Immunological Effects of Oenothein B, an Ellagitannin Dimer, on Dendritic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
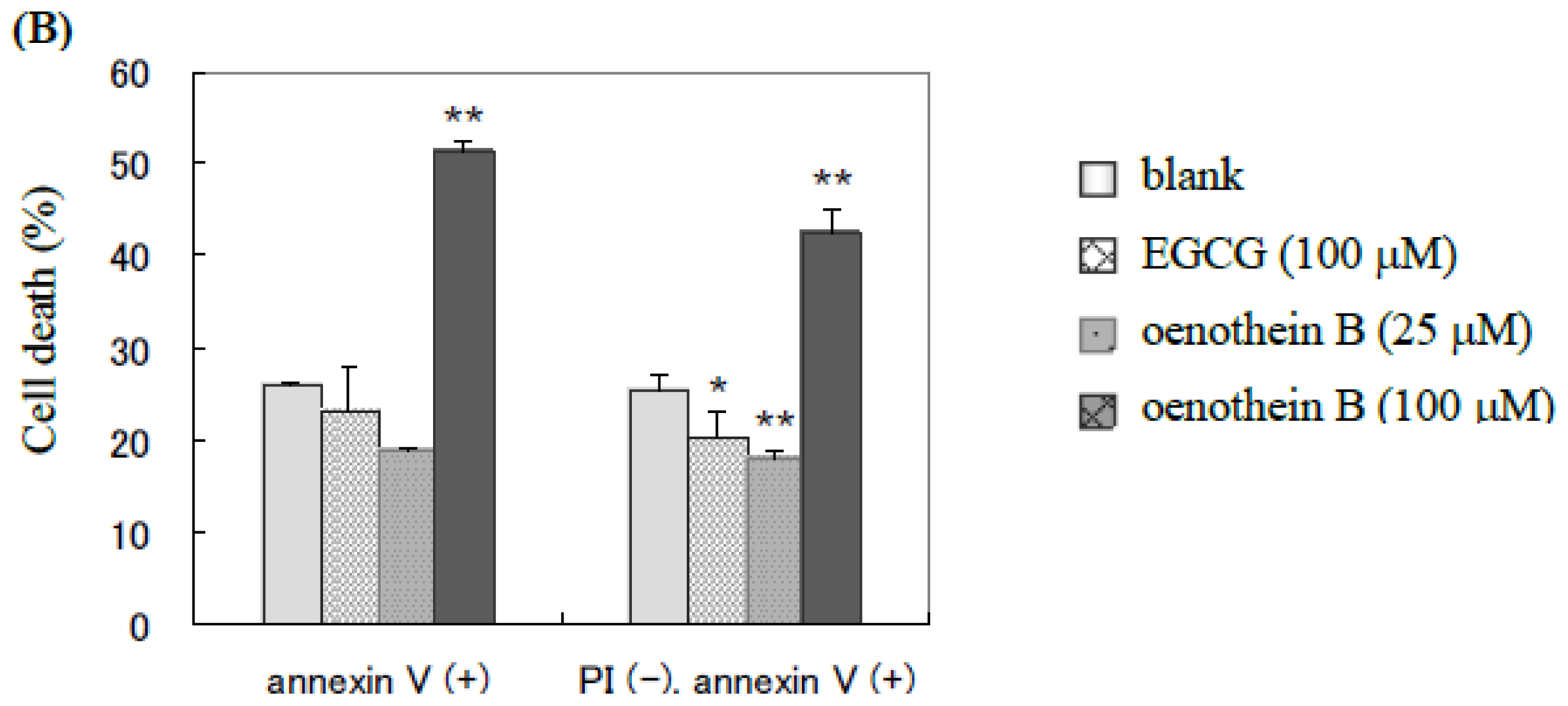
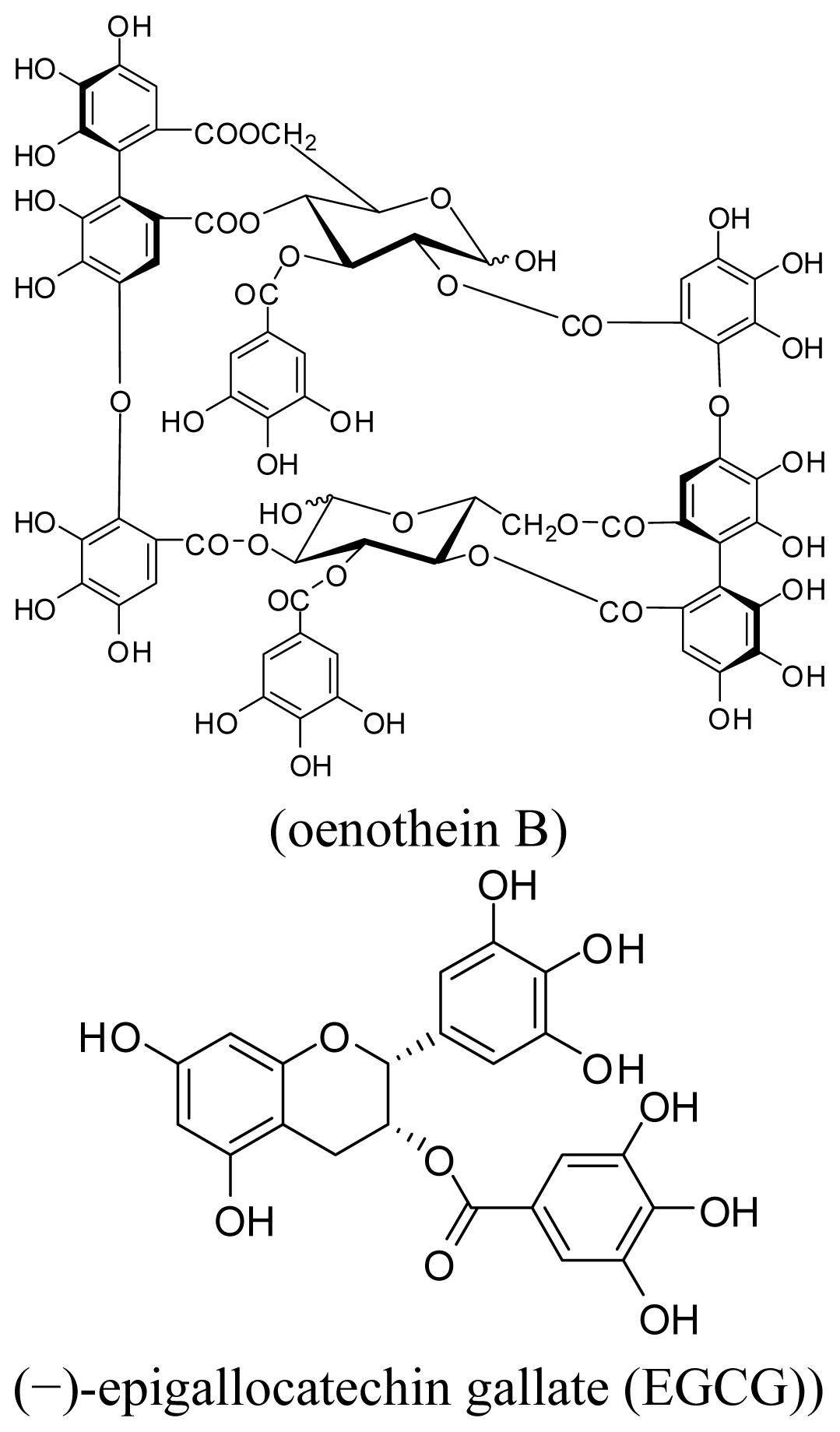
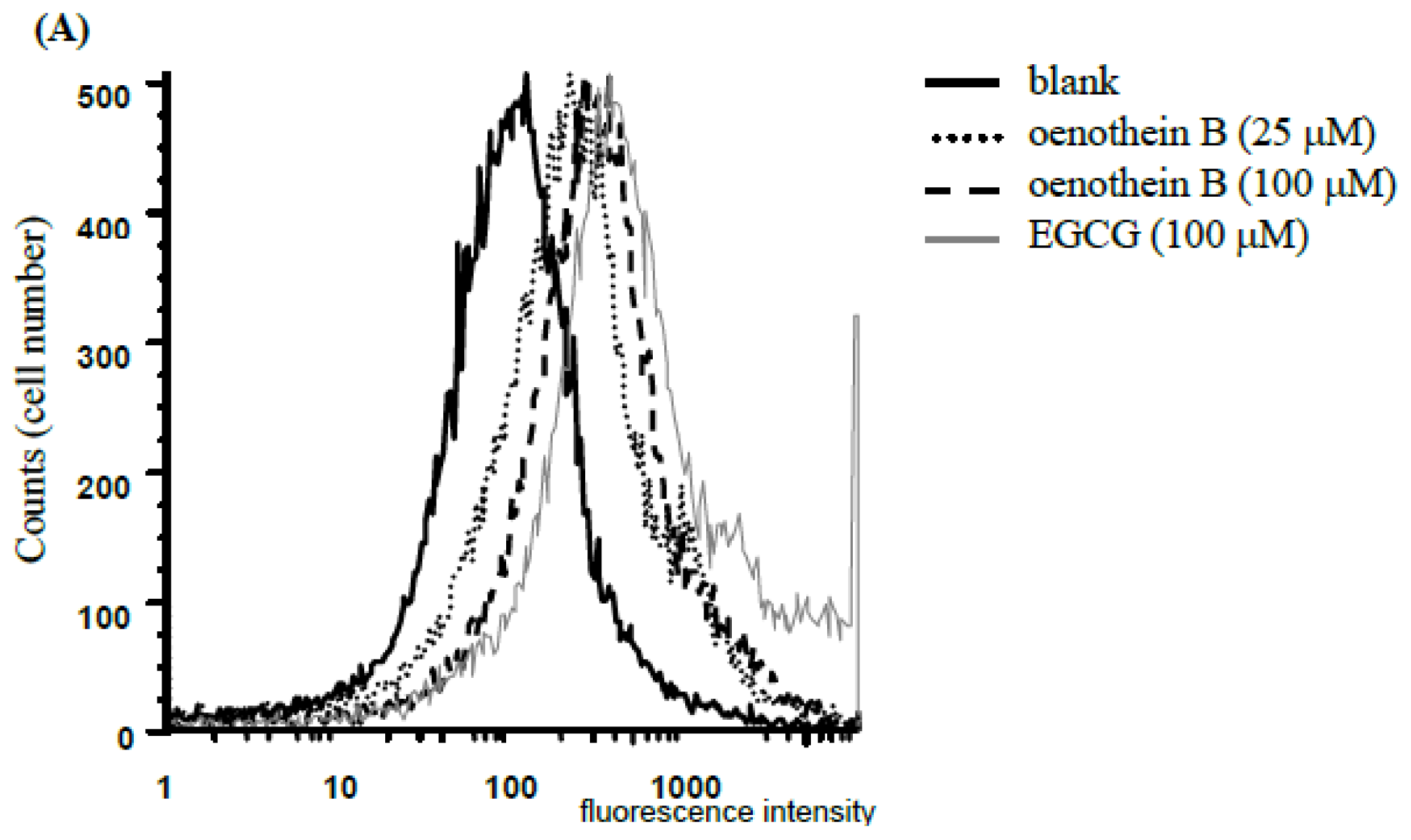
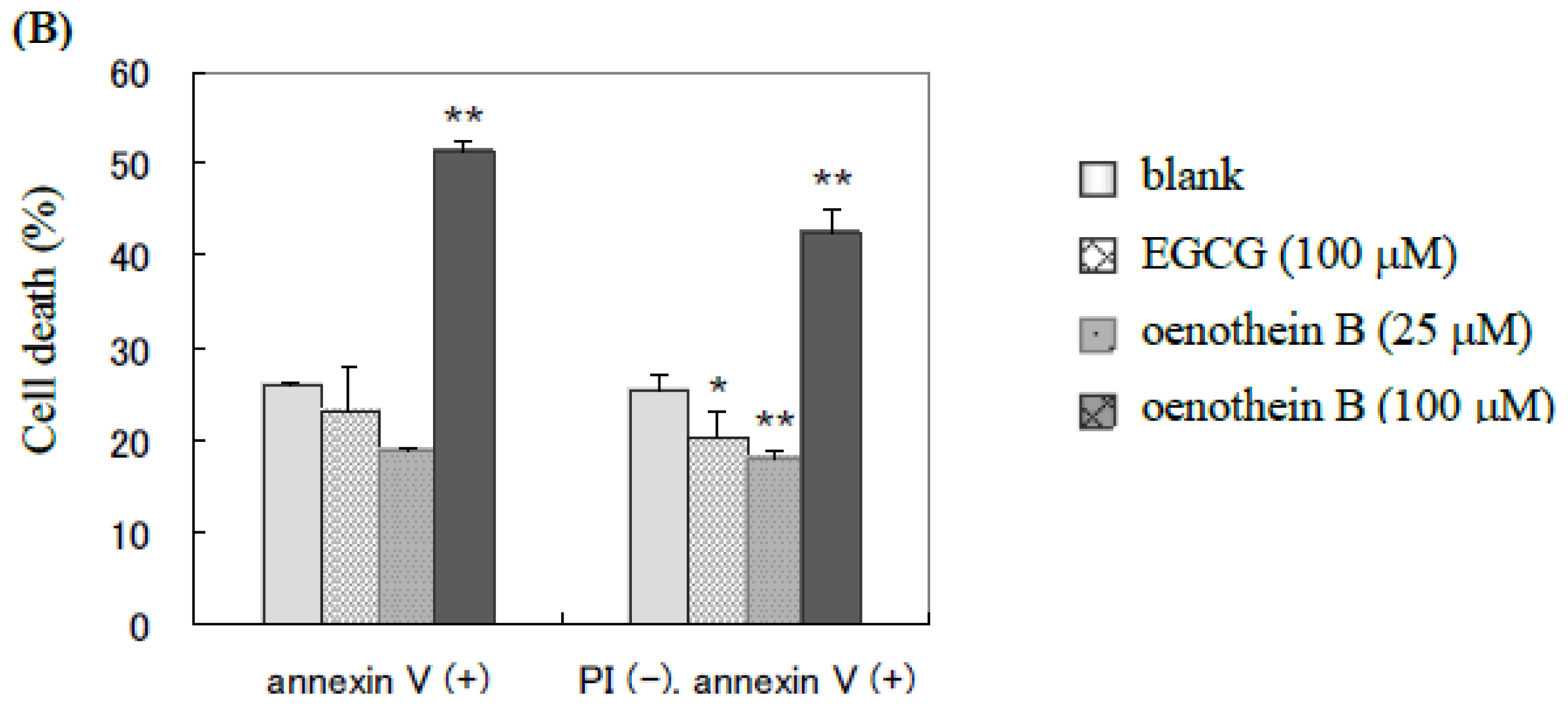
2.1. Analysis of Cell Apoptosis Using Flow-Cytometry
2.2. Flow-Cytometric Analyses of Cell Surface Molecules
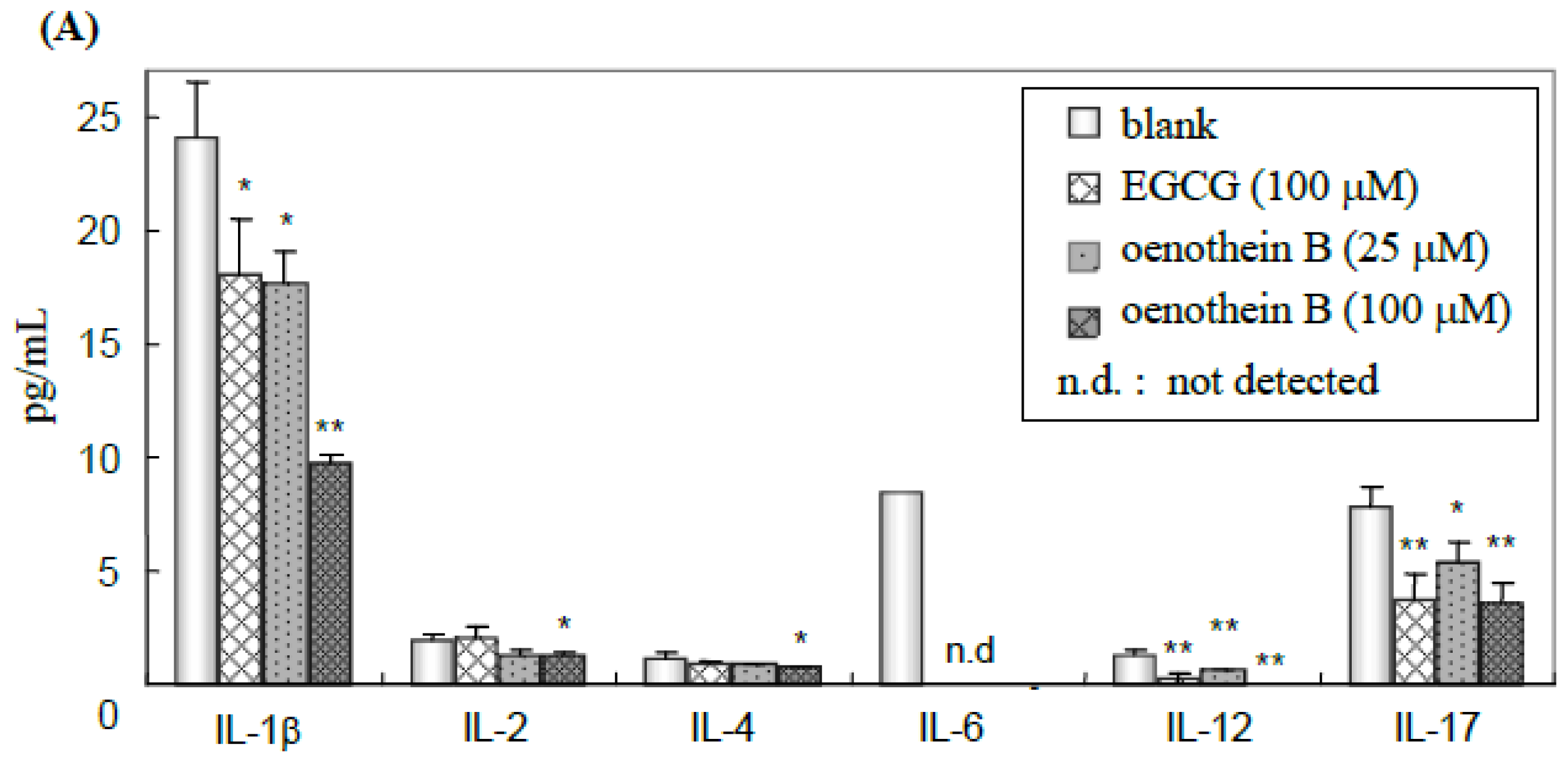
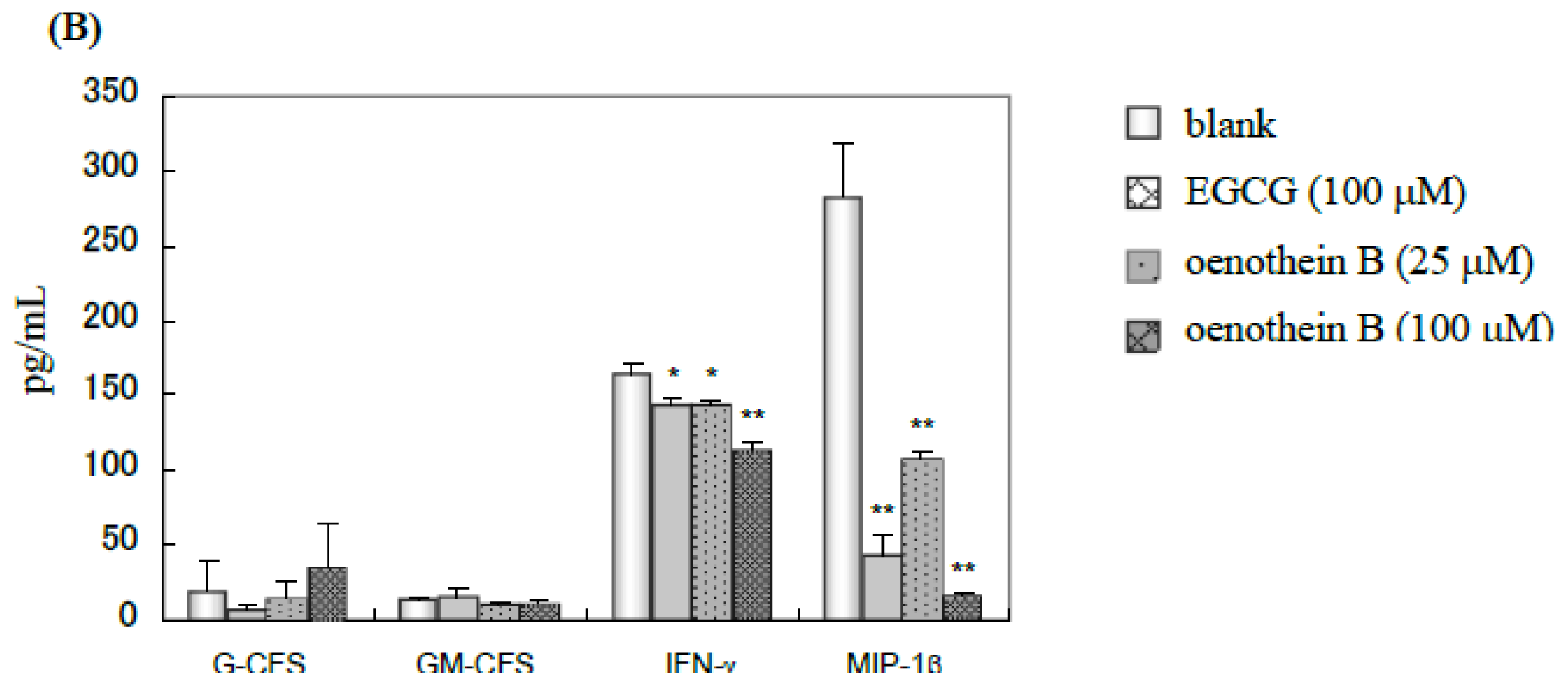
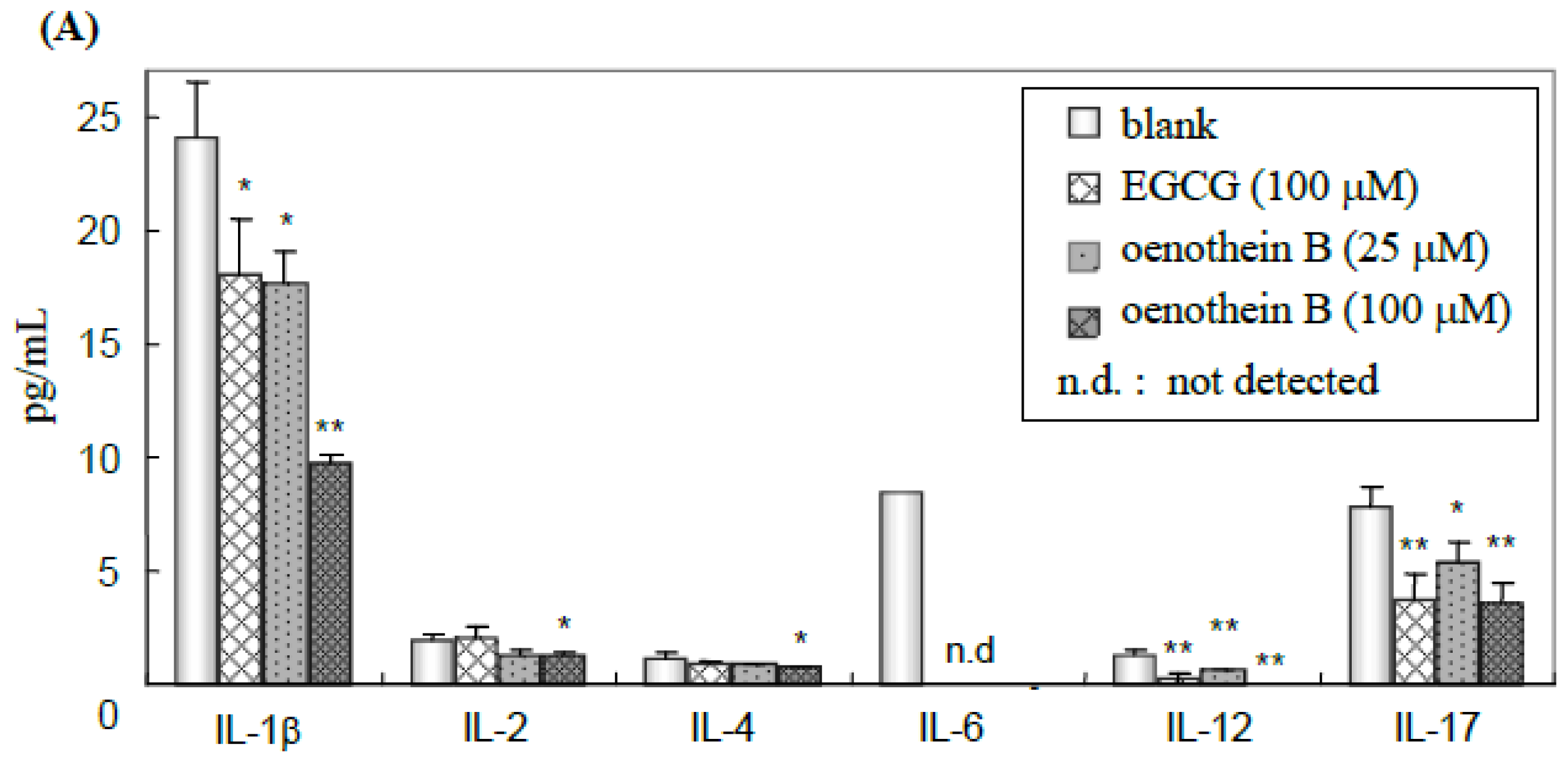
2.3. Quantification of Cytokines in Cell Culture Medium Supernatant
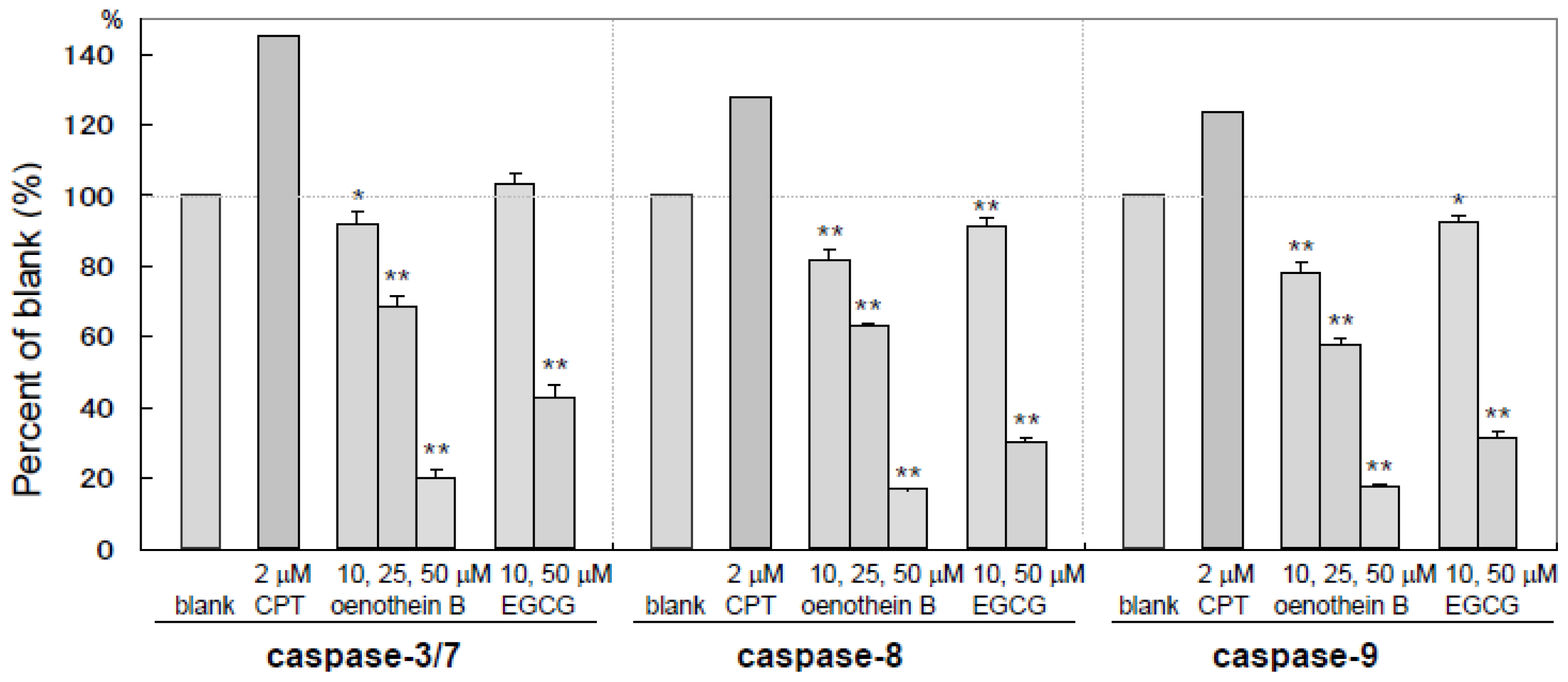
2.4. Measurement of Caspase Activities
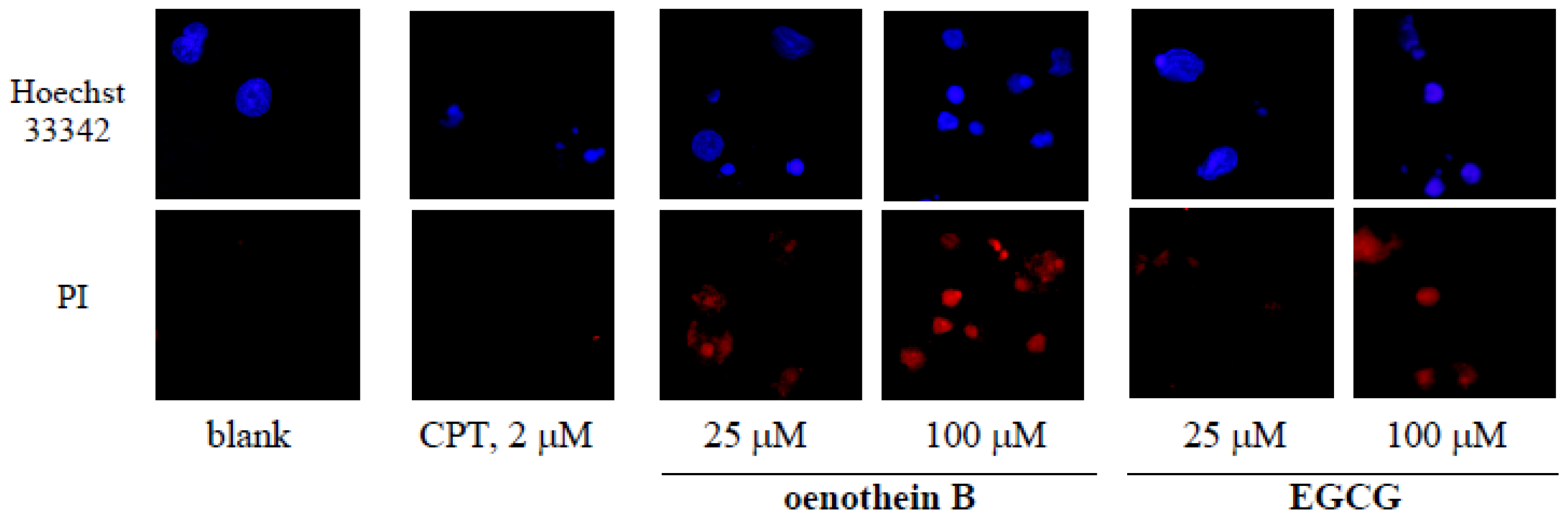
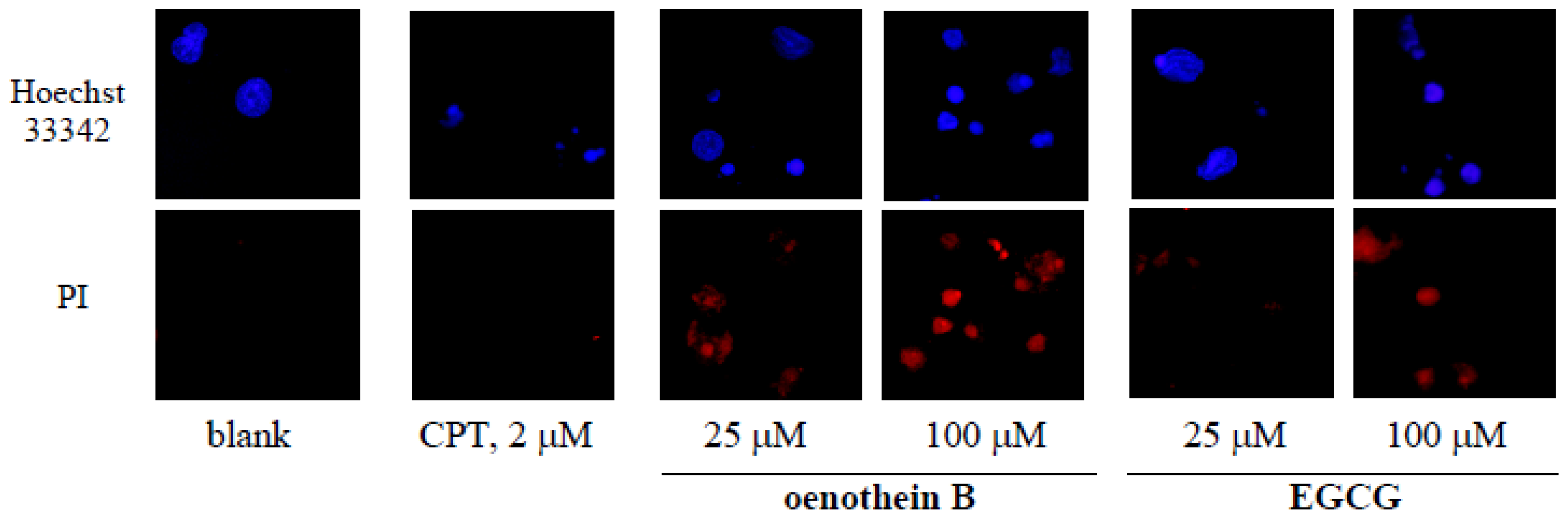
2.5. Morphological Analysis of Tannin-Treated DCs
3. Discussion
4. Experimental Section
4.1. Cell Line, Chemicals and Biochemicals
4.2. Cell Culture
4.3. Flow Cytometric Analyses
4.4. Quantification of Cytokines
4.5. Measurement of Caspase Activities
4.6. Observation of Morphological Changes
5. Conclusions
- Conflict of InterestThe authors declare no conflict of interest.
References
- Okuda, T.; Yoshida, T.; Hatano, T.; Ito, H. Ellagitannins Renewed the Concept of Tannin. Chemistry and Biology of Ellagitannins; Quideau, S., Ed.; World Scientific: Hackensack, NJ, USA, 2009; pp. 1–54. [Google Scholar]
- Miyamoto, K.; Kishi, N.; Koshiura, R.; Yoshida, T.; Hatano, T.; Okuda, T. Relationship between the structures and the antitumor activities of tannins. Chem. Pharm. Bull 1987, 35, 814–822. [Google Scholar]
- Kolodziej, H.; Kayser, O.; Kiderlen, A.F.; Ito, H.; Hatano, T.; Yoshida, T.; Foo, L.Y. Antileishmanial activity of hydrolyzable tannins and their modulatory effects on nitric oxide and tumor necrosis factor-α release in macrophages in vitro. Planta Med 2001, 67, 825–832. [Google Scholar]
- Fujiki, H.; Suganuma, M.; Kurusu, M.; Okabe, S.; Imayoshi, Y.; Taniguchi, S.; Yoshida, T. New TNF-α releasing inhibitors as cancer preventive agents from traditional herbal medicine and combination cancer prevention study with EGCG and sulindac or tamoxifen. Mutat. Res. 2003, 523–524, 119–125. [Google Scholar]
- Hatano, T.; Yasuhara, T.; Matsuda, M.; Yazaki, K.; Yoshida, T.; Okuda, T. Oenothein B, a dimeric, hydrolyzable tannin with macrocyclic structure, and accompanying tannins from Oenothera erythrosepala. J. Chem. Soc. Perkin Trans 1990, 10, 2735–2743. [Google Scholar]
- Yoshida, T.; Chou, T.; Nitta, A.; Okuda, T. Woodfordins A, B and C, dimeric hydrolyzable tannins from Woodfordia fruticosa flowers. Heterocycles 1989, 29, 2267–2271. [Google Scholar]
- Yoshida, T.; Amakura, Y.; Yoshimura, M. Structural features and biological properties of ellagitannins in some plant families of the order Myrtales. Int. J. Mol. Sci 2010, 11, 79–106. [Google Scholar]
- Schmid, D.; Gruber, M.; Piskaty, C.; Woehs, F.; Renner, A.; Nagy, Z.; Kaltenboeck, A.; Wasserscheid, T.; Bazylko, A.; Kiss, A.K.; et al. Inhibition of NF-κB-dependent cytokine and inducible nitric oxide synthesis by the macrocyclic ellagitannin oenothein B in TLR-stimulated RAW 264.7 macrophages. J. Nat. Prod 2012, 75, 870–875. [Google Scholar]
- Schepetkin, I.A.; Kirpotina, L.N.; Jakiw, L.; Khlebnikov, A.I.; Blaskovich, C.L.; Jutila, M.A.; Quinn, M.T. Immunomodulatory activity of oenothein B isolated from Epilobium angustifolium. J. Immunol 2009, 183, 6754–6766. [Google Scholar]
- Rescigno, M.; di Sabatino, A. Dendritic cells in intestinal homeostasis and disease. J. Clin. Invest 2009, 119, 2441–2450. [Google Scholar]
- Wenink, M.H.; Han, W.; Toes, R.E.; Radstake, T.R. Dendritic cells and their potential implication in pathology and treatment of rheumatoid arthritis. Handb. Exp. Pharmacol 2009, 188, 81–98. [Google Scholar]
- Miyamoto, Y.; Sano, M.; Haylor, J.L.; El-Nahas, A.M. (−)-Epigallocatechin 3-O-gallate (EGCG)-induced apoptosis in normal rat kidney interstitial fibroblast (NRK-49F) cells. J. Toxicol. Scis. 2008, 33, 367–370. [Google Scholar]
- Berchtold, S.; Mühl-Zürbes, P.; Heufler, C.; Winklehner, P.; Schuler, G.; Steinkasserer, A. Cloning, recombinant expression and biochemical characterization of the murine CD83 molecule which is specifically upregulated during dendritic cell maturation. FEBS Lett 1999, 461, 211–216. [Google Scholar]
- Yu, S.W.; Wang, H.; Poitras, M.F.; Coombs, C.; Bowers, W.J.; Federoff, H.J.; Poirier, G.G.; Dawson, T.M.; Dawson, V.L. Mediation of poly(ADP-ribose) polymerase-1-dependent cell death by apoptosis-inducing factor. Science 2002, 297, 259–263. [Google Scholar]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.C.; Fleshner, M.; Green, C.; Pedersen, B.K.; Hoffman-Goetz, L.; et al. Immune function and exercise. Exerc. Immunol. Rev 2001, 17, 6–63. [Google Scholar]
- Amakura, Y.; Yoshimura, M.; Sugimoto, N.; Yamazaki, T.; Yoshida, T. Marker constituents of the natural antioxidant Eucalyptus leaf extract for the evaluation of food additives. Biosci. Biotechnol. Biochem 2009, 73, 1060–1065. [Google Scholar]
- Rios, L.Y.; Bennett, R.N.; Lazarus, S.A.; Rémésy, C.; Scalbert, A.; Williamson, G. Cocoa procyanidins are stable during gastric transit in humans. Am. J. Clin. Nutr 2002, 76, 1106–1110. [Google Scholar]


© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yoshimura, M.; Akiyama, H.; Kondo, K.; Sakata, K.; Matsuoka, H.; Amakura, Y.; Teshima, R.; Yoshida, T. Immunological Effects of Oenothein B, an Ellagitannin Dimer, on Dendritic Cells. Int. J. Mol. Sci. 2013, 14, 46-56. https://doi.org/10.3390/ijms14010046
Yoshimura M, Akiyama H, Kondo K, Sakata K, Matsuoka H, Amakura Y, Teshima R, Yoshida T. Immunological Effects of Oenothein B, an Ellagitannin Dimer, on Dendritic Cells. International Journal of Molecular Sciences. 2013; 14(1):46-56. https://doi.org/10.3390/ijms14010046
Chicago/Turabian StyleYoshimura, Morio, Hiroshi Akiyama, Kazunari Kondo, Kozue Sakata, Hideki Matsuoka, Yoshiaki Amakura, Reiko Teshima, and Takashi Yoshida. 2013. "Immunological Effects of Oenothein B, an Ellagitannin Dimer, on Dendritic Cells" International Journal of Molecular Sciences 14, no. 1: 46-56. https://doi.org/10.3390/ijms14010046