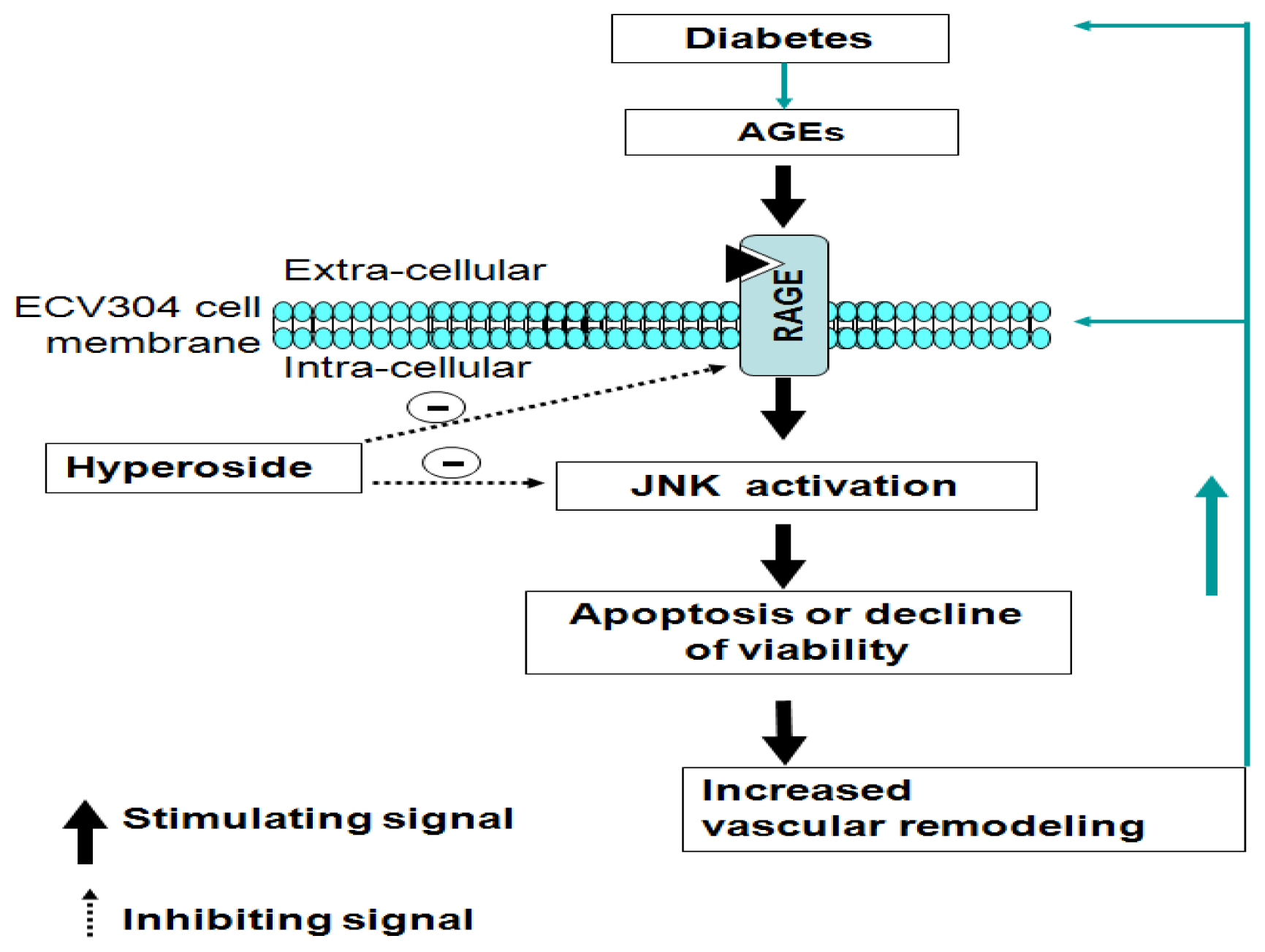
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Li, H.B.; Yi, X.; Gao, J.M.; Ying, X.X.; Guan, H.Q.; Li, J.C. The mechanism of hyperoside protection of ecv-304 cells against tert-butyl hydroperoxide-induced injury. Pharmacology 2008, 82, 105–113. [Google Scholar]
- Verma, N.; Amresh, G.; Sahu, P.K.; Mishra, N.; Rao Ch, V.; Singh, A.P. Pharmacological evaluation of hyperin for antihyperglycemic activity and effect on lipid profile in diabetic rats. Indian J. Exp. Biol 2013, 51, 65–72. [Google Scholar]
- Yamamoto, Y.; Yamamoto, H. Rage-mediated inflammation, type 2 diabetes, and diabetic vascular complication. Front. Endocrinol. (Lausanne) 2013, 4, 105. [Google Scholar]
- Tanaka, N.; Yonekura, H.; Yamagishi, S.; Fujimori, H.; Yamamoto, Y.; Yamamoto, H. The receptor for advanced glycation end products is induced by the glycation products themselves and tumor necrosis factor-alpha through nuclear factor-kappa b, and by 17beta-estradiol through sp-1 in human vascular endothelial cells. J. Biol. Chem 2000, 275, 25781–25790. [Google Scholar]
- Lin, W.D.; Lu, S.L.; Qing, C.; Yao, M.; Chen, X.F.; Xiang, J.; Qiao, L.; Liu, Y.K.; Dong, J.Y.; Shi, G.Y.; et al. Proliferation-inhibiting effect of advanced glycation end products modified human serum albumin to vascular endothelial cell ecv304. Zhonghua Yi Xue Za Zhi 2003, 83, 572–576. [Google Scholar]
- Chen, X.F.; Lin, W.D.; Lu, S.L.; Xie, T.; Ge, K.; Shi, Y.Q.; Zou, J.J.; Liu, Z.M.; Liao, W.Q. Mechanistic study of endogenous skin lesions in diabetic rats. Exp. Dermatol 2010, 19, 1088–1095. [Google Scholar]
- Neeper, M.; Schmidt, A.M.; Brett, J.; Yan, S.D.; Wang, F.; Pan, Y.C.; Elliston, K.; Stern, D.; Shaw, A. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J. Biol. Chem 1992, 267, 14998–15004. [Google Scholar]
- Brett, J.; Schmidt, A.M.; Yan, S.D.; Zou, Y.S.; Weidman, E.; Pinsky, D.; Nowygrod, R.; Neeper, M.; Przysiecki, C.; Shaw, A.; et al. Survey of the distribution of a newly characterized receptor for advanced glycation end products in tissues. Am. J. Pathol 1993, 143, 1699–1712. [Google Scholar]
- Schmidt, A.M.; Hori, O.; Brett, J.; Yan, S.D.; Wautier, J.L.; Stern, D. Cellular receptors for advanced glycation end products. Implications for induction of oxidant stress and cellular dysfunction in the pathogenesis of vascular lesions. Arterioscler. Thromb 1994, 14, 1521–1528. [Google Scholar]
- Schmidt, A.M.; Yan, S.D.; Wautier, J.L.; Stern, D. Activation of receptor for advanced glycation end products: A mechanism for chronic vascular dysfunction in diabetic vasculopathy and atherosclerosis. Circ. Res 1999, 84, 489–497. [Google Scholar]
- Yan, S.D.; Schmidt, A.M.; Anderson, G.M.; Zhang, J.; Brett, J.; Zou, Y.S.; Pinsky, D.; Stern, D. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J. Biol. Chem 1994, 269, 9889–9897. [Google Scholar]
- Lander, H.M.; Tauras, J.M.; Ogiste, J.S.; Hori, O.; Moss, R.A.; Schmidt, A.M. Activation of the receptor for advanced glycation end products triggers a p21(ras)-dependent mitogen-activated protein kinase pathway regulated by oxidant stress. J. Biol. Chem 1997, 272, 17810–17814. [Google Scholar]
- Shi, L.; Yu, X.; Yang, H.; Wu, X. Advanced glycation end products induce human corneal epithelial cells apoptosis through generation of reactive oxygen species and activation of jnk and p38 mapk pathways. PLoS One 2013, 8, e66781. [Google Scholar]
- Yamagishi, S.; Matsui, T.; Nakamura, K.; Takeuchi, M.; Inoue, H. Telmisartan inhibits advanced glycation end products (ages)-elicited endothelial cell injury by suppressing age receptor (rage) expression via peroxisome proliferator-activated receptor-gammaactivation. Protein Pept. Lett 2008, 15, 850–853. [Google Scholar]
- Liu, Y.; Liang, C.; Liu, X.; Liao, B.; Pan, X.; Ren, Y.; Fan, M.; Li, M.; He, Z.; Wu, J.; et al. Ages increased migration and inflammatory responses of adventitial fibroblasts via rage, mapk and nf-kappab pathways. Atherosclerosis 2010, 208, 34–42. [Google Scholar]
- de Bittencourt Pasquali, M.A.; Gelain, D.P.; Zeidan-Chulia, F.; Pires, A.S.; Gasparotto, J.; Terra, S.R.; Moreira, J.C. Vitamin a (retinol) downregulates the receptor for advanced glycation endproducts (rage) by oxidant-dependent activation of p38 mapk and nf-kb in human lung cancer a549 cells. Cell. Signal 2013, 25, 939–954. [Google Scholar]
- Pillai, S.S.; Sugathan, J.K.; Indira, M. Selenium downregulates rage and nfkappab expression in diabetic rats. Biol. Trace Elem. Res 2012, 149, 71–77. [Google Scholar]
- Wei, W.; Chen, M.; Zhu, Y.; Wang, J.; Zhu, P.; Li, Y.; Li, J. Down-regulation of vascular hmgb1 and rage expression by n-3 polyunsaturated fatty acids is accompanied by amelioration of chronic vasculopathy of small bowel allografts. J. Nutr. Biochem 2012, 23, 1333–1340. [Google Scholar]
- Cuccurullo, C.; Iezzi, A.; Fazia, M.L.; De Cesare, D.; Di Francesco, A.; Muraro, R.; Bei, R.; Ucchino, S.; Spigonardo, F.; Chiarelli, F.; et al. Suppression of rage as a basis of simvastatin-dependent plaque stabilization in type 2 diabetes. Arterioscler. Thromb. Vasc. Biol 2006, 26, 2716–2723. [Google Scholar]
- Marx, N.; Walcher, D.; Ivanova, N.; Rautzenberg, K.; Jung, A.; Friedl, R.; Hombach, V.; de Caterina, R.; Basta, G.; Wautier, M.P.; et al. Thiazolidinediones reduce endothelial expression of receptors for advanced glycation end products. Diabetes 2004, 53, 2662–2668. [Google Scholar]
- Wang, K.; Zhou, Z.; Zhang, M.; Fan, L.; Forudi, F.; Zhou, X.; Qu, W.; Lincoff, A.M.; Schmidt, A.M.; Topol, E.J.; et al. Peroxisome proliferator-activated receptor gamma down-regulates receptor for advanced glycation end products and inhibits smooth muscle cell proliferation in a diabetic and nondiabetic rat carotid artery injury model. J. Pharmacol. Exp. Ther 2006, 317, 37–43. [Google Scholar]
- Yan, F.L.; Zheng, Y.; Zhao, F.D. Effects of ginkgo biloba extract egb761 on expression of rage and lrp-1 in cerebral microvascular endothelial cells under chronic hypoxia and hypoglycemia. Acta Neuropathol 2008, 116, 529–535. [Google Scholar]
- Dou, L.; Lu, Y.; Shen, T.; Huang, X.; Man, Y.; Wang, S.; Li, J. Panax notogingseng saponins suppress rage/mapk signaling and nf-kappab activation in apolipoprotein-e-deficient atherosclerosis-prone mice. Cell. Physiol. Biochem 2012, 29, 875–882. [Google Scholar]
- Liu, R.L.; Xiong, Q.J.; Shu, Q.; Wu, W.N.; Cheng, J.; Fu, H.; Wang, F.; Chen, J.G.; Hu, Z.L. Hyperoside protects cortical neurons from oxygen-glucose deprivation-reperfusion induced injury via nitric oxide signal pathway. Brain Res 2012, 1469, 164–173. [Google Scholar]
- Kim, J.Y.; Park, H.K.; Yoon, J.S.; Kim, S.J.; Kim, E.S.; Ahn, K.S.; Kim, D.S.; Yoon, S.S.; Kim, B.K.; Lee, Y.Y. Advanced glycation end product (age)-induced proliferation of hel cells via receptor for age-related signal pathways. Int. J. Oncol 2008, 33, 493–501. [Google Scholar]
- Galler, A.; Muller, G.; Schinzel, R.; Kratzsch, J.; Kiess, W.; Munch, G. Impact of metabolic control and serum lipids on the concentration of advanced glycation end products in the serum of children and adolescents with type 1 diabetes, as determined by fluorescence spectroscopy and nepsilon-(carboxymethyl)lysine elisa. Diabetes Care 2003, 26, 2609–2615. [Google Scholar]
- Li, Y.; Liu, S.; Zhang, Z.; Xu, Q.; Xie, F.; Wang, J.; Ping, S.; Li, C.; Wang, Z.; Zhang, M.; et al. Rage mediates accelerated diabetic vein graft atherosclerosis induced by combined mechanical stress and ages via synergistic erk activation. PLoS One 2012, 7, e35016. [Google Scholar]
- Zhang, Z.; Zhang, M.; Li, Y.; Liu, S.; Ping, S.; Wang, J.; Ning, F.; Xie, F.; Li, C. Simvastatin inhibits the additive activation of erk1/2 and proliferation of rat vascular smooth muscle cells induced by combined mechanical stress and oxldl through lox-1 pathway. Cell. Signal 2013, 25, 332–340. [Google Scholar]
- Yamada, Y.; Ishibashi, K.; Bhutto, I.A.; Tian, J.; Lutty, G.A.; Handa, J.T. The expression of advanced glycation endproduct receptors in rpe cells associated with basal deposits in human maculas. Exp. Eye Res 2006, 82, 840–848. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Z.; Sethiel, M.S.; Shen, W.; Liao, S.; Zou, Y. Hyperoside Downregulates the Receptor for Advanced Glycation End Products (RAGE) and Promotes Proliferation in ECV304 Cells via the c-Jun N-Terminal Kinases (JNK) Pathway Following Stimulation by Advanced Glycation End-Products In Vitro. Int. J. Mol. Sci. 2013, 14, 22697-22707. https://doi.org/10.3390/ijms141122697
Zhang Z, Sethiel MS, Shen W, Liao S, Zou Y. Hyperoside Downregulates the Receptor for Advanced Glycation End Products (RAGE) and Promotes Proliferation in ECV304 Cells via the c-Jun N-Terminal Kinases (JNK) Pathway Following Stimulation by Advanced Glycation End-Products In Vitro. International Journal of Molecular Sciences. 2013; 14(11):22697-22707. https://doi.org/10.3390/ijms141122697
Chicago/Turabian StyleZhang, Zhengyu, Mosha Silas Sethiel, Weizhi Shen, Sentai Liao, and Yuxiao Zou. 2013. "Hyperoside Downregulates the Receptor for Advanced Glycation End Products (RAGE) and Promotes Proliferation in ECV304 Cells via the c-Jun N-Terminal Kinases (JNK) Pathway Following Stimulation by Advanced Glycation End-Products In Vitro" International Journal of Molecular Sciences 14, no. 11: 22697-22707. https://doi.org/10.3390/ijms141122697