A Genome-Wide Expression Profile of Salt-Responsive Genes in the Apple Rootstock Malus zumi


Abstract
:1. Introduction
2. Results
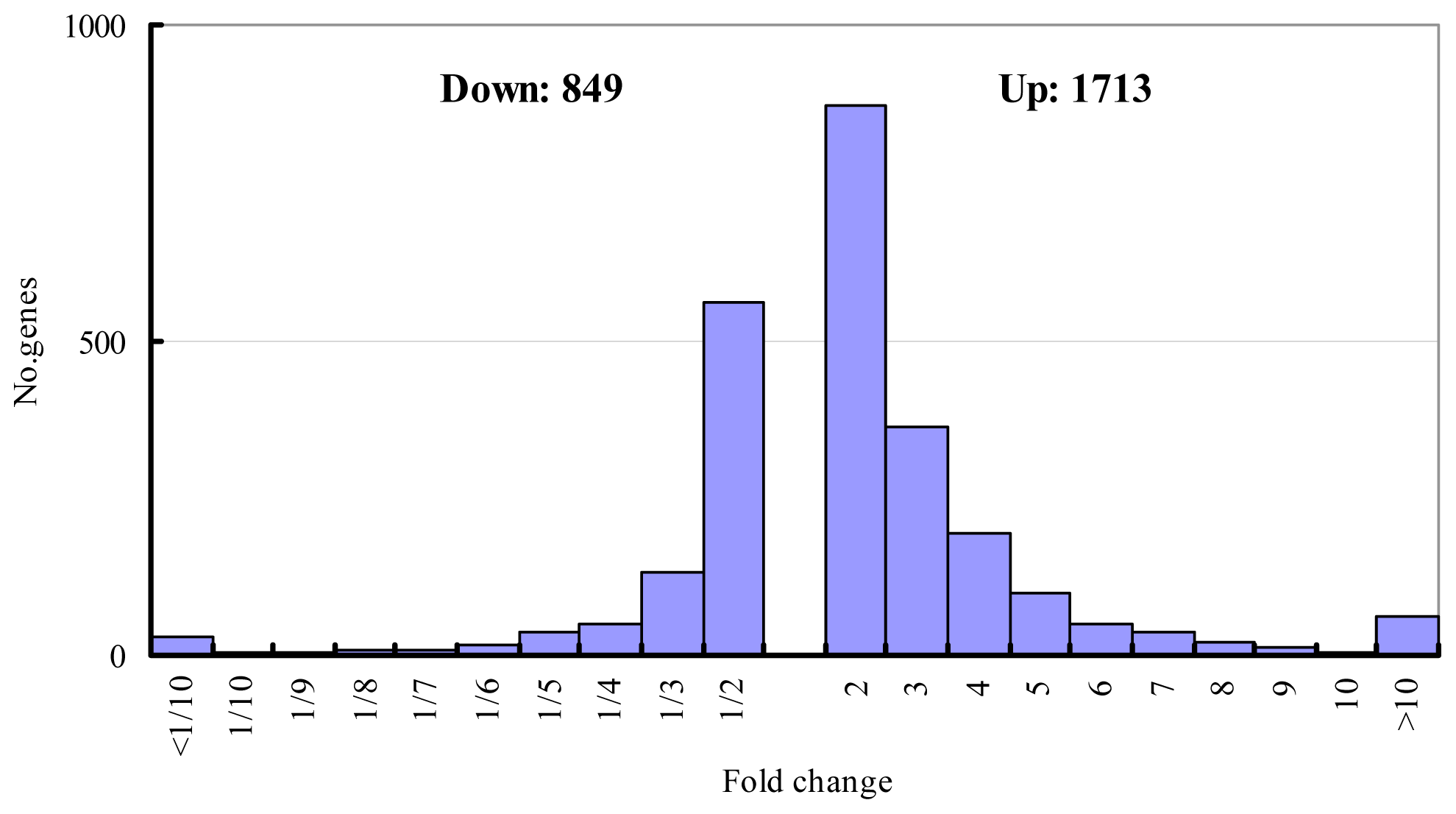
2.1. Salt-Responsive Genes Identified by Microarray Experiment and Semi-Quantitative RT-PCR Validation
2.2. Promoter Analysis
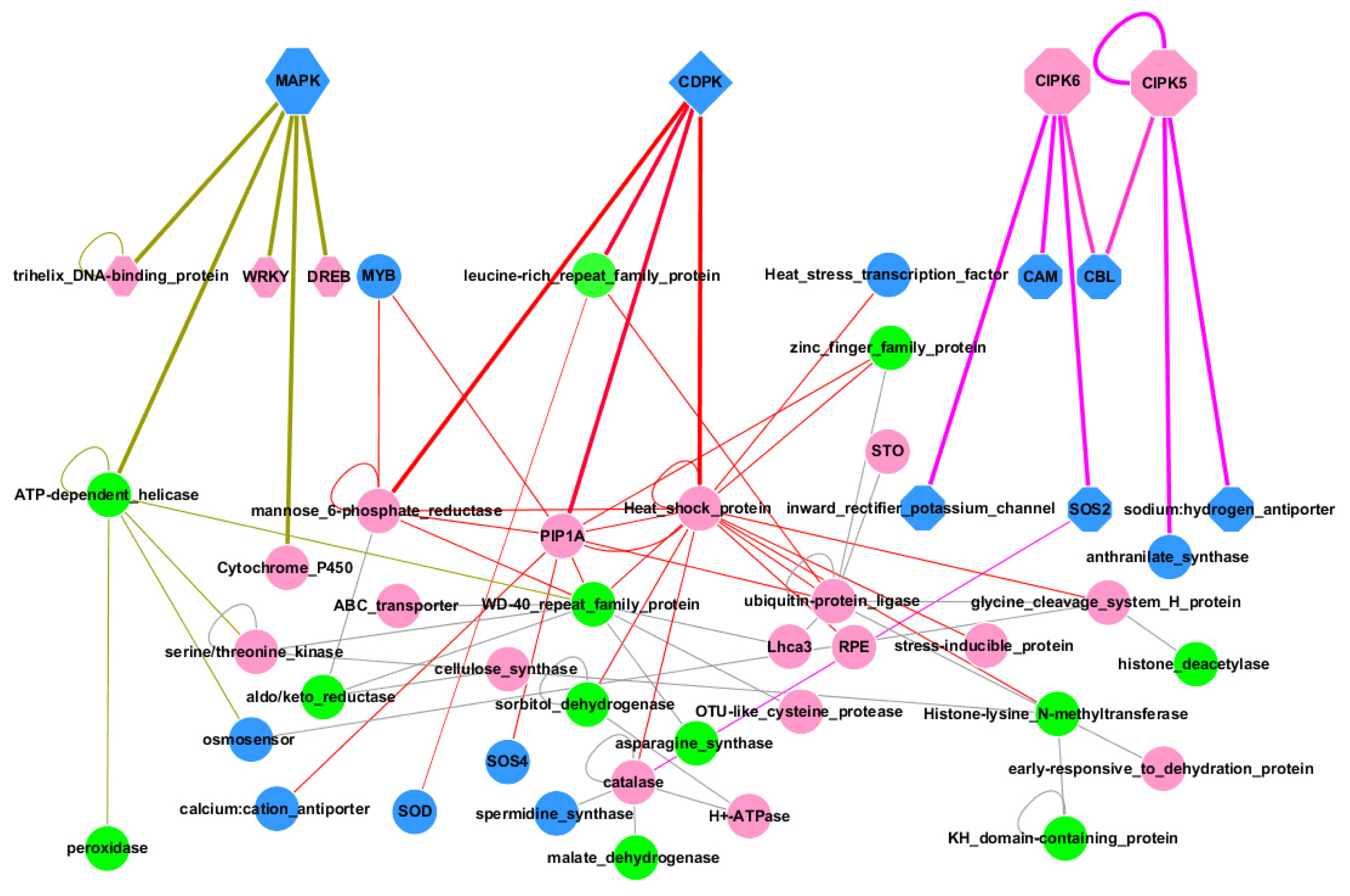
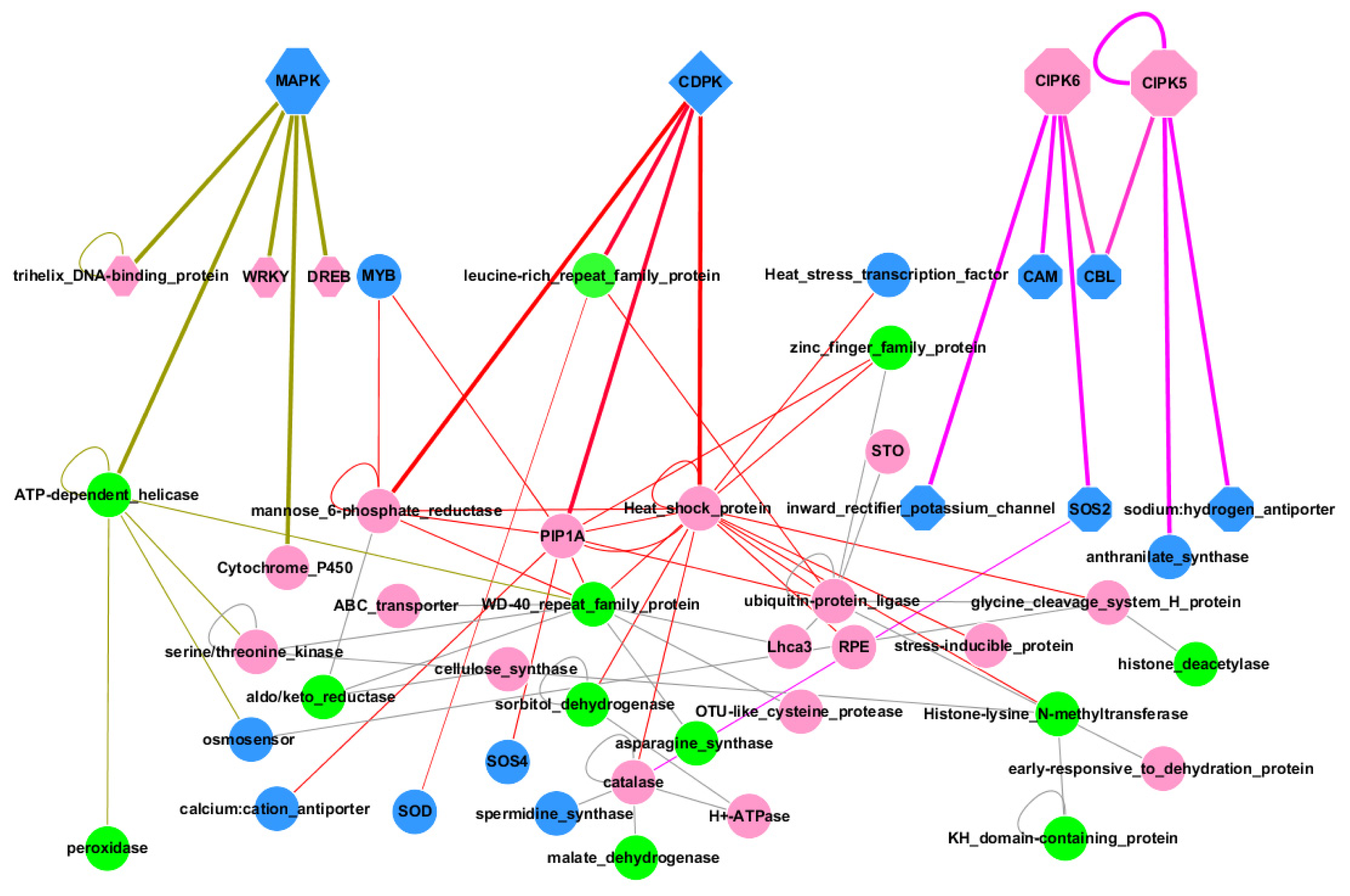
2.3. Regulatory Network during Salt Stress
3. Discussion
3.1. Characteristics of Constructed cDNA Library
3.2. Regulatory Network during Salt Stress
3.3. Salt Stress Signaling
3.4. Analysis of cis-Elements
3.5. Cellular Response Strategies to Salt Stress in Malus zumi
3.6. Stable Photosynthesis Suggests a Novel Salt Tolerance Mechanism in Malus zumi
3.7. Epigenetic and Novel Regulation in Salt Stress Response
4. Experimental Section
4.1. Plant Materials
4.2. RNA Extraction and Construction of the Normalized Full-Length cDNA Library of Malus zumi
4.3. Preparation of Slides for cDNA Microarrays
4.4. Probe Labeling, Slide Hybridization and Scanning
4.5. Data Analysis of Microarray Results
4.6. RT-PCR Analysis
4.7. Promoter Analysis
4.8. Exploring the Interaction Network
5. Conclusions
Supplementary Information
ijms-14-21053-s001.pdf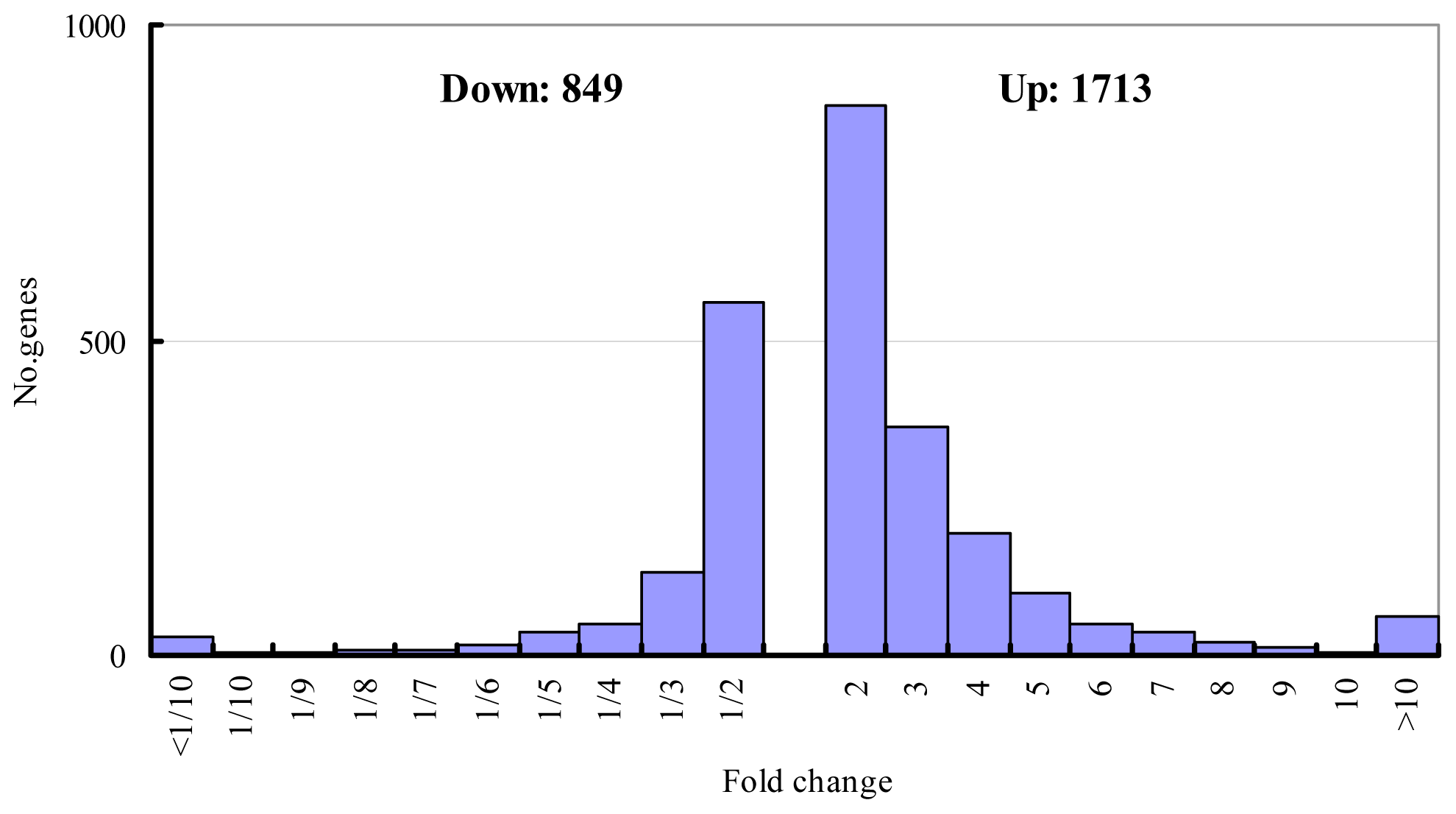


{kind=link}
{kind=link}
{kind=link}
| Functional category | Percentage of unigenes (%) |
|---|---|
| Photosynthesis | 16 |
| Transporter | 5 |
| Metabolism | 17 |
| Stress tolerance | 7 |
| Signal transduction | 6 |
| Protein related | 7 |
| ROS elimination | 6 |
| Osmoprotection | 5 |
| Cell maintenance and development | 6 |
| Others | 4 |
| Unknown protein | 21 |
| Cis-element | ABRE | ARE | TC-rich repeats | HSE | DRE | LTR | ERE | MBS | |
|---|---|---|---|---|---|---|---|---|---|
| Genes with the cis-element among the Regulatory genes | 9/14 | 13/14 | 10/14 | 9/14 | 0/14 | 8/14 | 7/14 | 10/14 | |
| Genes with the cis-element among the functional genes | ROS elimination | 7/10 | 5/10 | 8/10 | 8/10 | 0/10 | 4/10 | 3/10 | 7/10 |
| Osmoregulation | 7/9 | 7/9 | 7/9 | 2/9 | 1/9 | 1/9 | 2/9 | 7/9 | |
| Stress tolerance | 9/14 | 10/14 | 10/14 | 6/14 | 2/14 | 2/14 | 2/14 | 12/14 | |
| Photosynthesis | 21/26 | 20/26 | 17/26 | 13/26 | 1/26 | 12/26 | 7/26 | 18/26 | |
| Transporter | 7/9 | 7/9 | 6/9 | 7/9 | 0/9 | 3/9 | 1/9 | 7/9 | |
| Metabolism | 17/23 | 19/23 | 15/23 | 16/23 | 0/23 | 10/23 | 8/23 | 18/23 | |
| Protein related | 4/11 | 10/11 | 8/11 | 6/11 | 0/11 | 10/11 | 8/11 | 18/11 | |
| Cell maintenance and development | 2/9 | 7/9 | 4/9 | 7/9 | 0/9 | 3/9 | 1/9 | 2/9 | |
| Average number of the cis-elements per gene | 2.43 | 2.24 | 1.75 | 2.04 | 1.00 | 1.45 | 1.19 | 2.10 | |
| Number | Expression a | Putative annotation | Genebank accession | Identities | p-value b |
|---|---|---|---|---|---|
| Kinase | |||||
| 1 | I | leucine-rich repeat transmembrane protein kinase | NP_199948 | 64% | 5.41 × 10−5 |
| 1 | S | leucine-rich repeat family protein kinase | NP_179336 | 42% | 1.02 × 10−4 |
| 1 | I | tousled-like serine/threonine kinase | NP_568405 | 82% | 3.20 × 10−5 |
| 1 | I | CIPK5 | NP_568241 | 79% | 7.74 × 10−5 |
| 1 | S | CIPK6 | NP_194825 | 80% | 1.98 × 10−7 |
| 1 | S | protein kinase family protein | NP_194952 | 50% | 2.25 × 10−5 |
| Transcription factor | |||||
| 1 | I | IAA-LEUCINE RESISTANT3 | NP_200279 | 89% | 4.97 × 10−4 |
| 1 | I | IAA26 | NP_188271 | 80% | 3.58 × 10−6 |
| 1 | S | GT-like trihelix DNA-binding protein | NP_177814 | 37% | 3.04 × 10−6 |
| 1 | S | zinc finger (CCCH-type) family protein | NP_200670 | 59% | 8.13 × 10−5 |
| 1 | I | WRKY family transcription factor | NP_001078015 | 50% | 4.52 × 10−5 |
| 1 | S | GRAS family transcription factor | XP_002322514 | 52% | 3.58 × 10−4 |
| 2 | S | AP2 transcription factor | NP_173355 | 70% | 4.26 × 10−6 |
| 1 | I | SALT TOLERANCE homolog protein | NP_849598 | 68% | 5.37 × 10−7 |
| 1 | S | Auxin response factor | NP_182176 | 70% | 2.78 × 10−3 |
| Number | Expression a | Putative annotation | Genebank accession | Identities | p-value b |
|---|---|---|---|---|---|
| 1 | S | 1-deoxy-D-xylulose-5-phosphate synthase | NP_193291. | 41% | 5.20 × 10−4 |
| 1 | I | lil3 protein | NP_199522 | 51% | 2.56 × 10−7 |
| 2 | I | photosystem II subunit R | NP_178025 | 75% | 3.43 × 10−6 |
| 1 | I | CHLOROPHYLL A/B BINDING PROTEIN 1 | NP_174286 | 67% | 4.38 × 10−3 |
| 2 | I | PSAF (photosystem I subunit F) | NP_174418 | 72% | 3.75 × 10−5 |
| 1 | I | photosystem II CP43 chlorophyll apoprotein | YP_002149729 | 90% | 6.54 × 10−8 |
| 1 | I | photosystem I subunit D-2 | NP_171812 | 72% | 8.79 × 10−4 |
| 1 | I | Thylakoid membrane phosphoprotein of 14 kda | NP_566086 | 64% | 9.54 × 10−7 |
| 1 | I | Photosystem I light harvesting complex gene 3 | NP_176347 | 81% | 5.37 × 10−6 |
| 1 | I | cytochrome b6 | NP_051088 | 98% | 5.86 × 10−7 |
| 1 | I | ATP synthase gamma chain | NP_567265 | 77% | 7.35 × 10−4 |
| 1 | I | NADH dehydrogenase subunit 7 | NP_051115 | 87% | 2.36 × 10−7 |
| 2 | I | photosystem II 44 kDa protein | NP_051055 | 97% | 5.38 × 10−5 |
| 5 | I | light-harvesting complex I protein Lhca3 | XP_002321218 | 81% | 6.67 × 10−6 |
| 1 | I | Oxygen-evolving enhancer protein | NP_201458 | 83% | 4.31 × 10−3 |
| 1 | I | PSBP-1 (PHOTOSYSTEM II SUBUNIT P-1) | NP_172153 | 64% | 3.34 × 10−4 |
| 1 | I | FERREDOXIN-NADP(+)-OXIDOREDUCTASE 1 | NP_201420 | 76% | 4.53 × 10−5 |
| 1 | I | plastid-lipid-associated protein | NP_192311 | 44% | 5.61 × 10−4 |
| 65 | I | ribulose-1,5-bisphosphate carboxylase | CAA79857 | 91% | 5.64 × 10−6 |
| 1 | I | photosystem I P700 chlorophyll a apoprotein A2 | NP_051058 | 98% | 6.35 × 10−3 |
| 7 | I | light-harvesting complex II protein Lhcb2 | XP_002321186 | 92% | 6.89 × 10−7 |
| 1 | I | light-harvesting complex I protein Lhca4 | XP_002330127 | 75% | 8.32 × 10−6 |
| 1 | I | light-harvesting complex I protein Lhca2 | XP_002299309 | 94% | 3.57 × 10−4 |
| 1 | I | PHOTOSYSTEM II SUBUNIT O-2 | NP_190651 | 75% | 4.25 × 10−5 |
| 1 | S | PsbP domain-containing protein | NP_565131 | 65% | 6.31 × 10−7 |
| 5 | I | light-harvesting complex II protein Lhcb1 | XP_002316737 | 95% | 2.58 × 10−2 |
| 6 | I | light-harvesting complex II protein Lhcb6 | XP_002303160 | 82% | 5.34 × 10−5 |
| 1 | I | light-harvesting complex II protein Lhcb5 | XP_002329192 | 81% | 6.41 × 10−7 |
| 2 | I | photosystem II 22 kDa protein | NP_001150026 | 91% | 3.59 × 10−3 |
| 2 | I | protochlorophyllide reductase | NP_200230 | 65% | 5.34 × 10−7 |
| 1 | I | Clp protease proteolytic subunit 6 | NP_563893 | 45% | 4.37 × 10−5 |
| 2 | I | magnesium chelatase H subunit | ACO57443 | 100% | 1.82 × 10−8 |
| 1 | I | Photosystem I P700 chlorophyll A apoprotein | NP_001044491 | 60% | 2.59 × 10−4 |
| 1 | I | carbonic anhydrase | NP_186799 | 61% | 1.56 × 10−4 |
| 1 | I | starch synthase | NP_174566 | 36% | 8.24 × 10−5 |
| 1 | I | glucose-1-phosphate adenylyltransferase | NP_197423 | 81% | 2.34 × 10−7 |
| 1 | I | ribulose-phosphate 3-epimerase, RPE | NP_200949. | 83% | 5.20 × 10−4 |
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol 2008, 59, 651–681. [Google Scholar]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol 2003, 6, 441–445. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant Cellular and Molecular Responses to High Salinity. Rev. Plant Physiol. Plant Mol. Biol 2000, 51, 463–499. [Google Scholar]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol 2002, 53, 247–273. [Google Scholar]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol 2003, 21, 81–85. [Google Scholar]
- Wen, X.P.; Pang, X.M.; Matsuda, N.; Kita, M.; Inoue, H.; Hao, Y.J.; Honda, C.; Moriguchi, T. Over-expression of the apple spermidine synthase gene in pear confers multiple abiotic stress tolerance by altering polyamine titers. Transgenic Res 2008, 17, 251–263. [Google Scholar]
- Yin, R.; Bai, T.H.; Ma, F.W.; Wang, X.J.; Li, Y.H.; Yue, Z.Y. Physiological responses and relative tolerance by Chinese apple rootstocks to NaCl stress. Sci. Hortic 2010, 126, 247–252. [Google Scholar]
- Flowers, T.J. Salinisation and horticultural production. Sci. Hortic 1999, 78, 1–4. [Google Scholar]
- Xia, Y.; Huimin, H.; Shu, H.R.; Wang, T.M.; Liu, D.X.; Fang, Y.F.; Chai, C.H. Changes of leaf membrane penetration, proline and mineral nutrient contents of young apple tree under NaCl stress. J. Fruit Sci 2005, 22, 1–5. [Google Scholar]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet 2010, 42, 833–839. [Google Scholar]
- Janssen, B.J.; Thodey, K.; Schaffer, R.J.; Alba, R.; Balakrishnan, L.; Bishop, R.; Bowen, J.H.; Crowhurst, R.N.; Gleave, A.P.; Ledger, S.; et al. Global gene expression analysis of apple fruit development from the floral bud to ripe fruit. BMC Plant Biol 2008, 8, 16. [Google Scholar]
- Kawaura, K.; Mochida, K.; Ogihara, Y. Genome-wide analysis for identification of salt-responsive genes in common wheat. Funct. Integr. Genomics 2008, 8, 277–286. [Google Scholar]
- Zhao, J.; Wang, J.; An, L.; Doerge, R.W.; Chen, Z.J.; Grau, C.R.; Meng, J.; Osborn, T.C. Analysis of gene expression profiles in response to Sclerotinia sclerotiorum in Brassica napus. Planta 2007, 227, 13–24. [Google Scholar]
- Wei, G.; Tao, Y.; Liu, G.; Chen, C.; Luo, R.; Xia, H.; Gan, Q.; Zeng, H.; Lu, Z.; Han, Y.; et al. A transcriptomic analysis of superhybrid rice LYP9 and its parents. Proc. Natl. Acad. Sci. USA 2009, 106, 7695–7701. [Google Scholar]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar]
- Battaglia, C.; Hsu, C.M.; Soderstrom, K.; Escarre, J.; Haug, F.J.; Charriere, M.; Boccard, M.; Despeisse, M.; Alexander, D.T.; Cantoni, M.; et al. Light trapping in solar cells: Can periodic beat random? ACS Nano 2012, 6, 2790–2797. [Google Scholar]
- Tran, L.S.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar]
- Nagaoka, S.; Takano, T. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot 2003, 54, 2231–2237. [Google Scholar]
- Ding, D.; Zhang, L.F.; Wang, H.; Liu, Z.J.; Zhang, Z.X.; Zheng, Y.L. Differential expression of miRNAs in response to salt stress in maize roots. Ann. Bot 2009, 103, 29–38. [Google Scholar]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell. Biol 2006, 7, 847–859. [Google Scholar]
- Mundy, J.; Yamaguchi-Shinozaki, K.; Chua, N.H. Nuclear proteins bind conserved elements in the abscisic acid-responsive promoter of a rice rab gene. Proc. Natl. Acad. Sci. USA 1990, 87, 1406–1410. [Google Scholar]
- Ross, C.; Shen, Q.J. Computational prediction and experimental verification of HVA1-like abscisic acid responsive promoters in rice (Oryza sativa). Plant Mol. Biol 2006, 62, 233–246. [Google Scholar]
- von Koskull-Doring, P.; Scharf, K.D.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci 2007, 12, 452–457. [Google Scholar]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 2004, 9, 244–252. [Google Scholar]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol 2007, 143, 1739–1751. [Google Scholar]
- Martin, C.; Paz-Ares, J. MYB transcription factors in plants. Trends Genet 1997, 13, 67–73. [Google Scholar]
- Dolferus, R.; Jacobs, M.; Peacock, W.J.; Dennis, E.S. Differential interactions of promoter elements in stress responses of the Arabidopsis Adh gene. Plant Physiol 1994, 105, 1075–1087. [Google Scholar]
- Ashraf, M; F.M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot 2007, 59, 206–216. [Google Scholar]
- Tsugane, K.; Kobayashi, K.; Niwa, Y.; Ohba, Y.; Wada, K.; Kobayashi, H. A recessive arabidopsis mutant that grows photoautotrophically under salt stress shows enhanced active oxygen detoxification. Plant Cell 1999, 11, 1195–1206. [Google Scholar]
- Kanayama, Y.; Moriguchi, R.; Yamaki, S.; Deguchi, M.; Kanahama, K. Effects of environmental stresses and abscisic acid on Sorbitol-6-Phosphate dehydrogenase expression in Rosaceae fruit trees. Acta Hortic 2007, 375–381. [Google Scholar]
- Yamaki, S.I.K. Role of four sorbitol related enzymes and invertase in the seasonal alteration of sugar metabolism in apple tissue. J. Am. Soc. Hort. Sci 1986, 111, 134–137. [Google Scholar]
- Li, F.; Lei, H.J.; Zhao, X.J.; Tian, R.R.; Li, T.H. Characterization of Three Sorbitol Transporter Genes in Micropropagated Apple Plants Grown under Drought Stress. Plant Mol. Biol. Rep 2012, 30, 123–130. [Google Scholar]
- Pharr, D.M.; Stoop, J.M.H.; Williamson, J.D.; Feusi, M.E.S.; Massel, M.O.; Conkling, M.A. The Dual Role of Mannitol as Osmoprotectant and Photoassimilate in Celery. Hortscience 1995, 30, 1182–1188. [Google Scholar]
- Tarczynski, M.C.; Jensen, R.G.; Bohnert, H.J. Stress protection of transgenic tobacco by production of the osmolyte mannitol. Science 1993, 259, 508–510. [Google Scholar]
- Cubero, B.; Nakagawa, Y.; Jiang, X.Y.; Miura, K.J.; Li, F.; Raghothama, K.G.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. The phosphate transporter PHT4;6 is a determinant of salt tolerance that is localized to the Golgi apparatus of Arabidopsis. Mol. Plant 2009, 2, 535–552. [Google Scholar]
- Li, Q.T.; Zhao, Y.; Zhou, Q.Y.; Huang, L.F.; Jiang, Y.Z.; Chen, H.; Kong, J. Cloning and expression analysis of MzPIP1;1 gene from Malus zumi Mats. J. China Agric. Univ 2012, 17, 6. [Google Scholar]
- Bartholomew, D.M.; Van Dyk, D.E.; Lau, S.M.; O’Keefe, D.P.; Rea, P.A.; Viitanen, P.V. Alternate energy-dependent pathways for the vacuolar uptake of glucose and glutathione conjugates. Plant. Physiol 2002, 130, 1562–1572. [Google Scholar]
- Marrs, K.A.; Alfenito, M.R.; Lloyd, A.M.; Walbot, V. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar]
- Balakirev, M.Y.; Tcherniuk, S.O.; Jaquinod, M.; Chroboczek, J. Otubains: A new family of cysteine proteases in the ubiquitin pathway. EMBO Rep 2003, 4, 517–522. [Google Scholar]
- Hanna, J.; Meides, A.; Zhang, D.P.; Finley, D. A ubiquitin stress response induces altered proteasome composition. Cell 2007, 129, 747–759. [Google Scholar]
- Jentsch, S. The ubiquitin-conjugation system. Annu. Rev. Genet 1992, 26, 179–207. [Google Scholar]
- Singer, T.; Haefner, S.; Hoffmann, M.; Fischer, M.; Ilyina, J.; Hilt, W. Sit4 phosphatase is functionally linked to the ubiquitin-proteasome system. Genetics 2003, 164, 1305–1321. [Google Scholar]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot 2009, 103, 551–560. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Effects of salinity on sodium content and photosynthetic responses of rice seedlings differing in salt tolerance. J. Plant Physiol 2000, 157, 54–58. [Google Scholar]
- Downton, W.J.S. Photosynthesis in Salt-Stressed Grapevines. Aust. J. Plant. Physiol 1977, 4, 183–192. [Google Scholar]
- Ma, L.Q.; Han, Z.H.; Zhou, E.F.; Xu, X.F. Effect of salt stress on protective enzyme system of membrane in Malus zumi and M.baccata. J. Fruit Sci 2006, 23, 495–499. [Google Scholar]
- Qiu, N.W.; Lu, Q.T.; Lu, C.M. Photosynthesis, photosystem II efficiency and the xanthophyll cycle in the salt-adapted halophyte Atriplex centralasiatica. New Phytol 2003, 159, 479–486. [Google Scholar]
- Zhang, C.H.; Shen, Y.B.; Yin, W.L.; Pan, Q.H.; Zhao, Y.G. Effect of salt stress on photosynthesis and growth of four tree species seedings. Sci. Silvae Sin 2002, 38, 27–31. [Google Scholar]
- Chinnusamy, V.; Zhu, J.K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol 2009, 12, 133–139. [Google Scholar]
- Davey, S.; Nass, M.L.; Ferrer, J.V.; Sidik, K.; Eisenberger, A.; Mitchell, D.L.; Freyer, G.A. The fission yeast UVDR DNA repair pathway is inducible. Nucleic Acids Res 1997, 25, 1002–1008. [Google Scholar]
- Lee, G.J.; Vierling, E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein. Plant Physiol 2000, 122, 189–198. [Google Scholar]
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemannmerdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant Pathogenesis-Related Proteins and Their Role in Defense against Pathogens. Biochimie 1993, 75, 687–706. [Google Scholar]
- Zhang, Y.G.; Kong, J.; Wang, Y.; Xu, X.F.; Liu, L.L.; Li, T.Z.; Zhu, Y.J.; Han, Z.H. Isolation and characterisation of a nicotianamine synthase gene MxNas1 in Malus xiaojinensis. J. Hortic. Sci. Biotech 2009, 84, 47–52. [Google Scholar]
- Wilson, I.W.; Kennedy, G.C.; Peacock, J.W.; Dennis, E.S. Microarray analysis reveals vegetative molecular phenotypes of Arabidopsis flowering-time mutants. Plant Cell Physiol 2005, 46, 1190–1201. [Google Scholar]
- Mohr, P.G.; Cahill, D.M. Suppression by ABA of salicylic acid and lignin accumulation and the expression of multiple genes, in Arabidopsis infected with Pseudomonas syringae pv. tomato. Funct. Integr. Genomics 2007, 7, 181–191. [Google Scholar]
- Pollard, K.S.; Dut, S.; van der Laan, M.J. Multiple Testing Procedures: R Multtest Package and Applications to Genomics. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: Berlin, Germany, 2005; Volume Chapter 15. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. Roy. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Q.; Liu, J.; Tan, D.; Allan, A.C.; Jiang, Y.; Xu, X.; Han, Z.; Kong, J. A Genome-Wide Expression Profile of Salt-Responsive Genes in the Apple Rootstock Malus zumi. Int. J. Mol. Sci. 2013, 14, 21053-21070. https://doi.org/10.3390/ijms141021053
Li Q, Liu J, Tan D, Allan AC, Jiang Y, Xu X, Han Z, Kong J. A Genome-Wide Expression Profile of Salt-Responsive Genes in the Apple Rootstock Malus zumi. International Journal of Molecular Sciences. 2013; 14(10):21053-21070. https://doi.org/10.3390/ijms141021053
Chicago/Turabian StyleLi, Qingtian, Jia Liu, Dunxian Tan, Andrew C. Allan, Yuzhuang Jiang, Xuefeng Xu, Zhenhai Han, and Jin Kong. 2013. "A Genome-Wide Expression Profile of Salt-Responsive Genes in the Apple Rootstock Malus zumi" International Journal of Molecular Sciences 14, no. 10: 21053-21070. https://doi.org/10.3390/ijms141021053