Anti-Biofilm Performance of Three Natural Products against Initial Bacterial Attachment

Abstract
:1. Introduction
2. Results and Discussion
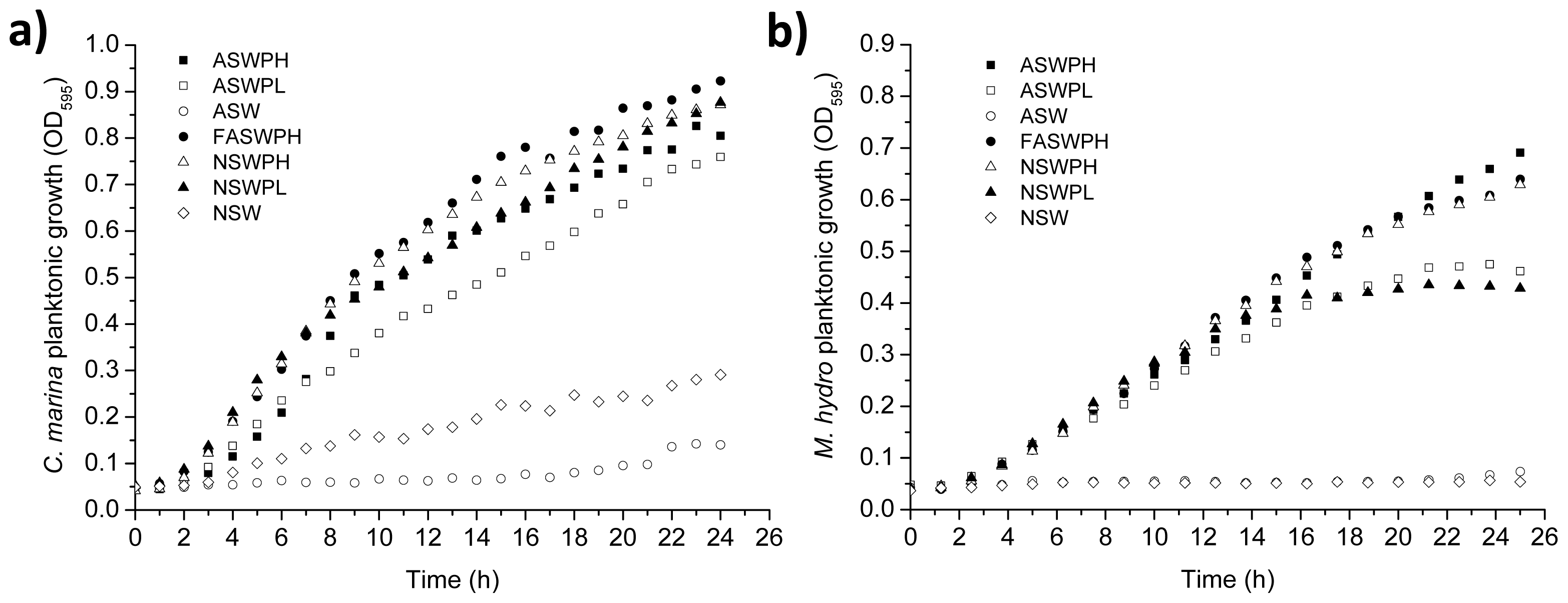
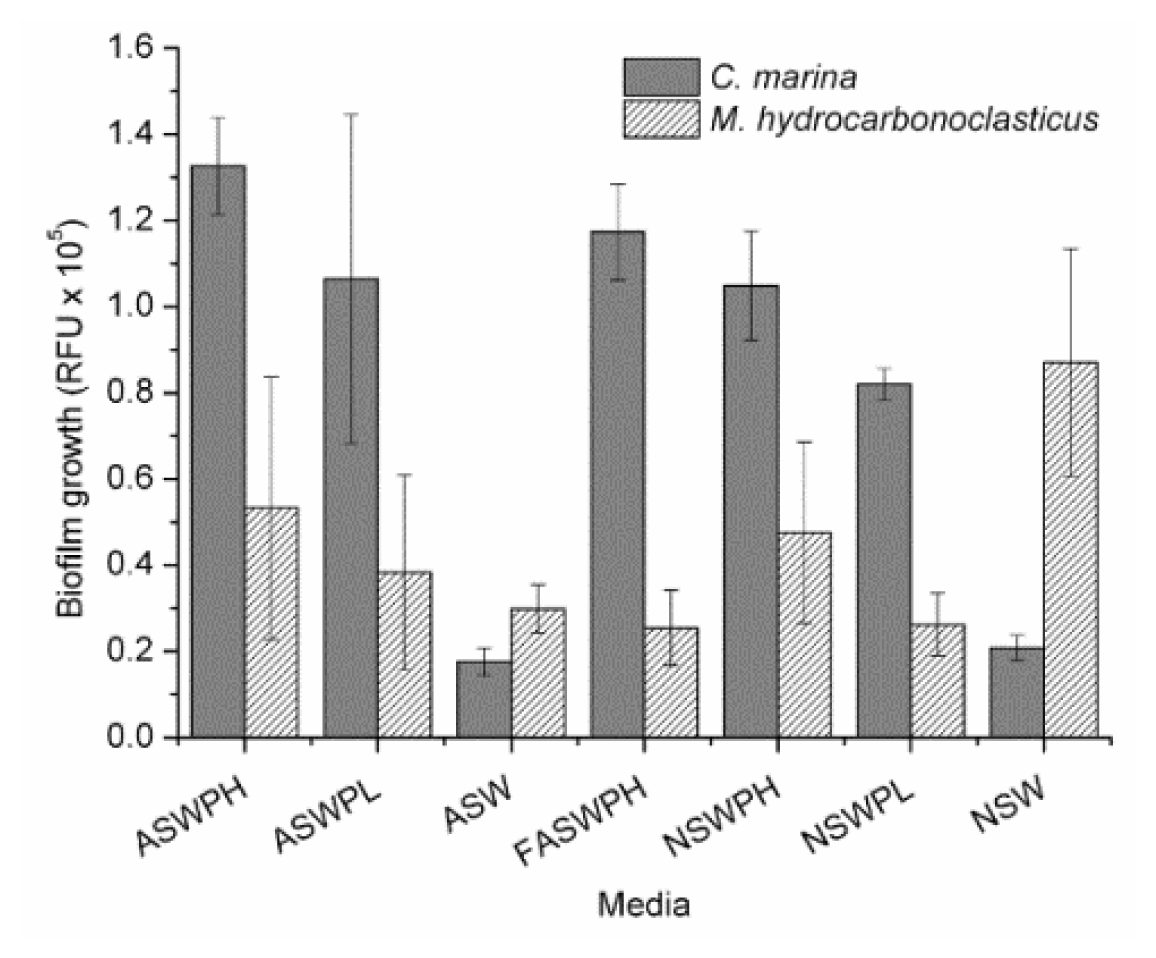
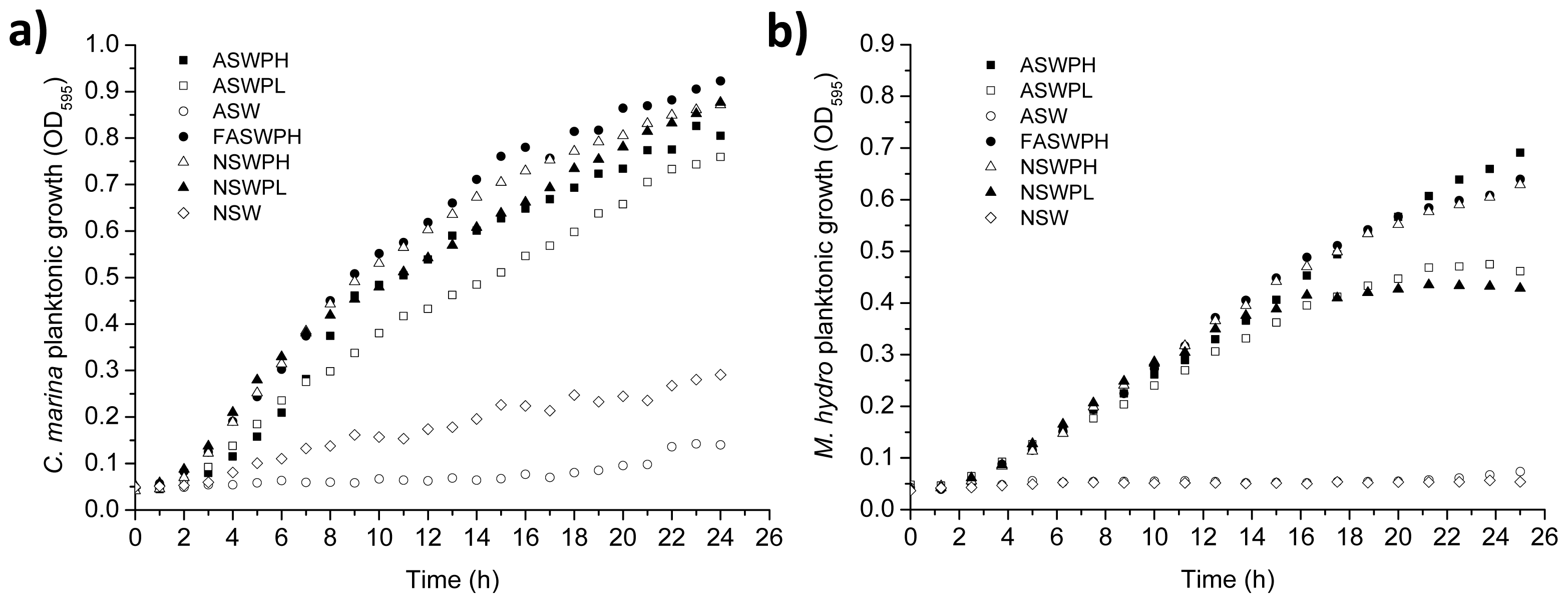
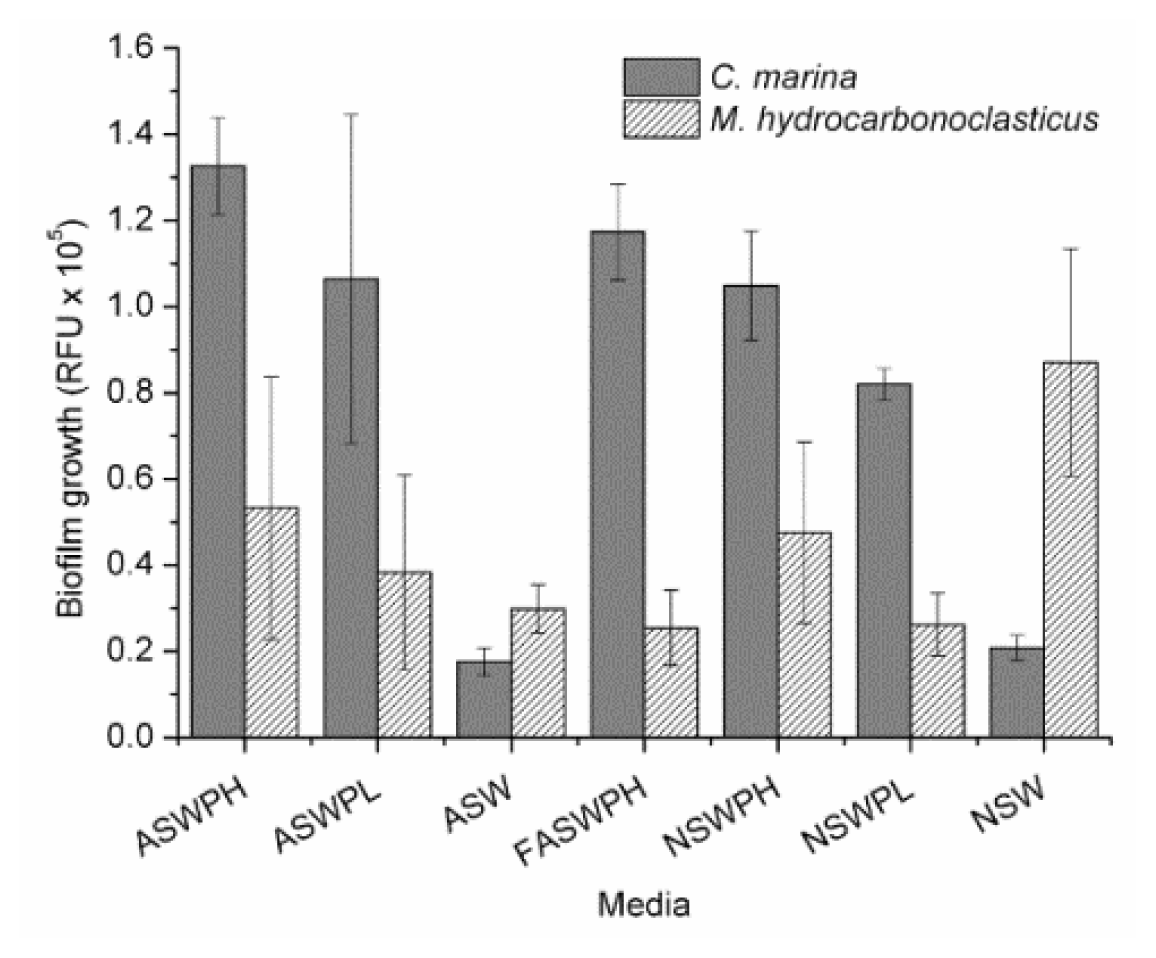
2.1. Growth Conditions (ASW vs. NSW)
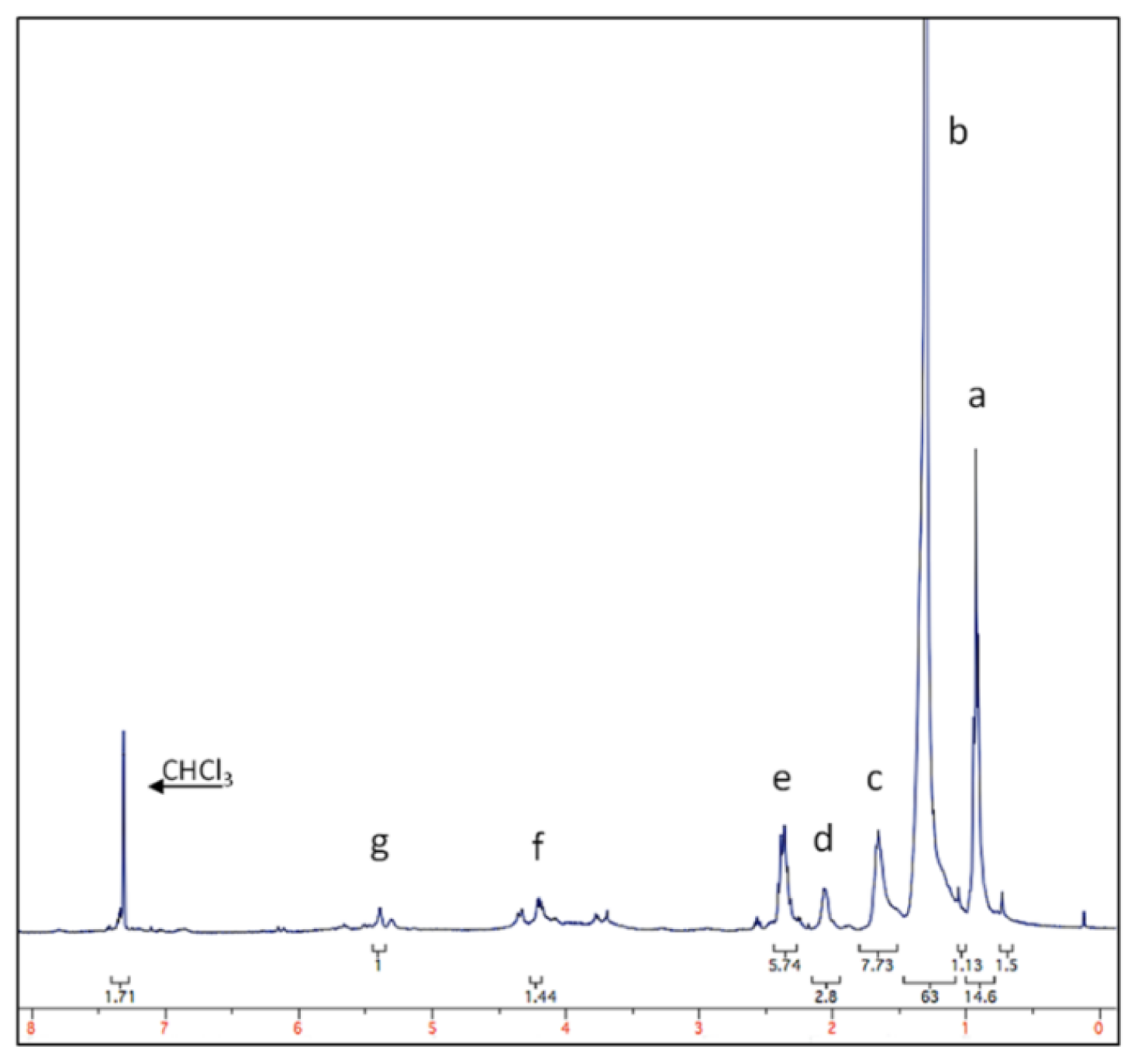
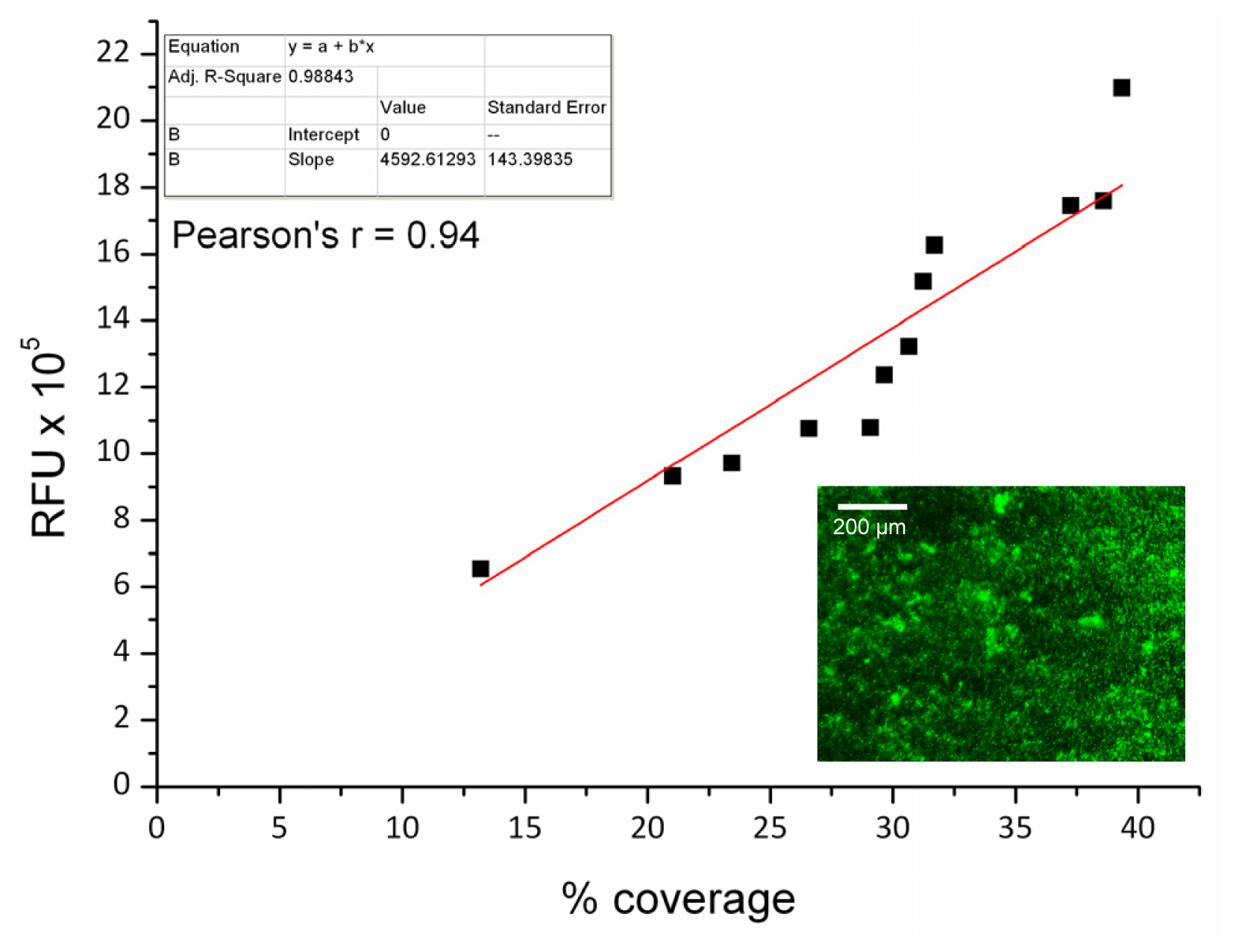
2.2. Protocol Verification
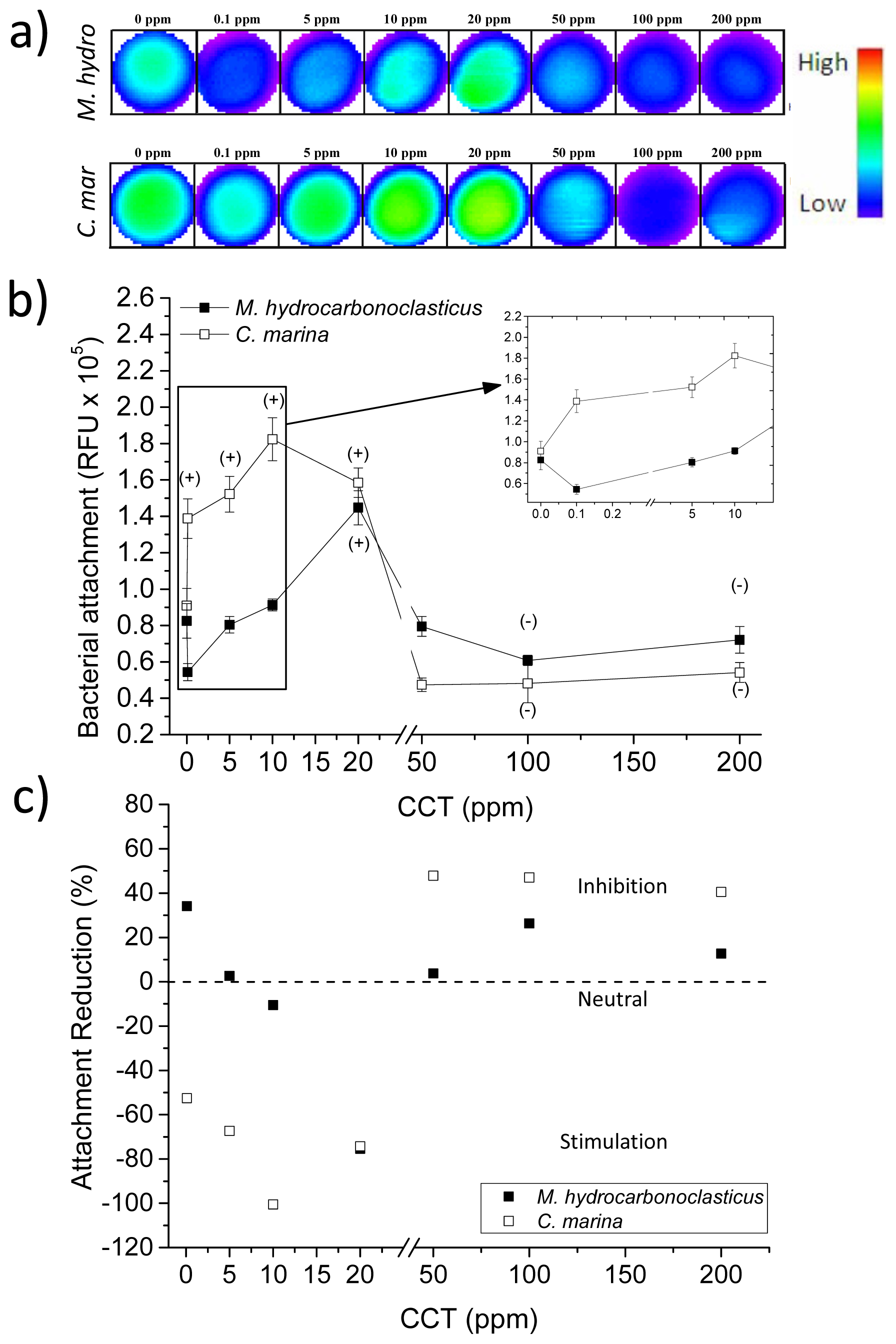
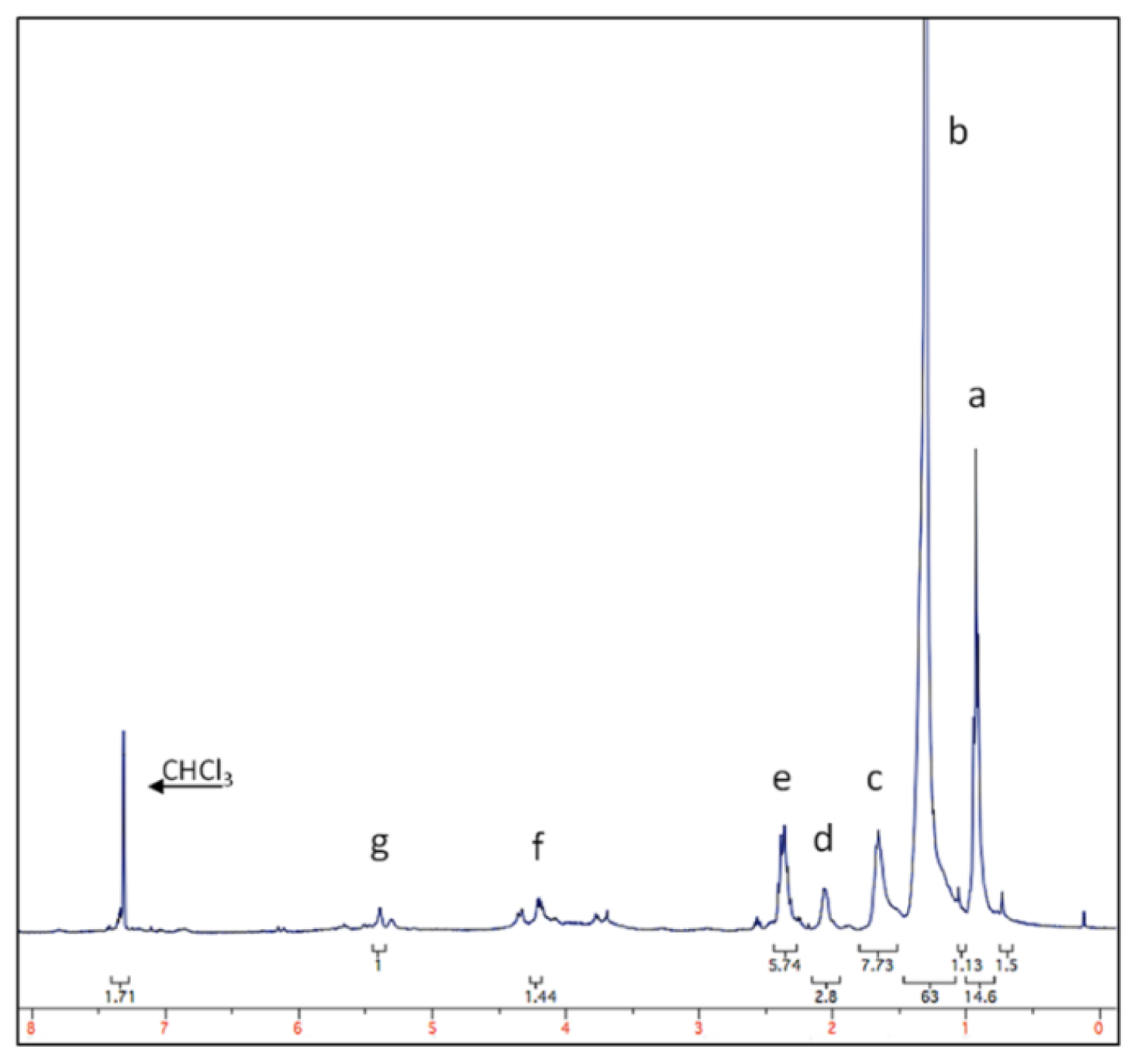
2.3. Chondrus crispus Extracts and Bacterial Attachment
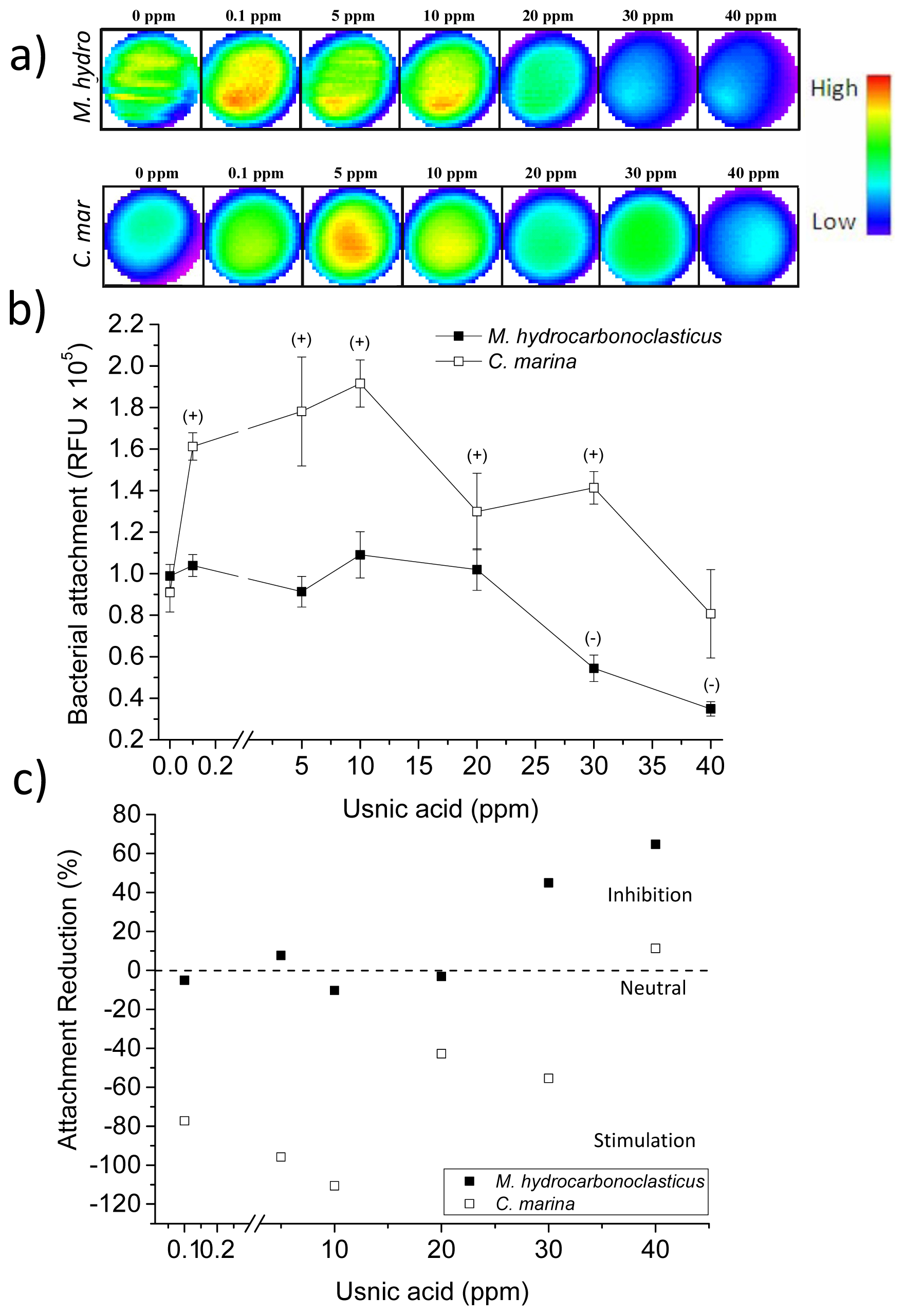
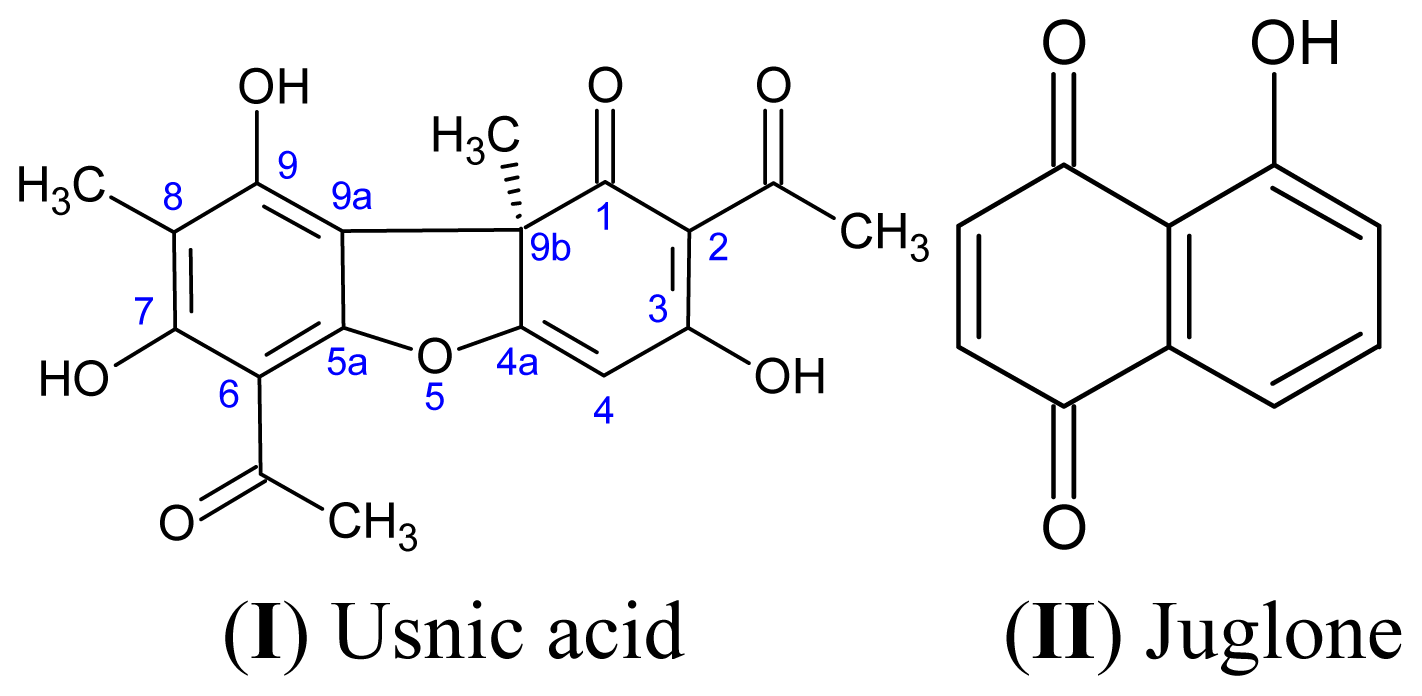
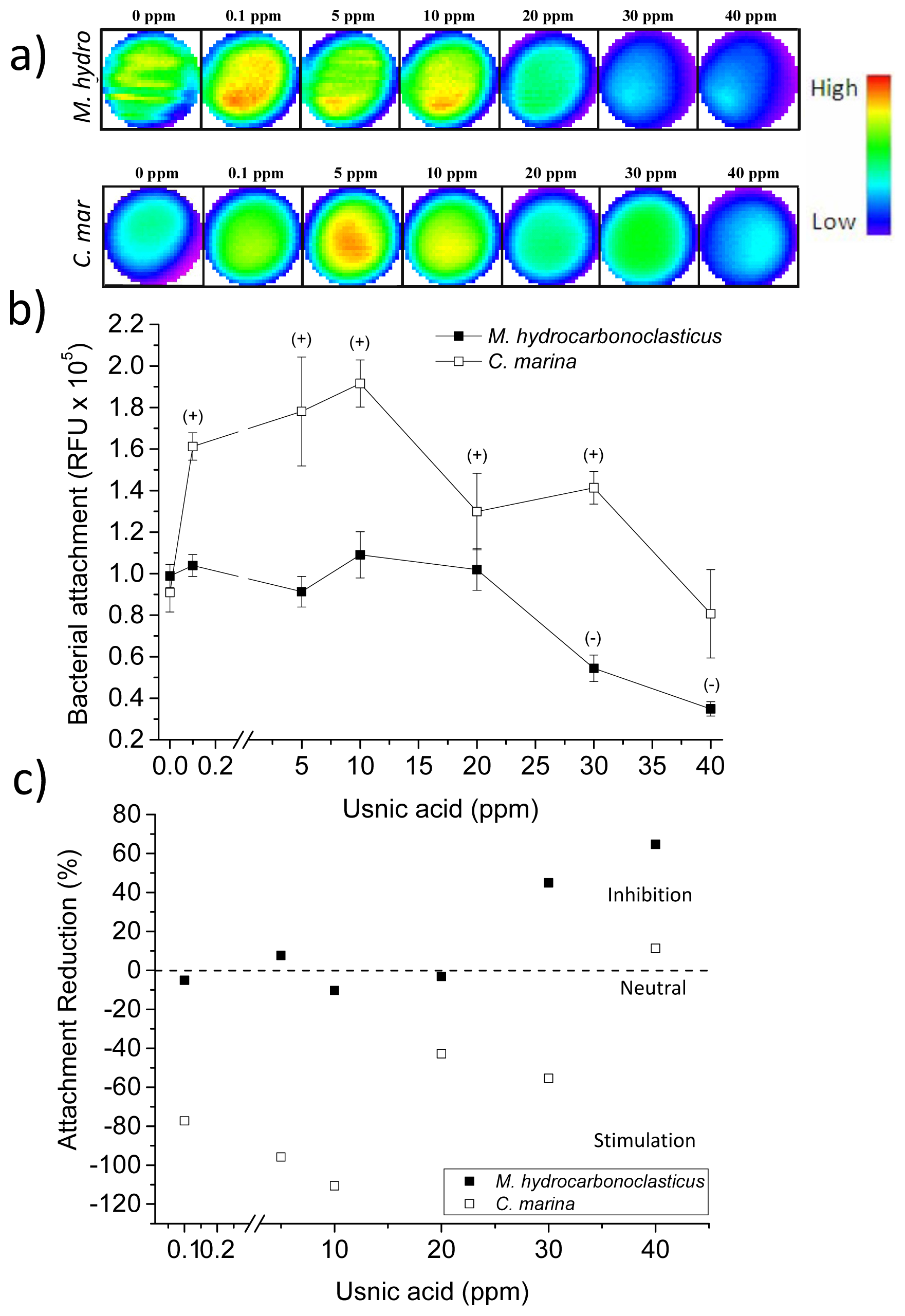
2.4. Usnic Acid and Bacterial Attachment
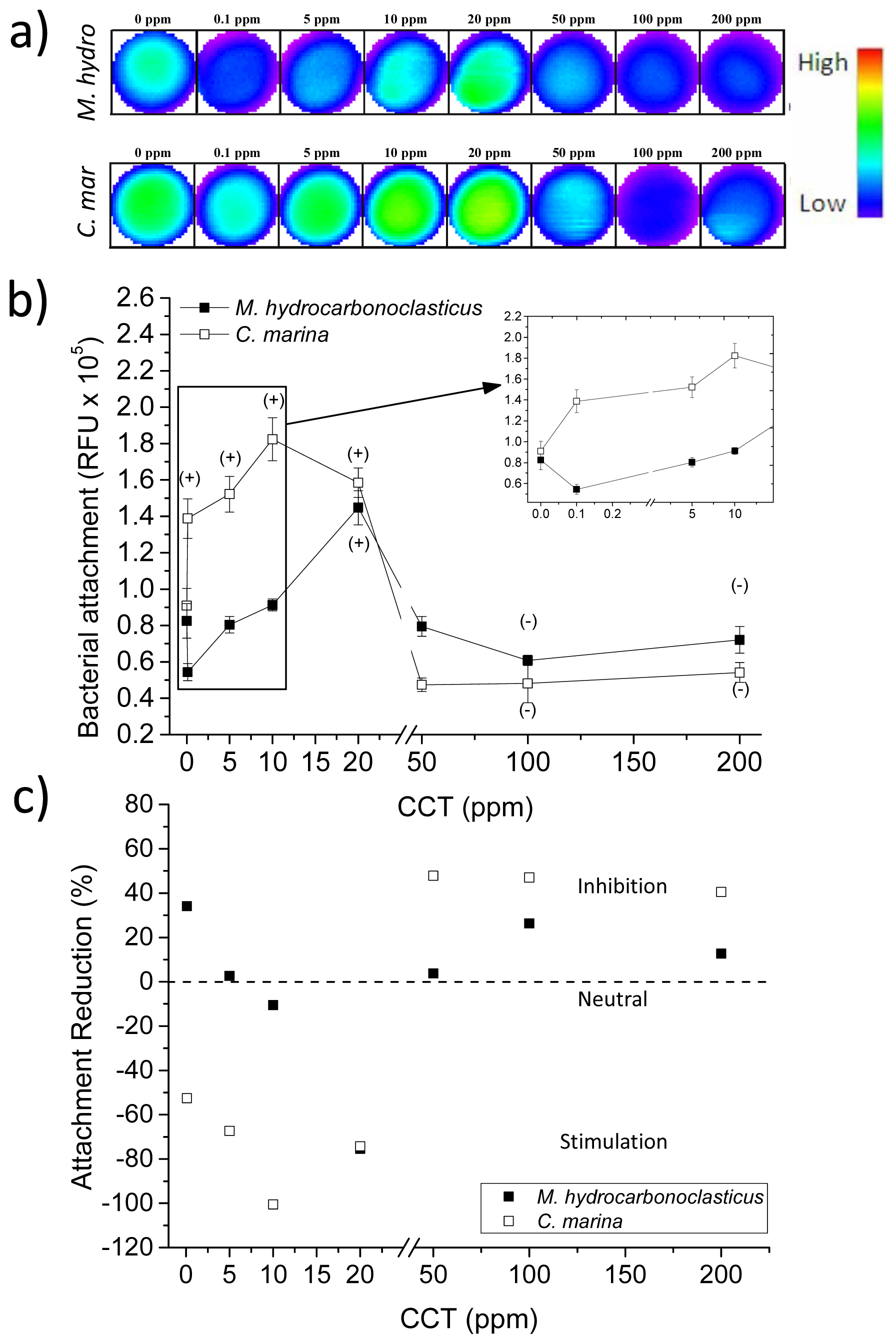
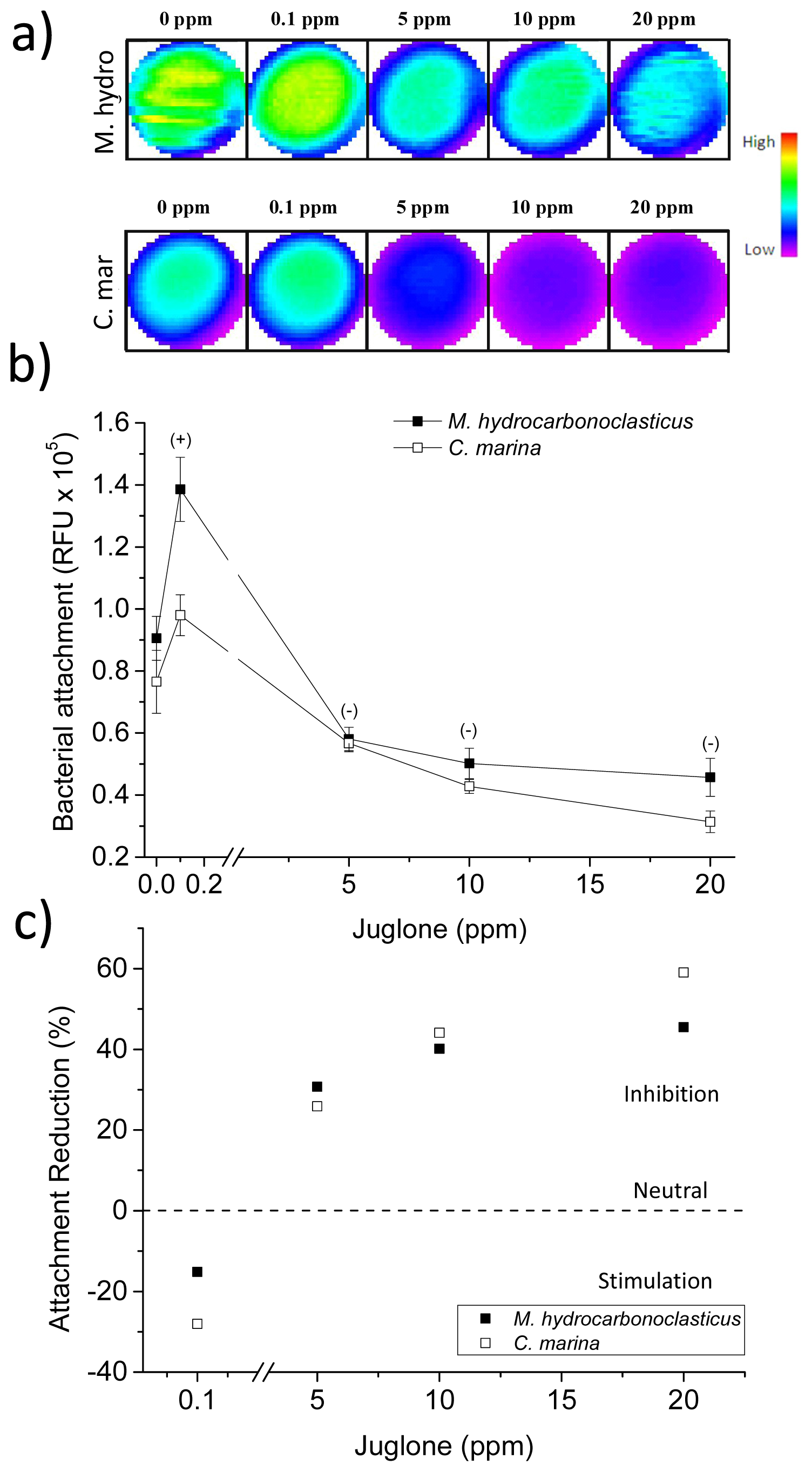
2.5. Juglone and Bacterial Attachment
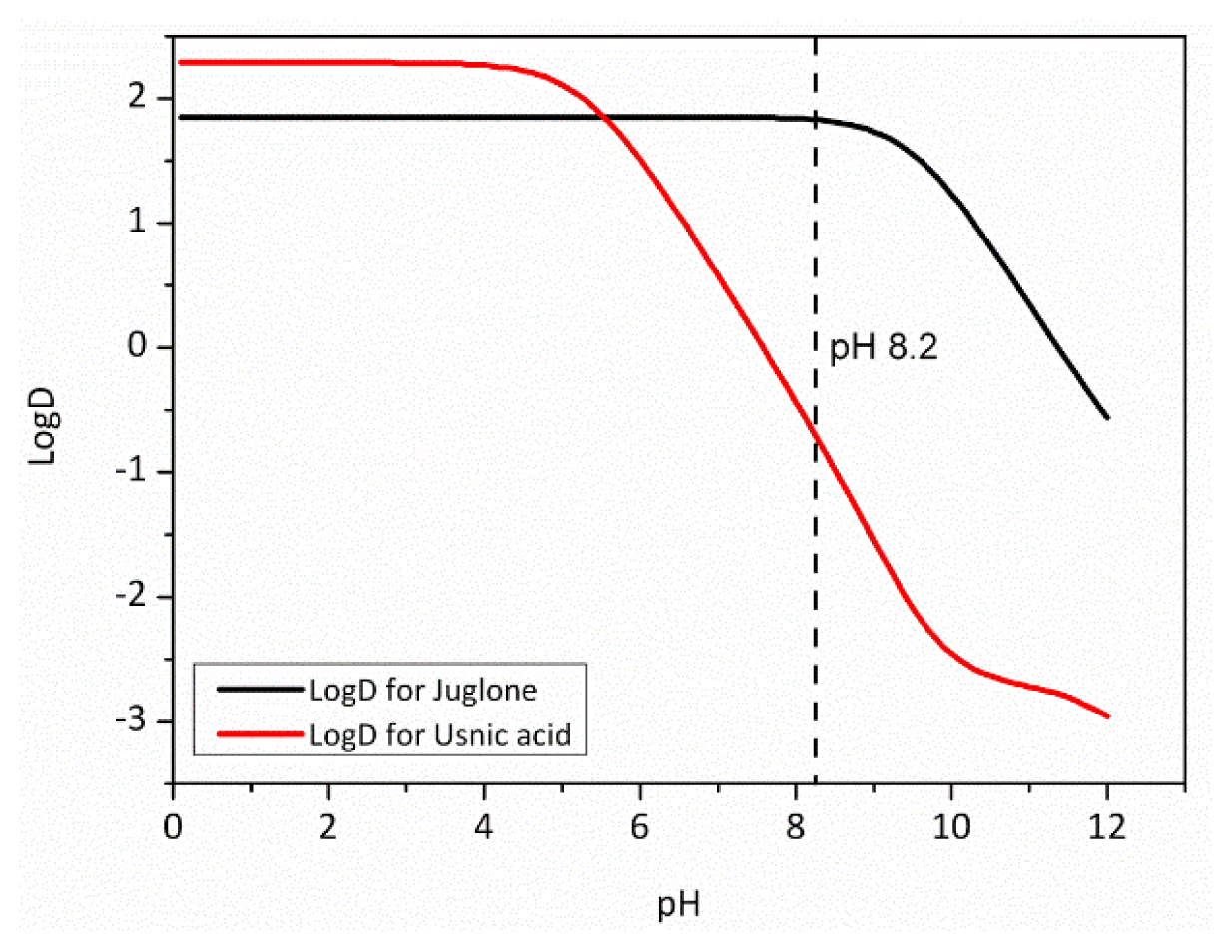
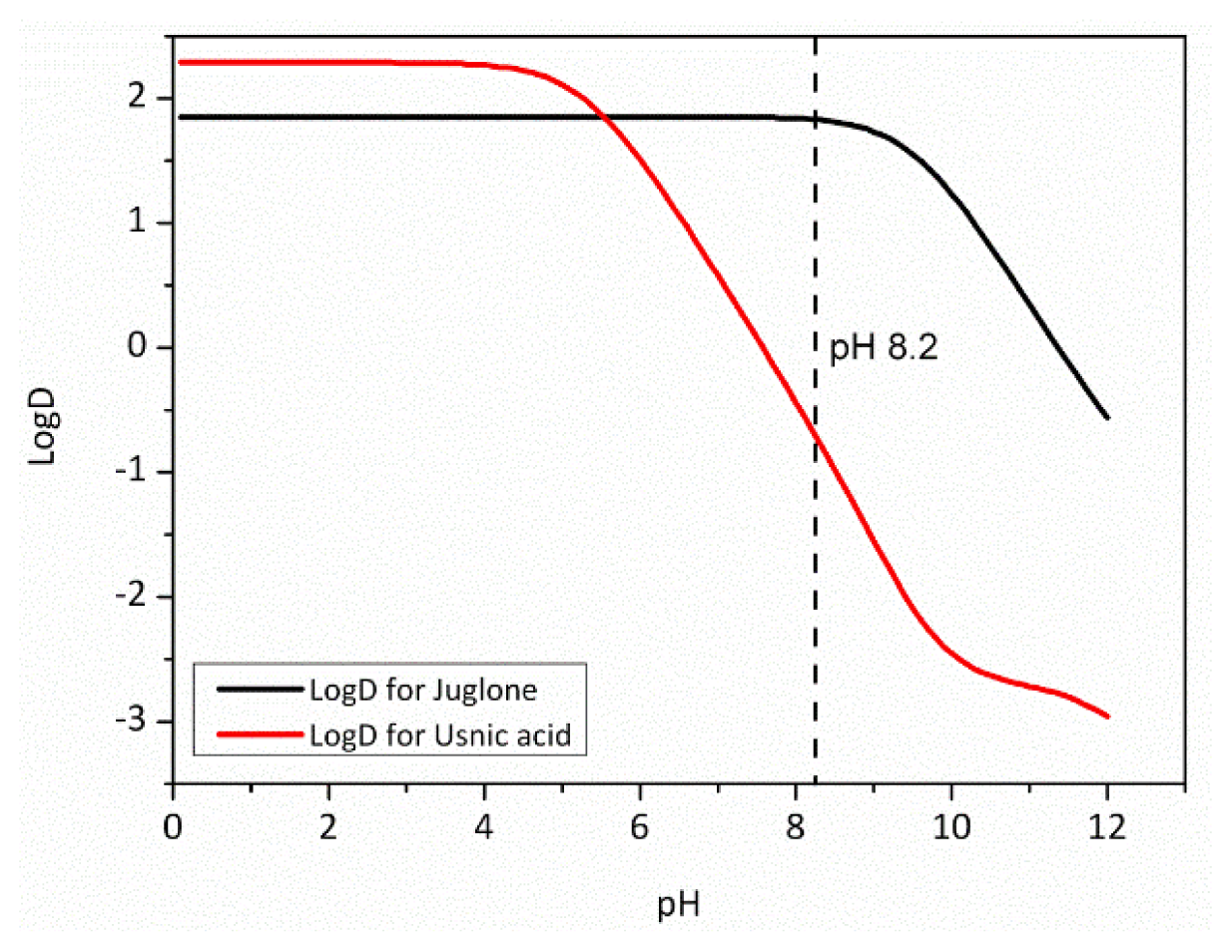
2.6. Physico-Chemical Parameters
3. Materials and Methods
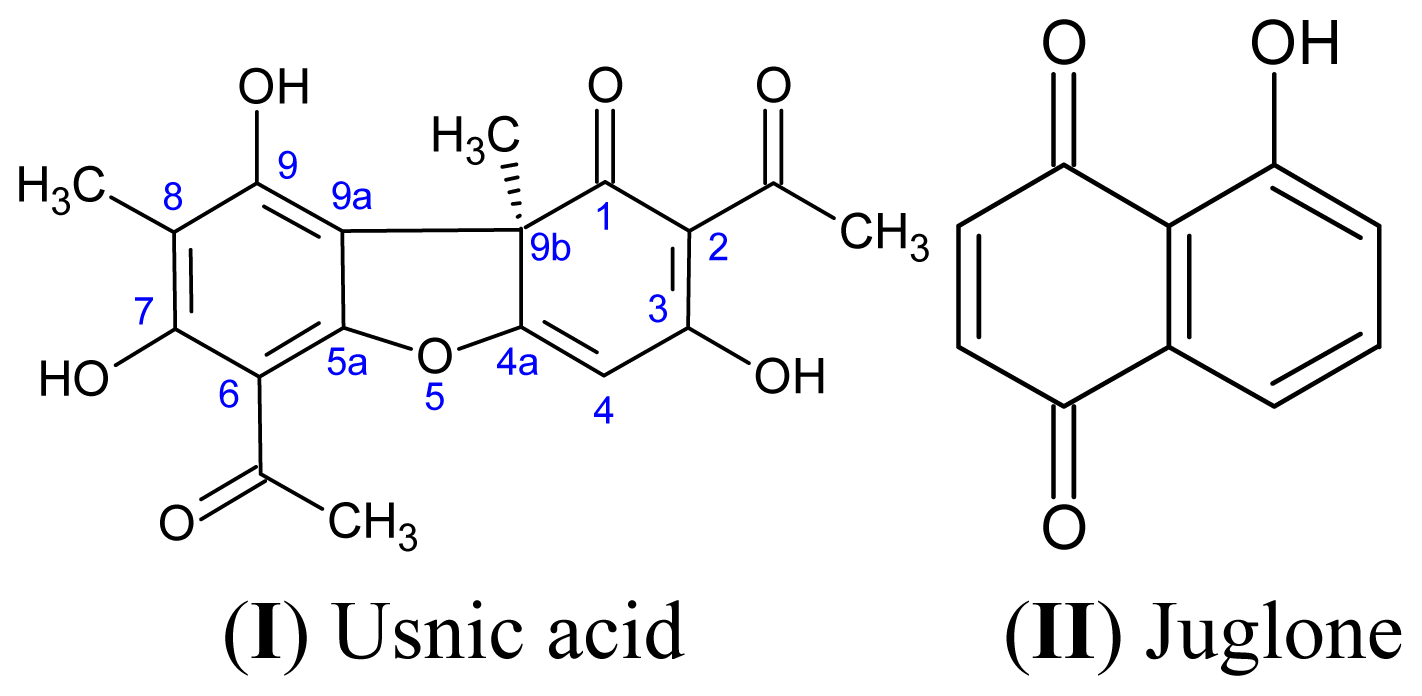
3.1. Natural Products
3.2. Marine Bacteria Test Species
3.3. Culture Conditions and Choice of Media
3.3.1. Planktonic Growth on Different Media
3.3.2. Biofilm Growth under Different Medium Conditions Using Nucleic Acid Staining
3.4. Attachment Bioassays
3.5. Syto©9 Validation (Microplate Reader-Epifluorescence Microscopy)
3.6. Physico-Chemical Parameters
3.7. Statistical Analysis
4. Conclusions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ASWPH | ASWPL | ASW | FASWPH | NSWPH | NSWPL | NSW | |
|---|---|---|---|---|---|---|---|---|
| M. hydro | Growth rate (h−1) | 0.115 ± 0.008 | 0.091 ± 0.006 | 0.022 ± 0.010 | 0.113 ± 0.007 | 0.109 ± 0.009 | 0.090 ± 0.002 | 0.013 ± 0.002 |
| Doubling time (h) | 6.03 ± 0.39 | 7.59 ± 0.52 | 35.66 ± 14.80 | 6.11 ± 0.36 | 6.35 ± 0.53 | 7.63 ± 0.22 | 53.38 ± 8.41 | |
| C. mar | Growth rate (h−1) | 0.079 ± 0.004 | 0.072 ± 0.002 | 0.055 ± 0.006 | 0.075 ± 0.002 | 0.073 ± 0.002 | 0.075 ± 0.002 | 0.06 ± 0.010 |
| Doubling time (h) | 8.77 ± 0.47 | 9.54 ± 0.30 | 13.77 ± 2.25 | 9.21 ± 0.31 | 9.39 ± 0.30 | 9.15 ± 0.27 | 11.29 ± 2.51 | |
| Peak | ppm | Group |
|---|---|---|
| a | 0.896 | –CH3 |
| b | 1.272 | –(CH2)n– |
| c | 1.625 | –OCO–CH2–CH2– |
| d | 2.027 | –CH2–CH=CH– |
| e | 2.331 | –OCO–CH2– |
| f | 4.168 | –CH2–OCOR |
| g | 5.360 | –CH=CH– |
| Parameters | Usnic acid | Juglone | Tolylfluanid | Dichlofluanid | DCOIT (SeaNine™) | Cybutryne |
|---|---|---|---|---|---|---|
| Solubility 1 (ppm) | 420 * | 160 * | 39 | 58 | 63 | 390 |
| LogP2 | 2.29 (RI = 0.63) | 1.85 (RI = 0.88) | 3.58 (RI = 0.82) | 3.32 (RI = 0.80) | 4.59 (RI = 0.62) | 3.45 (RI = 0.75) |
| pK a1 3 | 5.3 ± 0.8 | 9.5 ± 0.4 | −5.06 ± 0.5 | −5.37 ± 0.5 | −6.09 ± 0.6 | 4.12 ± 0.1 |
| pK a2 3 | 9.0 ± 0.9 | −16.79 ± 0.7 | −16.89 ± 0.7 | −2.55 ± 0.2 | ||
| pK a3 3 | 12.0 ± 0.9 | |||||
| LogD at pH 8.2 | −0.65 | 1.83 | 3.58 | 3.32 | 4.59 | 3.45 |
| Water | Peptone (g L−1) | Sterilisation | Acronym |
|---|---|---|---|
| ASW | 18 | Autoclaved | ASWPH * |
| 9 | Autoclaved | ASWPL | |
| 0 | Autoclaved | ASW | |
| 18 | Filtered (0.2 μm) | ASWPH | |
| NSW | 18 | Filtered (0.2 μm) | NSWPH |
| 9 | Filtered (0.2 μm) | NSWPL | |
| 0 | Filtered (0.2 μm) | NSW | |
| NP | Concentration (ppm) |
|---|---|
| CCT | 0.1, 5, 10, 20, 50, 100, 200 |
| UA | 0.1, 5, 10, 20, 30, 40 |
| JUG | 0.1, 5, 10, 20 |
Acknowledgments
Conflicts of Interest
References
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol 2013. [Google Scholar] [CrossRef]
- Schultz, M.P. Frictional resistance of antifouling coating systems. J. Fluids Eng 2004, 126, 1039–1047. [Google Scholar]
- Schultz, M. Effects of coating roughness and biofouling on ship resistance and powering. Biofouling 2007, 23, 331–341. [Google Scholar]
- International Convention on the Control of Harmfull Anti-fouling Systems on Ships. Available onlin: http://www.google.com.hk/url?sa=t&rct=j&q=International%20Convention%20on%20the%20Control%20of%20Harmfull%20Anti-fouling%20Systems%20on%20ships%20AFS%2FCONF%2F26&source=web&cd=1&ved=0CCsQFjAA&url=http%3a%2f%2fwww.safemedproject.org%2fdocuments%2freference-documents%2ftask1.1-important-documentation-for-maritime-administrations%2fafs%2fafs-convention-english-version%2fat_download%2ffile&ei=g1RbUvudBOPRiAfR_oDgBw&usg=AFQjCNGvTjN1Gq_Uh8yZwRJUU_KHk7YTXw&bvm=bv.53899372,d.aGc&cad=rjt (accessed on 14 October 2013).
- Evans, S.M.; Leksono, T.; McKinnell, P.D. Tributyltin pollution: A diminshing problem following legislation limiting the use of TBT-based anti-fouling paints. Mar. Pollut. Bull 1995, 30, 14–21. [Google Scholar]
- Salta, M.; Wharton, J.A.; Stoodley, P.; Dennington, S.P.; Goodes, L.R.; Werwinski, S.; Mart, U.; Wood, R.J.; Stokes, K.R. Designing biomimetic antifouling surfaces. Philos. Trans. R. Soc. A 2010, 368, 4729–4754. [Google Scholar]
- Scardino, A.J.; de Nys, R. Mini review: Biomimetic models and bioinspired surfaces for fouling control. Biofouling 2011, 27, 73–86. [Google Scholar]
- IMO. International Shipping and World Trade Facts and figures; Maritime Knowledge Centre. Available online: http://80.33.141.76/esatdor/index.php?option=com_content&view=article&id=94:international-shipping-and-world-trade-facts-and-figures-2009-&catid=11:key-eu-data-sources&Itemid=17 (accessed on 14 October 2013).
- Burns, E.; Ifrach, I.; Carmeli, S.; Pawlik, J.R.; Ilan, M. Comparison of anti-predatory defenses of Red Sea and Caribbean sponges. I. Chemical defense. Mar. Ecol. Prog. Ser 2003, 252, 105–114. [Google Scholar]
- Burns, E.; Ilan, M. Comparison of anti-predatory defenses of Red Sea and Caribbean sponges. II. Physical defense. Mar. Ecol. Prog. Ser 2003, 252, 115–123. [Google Scholar]
- Davis, A.R.; Targett, N.M.; McConnell, O.J.; Young, C.M. Epibiosis of marine algal and benthic invertebrates: Natural products chemistry and other mechanisms inhibiting settlement and overgrowth. Bioorgan. Mar. Chemistry 1989, 3, 85–114. [Google Scholar]
- Abarzua, S.; Jakubowski, S. Biotechnological investigation for the prevention of biofouling. 1. Biological and biochemical principles for the prevention of biofouling. Mar. Ecol. Prog. Ser 1995, 123, 301–312. [Google Scholar]
- Clare, A.S. Marine natural product antifoulants: status and potential. Biofouling 1996, 9, 211–229. [Google Scholar]
- Omae, I. General aspects of tin-free antifouling paints. Chem. Rev 2003, 103, 3431–3448. [Google Scholar]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep 2011, 28, 400. [Google Scholar]
- De Nys, R.; Steinberg, P.D.; Willemsen, P.; Dworjanyn, S.A.; Gabelish, C.L.; King, R.J. Broad spectrum effects of secondary metabolites from the red alga delisea pulchra in antifouling assays. Biofouling 1995, 8, 259–271. [Google Scholar]
- Sudatti, D.; Rodrigues, S.; Coutinho, R.; da Gama, B.A.P.; Salgado, L.T.; Filho, G.M.A.; Pereira, R.C. Transport and defensive role of elatol at the surface of the red seaweed Laurencia obtusa (Ceramiales, Rhodophyta). J. Phycol 2008, 44, 548–591. [Google Scholar]
- Hellio, C.; Bremer, G.; Pons, A.M.; le Gal, Y.; Bourgougnon, N. Inhibition of the development of microorganisms (bacteria and fungi) by extracts of marine algae from Brittany, France. Appl. Microbial. Biotechnol 2000, 54, 543–549. [Google Scholar]
- Hellio, C.; Marechal, J.P.; Véron, B.; Bremer, G.; Clare, A.S.; le Gal, Y. Seasonal variation of antifouling activities of marine algae from the Brittany coast (France). Mar. Biotechnol 2004, 6, 67–82. [Google Scholar]
- Burgess, J.G.; Boyd, K.G.; Armstrong, E.; Jiang, Z.; Yan, L.; Berggren, M.; May, U.; Pisacane, T.; Granmo, A.; Adams, D.R. The development of a marine natural product-based antifouling paint. Biofouling 2003, 19, 197–205. [Google Scholar]
- Shields, R.C.; Mokhtar, N.; Ford, M.; Hall, M.J.; Burgess, J.G.; ElBadawey, M.R.; Jakubovics, N.S. Efficacy of a marine bacterial nuclease against biofilm forming microorganisms isolated from chronic rhinosinusitis. PLoS One 2013, 8, e55339. [Google Scholar]
- Qian, P.Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. In Biofouling; 2010; Volume 26, pp. 223–234. [Google Scholar]
- Chambers, L.D.; Hellio, C.; Stokes, K.R.; Dennington, S.P.; Goodes, L.R.; Wood, R.J.K.; Walsh, F.C. Investigation of Chondrus crispus as a potential source of new antifouling agents. Int. Biodeterior. Biodegrad. 2011, 65, 939–946. [Google Scholar]
- Francolini, I.; Norris, P.; Piozzi, A.; Donelli, G.; Stoodley, P. Usnic acid, a natural antimicrobial agent able to inhibit bacterial biofilm formation on polymer surfaces. Antimicrob. Agents Chemother 2004, 48, 4360–4365. [Google Scholar]
- Wright, D.A.; Dawson, R.; Cutler, S.J.; Cutler, H.G.; Orano-Dawson, C.E.; Graneli, E. Naphthoquinones as broad spectrum biocides for treatment of ship’s ballast water: Toxicity to phytoplankton and bacteria. Water Res 2007, 41, 1294–1302. [Google Scholar]
- Gauthier, M.J.; Lafay, B.; Christen, R.; Fernandez, L.; Acquaviva, M.; Bonin, P.; Bertrand, J.C. Marinobacter hydrocarbonoclasticus gen. nov., sp. nov., a new, extremely halotolerant, hydrocarbon-degrading marine bacterium. Int. J. Syst. Bacterial 1992, 42, 568–576. [Google Scholar]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol 1998, 30, 295–304. [Google Scholar]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol 1985, 22, 996–1006. [Google Scholar]
- Li, X.; Yan, Z.; Xu, J. Quantitative variation of biofilms among strains in natural populations ofCandida albicans. Microbiology 2003, 149, 353–362. [Google Scholar]
- Nylund, G.M.; Pavia, H. Inhibitory effects of red algal extracts on larval settlement of the barnacleBalanus improvisus. Mar. Biol 2003, 143, 875–882. [Google Scholar]
- Trevor, R.; Pettitt, A.; Jones, L.; Harwood, J.L. Lipids of the marine red algae, Chondrus crispus and Polysiphonia lanosa. Phytochemistry 1989, 28, 399–405. [Google Scholar]
- Guillen, A.; Legchenko, A. Inversion of surface nuclear magnetic resonance data by an adapted Monte Carlo method applied to water resource characterization. J. Appl. Geophys 2002, 50, 193–205. [Google Scholar]
- Guillén, M.D.; Ruiz, A. High resolution 1H nuclear magnetic resonance in the study of edible oils and fats. Trends Food Sci. Technol 2001, 12, 328–338. [Google Scholar]
- Baumann, L.; Baumann, P.; Mandel, M.; Allen, R.D. Taxonomy of aerobic marine eubacteria. J. Bacteriol 1972, 110, 402–429. [Google Scholar]
- Riber, H.; Wetzel, R. Boundary-layer and internal diffusion effects on phosphorus fluxes in lake periphyton. Limnol. Oceanogr 1987, 32, 1181–1194. [Google Scholar]
- Burkholder, J.; Wetzel, R.; Klomparens, K. Direct comparison of phosphate uptake by adnate and loosely attached microalgae within an intact biofilm matrix. Appl. Environ. Microbiol 1990, 56, 2882–2890. [Google Scholar]
- Wetzel, R. Microcommunities and microgradients: Linking nutrient regeneration, microbial mutualism, and high sustained aquatic primary production. Aquat. Ecol 1993, 27, 3–9. [Google Scholar]
- Haack, T.; McFeters, G. Nutritional relationships among microorganisms in an epilithic biofilm community. Microb. Ecol 1982, 8, 115–126. [Google Scholar]
- Kirchman, D.; Mazzella, L.; Alberte, R.S.; Mitchell, R. Epiphytic bacterial production onZostera marina. Mar. Ecol. Prog. Ser 1984, 15, 117–123. [Google Scholar]
- Neely, R.; Wetzel, R. Simultaneous use of 14C and 3H to determine autotrophic production and bacterial protein production in periphyton. Microb. Ecol 1995, 30, 227–237. [Google Scholar]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish Seaweeds. Int. Food Res. J 2010, 17, 205–220. [Google Scholar]
- Kristensen, J.B.; Olsen, S.M.; Laursen, B.S.; Kragh, K.M.; Poulsen, C.H.; Besenbacher, F.; Meyer, R.L. Enzymatic generation of hydrogen peroxide shows promising antifouling effect. Biofouling 2010, 26, 141–153. [Google Scholar]
- Stupak, M.E.; García, M.T.; Perez, M.C. Non-toxic alternative compounds for marine antifouling paints. Int. Biodeterior. Biodegrad 2003, 52, 49–52. [Google Scholar]
- Sullivan, J.D., Jr.; Ikawa, M. Purification and characterization of hexose oxidase from the red alga Chondrus crispus. Biochim. Biophys. Acta 1973, 309, 11–22. [Google Scholar]
- Hansen, O.; Stougaard, P. Hexose oxidase from the red alga Chondrus crispus. J. Biol. Chem 1997, 272, 11581. [Google Scholar]
- Savary, B.J.; Hicks, K.B.; O’Connor, J.V. Hexose oxidase from Chondrus crispus: Improved purification using perfusion chromatography. Enzyme Microb. Technol 2001, 29, 42–51. [Google Scholar]
- Howell, D. Testing the Impact of Biofilms on the Performance of Marine Antifouling Coatings. In Advances in Marine Antifouling Coatings and Technologies; Hellio, C., Yebra, D., Eds.; Woodhead: Cambridge, UK, 2009. [Google Scholar]
- Olsen, S.M.; Kristensen, J.B.; Laursen, B.S.; Pedersen, L.T.; Dam-Johansen, K.; Kiil, S. Antifouling effect of hydrogen peroxide release from enzymatic marine coatings: Exposure testing under equatorial and Mediterranean conditions. Prog. Org. Coatings 2010, 68, 248–257. [Google Scholar]
- Lauterwein, M.; Oethinger, M.; Belsner, K.; Peters, T.; Marre, R. In vitro activities of the lichen secondary metabolites vulpinic acid,(+)-usnic acid, and (−)-usnic acid against aerobic and anaerobic microorganisms. Antimicrob. Agents Chemother 1995, 39, 2541–2543. [Google Scholar]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar]
- Ingólfsdóttir, K. Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar]
- Campanella, L.; Delfini, M.; Ercole, P.; Iacoangeli, A.; Risuleo, G. Molecular characterization and action of usnic acid: a drug that inhibits proliferation of mouse polyomavirus in vitro and whose main target is RNA transcription. Biochimie 2002, 84, 329–334. [Google Scholar]
- Soderquist, C.J. Juglone and allelopathy. J. Chem. Educ 1973, 50, 782. [Google Scholar]
- O’Brien, P. Molecular mechanisms of quinone cytotoxicity. Chem. Biol. Interact 1991, 80, 1–41. [Google Scholar]
- Inbaraj, J.; Chignell, C. Cytotoxic action of juglone and plumbagin: A mechanistic study using HaCaT keratinocytes. Chem. Res. Toxicol 2004, 17, 55–62. [Google Scholar]
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar]
- Chobot, V.; Hadacek, F. Milieu-dependent pro-and antioxidant activity of juglone may explain linear and nonlinear effects on seedling development. J. Chem. Ecol 2009, 35, 383–390. [Google Scholar]
- De Haan, E.J.; Charles, R. The mechanism of uncoupling of oxidative phosphorylation by 2-methyl-1,4-naphthoquinone. Biochim. Biophy. Acta 1969, 180, 417–419. [Google Scholar]
- Riffel, A.; Medina, L.F.; Stefani, V.; Santos, R.C.; Bizani, D.; Brandelli, A. In vitro antimicrobial activity of a new series of 1, 4-naphthoquinones. Braz. J. Med. Biol. Res 2002, 35, 811–818. [Google Scholar]
- Tran, T.; Saheba, E.; Arcerio, A.V.; Chavez, V.; Li, Q.Y.; Martinez, L.E.; Primm, T.P. Quinones as antimycobacterial agents. Bioorg. Med. Chem 2004, 12, 4809–4813. [Google Scholar]
- Jeon, J.-H.; Lee, C.-H.; Kim, M.K.; Lee, H.-S. Antibacterial effects of juglone and its derivatives against oral pathogens. J. Korean Soc. Appl. Biol. Chem 2009, 52, 720–725. [Google Scholar]
- Lim, M.-Y.; Jeon, J.-H.; Jeong, E.-Y.; Lee, C.-H.; Lee, H.-S. Antimicrobial activity of 5-hydroxy-1, 4-naphthoquinone isolated from Caesalpinia sappan toward intestinal bacteria. Food Chem 2007, 100, 1254–1258. [Google Scholar]
- Belson, N.A.; Cutler, H.G.; Dawson, R.; Wright, D.A. Method for Treating Aquatic Pests U.S. Patent US6164244 A, 2000.
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar]
- Calabrese, E.J. Nitric oxide: Biphasic dose responses. Crit. Rev. Toxicol 2001, 31, 489–501. [Google Scholar]
- Kristmundsdóttir, T.; Jónsdóttir, E.; Ogmundsdóttir, H.M.; Ingólfsdóttir, K. Solubilization of poorly soluble lichen metabolites for biological testing on cell lines. Eur. J. Pharm. Sci 2005, 24, 539–543. [Google Scholar]
- Riedl, J.; Altenburger, R. Physicochemical substance properties as indicators for unreliable exposure in microplate-based bioassays. Chemosphere 2007, 67, 2210–2220. [Google Scholar]
- Schreiber, R.; Altenburger, R.; Paschke, A.; Küster, E. How to deal with lipophilic and volatile organic substances in microtiter plate assays. Environ. Toxicol. Chem 2008, 27, 1676–1682. [Google Scholar]
- Thomas, K.; Brooks, S. The environmental fate and effects of antifouling paint biocides. Biofouling 2010, 26, 73–88. [Google Scholar]
- Comber, S.D.; Franklin, G.; Gardner, M.J.; Watts, C.D.; Boxall, A.B.; Howcroft, J. Partitioning of marine antifoulants in the marine environment. Sci. Total Environ 2002, 286, 61–71. [Google Scholar]
- Sharma, R.K.; Jannke, P.J. Acidity of usnic acid. Ind. J. Chem 1966, 5, 16–18. [Google Scholar]
- Stark, J.B.; Walter, E.D.; Owens, H.S. Method of isolation of usnic acid from Ramalina reticulata. J. Am. Chem. Soc 1950, 72, 1819–1820. [Google Scholar]
- Dana, M.N.; Lerner, B.R. Black Walnut Toxicity. Available online: http://www.hort.purdue.edu/ext/HO-193.pdf (accessed on 14 October 2013).
- National Institutes of Health. U.S. Summary of Data for Chemical Selection Juglone. Available online: http://findpdf.net/reader/SUMMARY-OF-DATA-FOR-CHEMICAL-SELECTION-Juglone-48139.html (accessed on 31 October 2013).
- Ekblad, T.; Bergström, G.; Ederth, T.; Conlan, S.L.; Mutton, R.; Clare, A.S.; Wang, S.; Liu, Y.; Zhao, Q.; D’Souza, F.; et al. Poly (ethylene glycol)-containing hydrogel surfaces for antifouling applications in marine and freshwater environments. Biomacromolecules 2008, 9, 2775–2783. [Google Scholar]
- Akesso, L.; Pettitt, M.E.; Callow, J.A.; Callow, M.E.; Stallard, J.; Teer, D.; Liu, C.; Wang, S.; Zhao, Q.; D’Souza, F.; et al. The potential of nano-structured silicon oxide type coatings deposited by PACVD for control of aquatic biofouling. Biofouling 2009, 25, 55–67. [Google Scholar]
- D’Souza, F.; Bruin, A.; Biersteker, R.; Donnelly, G.; Klijnstra, J.; Rentrop, C.; Willemsen, P. Bacterial assay for the rapid assessment of antifouling and fouling release properties of coatings and materials. J. Ind. Microbiol. Biotechnol 2010, 37, 363–370. [Google Scholar]
- Faÿ, F.; Linossier, I.; Carteau, D.; Dheilly, A.; Silkina, A.; Vallée-Rééhel, K. Booster biocides and microfouling. Biofouling 2010, 26, 787–798. [Google Scholar]
- Camps, M.; Briand, J.F.; Guentas-Dombrowsky, L.; Culioli, G.; Bazire, A.; Blache, Y. Antifouling activity of commercial biocides vs. natural and natural-derived products assessed by marine bacteria adhesion bioassay. Mar. Pollut. Bull 2011, 62, 1032–1040. [Google Scholar]
- Jacob, S.W.; Bischel, M.; Herschler, R.J. Dimethyl Sulfoxide (DMSO): A new concept in pharmacotherapy. Curr. Ther. Res. Clin. Exp 1964, 6, 134–135. [Google Scholar]
- Wood, D.C.; Wood, J. Pharmacologic and biochemical considerations of dimethyl sulfoxide. Ann. N. Y. Acad. Sci 1975, 243, 7–19. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Salta, M.; Wharton, J.A.; Dennington, S.P.; Stoodley, P.; Stokes, K.R. Anti-Biofilm Performance of Three Natural Products against Initial Bacterial Attachment. Int. J. Mol. Sci. 2013, 14, 21757-21780. https://doi.org/10.3390/ijms141121757
Salta M, Wharton JA, Dennington SP, Stoodley P, Stokes KR. Anti-Biofilm Performance of Three Natural Products against Initial Bacterial Attachment. International Journal of Molecular Sciences. 2013; 14(11):21757-21780. https://doi.org/10.3390/ijms141121757
Chicago/Turabian StyleSalta, Maria, Julian A. Wharton, Simon P. Dennington, Paul Stoodley, and Keith R. Stokes. 2013. "Anti-Biofilm Performance of Three Natural Products against Initial Bacterial Attachment" International Journal of Molecular Sciences 14, no. 11: 21757-21780. https://doi.org/10.3390/ijms141121757